

Abstract
Eclipta alba (L.) Hassk, Asiasarum sieboldii (Miq.) F. Maek (Asiasari radix), and Panax ginseng C. A. Mey (red ginseng) are traditionally acclaimed for therapeutic properties of various human ailments. Synergistic effect of each standardized plant extract was investigated for hair growth potential on nude mice, as these mutant mice genetically lack hair due to abnormal keratinization. Dried plant samples were ground and extracted by methanol. Topical application was performed on the back of nude mice daily up to completion of two hair growth generations. The hair density and length of Eclipta alba treated mice were increased significantly (P > 0.001) than control mice. Hair growth area was also distinctly visible in Eclipta alba treated mice. On the other hand, Asiasari radix and Panax ginseng treated mice developing hair loss were recognized from the abortive boundaries of hair coverage. Histomorphometric observation of nude mice skin samples revealed an increase in number of hair follicles (HFs). The presence of follicular keratinocytes was confirmed by BrdU labeling, S-phase cells in HFs. Therefore, Eclipta alba extract and/or phytochemicals strongly displayed incomparability of hair growth promotion activity than others. Thus, the standardized Eclipta alba extract can be used as an effective, alternative, and complementary treatment against hair loss.
1. Introduction
Hair is a dynamic mini-organ of mammalian skin that originates from the hair follicle and serves multiple functions throughout adult life. Inside the skin every HF is particularly active and is composed of fully keratinized epithelial cells. Hair growth is coordinated by hormones and it commands the follicle to undergo appropriate changes during this process. This process is followed by specific cyclic order and characterized by anagen (growth phase), catagen (regression), and telogen (resting phase) [1]. Human hair follicles also pose a unique paradox as the hormones, androgens, growth factor may cause stimulation of hair growth. On the other hand, genetically acquired disorder inhibit to generate well differentiated hair follicles or shorten anagen phase; these phenomena may cause baldness pattern [2, 3]. Although human hair loss is not life threatening, alopecia (hair loss/balding) may cause serious psychological distress [4]. These may reflect on some sociopsychological contexts. Novel therapies are therefore required to prevent hair loss and promote hair growth. In this context, developing alternative medicine has increased interest in the recent years. Until now, complete lack of strong scientific evidence of effective component of natural plant extract and limited to their hair promotion mechanisms has kept natural hair care away from mainstream research. Although the search for the treatment hair loss research resulted in some synthetic drugs, associated side effects are highly remarkable. Therefore, herbal products now emerged as an alternate to the synthetic drugs in this current state.
Studies of various research groups reported that hundreds of plants or substances are having potential to promote hair growth as well. In particular only little candidate plant plays great role to enhance hair growth whose efficacies are now under investigation. Therefore, recent research was under taken to investigate of some known plants those were traditionally acclaimed and purported in oriental medicine such as A. radix [5], P. ginseng [6, 7], and E. alba [8, 9] exert to promote hair growth.
Moreover, E. alba has also been reported with multiple medicinal uses and is known for numerous biological activities. It is an enormous source of several secondary metabolites, such as polypeptides, polyacetylenes and triterpenes [10], flavonoids, phytosterols, and coumestans [11]. Phytochemical coumestans represent an important class of natural oxygenated aromatic compound, including wedelolactone and demethylwedelolactone, and saponins, responsible for the main medicinal effects of E. alba, such as its antihepatitis C virus [12], antitumor activity [13], and neuropharmacological activity [14] and its anticancer effect shows antiproliferative and cytotoxic effects determined using MTT assay [15].
Taken together this bioresource information, we undertake nude mice as a model to get more precise result for hair biology from different well-known medicinal plant extracts, as these mice express the role of Foxn1 in mammalian skin biology responsible for structural abnormality of hair shaft due to abnormal keratinization [16]. Moreover, nude mice are also frequently used in skin pharmacology to serve for skin penetration assays [17] as the follicular penetration route acts as an entry pathway for compounds into the skin [18].
E. alba is widely reported traditional medicine and wellbeing functional food, thus it prompted us to evaluate hair growth promoting activity along with other plants. Therefore, this approach may incorporate some novel findings that may promote us to explore extensive bioactive compounds from crude extract for prevention of hair loss as well as enhancing hair growth.
2. Materials and Methods
2.1. Plant Materials and Preparation of Extraction
Aerial parts (leaves, stems, and blooms) of E. alba and dried roots of A. radix and P. ginseng were purchased from the Jecheon Medicinal Herb Association, South Korea. All the samples were authenticated by Associate Professor Ki Hwan Bae (College of Pharmacy, Chungnam National University) where the voucher specimens were deposited. The dried plant sampleswere ground into powder (500 g) and extracted with 95% methanol at 60°C. The methanol extract was concentrated and dried in a vacuum evaporator under reduced pressure below 60°C. The resulting residue was weighed and dissolved in a vehicle mixture (propylene glycol : ethanol : dimethyl sulfoxide, 67 : 30 : 3% v/v). The extract yields were determined as “%” of dry crude extracts (Table 1).
