

Abstract
• Background and Aims Rheum, a highly diversified genus with about 60 species, is mainly confined to the mountainous and desert regions of the Qinghai–Tibetan plateau and adjacent areas. This genus represents a good example of the extensive diversification of the temperate genera in the Qinghai–Tibetan plateau, in which the forces to drive diversification remain unknown. To date, the infrageneric classification of Rheum has been mainly based on morphological characters. However, it may have been subject to convergent evolution under habitat pressure, and the systematic position of some sections are unclear, especially Sect. Globulosa, which has globular inflorescences, and Sect. Nobilia, which has semi-translucent bracts. Recent palynological research has found substantial contradictions between exine patterns and the current classification of Rheum. Two specific objectives of this research were (1) to evaluate possible relationships of some ambiguous sections with a unique morphology, and (2) to examine possible occurrence of the radiative speciation with low genetic divergence across the total genus and the correlation between the extensive diversification time of Rheum and past geographical events, especially the recent large-scale uplifts of the Qinghai–Tibetan Plateau.
• Methods The chloroplast DNA trnL-F region of 29 individuals representing 26 species of Rheum, belonging to seven out of eight sections, was sequenced and compared. The phylogenetic relationships were further constructed based on the sequences obtained.
• Key Results Despite the highly diversified morphology, the genetic variation in this DNA fragment is relatively low. The molecular phylogeny is highly inconsistent with gross morphology, pollen exine patterns and traditional classifications, except for identifying all samples of Sect. Palmata, three species of Sect. Spiciformia and a few species of Sect. Rheum as corresponding monophyletic groups. The monotypic Sect. Globulosa showed a tentative position within the clade comprising five species of Sect. Rheum. All of the analyses revealed the paraphyly of R. nobile and R. alexandrae, the only two species of Sect. Nobilia circumscribed by the possession of large bracts. The crude calibration of lineages based on trnL-F sequence differentiation implied an extensive diversification of Rheum within approx. 7 million years.
• Conclusions Based on these results, it is suggested that the rich geological and ecological diversity caused by the recent large-scale uplifts of the Qinghai–Tibetan Plateau since the late Tertiary, coupled with the oscillating climate of the Quaternary stage, might have promoted rapid speciation in small and isolated populations, as well as allowing the fixation of unique or rare morphological characters in Rheum. Such a rapid radiation, combined with introgressive hybridization and reticulate evolution, may have caused the transfer of cpDNA haplotypes between morphologically dissimilar species, and might account for the inconsistency between morphological classification and molecular phylogeny reported here.
Keywords: Rheum, phylogeny, trnL-F, the Qinghai–Tibetan Plateau, radiation
INTRODUCTION
The Qinghai–Tibetan Plateau is the highest and largest plateau in the world. The rapid uplift of the plateau since the Pliocene overturned the forest vegetation present here, and its subsequent replacement by alpine meadow in most areas greatly reduced the number of its floral components (Wu, 1987). These trends are likely to have continued into the Quaternary, during which climatic fluctuations (Shi et al., 1998) should theoretically have caused large-scale recession and extinction of the biota distributed in the region because of the growth limitations at the high altitudes during the glacial ages. Despite this, the plateau, especially its eastern part which is usually referred to as the eastern Himalayan ‘biodiversity hotspot’ or Hengduan Mountains ‘hotspot’, now has an exceptionally diverse flora (Wilson, 1992), and is the major component of the south-central ‘biodiversity hotspot’: one of 25 areas recognized globally as featuring exceptional concentrations of endemic species (Myers et al., 2000). The region comprises a series of spectacular north–south trending ridges alternating with deep valleys, with altitudes ranging from 2000 to 6000 m a.s.l. (Li et al., 1995). This area (Fig. 1) contains more than 12 000 species of plants and is especially rich in endemic species and genera (Wang et al., 1993). However, the forces underlying the production of such biotic diversity have not yet been clarified, especially within particular species-rich lineages. A combination of two hypotheses has been proposed for the production of such high diversity (Wu, 1988; Wu and Wu, 1996). On the one hand, the south-east part of the plateau might have served as a refuge for ancient species while habitats changed and the climate oscillated because of its complex topology. On the other hand, more species may have been produced from rapid adaptive speciation while the habitats changed and climate oscillated.
Fig. 1.

The total distribution range of Rheum (left) in the world and the collection sites (circles) of species studied (right) except for R. rhaponticum, which is distributed in Europe. Most samples were collected from the Qinghai–Tibetan Plateau (shaded area).
Rheum (Polygonaceae), a highly diversified genus with about 60 species, is mainly distributed in the mountainous and desert regions of the Qinghai–Tibetan Plateau area and Asian interior (Losina-Losinskaya, 1936). These two adjacent areas are putatively centres of both origin and diversification of Rheum, due to its extremely diversified morphology and high endemism at both species and section level (Li, 1988). Nine sections were recognized under Rheum by Losina-Losinskaya (1936). She further suggested that Sect. Palmata is closely related to Sect. Rheum, and that both sections are primitive groups of the genus. Kao and Cheng (1975) acknowledged only five of Losina-Losinskaya's sections and proposed two new sections: Sect. Acuminata based on the cordiform leaves of several species originally placed in Sect. Rheum; and the monotypic Sect. Globulosa, which has spherical inflorescences, but lacks distinct stems. To date, eight sections have been established and acknowledged under Rheum, according to Li (1998). He further accepted Losina-Losinskaya's phylogenetic hypothesis of Rheum (Losina-Losinskaya, 1936), although no new data were provided to support it. However, the phylogenetic relationships of some sections, e.g. Sect. Nobilia and Sect. Globulosa, are difficult to infer if exclusively based on their gross morphology, because they embrace unique morphologies, which show no distinct connections with other sections (Kao and Cheng, 1975; Li, 1988). Recent palynological research has revealed diverse exine ornamentation in Rheum (Table 1) (Yang et al., 2001), but the variations in ornamentation are not consistent with the morphological classification. Some species with distinctly different morphology in different sections share similar types of pollen ornamentation while some species with very similar morphology have contrasting pollen ornamentation. For example, microechinate pollen ornamentation has been found in species of both Sect. Rheum and Sect. Palmata, while two species of Sect. Nobilia have very different ornamentation, pollen being densely microechinate and sparsely perforate in R. globulosum, but rugulate, verrucate and high-relief in the other species, R. nobile. Three of the species (R. officinale, R. palmatum and R. tanguticum) are highly regarded medicinal plants in China, and therefore widely cultivated. Rhubarb is known as the ‘lord’ or ‘king of herbs’. Chinese people have used it for over 2000 years as a purgative medicine, although some scientists consider it a medical enigma.
