

Abstract
Despite enthusiastic efforts directed at elucidating critical underlying mechanisms towards the identification of novel therapeutic targets for severe acute pancreatitis (SAP), the disease remains without a specific therapy to be executed within the first hours to days after onset of symptoms. Although earlier management for SAP should aim to either treat organ failure or reduce infectious complications, the current standard of care for the general management of AP in the first hours to days after onset of symptoms include intravenous fluid replacement, nutritional changes, and the use of analgesics with a close monitoring of vital signs. Furthermore, repeated evaluation of severity is very important, as the condition is particularly unstable in the early stages. In cases where biliary pancreatitis is accompanied by acute cholangitis or in cases where biliary stasis is suspected, an early endoscopic retrograde cholangiopancreatography is recommended. However, practice guidelines regarding the treatment of pancreatitis are suboptimal. In chronic pancreatitis, conservative management strategies include lifestyle modifications and dietary changes followed by analgesics and pancreatic enzyme supplementation. Recently, attention has been focused on phytoceuticals or antioxidants as agents that could surpass the limitations associated with currently available therapies. Because oxidative stress has been shown to play an important role in the pathogenesis of pancreatitis, antioxidants alone or combined with conventional therapy may improve oxidative-stress-induced organ damage. Interest in phytoceuticals stems from their potential use as simple, accurate tools for pancreatitis prognostication that could replace older and more tedious methods. Therefore, the use of antioxidative nutrition or phytoceuticals may represent a new direction for clinical research in pancreatitis. In this review article, recent advances in the understanding of the pathogenesis of pancreatitis are discussed and the paradigm shift underway to develop phytoceuticals and antioxidants to treat it is introduced. Despite the promise of studies evaluating the effects of antioxidants/phytoceuticals in pancreatitis, translation to the clinic has thus far been disappointing. However, it is expected that continued research will provide solid evidence to justify the use of antioxidative phytoceuticals in the treatment of pancreatitis.
Keywords: Acute pancreatitis, Chronic pancreatitis, Severe acute pancreatitis, Antioxidants, Phytoceuticals
Core tip: In this review, the paradigm shift regarding the development of phytoceuticals and antioxidants is introduced following a comprehensive description of newer information pertaining to the pathogenesis of pancreatitis. Several animal models are discussed with regard to their role in efforts to develop efficient strategies against pancreatitis. Subsequently, newer therapeutic options with an emphasis on nutrients and phytoceuticals are reviewed. Further discussion also focuses on the promise of studies evaluating the effects of antioxidants/phytoceuticals in pancreatitis, the disappointing nature of translation of these agents to clinical settings, and the expected research advances that may support the use of antioxidative phytoceuticals in the treatment of pancreatitis.
INTRODUCTION
Acute pancreatitis (AP) is a relatively common clinical condition, presenting with variable severity from mild and self-limited attacks to severe attacks that contribute to mortality[1]. Severity is associated mechanistically with the underlying pathogenesis of AP which includes pancreatic acinar cell injury in early stages after a local inflammatory reaction, subsequent acinar cell death in the form of apoptosis and necrosis, and the initiation of systemic inflammatory response syndrome (SIRS). An excessive SIRS leads to distant organ damage referred to as multiple organ dysfunction syndrome (MODS)[2]. Recent insights changed a paradigm shift in understanding of AP that intra-acinar trypsinogen activation might lead to early pancreatic injury, but the inflammatory response of AP develops independently driven by early activation of enzyme activation[3]. Whereas, though still effective, the concept that the pancreatic injury is initiated within pancreatic acinar cells subsequent to premature intracellular activation of digestive enzymes and these zymogen activations within acini early during AP was shown to be sufficient to induce AP, finally contributed to the development of chronic pancreatitis[4,5]. Recently, Sah et al[6] found that cerulean-induced chronic pancreatitis (CP) did not require intra-acinar activation of trypsinogen, whereas regulation of the inflammatory response by nuclear factor kappa B (NF-κB) might be involved in the pathogenesis of CP. Collectively, these data suggest a need for the development of novel compounds to either block the early activation of pancreatic enzymes or to ameliorate inflammation in order to limit or prevent complications of AP or inhibit the progression to CP or inflammation-associated fibrosis or carcinogenesis. The delay between the onset of pancreatitis and the development of the systemic response makes AP an ideal experimental and clinical model with which to study the role of inflammatory mediators and to test novel therapies, as the elucidation of the key mediators involved in the pathogenesis of AP will facilitate the development of clinically effective anti-inflammatory therapies[7].