Table 1.
Botanical origins, plant parts, and percentage yields of investigated specimens.
| Botanical name (family) | Chinese name | Common name | Plant part | Extraction yield (% w/w) |
|---|---|---|---|---|
| Eclipta alba (Asteraceae) | Han lian cao | False daisy | Leaves, stems, and blooms | 12.40 |
| Asiasarum sieboldii (Aristolochiaceae) | Shashin | Chinese wild ginger root | Roots | 7.52 |
| Panax ginseng (Araliaceae) | Hong-Shen | Red ginseng | Roots | 38.33 |
2.2. Chromatographic Analysis
HPLC-grade acetonitrile was purchased from Merck Co. (Merck, Darmstadt, Germany). Deionized water was purified by Milli-Q system (Millipore, Bedford, MA, USA). Other chemicals were of reagent grade. Standard wedelolactone powder was purchased from the Sigma-Aldrich Co. The purity of the standard was over 98% as indicated by the manufacturer. Calibration standard samples were prepared from the stock solutions by adding 0.5–1.5 mg/mL of wedelolactone in MeOH to obtain appropriated dilutions which were filtered through Millipore membrane before use. Chromatographic analysis was performed on the Shimadzu (Japan) HPLC apparatus, with a column of Discovery C18 (ODS, 25 cm × 4.6 mm, 5 μm, Supelco, USA) and a Guard Column: Security Guard Cartridge system (Phenomenex, USA). The mobile phase consisted of acetonitrile: water (ACN%, v/v) at a flow rate of 1 mL/min as follows: 20–45% (up to 15 min), 45–100% (up to 30 min), and 100% (up to 40 min). The method was performed with an injection volume of 20 μL and detection wavelength at 365 nm.
2.3. Toxicity Studies
Toxicity studies were carried out by applying the extract to the nude mice skin in concentrations of up to 5% for 7 days and did not show any toxic side effects like erythema on skin surface. Thus, the prepared extracts were considered safe for topical application on nude mice.
2.4. Balb/c Nude Mice
Athymic Balb/c male nude mice at 7 weeks of age were purchased from Dae-Han Biolink (Eumsung, Korea) and maintained under 12 h light: dark periods at 24 ± 2°C and were housed and handled under aseptic conditions. Experiment was performed in accordance with approved institutional protocols by the Institutional Animal Care and Use Committee (IACUC), Chungnam National University, Daejeon, South Korea. The authorization code number is CNU-00244.
2.5. Treatment Protocol of Different Natural Extracts on Nude Mice
Five animals were allocated to five randomized groups for evaluation of hair growth and hair changing pattern of nude mice. Topical application was performed daily with a solution of A. radix, E. alba, and P. ginseng extracts (2.5%) in a vehicle formulation and/or minoxidil (2%) until completion of two full hair growth cycles.
2.6. Measurement of Hair Density and Length
After daily treatment, in each hair growth cycle the images from specific regions of neck, back, and hip were captured two times per week using KONG, Bom-Viewer Plus (Seoul, Korea). About six images of 80× and 200× were exploited to measure the hair length and hair density, respectively, using Bom-Viewer Image Analyzer.
2.7. Evaluation of Hair Existing Area
The hair existing area of nude mice was systematically evaluated for each mouse three times per week. At this time point, mice were scored on a scale of “0” to “4” for percent body coverage of hair “0” = complete alopecia; “1” = <25% hair coverage; “2” = 25–50% hair coverage; “3” = 50–75% hair coverage; “4” = normal hair density and full hair coverage.
2.8. Histologic Assessment of Hair Growth
Mice were sacrificed in anagen phase and skin samples were fixed in 10% buffered formalin for histological analysis. Paraffin-embedded 4 μm sections were stained with hematoxylin and eosin (H&E). The nude mice HF morphology and the number of HFs were evaluated microscopically in 5 fields per section of the dorsal skin at a magnification of ×100.
2.9. BrdU Immunohistochemistry
BrdU labeling cell proliferation was detected by intraperitoneal injection of bromodeoxyuridine (BrdU; Sigma, USA) with 50 mg/kg body weight at a concentration of 5 mg/mL in PBS. BrdU incorporation was detected by IHC staining of paraffin-embedded sections with mouse anti-BrdU primary antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA). Skin sections that were not incubated with primary antibodies were used as a negative control. Detection of follicular keratinocyte BrdU labeling was done at magnification ×100 and ×400.