Table 1.
List of taxa, sources of plant material analysed and GenBank accession numbers
| Section* |
Species |
Locations |
Accession number in GenBank |
Pollen types† |
||||
|---|---|---|---|---|---|---|---|---|
| Sect. I. Rheum (RHEU) | Rheum undulatum L. | Botanical Garden, IPS* Y990615 | AF303435 | 3 | ||||
| R. australe D. Don | Dêqên, Xizang, Liu JQ 1101 | AY566459 | 3 | |||||
| R. webbianum Royle | Cuori, Xizang, Sn31 | AY566450 | 3 | |||||
| R. hotaoense C. Y. Cheng et Kao | Ledu, Qinghai, Yang MH 99130–1 | AY566445 | 1 | |||||
| R. wittrockii Lundstr. | Yili, Xingjiang, Yang MH 99059 | AY566464 | 1 | |||||
| R. forrestii Diels | Dali, Yunnan, Liu JQ 2175 | AY566447 | 3 | |||||
| R. likiangense (L.) Sam. | Yushu, Qinghai, Q99147 | AF303438 | 3 | |||||
| R. lhasaense A. J. Li et P. K. Hsiao | Sangri, Xizang, Liu JQ 1133 | AY566463 | 3 | |||||
| R. compactum L. | Hami, Xinjiang, X99006 | AF303439 | 1 | |||||
| R. rhaponticum L. | Botanical Garden, Geneva, Switzerland Liu JQ, GG001 | AY566446 | ||||||
| Sect. II. Palmata A. Los. (PALM) | R. officinale Baill. | Nanchuan, Chongqing, M991013 | AF303431 | 2 | ||||
| R. palmatum L. | Huzhu, Qinghai, Yang MH Q99134 | AF303433 | ||||||
| Kangding, Sichuan, Liu JQ 2082 | AY566453 | 1 | ||||||
| R. tanguticum Maxim | Maqin, Qinghai, Q99155 | AF303434 | 3 | |||||
| R. tanguticum var. liupanshanense C. Y. Cheng et Kao | Gande, Qinghai, Liu JQ 1773 Kangding, Sichuan, Liu JQ 2060 | AY566452 AY566457 | 2 | |||||
| Sect. III. Acuminata | R. kialense Franch. | Kangding, Sichuan, Liu JQ 2050 | AY566458 | 3 | ||||
| C. Y. Cheng et Kao (ACUM) | ||||||||
| Sect. IV. Deserticola | R. sublanceolatum C. Y. Cheng et Kao | Qamdo, Qinghai, Liu JQ 847 | AY566454 | 3 | ||||
| Maxim (DESE) | R. pumilum Maxim. | Qamdo, Qinghai, Yang MH 99145 | AF303441 | 1 | ||||
| R. nanum Siev. ex Pall. | Balikun, Xinjiang, Yang MH 99129–1 | AY566444 | 1 | |||||
| R. tibeticum Maxim. ex Hook.f. | Qushui, Xizang, Liu JQ 1112 | AY566455 | 3 | |||||
| Sect. V. Spiciforma A. Los. (SPIC) | R. spiciforme Royle | Zhidoi, Qinghai, Liu JQ 689 | AY566461 | 3 | ||||
| R. moorcroftianum Royle | Zhidoi, Qinghai, Liu JQ 688 | AY566460 | 1 | |||||
| R. przewalskyi A. Los. | Huzhu, Qinghai, Q99136 | AF303442 | 4 | |||||
| R. rhizostachyum Schrenk | Sunan, Gansu, Liu JQ 1506 | AY566448 | 1 | |||||
| R. reticulatum A. Los. | Madoi, Qinghai, Liu JQ 820 | AY566462 | 3 | |||||
| Sect. VI. Globulosa C. Y. Cheng et Kao | R. globulosum Gage | Dazi, Xizang, SN221 | AY566449 | 5 | ||||
| Sect. VII. Nobilia | R. nobile Hook.f.et Thoms | Linzhi, Qinghai, Liu JQ 1206 | AY566465 | 6 | ||||
| A. Los. (NOBI) | R. alexandrae Batal. | Kangding, Sichuan, Liu JQ 2051 | AY566456 | 3 | ||||
The infrageneric classification was referred to Li (1988) and the material of Sect. V. Orbiccularia A. Los consisting of the only species, R. tataricum L. f., was unavailable.
Pollen types: 1, microechinate, tectum smooth and densely perforated; 2, microechinate, microreticulate, relatively wide muri, large lumina; 3, microechinate, tectum uneven, smooth sparsely foveolate to perforate; 4, indistinctly rugulate, low relief; 5, distinctly and densely microechinate, sparsely perforate; 6, distinctly rugulate, high relief, verrucate.
The trnL-F region sequence usually comprises two non-coding chloroplast DNA sequences, the trnL intron and trnL/trnF intergenic spacers (Taberlet et al., 1991). This DNA fragment has been widely used in studies of phylogenetic relationships at inter- or intraspecific level due to its fast rate of evolution and great variation (Bakker et al., 2000; Fukuda et al., 2001, 2003; Liu et al., 2002; Hilger et al., 2004) and to estimate the divergence time of species within a given lineage (Richardson et al., 2001a, b). In the study presented here the aim was to establish the possible phylogeny of Rheum based on sequence analysis of the chloroplast trnL-F region. This phylogeny provides a basis for elucidating evolutionary trends of morphology and speciation patterns (such as gradual or rapid speciation) of Rheum. Two specific objectives of this research were (1) to evaluate possible relationships of some ambiguous sections with a unique morphology; and (2) to examine possible occurrence of radiative speciation with low genetic divergence across the total genus and the correlation between the extensive diversification time of Rheum and past geographical events, especially the recent large-scale uplifts of the Qinghai–Tibetan Plateau.