Recent advances in understanding the pathogenesis of pancreatitis-induced SIRS and its complications
AP is an inflammatory disorder, as inflammation not only affects pathogenesis, but also determines the course of the disease from pancreatic acinar cell injury and death to the initiation of SIRS[8]. As excessive SIRS culminates in the primary cause of morbidity and mortality associated with AP, distant organ damage (MODS), it is important to identify the molecules and factors involved in this process. Phospholipase A2 (PLA2), tumor necrosis factor-α (TNF-α), interleukin (IL)-1β, IL-6, IL-8, CINC/GRO-α, MCP-1, platelet activating factor (PAF), IL-10, CD40L, C5a, ICAM-1, MIP1-α, CCL5 (RANTES), substance P, and hydrogen sulfide (H2S) have all been shown to play critical roles[9]. The systemic effects of AP are similar to those of other conditions such as septicemia, severe burns, and trauma. For instances, AP in its severe form is complicated by MODS, most importantly by pulmonary complications which include hypoxia, acute respiratory distress syndrome, atelectasis, and pleural effusion[10].
Novel pathogenic mechanisms relevant to newer therapeutics: Autophagy, apoptosis, and redox-associated transcriptional activators
Autophagy, the principal cellular degradative pathway for cellular protection, is impaired in pancreatitis and is associated with defective lysosomal function[11]. Although research on autophagy in pancreatitis is now in its early stages, it is hoped that data regarding upstream mechanisms mediating autophagic dysfunction and downstream links to pancreatitis pathologies may provide insights into novel molecular targets and therapeutic strategies for the treatment of pancreatitis[12]. In their detailed explanation of a profound dysfunction of key cellular organelles (lysosomes and mitochondria) in pancreatitis, Gukovsky et al[13] described the cause of impaired autophagy in AP and attributed it to inefficient flux resulting from defective lysosomes. Additionally, they suggested that lysosomal dysfunction in pancreatitis could be attributed to either abnormal processing and activation of major lysosomal hydrolases such as cathepsins, or via a decrease in pancreatic levels of the key lysosomal membrane proteins LAMP-1 and LAMP-2. NF-κB inactivation is an additional key pathogenic concern in pancreatitis[14]. NF-κB is a nuclear transcription factor responsible for regulating the transcription of a wide variety of genes involved in immunity and inflammation and plays a critical role in pancreatitis as well as extrapancreatic complications and pancreatic cancer[15]. As seen in several animal models of pancreatitis, NF-κB has been critically implicated in either initiation or propagation of pancreatic inflammations, cerulean-induced pancreatitis[16], taurocholate-induced pancreatitis[17], and arginine-induced pancreatitis[18]. Relevant to autophagy, NF-κB pathway activation stimulated autophagy during induction of acute necrotizing pancreatitis, after which targeted inhibition of the NF-κB pathway may provide novel therapeutic strategies for reducing the severity of pancreatitis[19]. An additional novel mechanism relevant to newer therapeutics involves apoptosis. To test the hypothesis that preventive apoptosis execution would limit the propagation of necro-inflammations in pancreatitis, our group[20] investigated the ability of natural products to induce apoptosis and ameliorate cerulean-induced pancreatitis. Bhatia[21,22] concluded that apoptosis could be a favorable response to acinar cells and that interventions that favor induction of apoptotic, as opposed to necrotic, acinar cell death might reduce the severity of an attack of AP. Aside from pancreatic damage, accelerated acinar cell apoptosis can limit SIRS, as exemplified by honokiol, a low molecular weight natural product similar to Artemisia[23]. The pathogenic roles of transforming growth factor-β (TGF-β) signaling[24], H2S bio-gas, and substance P have also come under scrutiny in order to identify potential therapeutic targets. H2S, which plays important physiologic roles in the cardiovascular, central nervous, and gastrointestinal (GI) systems, has been associated with inflammation, especially gastritis and pancreatitis, through vasomodulation and neuromodulation[25,26]. Substance P, a neuropeptide released from nerve endings after binding to neurokinin-1 (NK-1) receptors on the surface of effector cells, plays important roles in several inflammatory states including asthma, immune-complex-mediated lung injury, experimental arthritis, and inflammatory bowel disease, as well as A/CP through increasing microvascular permeability, promoting plasma extravasation, and mediating pain[27]. Bhatia et al[28] investigated the interplay between the pro-inflammatory effects of H2S and substance P in a murine model of cerulein-induced AP and suggested that the pro-inflammatory effects of H2S may have been be mediated by the substance P-NK-1 receptor pathway in AP. Lastly, oxygen free radicals in excessively high amounts are all very reactive chemically and can impose a detrimental influence on living organisms by provoking oxidative stress that can damage the pancreas[28].