2.10. Statistical Analysis
Statistical analysis was performed using SAS Systems ver. 9.3, and analysis of variance (ANOVA) was used for comparisons among all groups. All data are expressed as means ± SD unless otherwise is noted. A probability level of 5% was considered significant. Differences between two groups were analyzed by t-test. ANOVA was followed after Duncan's multiple comparison tests.
3. Results
3.1. Differences in Activity of Plant Extracts on Changing Hair Growth Pattern of Nude Mice
After daily treatment, hair growth was documented on experimental day 7. Systematic monitoring of hair growth showed that A. radix (Figure 1(c)) and vehicle treated control mice (Figure 1(a)) exhibit extremely sparse and transient hair coat. On the other hand, minoxidil and E. alba extract treated mice hair growth first documented from the frontal part of the head as early as 7 days and became dense, thicker, and longer that evenly extended to the shoulder and posterior part of the back during experimental day 12. The maximum hair coat was evident in E. alba extract treated group on experimental day 16 (Figures 1(d) and 1(b)). In contrast, A. radix (Figure 1(c)), and P. ginseng (Figure 1(e)) treated mice found rapid hair loss with increasing time point and eventually remained only few sparse, abortive short regional hair that moved caudally to the tail region.
Figure 1.

Stimulation of hair growth by different natural extract in nude mice. Specific concentrations of different extract in vehicle formulation were daily applied topically (n = 5) on the back of nude mice skin (a) control, (b) minoxidil, (c) A. radix, (d) E. alba, and (e) P. ginseng during one hair growth phase.
3.2. Effect on Hair Coverage Area
The effects on hair coverage area of all the treated groups were precisely estimated for each mouse in both first and second hair growth cycle by giving them a score from “0” to “4.” The maximum hair growth score was significantly (P < 0.001) increased in mice treated with minoxidil and E. alba than control group during first and second hair growth cycles (Figure 2). On the other hand, A. radix treated group also consistently effective for inducing hair coverage area (2.25 ± 0.82) in first hair cycle but transient hair coat and progressive hair loss turn to similar score (1.25 ± 0.5) with that of control group (1.25 ± 0.5) in second hair cycle. There was no significant difference in terms of the hair coverage area among P. ginseng and control group.
Figure 2.
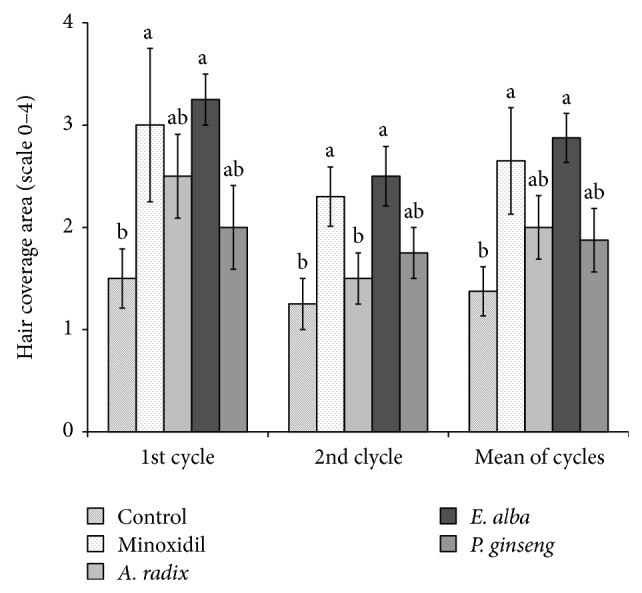
Effect of different plant extract on hair coverage area in athymic nude mice. Mice were evaluated on a scale of “0–4” parameters as follows: “0” = complete alopecia; “1” = <25% hair coverage; “2” = 25–50% hair coverage; “3” = 50–75% hair coverage; “4” = full hair coverage. The different letters (a and b) within a column indicate significant differences (P < 0.05) determined by Duncan's multiple range test.
3.3. Effect on Hair Length and Hair Density
In terms of the measurement of hair length, the remarkable effect was observed in E. alba extract treated group. A significant increase (P < 0.0001) of hair length was observed in both first and second hair growth cycles while being compared with that of control group (Figure 3(F)). Similar effect was also found while evaluating hair density (Figure 3(G)). Although minoxidil treated group showed similar hair density with E. alba treated group (Figure 3(G)), the irregular and deformed hair was found in minoxidil treated mice (Figure 3(B)). On the other hand, E. alba treated mice hair (Figures 3(D) and 3(D′)) was straight and thicker and smother than other treatment groups. Although the hair length of A. radix and P. ginseng extract treated mice increased significantly than control group in earlier hair cycle, it became similar with that of control group afterward (Figure 3(F)). On the other hand, in terms of hair density A. radix treated group found significant effect compared to control but there was no difference of hair density of P. ginseng and control group (Figure 3(G)). Moreover, irregularly distributed single deformed hair shaft was also observed on the skin surface in both A. radix (Figures 3(C) and 3(C′)) and P. ginseng (Figures 3(E) and 3(E′)) treated mice similar to control group (Figures 3(A) and 3(A′)).