MATERIALS AND METHODS
Plant materials
Twenty-six species, belonging to seven out of eight sections of Rheum L. were sampled (Table 1). These species cover the morphological range in this genus. Most species were collected from the Qinghai–Tibetan Plateau (Fig. 1). Voucher specimens are deposited in the Herbaria of the Northwest Plateau Institute of Biology, the Chinese Academy of Sciences (HNWP) and the School of Pharmaceutical Sciences, Peking University.
DNA extraction, amplification and sequencing
Total DNA was extracted from silica gel-dried leaves using the CTAB method following (Doyle and Doyle, 1987). The trnL-F region was amplified with the ‘c’ (5′-CGGAATTGGTAGACGCTACG) and ‘f’ (5′-ATTTGAACTGGTGACACGAG) primers of Taberlet et al. (1991). The PCR reaction was performed in a 25 μL reaction mixture with 10–40 ng template DNA, containing 19 μL sterile double-distilled water, 2·5 µl of 10× Taq polymerase reaction buffer, 0·5 mm of MgCl2, 0·2 µM of each of the ‘c’ and ‘f’ primers and 1 unit TaqDNA polymerase. Initial template denaturation was programmed at 94 °C for 2 min, followed by 38 cycles of 94 °C for 1 min, 56 °C for 50 s, 72 °C for 1·75 min plus a final extension of 72 °C for 8 min. The PCR products were excised from 1·2 % agarose gels and purified using a CASpure PCR Purification Kit (CASARRAY) to remove the non-incorporated primers and nucleotides. Sequencing reactions were carried out using amplified ‘c’ and ‘f’ primers, products were purified, concentrated by EtOH precipitation and then run on a Megabase 500 Automated DNA Analysis System using dye-terminator chemistry following the manufacturer's protocols. All DNA sequences were submitted to GenBank (Table 1).
Analysis of sequence data
Sequences were aligned using CLUSTAL X software (Thompson et al., 1997) and then refined by hand. All gaps were treated as missing characters. Phylogenetic analyses were performed with PAUP 4.010a (Swofford, 2000) with all characters unweighted and MrBayes (Huelsenbeck and Ronquist, 2001). Heuristic parsimony searches were conducted with 100 replicates of random addition of sequences, in combination with ACCTRAN character optimization, MULPARS+TBR branch-swapping and STEEPEST DESCENT options to search for multiple islands of most parsimonious trees. The 50 % majority rule consensus tree calculated from 40 000 trees is shown in Fig. 2. Although trees are unrooted, phylogenetic analysis using Oxyria digyna and Rumex patientia as outgroups did not collapse these groups. Bootstrap analyses (Felsenstein, 1985) under MP analyses were performed to assess the relative support of the branches. Bootstrap values (BS) were calculated from 1000 replicates using a heuristic search with simple addition of sequences, TBR branch-swapping and MULPARS options.
Fig. 2.
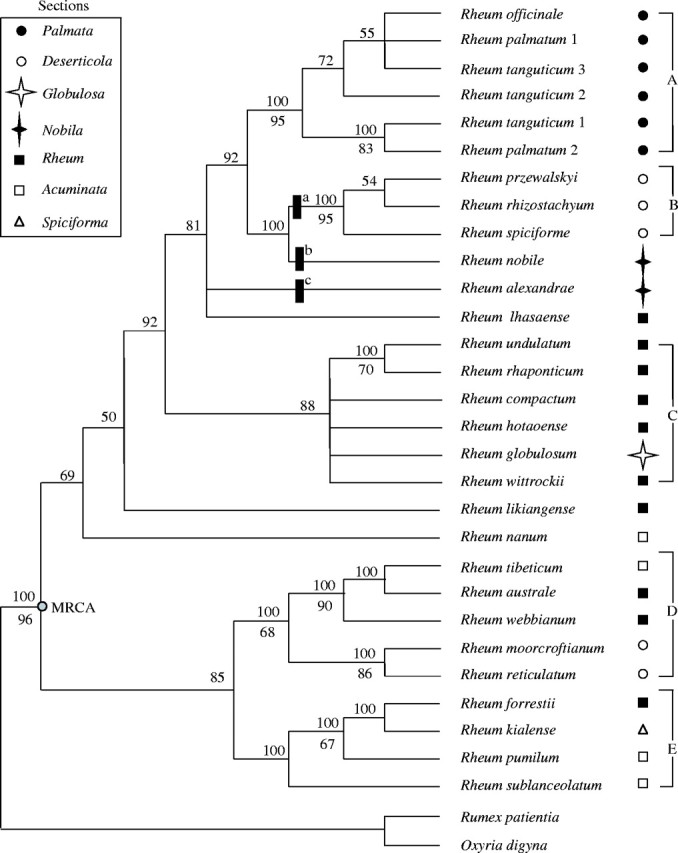
The 50 % majority-rule consensus of 40 000 most parsimonious trees (length = 246, CI = 0·821, RI = 0·86) from parsimony analysis of trnL-F region data (gaps coded as missing values). Bootstrap values with supports >50 % are noted below branches. The sections following each species are indicated. Three indels are marked: a = ‘AAAAAGAT’; b = ‘GTCTTGTGATG’; c = ‘AAAAAAGGAAGAAT’. MRCA, the onset of diversification in the most recent common ancestor of the Rheum.