Recent updates on the pathogenesis of CP relevant to pancreatic inflammation
CP is an inflammatory disease of the pancreas characterized by progressive fibrotic destruction of the pancreatic secretory parenchyma. Genetic studies of hereditary, familial, and idiopathic forms of CP have provided much-needed insight into the pathogenesis of CP. The pivotal role of prematurely activated trypsin within the pancreas in the etiology of CP has been firmLy established based on the identification of gain-of-function missense and copy number mutations in the cationic trypsinogen gene and loss-of-function variants in both the pancreatic secretory trypsin inhibitor and chymotrypsinogen C genes. In particular, variants in the gene encoding carboxypeptidase A1, CPA1, were found to be strongly associated with early onset CP[29-31]. Additionally, loss-of-function variants in the cystic fibrosis transmembrane conductance regulator and calcium-sensing receptor genes have also been shown to increase the risk of CP[32]. In addition to these genetic preponderances, necrosis or apoptosis, and inflammation or pancreatic duct obstruction are known to be involved in the pathogenesis of CP. Furthermore, the fibrosing process ultimately leads to progressive loss of the lobular morphology and structure of the pancreas, deformation of the large ducts, and severe changes in the arrangement and composition of the islets. These changes in turn lead to pancreatic insufficiency and predispose patients to changes associated with carcinoma. Irrespective of etiological factors such as heredity, alcohol or nicotine consumption, and nutritional, efferent duct, immunological, and rare metabolic factors, the underlying inflammation and associated subsequent fibrotic destruction of the pancreatic secretory parenchyma are common pathogenic factors in CP that represent targets for prevention through modulation of pancreatic inflammation[33]. Our understanding of CP pathogenesis has improved in recent years through important advances regarding the delineation of mechanisms responsible for the development of pancreatic fibrosis following repeated acute attacks of pancreatic necro-inflammation, also referred to as the necrosis-fibrosis concept[34]. Although steroids can rapidly reduce symptoms in patients with autoimmune CP and micronutrient therapy to correct electrophilic stress is emerging as a promising treatment in the other patients[35], steatorrhea, diabetes, local complications, and psychosocial issues associated with the disease represent additional therapeutic challenges. Such challenges may be resolved in part through intervention with potent anti-inflammatory/anti-oxidative phytoceuticals. In this review, newer therapeutic nutrient-based options and phytoceuticals will be introduced.
ANIMAL MODELS OF PANCREATITIS FOCUSED ON THE DEVELOPMENT OF NEW THERAPEUTICS
Failure to decrease the mortality rate attributable to pancreatitis or improve strategies to prevent CP over the past few decades indicate that current treatment options are limited and predominantly dependent on supportive therapy[36]. Because a key feature of severe AP (SAP) is the presence of extensive tissue necrosis accompanied by inflammatory response syndromes, animal models of AP have become an essential investigative tool for developing potent anti-inflammatory agents. Therefore, a better understanding of the underlying pathophysiology of SAP may lead to more targeted therapeutic options, potentially leading to improved survival. Diverse animal models of AP, from the non-invasive gene knockout and L-arginine models as well as the hormone [cerulenin as a cholecystokinin (CCK) analog]-, alcohol-, and immune-mediated-diet [choline deficient, ethionine supplemented, (CDE)]-induced models, to invasive models including pancreatic duct ligation (PDL), antegrade pancreatic duct perfusion, biliopancreatic duct injection of sodium taurocholate, combination of secretory hyperstimulation with minimal intraductal bile acid exposure, vascular-induced, ischaemia/reperfusion and duct ligation, are available[37] (Table 1). Potential therapeutics can be developed with these animal models, as they share common aspects including the aforementioned pathogenesis of intracellular chemical activation, pancreatic secretion reflux, intracellular production of reactive oxygen species (ROS), and intracellular production of free radicals. As in CP, a special focus on pancreatic duct ligation, repetitive overstimulation with cerulein and chronic alcohol feeding, as well as specific genetic models has been applied[38]. In this review, we will describe some of the animal models used in our efforts to develop efficient strategies against pancreatitis.