Figure 3.

Evaluation of hair length and hair density in cycles. Image of nude mice hair on skin surface, treated with control ((A), (A′)), minoxidil ((B), (B′)), A. radix ((C), (C′)), E. alba ((D), (D′)), and P. ginseng ((E), (E′)) in first and second hair cycles. Hair length (F) and hair density (G) shown in column. The different letters (a, b, c, and d) within a column indicate significant differences (P < 0.0001) determined by Duncan's multiple range test. Digital images were takenby KONG, Bom-Viewer Plus with 80× magnification lens. Hair length and hair density were measured using Bom-Viewer Image analyzer (n = 10).
3.4. Effect of Plant Extracts on the Development and Morphogenesis of Nude Mice HF
Histological examination of the skin specimen revealed differences between control and treated mice in the morphology of the developing HFs. On experimental day 10, minoxidil (Figure 4(B′)), A. radix (Figure 4(C′)), and E. alba treated mice (Figure 4(D′)) showed an early onset of anagen hair morphogenesis compared with that of control (Figure 4(A′)), confirming the macroscopic observation (Figures 4(B), 4(C), and 4(D)). On the other hand, P. ginseng extract treated mice skin specimen (Figure 4(E′)) showed similar stage of hair morphogenesis with that of control mice. Following the guidelines proposed by [19], we determined that the control and P. ginseng treated mice HFs were on stage 6 (Figures 4(A′) and 4(E′)) whereas E. alba extract treated mice HFs were clearly at stage 8 (Figure 4(D′)). Negligible numbers of hair shafts, which emerged from the HFs and moderately atrophic, were also observed in the skin of A. radix, P. ginseng, and vehicle treated mice (Figures 4(C), 4(E), and 4(A)). Moreover, the hair shafts of those were defective (often twisted) with dilation of the hair canal and apparently had insufficient structural rigidity to emerge through the epidermis (Figures 4(A′) and 4(E′)). On the other hand, E. alba treatedmice seemed to have a propensity for follicular accumulation of keratin and structural rigidity to penetrate the epidermis (Figure 4(D)).
Figure 4.

Histological features of the skin specimen of different extract and/or vehicle control treated mice. Macroscopic observation of hair in the skin surface of (A) control, (B) minoxidil, (C) A. radix, (D) E. alba, and (E) P. ginseng in early hair existing phase. Microscopic observation of longitudinal sections of (A′) control, (B′) minoxidil, (C′) A. radix, (D′) E. alba, and (E′) P. ginseng treated mice skin was harvested and fixed in buffered formalin for routine hematoxylin and eosin (H&E) stained histology (n = 7). The broken line indicates defective HFs at the level of sebaceous gland ((A′), (B′), (C′), and (E′)). Note that the straight hair shafts that emerge from the skin surface of minoxidil and E. alba treated mouse ((B), (D) and Arrows (B′), (D′)). Scale bar 100 μm.
3.5. Number of Hair Follicles
Histologic assessment was performed to investigate the progression of hair follicles in nude mice, since there was an increase in the number of hair follicles observed in the deep subcutis during the anagen phase. In the representative transverse sections, the numbers of hair follicles were increased significantly in E. alba (P < 0.0001) and minoxidil (P < 0.005) extract treated groups compared to the control (Figure 5(f)).
Figure 5.

Increase in number of HFs in the dorsal skin of nude mice treated with different plant extract. Representative photomicrographs of the transverse sections of nude mice skin: (a) control, (b) minoxidil, (c) A. radix, (d) E. alba, and (e) P. ginseng, were stained with hematoxylin-eosin (H&E). (f) The number of hair follicles in subcutis with respect to the total number of HFs per field (n = 10). Values are mean ± standard deviation (S.D). The different letters (a, b, c, and d) within a column indicate significant differences (P < 0.0001) determined by Duncan's multiple range test. Scale bar 100 μm.