Modeltest 3.06 (Posada and Crandall, 1998) was then used to select the best model for the evolution of Rheum according to the trnL-F dataset. With the selected model, the phylogeny tree was constructed by the maximum likelihood (ML) method using PAUP 4.010a with simple addition, TBR branch swapping, MULTREES and COLLAPSE options. Bayesian analysis was also performed, based on a general time reversible model with gamma distribution in MrBayes (Huelsenbeck and Ronquist, 2001). For this analysis, four simultaneous Monte Carlo Markov Chains were run for 2 000 000 generations, saving a tree every 100 generations. The last 150 000 trees were used to calculate posterior probability support (PS).
Molecular calibration
In the absence of a fossil record, trnL-F sequences were used to infer the onset of diversification in the most recent common ancestor (MRCA) of the Rheum. The hypothesis of rate constancy was evaluated with a likelihood ratio test that is twice the difference in log likelihood of branch lengths between a rate-constrained tree (forcing the molecular clock in PAUP) and a tree that has no constraints on branch lengths. The molecular clock was rejected because constrained and unconstrained analyses differed significantly, so the average value and deviation of MRCA node divergence was estimated under TreeEdit (Rambaut and Charleston, 2000) based on ML branches without molecular constraints. The time of MRCA onset was calculated as the value of sequence divergence divided by an evolutionary rate of trnL-F following Richardson et al. (2001a).
RESULTS
Phylogenetic analyses
The greatest pairwise distance within Rheum ranged from 0 to 5·597 %, with the largest distance between R. webbianum and R. globulosum, and identical sequences were found in the following pairs of species, R. tibeticcum vs. R. australe, R. przewalskyi vs. R. rhizostachyum vs. R. spiciforme, and R. undulatum vs. R. rhaponticum. The ingroup Rheum species showed a pairwise distance variation of 7·05–9·50 % with two outgroups, Oxyria and Rumex.
The total alignment of trnL-F covered 968 positions, of which 789 were constant, 110 variable but parsimony-uninformative, and 69 only informative when indels were excluded. The heuristic search identified 120 222 most parsimonious trees (length = 246, RI = 0·866, CI = 0·821) and the 50 % strict consensus tree is depicted in Fig. 2, with BS values noted below branches. Most clades were further recovered in the Bayesian analysis (Fig. 3), and posterior PS values were greatly elevated (compared with BS). However, the positions of R. alexandrae, R. nobile, R. nanum and R. likiangense and the phylogenetic relationships of major clades differed from those of the MP analysis. Phylogenetic relationships of species and major clades recovered in the ML analysis (not shown; –lnL = 2727.84404, the best-fit model TrN + G) agree well with Bayesian analysis (Fig. 3).
Fig. 3.

Fifty per cent majority rule consensus tree derived from Bayesian analysis of trnL-F data.
In both MP and strict Bayesian trees (Figs 2 and 3), five tentative groups (A, B, C, D and E) were identified. The first (Group A) consisted of six samples of three species from Sect. Palmata (BS = 95; PS = 98). Group B comprised three species of Sect. Spiciformia, R. przewalskyi, R. rhizostachyum and R. spiciforme with high support (BS = 95; PS = 100). However, the other two species of this section, R. morocroftianum and R. reticulatum, clustered as a subclade (BS = 86; PS = 100) of the well-supported Group D (BS = 68; PS = 98), sister to the other subclade containing R. tibeticum, R. australe and R. webbianum of Sect. Rheum and Sect. Deserticola. Group C comprised six species of Sect. Rheum and the monotypic Sect. Globulosa and received low support (BS < 50; PS = 79). The last tentative group without robust statistical support, Group E, comprised four species in MP analyses, but five species in Bayesian analyses. In the strict MP tree, R. kialense of Sect. Acuminata, R. forrestii of Sect. Rheum and R. pumilum of Sect. Deserticola clustered together as a moderately confident subclade (BS = 67), sister to R. sublanceolatum of Sect. Deserticola. However, R. likiangense clustered with R. kialense, R. forrestii and R. pumilum in the Bayesian analyses with moderate support (PS = 72).
The phylogenetic relationships of the five groups varied in MP and Bayesian analyses: in the former, A, B and C comprised one lineage while D and E formed another (without BS support), while in the latter A and C comprised one lineage (PS = 98) and B, D and E the other (PS = 64). Rheum lhasaense of Sect. Rheum showed tentative relationships with Groups A and B in both analyses and R. nanum sited at the base of the lineage comprising A, B and C in MP analyses, but nested within the lineage containing B, D and E in Bayesian analyses. Rheum nobile of Sect. Nobilia is sister to Group B in the strict MP tree, but nested within the lineage containing Groups B and D in Bayesian analysis with a low support of PS = 67. The other species of this section, R. alexandrae, comprised a lineage with Group A, Group B + R. nobile and R. lhasaense, but nested within the lineage containing Group A, Group C and R. lhasaense with a high support of PS = 98. Because the statistical support in the Bayesian analysis was elevated, the phylogenetic implications of this analysis are more confident.
In the total alignment, a tandem repeat indel (ranging from 0 to 72 bp), and four indels (ranging from 0 to 15 bp) were found from sites 220 to 317 of the trnL intron. As well as these indels, as shown in Fig. 2, R. alexandrae has a unique deletion ‘AAAAAAGGAAGAAT’ at site 64 of the trnL intron, while R. nobile has a unique deletion ‘GTCTTGTGATG’ in the trnL-trnF intergenic spacer. These two different deletions further support the isolated position of R. nobile and R. alexandrae. In addition, three species of Sect. Spiciforma (R. przewalskyi, R. rhizostachyum and R. spiciforme) share a deletion ‘AAAAAGAT’ in the trnL-trnF intergenic spacer, which is phylogenetically informative and unambiguously supports the monophyletic clade comprising these three species (Figs 2 and 3).
Dating the diversification onset of MRCA
The hypothesis of rate constancy was evaluated with a likelihood ratio test that is twice the difference in log likelihood of branch lengths between a rate-constrained tree (forcing the molecular clock in PAUP) (−lnL = 2727·8440) and a tree that has no constraints on branch lengths (−lnL = 2789·98039). The log likelihoods obtained with and without forcing the molecular clock were significantly different, so the rate constancy hypothesis was rejected at a probability level <0·005. The ML branches were saved and the average genetic distance from the MRCA node to each branch tip was estimated under TreeEdit version 1.0 alpha 10 (Rambaut and Charleston, 2000).