Table 1.
Rodent model to study acute and chronic pancreatitis
| Acute pancreatitis |
| Cerulein ± lipopolysaccharide (LPS) or ethanol |
| Bile salt duct infusion |
| Duct obstruction ± secretagogues |
| Diet [choline-deficinet ethionine-supplemented (CDE)] |
| Cytokines |
| Coxsackie virus group B (CVB) |
| Chronic pancreatitis |
| Cerulein (repeated dosing) |
| Alcohol |
| Duct infusion such as trinitrobenzene sulfonic acid or sodium taurocholate or dibutyltin dichloride |
| Duct obstruction |
| Genetic; Cox-2, CFTR, IKK2, LXRb, PERK, TGF-β1 |
| Immunologic |
| Diet (CDE) |
| CVB |
Cerulein-induced pancreatitis
Intravenous infusion of the synthetic CCK analog cerulean at a dose of 0.25 μg/kg per hour causes maximal stimulation of pancreatic exocrine secretion[39]. The infusion of supramaximal doses of cerulein (5 μg/kg per hour and 10 μg/kg per hour) induces a significant increase in pancreatic enzymes in blood, as well as interstitial edema and inflammatory cell infiltration that leads to cerulein-induced edematous pancreatitis in rats, mice, dogs, and hamsters. Aside from intravenous infusion, repeated intraperitoneal injections can also be used to induce pancreatitis. In the early phase, large autophagic vacuoles result from fusion of zymogen granules, accompanied by an increase in lysosomal enzyme activity and activation of trypsinogen. However, since the degree of pancreatitis is generally mild, all animals survive the induction of pancreatitis and resolve completely within 6 d after induction. This model of experimental pancreatitis favors the analysis of intracellular events in the early phase of pancreatitis as seen in Figure 1A, which shows edematous pancreatitis, however, the addition of lipopolysaccharide injection or bile duct ligation can to worsen simple edematous mild pancreatitis as well as oxidative stress and result in acute hemorrhagic pancreatitis[40-42].
Figure 1.
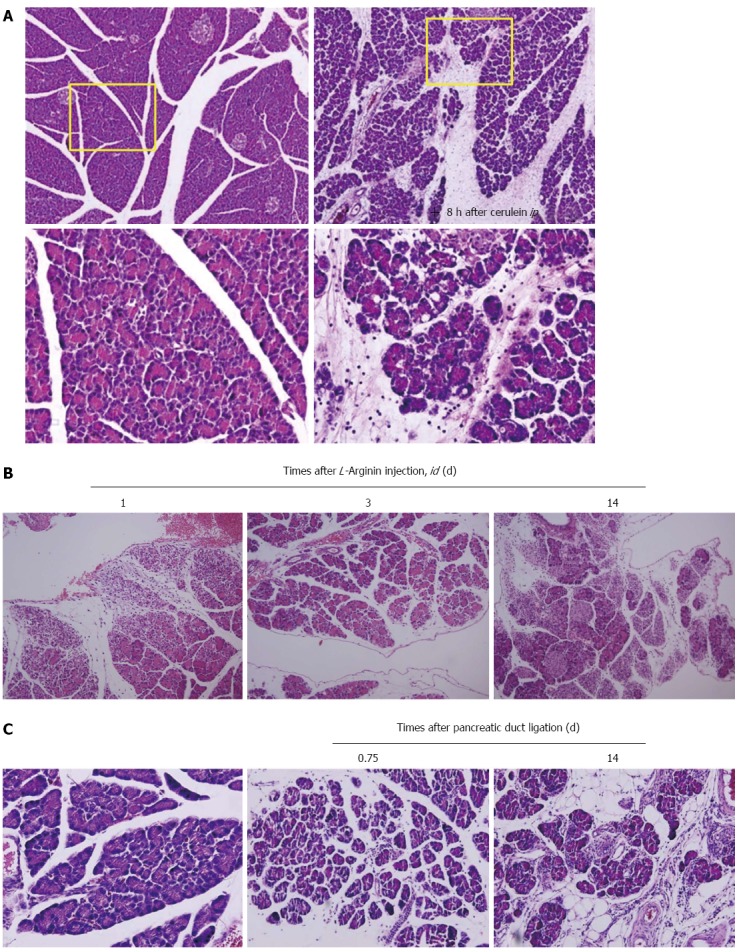
Animal models for pancreatitis. A: Cerulein-induced edematous pancreatitis. Caerulein-induced pancreatitis is a valuable experimental model for studying altered intracellular transport, compartmentation of lysosomal, and digestive enzymes, resulting in edematous pancreatitis. The formation of enlarged secretory vacuoles containing lysosomal and digestive enzymes is paralleled by the activation of lysosomes and degradation of cellular organelles in autophagosomes. On the level of secretory and autophagic vacuoles, activation of serine proteases occurs, which in addition to increasing lysosomal enzyme activities can represent the initial stage for acinar cell destruction and the development of pancreatitis; B: L-arginine-induced necrotizing pancreatitis. Parenchymal hemorrhage and widespread acinar cell necrotic changes were noted with L-arginine; C: Pancreatic duct ligation-induced pancreatitis. Morphologic examination of the pancreas showed massive interstitial edema, apoptosis, and necrosis of acinar cells with infiltration of neutrophil granulocytes and monocytes 0.75 d after pancreatic duct ligation. Two weeks later after periodontal ligament, the destructed parenchyma with fat replacement as well as some fibrotic changes were seen.