3.6. BrdU Immunohistochemistry
To further investigate the molecular mechanism underlying the ability of E. alba extract to form nude mice hair follicles, we performed immunohistochemistry with BrdU antibodies of the back skin of nude mice. We found that E. alba treated mice skin exhibits BrdU positive keratinocyte in the HFs, particularly a single cell layer that surrounds the innermost layer of the outer root sheath, as shown in Figures 6(B) and 6(B′). Moreover, E. alba treated nude mice exhibited a significant (P < 0.0001) increase in the number of BrdU-labeled keratinocyte per anagen follicle versus control mice (Figure 6(C)). On the other hand, there was no significant difference in the number of BrdU-labeled epithelial cells per sebaceous gland.
Figure 6.
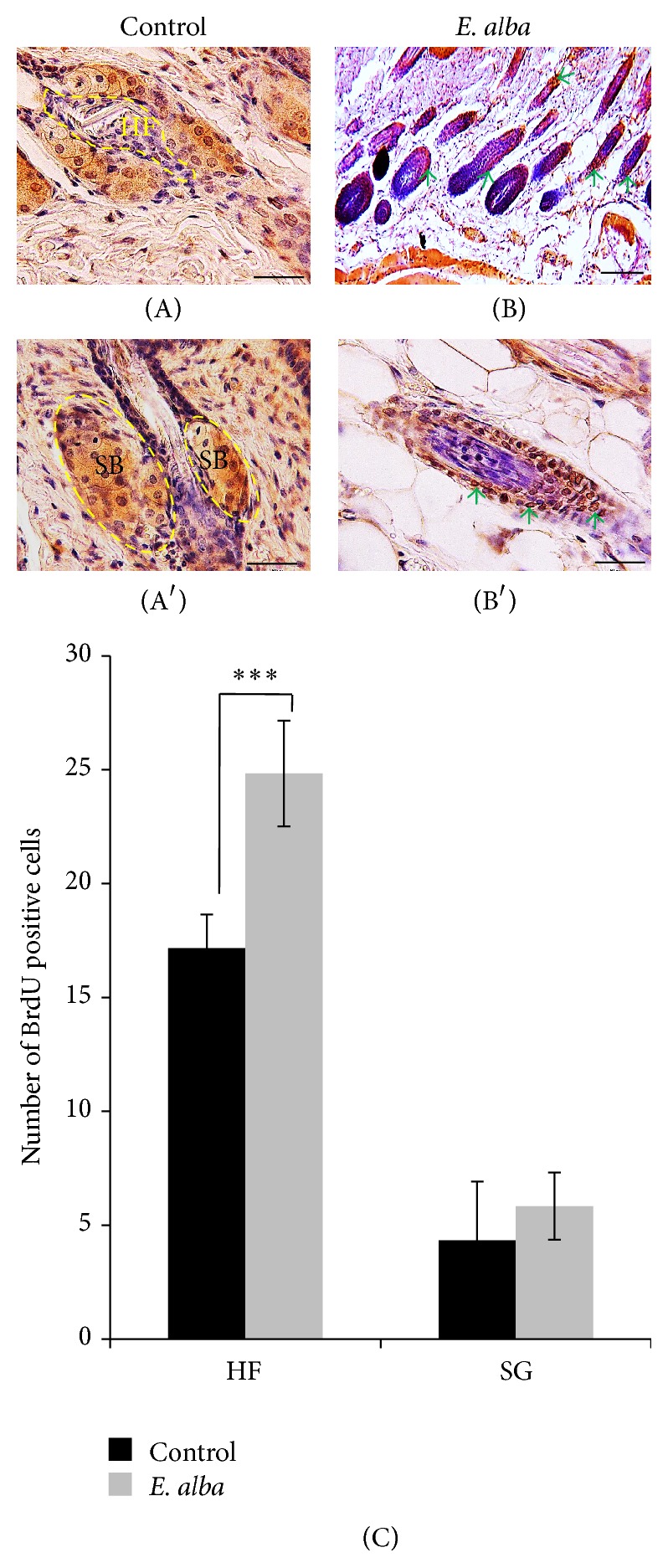
BrdU-labeled proliferating cells in anagen hair follicles. Control (A) and E. alba extract treated (B) follicles are nuclear BrdU staining (Arrows, red-brown). Scale bar, 100 μm (B) and 50 μm ((A), (A′), and (B′)). Number of BrdU positive cells (C). Data shown are means of five replicate samples (*** P < 0.001).
3.7. Identification and Quantification of Wedelolactone in E. alba Extract
Additionally, HPLC analysis was performed to illustrate the methanol extract of E. alba (Figure 7) and was found to contain principle constituents like wedelolactone. The existence of the wedelolactone was quantified by 1.8% in methanol extract. It has been reported that various bioactive phytochemicals found in the E. alba extract, including wedelolactone, demethylwedelolactone, and saponins, are responsible for the main medicinal effects of E. alba, such as antitumor activity [13], antihepatitis C virus [12], and neuropharmacological activity [14] and its anticancer effect shows antiproliferative and cytotoxic effects determined using MTT assay [15].