The calibrated substitution rates of the trnL-F region ranged from 1·00 × 10−9 to 8·24 × 10−9 substitutions per site (Richardson et al., 2001a). A relatively fast rate of 8·24 × 10−9 was chosen in Aichryson of the Crassulaceae to estimate the diversification onset times of Rheum. The species in both genera are perennials, thus minimizing the effect of generation time on substitution rates, while all other slower rates were calibrated for trees or shrubs. The diversification onset of MRCA based on the average branch distances (0·056) was dated to 6·796 million years ago (Myr) when divided by the rate of 8·24 × 10−9 substitutions per site per year. Molecular calibration of branching time in phylogenetic trees is controversial and should be treated with caution (Sanderson, 1997), but when paleontological data are lacking, molecular estimates provide the only means of inferring lineage ages (Li, 1997). It must be pointed out that the present calibration is very crude, and subject to many potential errors, which might arise from inappropriate calibration rates and other factors (Li, 1997).
DISCUSSION
High inconsistency between gross morphology and the chloroplast DNA trnL-F phylogeny
Li (1988) recognized eight sections in the genus Rheum, based on panicle shape, distinct or indistinct stems and leaf variations. The constructed phylogeny based on trnL-F sequence data coincided with no infrageneric entities based on gross morphology except for Sect. Palmata (Figs 2 and 3). The three species of this section have concordant gross morphology, especially their lobed leaves, but their pollen exine patterns are different (Table 1). This discordance between pollen exine patterns and trnL-F phylogeny also prevails in the other groups identified, i.e. Groups B and D. Despite the poor resolution of the phylogenetic relationships of the five major groups, the present analyses based on trnL-F sequences indicate that Sect. Spiciformia, Sect. Rheum and Sect. Deserticola are paraphyletic. For example, four species of Sect. Deserticola, which is circumscribed by the plants' terminal panicles and indistinct stems, scattered within two groups—D and E.
It is surprising to find that R. globulosum, a species with an unclear position in the previous studies, nested within group C. This species, endemic to the central Qinghai–Tibetan Plateau, differs from all other species of Rheum, having a capitulum-like panicle and short individuals <10 cm in height. In contrast to the sinuate leaf margins of the other species in group C, its leaves are reniform-orbicular with entire margins. The only species of Sect. Acuminata with cordate leaves, R. kialense, was found to be a member of Group E, the other species of which have different leaf shapes, i.e. lanceolate, ovate or elliptic. The paraphyly of only two species of Sect. Nobilia revealed by the trnL-F phylogeny is further supported by the recent finding of their marked differences in bract colour and anatomy (Tsukaya, 2002). Bracts of R. nobile are yellowish and 110–170 µm thick. The mesophyll tissue consists of two or three cell layers and is not differentiated into palisade and spongy parenchyma that lacks distinct plastids and intercellular spaces. However, the bracts of R. alexandrae are creamy white and 230–390 µm thick. The mesophyll tissue of this species consists only of spongy parenchyma with plastids and intercellular spaces.
Recent radiation and evolution of morphology
The samples used in this study represented seven out of eight sections in Rheum, covering a diverse array of morphology, but trnL-F sequence divergence was generally relatively low, ranging from 0·00 % in many cases to 5·597 %. Because of the low number of mutations the parsimonious trees were relatively poorly resolved, with short internal branches (Fig. 2), in contrast to the long terminal braches found in trees for derived mutation-rich groups. All trees had short branch lengths between the most recent common ancestor node, where diversification began, and the branch tips. Such tree topology indicates that recent diversification and rapid radiation has occurred in Rheum (Richardson et al., 2001a).
Previous studies have revealed that similarly low sequence mutation rates and adaptive radiation have occurred in most island archipelago biomes, such as Arygyranthemum in Macronesia (Francisco-Ortega et al., 1997) and silverswords (Baldwin and Sanderson, 1998). These rapid radiations are often hypothesized to have been driven by low levels of competition in newly occupied habitats (Liem, 1990). However, evidence of recent diversity and radiation was recently found in a tropical continental tree genus, Inga, and the causes of its radiation were suggested to be associated with the recent major uplifting of the Andes, the bridging of the Isthmus of Panama, and Quaternary climate oscillations (Richardson et al., 2001a). Geological evidence indicates that extensive habitat changes occurred in the Qinghai–Tibetan Plateau and adjacent areas due to the recent large-scale upliftings of the Qinghai–Tibetan Plateau in the late Tertiary and climate oscillations in the Quaternary within 10 Myr (Harrison et al., 1992; Li et al., 1995; Shi et al., 1998). The crude calibration of MRCA indicates that Rheum began to diversify around 7 Myr. Therefore, it is suggested that the adaptive radiation of Rheum might have been triggered by the recent uplifts of the plateau and the Quaternary climate oscillations. This hypothesis is further strengthened by the expansion of the habitats of its current species. Most Rheum species occur in the Qinghai–Tibetan Plateau and other parts of central Asia, indicating that this area might also be the centre of diversification of the genus (Yang et al., 2001). The habitats preferred by most species are cold and dry alpine meadow, steppe desert and dry slopes. Geological evidence indicates that these arid habitats formed recently as a consequence of the uplifting of the Qinghai–Tibetan Plateau (Shi et al., 1998). The drier climate in central Asia was also created by the uplifting of the Qinghai–Tibetan Plateau (An et al., 2001). In addition, in response to the global climate oscillations that occurred between the late Pliocene and Holocene, the vegetation of the Qinghai–Tibetan Plateau alternated between desert-steppes and forests (Tang and Shen, 1996). The rich geological and ecological diversity of the plateau and adjacent areas of central Asia, together with habitat isolation due to changing climatic conditions during and after the uplifts of the plateau, might well have promoted rapid speciation and radiation of Rheum in small, isolated populations. Such a rapid speciation could have resulted in small numbers of synapomorphic nucleotide substitutions.