Figure 7.
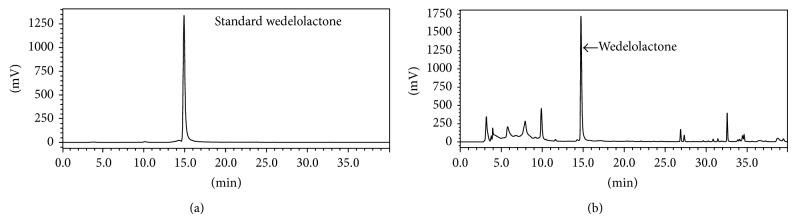
HPLC chromatogram. (a) Standard wedelolactone; (b) HPLC chromatogram of methanol extract of E. alba.
4. Discussion
Medicinal plants may have potential by identifying active biological components and their successful use might open reverse pharmacology, a new approach to drug discovery. Given the known therapeutic properties of E. alba extract, we have performed enormous screening of different plant extracts on Balb/C nude mice model, and this has been used as a unique model to study the hair growth mechanism [20, 21].
Among all the plant extracts, E. alba showed outstanding hair growth promoting potential. Topical application was performed on the back of nude mice skin daily up to completion of two hair growth generations. The macroscopic appearance and the local distribution of hair were precisely observed in each individual group and showed clearly distinguishable feature. The regional distribution of the “abortive” anagen hair pattern in nude strains corresponded to the wave-like pattern of hair generations [22]. The hair growth area of E. alba treated mice is distinctly visible and simultaneously covered the maximum part of the back. In contrast, A. radix, P. ginseng treated mice with increasing age rapidly developed hair loss recognizable from the boundaries of hair covering. However, a few uneven hairs are visible particularly in nude mice skin surface, which referred to as “abortive hair growth” [23]. During anagen growth phase, we were able to observe a comparable growth pattern in E. alba treated mice with other treated and control mice (Figure 1). These phenomena may suggest that E. alba improved the disordered keratinization of nude mouse hair follicles, because original control hair shafts are short and irregular in shape suggesting some disordered keratinization of the HF [24, 25]. The precise genetic defect of nude mice, Foxn1 mutation expression, involved abnormal keratinization as the cause for defective cell type in the hair development [26–28]. Our observation suggests that treatment with E. alba acts transiently to normalize keratinocyte of nude mice HF, leading to fully formed visible HF with increased hair coverage area. An increase in the number and size of HFs has been considered as an indicator for the transition of hair growth from telogen to anagen [9, 19]. These results suggest that topical application of E. alba extract could stimulate the formation of hair follicles and induce an earlier anagen phase, compared to either the control or other treated groups (Figure 4). To elucidate the molecular mechanism underlying the ability of E. alba extract, the BrdU immunohistochemistry result suggests an enhanced keratinocyte proliferation in anagen hair follicles [29]. In our present studies, macroscopic analysis of hair growth parameters included changing pattern of nude mice hair growth, hair coverage area, and hair length. Histological study of different plant extracts along with control and minoxidil groups was investigated and found to have fully formed HFs in E. alba treated group as well as increasing the number of HFs. The active phase of HF cycling is also accompanied by increase in size and number of follicles resulting in an enlargement of the subcutis layer [19]. Given the vital role of hair restorer efficacy of E. alba extract, further HPLC analysis was carried out and showed that wedelolactone and other components exist in methanol extract. In this experiment, none of the plant extracts caused adverse dermatological effects, such as erythema, redness, drying or scaling, and dermatitis as it was case with minoxidil group at the skin site of nude mice.
It has been argued that combination of several constituents of a plant extract may act synergistically rather than single compound [30]. The interaction is possibly due to the binding of more than one compound at different sites on the same target or receptor. Further detailed clinical trials and studies will be necessary to investigate what components in E. alba extract contribute to its efficacy, since whole E. alba extract, rather than individual components, was used here to establish its biological activity against alopecia.
5. Conclusion
E. alba extract containing active ingredients provided more safety and compatibility which has more accessibility in pharmaceutical and cosmetic science and may promote the herbal medicine with minimal or no side effects or toxicities for hair growth therapy. Taking together all the investigation indicates that plant extract with suitable vehicle may be useful as an alternative topical medicine to minoxidil therapy.
Acknowledgments
The authors would like to express special thanks to Dr. Kang Ju Choi, a great generous person, for sharing pearls of wisdom with them which is gratefully acknowledged. This study was supported by a grant from the DBIO Science and Technology Corporation.
Conflict of Interests
The authors declare that there is no conflict of interests regarding the publication of this paper.
References
- 1.Tiede S., Kloepper J. E., Whiting D. A., Paus R. The “follicular trochanter”: an epithelial compartment of the human hair follicle bulge region in need of further characterization. British Journal of Dermatology. 2007;157(5):1013–1016. doi: 10.1111/j.1365-2133.2007.08138.x. [DOI] [PubMed] [Google Scholar]
- 2.Randall V. A. Androgens and human hair growth. Clinical Endocrinology. 1994;40(4):439–457. doi: 10.1111/j.1365-2265.1994.tb02483.x. [DOI] [PubMed] [Google Scholar]
- 3.Park H.-J., Zhang N., Park D. K. Topical application of Polygonum multiflorum extract induces hair growth of resting hair follicles through upregulating Shh and β-catenin expression in C57BL/6 mice. Journal of Ethnopharmacology. 2011;135(2):369–375. doi: 10.1016/j.jep.2011.03.028. [DOI] [PubMed] [Google Scholar]
- 4.Girman C. J., Rhodes T., Lilly F. R. W., Guo S. S., Siervogel R. M., Patrick D. L., Chumlea W. C. Effects of self-perceived hair loss in a community sample of men. Dermatology. 1998;197(3):223–229. doi: 10.1159/000018001. [DOI] [PubMed] [Google Scholar]
- 5.Rho S. S., Park S. J., Hwang S. L., Lee M. H., Chang D. K., Lee I. H., Chang S. Y., Rang M. J. The hair growth promoting effect of Asiasari radix extract and its molecular regulation. Journal of Dermatological Science. 2005;38(2):89–97. doi: 10.1016/j.jdermsci.2004.12.025. [DOI] [PubMed] [Google Scholar]
- 6.Matsuda H., Yamazaki M., Asanuma Y., Kubo M. Promotion of hair growth by ginseng radix on cultured mouse vibrissal hair follicles. Phytotherapy Research. 2003;17(7):797–800. doi: 10.1002/ptr.1241. [DOI] [PubMed] [Google Scholar]
- 7.Park S., Shin W.-S., Ho J. Fructus panax ginseng extract promotes hair regeneration in C57BL/6 mice. Journal of Ethnopharmacology. 2011;138(2):340–344. doi: 10.1016/j.jep.2011.08.013. [DOI] [PubMed] [Google Scholar]
- 8.Roy R. K., Thakur M., Dixit V. K. Hair growth promoting activity of Eclipta alba in male albino rats. Archives of Dermatological Research. 2008;300(7):357–364. doi: 10.1007/s00403-008-0860-3. [DOI] [PubMed] [Google Scholar]
- 9.Datta K., Singh A. T., Mukherjee A., Bhat B., Ramesh B., Burman A. C. Eclipta alba extract with potential for hair growth promoting activity. Journal of Ethnopharmacology. 2009;124(3):450–456. doi: 10.1016/j.jep.2009.05.023. [DOI] [PubMed] [Google Scholar]
- 10.Wagner H., Geyer B., Kiso Y., Hikino H., Rao G. S. Coumestans as the main active principles of the liver drugs Eclipta alba and Wedelia calendulacea . Planta Medica. 1986;5:370–374. [PubMed] [Google Scholar]
- 11.Pereira A. M. S., Bertoni B. W., Menezes A., Jr., Pereira P. S., Franca S. C. Soil pH and production of biomass and wedelolactone in field grown Eclipta alba . Journal of Herbs, Spices and Medicinal Plants. 1998;6(1):43–48. doi: 10.1300/J044v06n01_06. [DOI] [Google Scholar]
- 12.Manvar D., Mishra M., Kumar S., Pandey V. N. Identification and evaluation of anti Hepatitis C virus phytochemicals from Eclipta alba . Journal of Ethnopharmacology. 2012;144(3):545–554. doi: 10.1016/j.jep.2012.09.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Liu Q.-M., Zhao H.-Y., Zhong X.-K., Jiang J.-G. Eclipta prostrata L. phytochemicals: isolation, structure elucidation, and their antitumor activity. Food and Chemical Toxicology. 2012;50(11):4016–4022. doi: 10.1016/j.fct.2012.08.007. [DOI] [PubMed] [Google Scholar]
- 14.Thakur V. D., Mengi S. A. Neuropharmacological profile of Eclipta alba (Linn.) Hassk. Journal of Ethnopharmacology. 2005;102(1):23–31. doi: 10.1016/j.jep.2005.05.037. [DOI] [PubMed] [Google Scholar]