As they adapted to the stable aridity of the Qinghai–Tibetan Plateau and adjacent areas after the uplift of the plateau and climatic oscillations, species from different lineages might have been subject to similar selection pressures and thus evolved similar morphologies. This would account for the convergent morphology of the species of polyphyletic sections such as Sect. Deserticola and Sect. Spiciformia. In fact, the morphological characters that define some of these sections are found in diverse families of alpine plants, and have been demonstrated to be of great adaptive value (Ohba and Malla, 1988; Korner, 1999). For example, the indistinct stems and spiciform-panicles that were respectively or collectively used to circumscribe Sect. Deserticola and Sect. Spiciformia, are prevalent among alpine species of many genera, and have been found to result from adaptation to the arid habitats of the plateau or extremely dry parts of central Asia. The colourful bracts shared by two species of Sect. Nobilia (Fig. 2), were shown to have a similar warming effect and to protect reproductive organs from damage by the high levels of UV-B radiation associated with the high altitude of the plateau (Terashima et al., 1993; Omori and Ohba, 1996; Omori et al., 2000). This type of bract also occurs in other alpine plants, e.g. Saussurea of the Asteraceae, and species with this type of bract have also been demonstrated to be paraphyletic (Wang and Liu, 2004b).
Furthermore, such rapid speciation in small and isolated populations driven by selective pressures might promote the fixation of unique or rare morphological characters in some species (Kadereit, 1994), which might cause them to be taxonomically treated at a higher rank than is warranted by the relatively low number of genetic mutations separating them. This could account for the unique morphology of R. globulosum of the monotypic Sect. Globulosa. Many similar examples have been reported from the Qinghai–Tibetan Plateau. For example, the plateau-endemic Milula (Alliaceae), a monotypic genus represented by M. spicata, differs from Allium by having a distinctly elongated, spicate inflorescence instead of the capitate or umbellate inflorescences of the rest of the genus. Molecular data have shown that it is closely related to Allium cyathophorum of the subgenus Rhizirideum with low genetic differentiation (Friesen et al., 2000). Sinadoxa, a monotypic genus endemic to the Qinghai–Tibetan Plateau, differs from its progenitor Adoxa by having a unique and highly complex inflorescence like a spike with several glomerate interrupted clusters, but the ITS sequence divergence between them is only 3·4 % (Liu et al., 2000). The third example involves the plateau endemic Lomatogoniopsis in Gentianaceae (Liu et al., 2001). This genus is distinct from Lomatogonium in having protruding glands at the corolla base and non-vascularized scales at the inner lobes, but very low genetic mutations based on ITS sequences (<2 %) were detected between these genera, despite the distinctive differences in corolla morphology. Both the convergent evolution and random fixation of unique morphological characters might partly explain the substantial inconsistencies among gross morphology, pollen exine pattern and trnL-F phylogeny of Rheum revealed by the present investigation.
The second possible cause of low sequence divergence is cytoplasmic gene flow and chloroplast capture due to ancient or recent hybridization. Ancient introgression could cause replacements of cpDNA-types (Rieseberg and Carney, 1998), leading to the identical or low divergence of trnL-F sequences. More importantly, recent hybridization and introgression could cause morphologically dissimilar species to group together on cytoplasmic DNA phylogeny trees (Sang et al., 1997; Tsukaya et al., 2003). In the present trnL-F phylogeny tree of Rheum, the unexpected position of some species, e.g. the grouping of R. kialense with species with non-cordate leaves, might be due to recent hybridization and introgression. This possibility has been strengthened by the results of ongoing nuclear ITS DNA fragment analysis, showing that individuals of most species have copies of more than one different ITS sequence (A. Wang, M. Yang and Jianquan Liu, unpubl. res.). The nr ITS of plants consists of highly repeated tandemly arranged sequences, and repeats derived from different parental genomes in ancient hybridization events can be used to demonstrate the hybrid origins of some species (Baldwin et al., 1995). The ploidy levels of the species investigated here remain unknown, but new species produced through polyploidization- or diploid-hybridization—would probably have been more likely to survive if they were located in different habitats from their parents (i.e. if the hybrid offspring were allopatrically isolated). The complex topography and diverse habitats of the Qinghai–Tibetan Plateau due to the uplifting and climatic oscillations clearly provided such opportunities.
In conclusion, the present molecular examination of Rheum, a species-rich genus with a diverse array of morphology, provided evidence of a recent adaptive radiation of species with low genetic variation in this genus within a relatively short timeframe. This radiation might be correlated with the extensive habitat changes in the plateau and adjacent areas that followed the large-scale upliftings of the plateau and subsequent climatic oscillations in the Quaternary. These habitat changes may not only have promoted rapid allopatric speciation, but may also have provided opportunities for the production of new species through polyploidization- or diploid-hybridization. Other genera that also have a centre of diversity in the Qinghai–Tibetan Plateau have been shown to have similar patterns to that reported here for Rheum, e.g. Saussurea (Wang and Liu, 2004a, b; Wang et al., 2005) and Nannoglottis (Liu et al., 2002). Therefore, recent diversification and radiation triggered by the uplift of the Qinghai–Tibetan Plateau and subsequent climatic oscillation seem to be common patterns for temperate taxa with the greatest diversity of species in the Qinghai–Tibetan Plateau and adjacent areas. Identifying which mode of speciation contributed most to the diversity of these genera, allopatric or hybridization, remains an interesting issue for further research. For this purpose, low copy nuclear genes with a fast mutation rate might be more promising candidates for study (Ferguson and Sang, 2001). However, the radiation, hybridization, convergent evolution and random fixation of unique characters complicate the establishment of a natural classification system that both reflects the phylogeny of the genus and facilitates identification of its member species for general users.
Acknowledgments
Support for this research was provided Key Innovation Plan KSCX2-SW-106, the Special Fund of Outstanding PhD Dissertation, FANEDD 200327, and the National Science Foundation of China (3000012).