- 15.Chaudhary H., Dhuna V., Singh J., Kamboj S. S., Seshadri S. Evaluation of hydro-alcoholic extract of Eclipta alba for its anticancer potential: an in vitro study. Journal of Ethnopharmacology. 2011;136(2):363–367. doi: 10.1016/j.jep.2011.04.066. [DOI] [PubMed] [Google Scholar]
- 16.Mecklenburg L., Tychsen B., Paus R. Learning from nudity: lessons from the nude phenotype. Experimental Dermatology. 2005;14(11):797–810. doi: 10.1111/j.1600-0625.2005.00362.x. [DOI] [PubMed] [Google Scholar]
- 17.Simon G. A., Maibach H. I. Relevance of hairless mouse as an experimental model of percutaneous penetration in man. Skin Pharmacology and Applied Skin Physiology. 1998;11(2):80–86. doi: 10.1159/000029812. [DOI] [PubMed] [Google Scholar]
- 18.Toll R., Jacobi U., Richter H., Lademann J., Schaefer H., Blume-Peytavi U. Penetration profile of microspheres in follicular targeting of terminal hair follicles. Journal of Investigative Dermatology. 2004;123(1):168–176. doi: 10.1111/j.0022-202X.2004.22717.x. [DOI] [PubMed] [Google Scholar]
- 19.Paus R., Müller-Röver S., van der Veen C., et al. A comprehensive guide for the recognition and classification of distinct stages of hair follicle morphogenesis. Journal of Investigative Dermatology. 1999;113(4):523–532. doi: 10.1046/j.1523-1747.1999.00740.x. [DOI] [PubMed] [Google Scholar]
- 20.Sawada M., Terada N., Taniguchi H., Tateishi R., Mori Y. Cyclosporin A stimulates hair growth in nude mice. Laboratory Investigation. 1987;56(6):684–686. [PubMed] [Google Scholar]
- 21.Watanabe S., Mochizuki A., Wagatsuma K., Kobayashi M., Kawa Y., Takahashi H. Hair growth on nude mice due to cyclosporin A. The Journal of Dermatology. 1991;18(12):714–719. doi: 10.1111/j.1346-8138.1991.tb03162.x. [DOI] [PubMed] [Google Scholar]
- 22.Militzer K. Hair growth pattern in nude mice. Cells Tissues Organs. 2001;168(4):285–294. doi: 10.1159/000047845. [DOI] [PubMed] [Google Scholar]
- 23.Iversen U., Iversen O. H. Cycles of hair growth in hairless mice. Acta Pathologica Microbiologica Scandinavica. 1967;69(1):50–62. [Google Scholar]
- 24.Rigdon R. H., Packchanian A. A. Histologic study of the skin of congenitally athymic “nude” mice. Texas Reports on Biology & Medicine. 1974;32(3-4):711–723. [PubMed] [Google Scholar]
- 25.Hozumi Y., Imaizumi T., Kondo S. Effect of cyclosporin on hair-existing area of nude mice. Journal of Dermatological Science. 1994;7:S33–S38. doi: 10.1016/0923-1811(94)90033-7. [DOI] [PubMed] [Google Scholar]
- 26.Flanagan S. P. “Nude”, a new hairless gene with pleiotropic effects in the mouse. Genetical Research. 1966;8(3):295–309. doi: 10.1017/S0016672300010168. [DOI] [PubMed] [Google Scholar]
- 27.Brissette J. L., Li J., Kamimura J., Lee D., Dotto G. P. The product of the mouse nude locus, whn, regulates the balance between epithelial cell growth and differentiation. Genes and Development. 1996;10(17):2212–2221. doi: 10.1101/gad.10.17.2212. [DOI] [PubMed] [Google Scholar]
- 28.Mecklenburg L., Paus R., Halata Z., Bechtold L. S., Fleckman P., Sundberg J. P. FOXN1 is critical for onycholemmal terminal differentiation in nude (Foxn1 nu) mice. Journal of Investigative Dermatology. 2004;123(6):1001–1011. doi: 10.1111/j.0022-202X.2004.23442.x. [DOI] [PubMed] [Google Scholar]
- 29.Pinkus H., Iwasaki T., Mishima Y. Outer root sheath keratinization in anagen and catagen of the mammalian hair follicle. A seventh distinct type of keratinization in the hair follicle: trichilemmal keratinization. Journal of Anatomy. 1981;133(1):19–35. [PMC free article] [PubMed] [Google Scholar]
- 30.Wagner H., Ulrich-Merzenich G. Synergy research: approaching a new generation of phytopharmaceuticals. Phytomedicine. 2009;16(2-3):97–110. doi: 10.1016/j.phymed.2008.12.018. [DOI] [PubMed] [Google Scholar]