Footnotes
Both authors contributed equally to this work.
LITERATURE CITED
- An ZS, Kutzbach JE, Prell WL, Port SC. 2001. Evolution of Asian monsoons and phased uplift of the Himalayan-Tibetan Plateau since late Miocene times. Nature 411: 62–66. [DOI] [PubMed] [Google Scholar]
- Bakker FT, Culham A, Gomez-Martinez R, Carvalho J, Compton J, Dawtrey R, et al. 2000. Patterns of nucleotide substitution in angiosperm cpDNA trnL (UAA)-trnLF(GAA) regions. Molecular Biology and Evolution 17: 1146–1155. [DOI] [PubMed] [Google Scholar]
- Baldwin BG, Sanderson MJ. 1998. Age and rate of diversification of the Hawaiian silversword alliance (Compositae). Proceedings of the National Academy of Sciences of the USA 95: 9402–9406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baldwin BG, Sanderson MJ, Porter JM, Wojciechowski MF, Campbel CS, Donoghue MJ. 1995. The ITS region of nuclear ribosomal DNA: a valuable source of evidence on angiosperm phylogeny. Annals of the Missouri Botanical Garden 82: 247–277. [Google Scholar]
- Doyle JJ, Doyle JL. 1987. A rapid DNA isolation procedure for small quantities of fresh leaf material. Phytochemistry Bulletin 19: 11–15. [Google Scholar]
- Felsenstein J. 1985. Confidence limits on phylogenies: an approach using the bootstrap. Evolution 39: 783–791. [DOI] [PubMed] [Google Scholar]
- Ferguson D, Sang T. 2001. Speciation though homoploid hybridization between allotetraploids in peonies (Paeonia). Proceedings of the National Academy of Sciences of the USA 98: 3915–3919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Francisco-Ortega J, Santos-Guerra A, Hines A, Jansen RK. 1997. Molecular evidence for a Mediterranean origin of the Macaronesian endemic genus Argyranthemum (Asteraceae). American Journal of Botany 84: 1595–1613. [PubMed] [Google Scholar]
- Friesen N, Fritsch RM, Pollner S, Blattner FR. 2000. Molecular and morphological evidence for an origin of the aberrant genus Milula within Himalayan species of Allium (Alliaceae). Molecular Phylogenetics and Evolution 17: 209–218. [DOI] [PubMed] [Google Scholar]
- Fukuda T, Yokoyama J, Ohashi H. 2001. Phylogeny and biogeography of the genus Lycium (Solanaceae): inferences from chloroplast DNA sequences. Molecular Phylogenetics and Evolution 19: 246–258. [DOI] [PubMed] [Google Scholar]
- Fukuda T, Yokoyama J, Tsukaya H. 2003. Phylogenetic relationships among species in the genera Chisocheton and Guarea that have unique indeterminate leaves as inferred from sequences of chloroplast DNA. International Journal of Plant Sciences 164: 13–24. [Google Scholar]
- Harrison TM, Copeland P, Kidd WSF, Yin A. 1992. Raising Tibet. Science 255: 1663–1670. [DOI] [PubMed] [Google Scholar]
- Hilger HH, Selvi F, Papini A, Bigazzi M. 2004. Molecular systematics of Boraginaceae Tribe Boragineae based on ITS1 and trnL sequences, with special reference to Anchusa s.l. Annals of Botany 94: 201–212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huelsenbeck JP, Ronquist F. 2001. MTBAYES: Bayesian inference of phylogeny. Bioinformatics 17: 754–755. [DOI] [PubMed] [Google Scholar]
- Kadereit JW. 1994. Molecules and morphology, phylogenetics and genetics. Botanica Acta 107: 369–373. [Google Scholar]
- Kao TC, Cheng CY. 1975. Synopsis of the Chinese Rheum Acta Phytotaxonomica Sinica. 13: 69–82. [Google Scholar]
- Korner C. 1999.Alpine plant life—functional plant ecology of high mountain ecosystems. Berlin: Springer-Verlag. [Google Scholar]
- Li AR. 1998.Flora Republicae popularis Sinicae. Beijing: Science Press. [Google Scholar]
- Li JJ, Shi YF, Li BY. 1995.Uplift of the Qinghai-Xizang (Tibet) Plateau and Global Change. Lanzhou: Lanzhou University Press. [Google Scholar]
- Li WH. 1997.Molecular evolution. Sunderland, MA: Sinauer Associates. [Google Scholar]
- Liem KF. 1990. Key evolutionary innovations, differential diversity, and symecomorphosis. In: Nitecki M, ed. Evolutionary innovations. Chicago: University of Chicago Press, 147–170. [Google Scholar]
- Liu JQ, Chen ZD, Lu AM. 2000. The phylogenetic relationships of the Qinghai–Tibet endemic Sinadoxa, revealed by the ITS data. Acta Phytotaxonomica Sinica 42: 656–658. [Google Scholar]
- Liu JQ, Chen ZD, Lu AM. 2001. A preliminary study of the phylogeny of the Swertiinae (Gentianaceae) based on ITS data. Israel Journal of Plant Sciences 43: 301–308. [Google Scholar]
- Liu JQ, Gao TG, Chen ZD, Lu AM. 2002. Molecular phylogeny and biogeography of the Qinghai-Tibet Plateau endemic Nannoglottis (Asteraceae). Molecular Phylogenetics and Evolution 23: 307–325. [DOI] [PubMed] [Google Scholar]
- Losina-Losinskaya AS. 1936. The genus Rheum and its species. Acta Instituti Botanici Academiae Scientiarum Unionis Rerum Publicarum Soveticarum Socialisticarum Ser. 1: 5–141. [Google Scholar]
- Myers N, Mittermeier RA, Mittermeier CG, da Fonseca GAB, Kent J. 2000. Biodiversity hotspots for conservation priorities. Nature 403: 853–858. [DOI] [PubMed] [Google Scholar]
- Ohba H, Malla SB. 1988.The Himalayan plants. Tokyo: University of Tokyo Press. [Google Scholar]
- Omori Y, Ohba H. 1996. Pollen development of Rheum nobile Hook. F. & Thomson (Polygonaceae), with reference to its sterility induced by bract removal. Botanical Journal of the Linnean Society 122: 269–278. [Google Scholar]
- Omori Y, Takayama H, Ohba H. 2000. Selective light transmittance of translucent bracts in the Himalayan giant glasshous plant Rheum nobile Hook. F. & Thomson (Polygonaceae). Botanical Journal of the Linnean Society 132: 19–27. [Google Scholar]
- Posada D, Crandall KA. 1998. MODELTEST: testing the model of DNA substitution. Bioinformatics 14: 817–818. [DOI] [PubMed] [Google Scholar]
- Rambaut A, Charleston M. 2000. TreeEdit version 1.0 alpha 10 (http://evolve.zoo.ox.ac.uk/software/TreeEdit/TreeEdit). [Google Scholar]
- Richardson JE, Pennington RT, Pennington TD, Hollingsworth PM. 2001. Rapid diversification of a species-rich genus of neotropical rain forest trees. Science 293: 2242–2245. [DOI] [PubMed] [Google Scholar]
- Richardson JE, Weitz FM, Fay MF, Cronk QC, Linder HP, Reeves G, Chase MW. 2001. Rapid and recent origin of species richness in the Cape flora of South Africa. Nature 412: 181–183. [DOI] [PubMed] [Google Scholar]
- Rieseberg LH, Carney S. 1998. Plant hybridization. New Phytologist 140: 599–624. [DOI] [PubMed] [Google Scholar]
- Sanderson MJ. 1997. A nonparametric approach to estimating divergence times in the absence of rate constancy. Molecular Biology and Evolution 14: 1218–1232. [Google Scholar]
- Sang T, Crawford J, Stuessy TF. 1997. Chloroplast DNA phylogeny, reticulate evolution, and biogeography of Paeonia (Paeoniaceae). American Journal of Botany 84: 1120–1136. [PubMed] [Google Scholar]
- Shi YF, Li JJ, Li BY. 1998.Uplift and environmental changes of Qinghai–Tibetan Plateau in the late Cenozoic. Guangzhou: Guangdong Science and Technology Press. [Google Scholar]
- Swofford DL. 2000.PAUP*. Phylogenetic analysis using parsimony (*and other methods), Version 4. Sunderland, MA: Sinauer Associates. [Google Scholar]
- Taberlet PT, Gielly L, Patou G, Bouvet J. 1991. Universal primers for amplification of three non-coding regions of chloroplast DNA. Plant Molecular Biology 17: 1105–1109. [DOI] [PubMed] [Google Scholar]
- Tang LY, Shen CM. 1996. Late Cenozoic vegetational history and climatic characteristics of Qinghai-Xizang Plateau. Acta Micropalaeont Sinica 13: 321–337. [Google Scholar]
- Terashima I, Masuzawa T, Ohba H. 1993. Photosynthetic characteristics of a giant alpine plant Rheum nobile Hook. F. & Thomson and of some other alpine species measured at 4300 m, in the eastern Himalayan, Nepal. Oecologia 95: 194–201. [DOI] [PubMed] [Google Scholar]
- Thompson JD, Gibson TJ, Plewinak F, Jeanmougin F, Higgins DG. 1997. The Clustal X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Research 24: 4876–4882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tsukaya H. 2002. Optical and anatomical characteristics of bracts from the Chinese ‘glasshouse’ plant, Rheum alexandrae Batalin (Polygonaceae), Yunnan, China. Journal of Plant Research 115: 59–63. [DOI] [PubMed] [Google Scholar]
- Tsukaya H, Fukuda T, Yokoyama J. 2003. Hybridization and introgression between Callicarpa japonica and C. mollis (Verbenaceae) in central Japan, as inferred from nuclear and chloroplast DNA sequences. Molecular Ecology 12: 3003–3012. [DOI] [PubMed] [Google Scholar]
- Wang WT, Wu SG, Lang KY, Li PQ, Pu FT, Chen SK. 1993.Vascular plants of the Hengduan Mountians, Vol. 1. Pteridophyta, Gymnospermae, Dicotyledoneae (Saussuraceae to Cornaceae). Beijing: Science Press. [Google Scholar]
- Wang YJ, Liu JQ. 2004. Phylogenetic analyses of Saussurea sect. Pseudoeriocoryne (Asteraceae: Cardueae) based on chloroplast DNA trnL-F sequences. Biochemical Systematics and Ecology 32: 1009–1023. [Google Scholar]
- Wang YJ, Liu JQ. 2004. A preliminary investigation on the phylogeny of Saussurea (Asteraceae: Cardueae) based on chloroplast DNA trnL-F sequences. Acta Phytotaxonomica Sinica 42: 136–153. [Google Scholar]
- Wang YJ, Pan JT, Liu SW, Liu JQ. 2005. A new species of Saussurea (Asteraceae) from Tibet in west China and its systematic position according to ITS sequence analysis. Botanical Journal of the Linnean Society 147: 349–356. [Google Scholar]
- Wilson EO. 1992.The diversity of life. Cambridge, MA: Belknap Press of Harvard University. [Google Scholar]
- Wu CY. 1987.Flora of Tibet. Beijing: Science Press. [Google Scholar]
- Wu CY. 1988. Hengduan Mountains flora and its significance. Journal of Japanese Botany 63: 297–311. [Google Scholar]
- Wu CY, Wu SG. 1996. A proposal for new floristic kingdom (realm). In: Zhang AL, Wu SG, eds. Floristic characteristics and diversity of East Asian plants. Beijing: Springer-Verlag, Hongkong China Higher Education Press, 3–42. [Google Scholar]
- Yang MH, Zhang DM, Zheng JH, Liu JQ. 2001. Pollen morphology and its systematic and ecological significance in Rheum (the Rhuburb genus, Polygonaceae) from China. Nordic Journal of Botany 21: 411–418. [Google Scholar]