Sodium taurocholate infusion; intraparenchymal or intrapancreatic ductal injection
Paran et al[43] are credited with the initial attempt to develop acute necrotizing pancreatitis through intraparenchymal injection of sodium taurocholate in rats. Sodium taurocholate was injected at a dose of 0.3 mL/100 g body weight in concentrations of 5% and 10% into the pancreatic parenchyma of 32 Wistar rats. Early pathological changes observed in the pancreas were focal hemorrhages, parenchymal necrosis, and neutrophil infiltration and at 72 h, the changes observed were acinar necrosis, edema, fibrin deposition and inflammatory cell infiltration. At later time points, changes such as fibrinoid necrosis and fibroblast proliferation were observed[44]. High-pressure infusion of sodium taurocholate into the bilio-pancreatic duct of rats resulted in significant pancreatic and lung alterations[45]. Taurocholate-induced pancreatitis is therefore a reliable model for severe necrotizing pancreatitis in mice with significantly greater pancreatic damage and systemic inflammatory responses as compared to cerulein-induced pancreatitis and correlate with the clinical observations of multisystem organ failure in AP and early changes in affected organs, suggesting that careful observation should be mandatory in patients with AP in order to institute supportive treatment[46].
L-arginine-induced pancreatitis
In 1984, Hegyi et al[47] developed a new type of experimental necrotizing pancreatitis model in rats through the use of a high dose of L-arginine administered via intraperitoneal administration. This non-invasive model is highly reproducible and produces selective, dose-dependent acinar cell necrosis. Not only is this a good model to study the pathogenic mechanisms of acute necrotizing pancreatitis, but it is also excellent with regard to observing and influencing time course changes of the disease (Figure 1B). Subsequent intraperitoneal injection of 3 g/kg L-ornithine caused SAP and higher doses (4 to 6 g/kg) were lethal within hours[48]. Serum and ascitic amylase activities were significantly increased and the increase in pancreatic trypsin activity correlated with the degradation of IκB proteins and elevated IL-1β levels. Oxidative stress in the pancreas was evident from 6 h, making this a simple, noninvasive model of acute necrotizing pancreatitis in rats via intraperitoneal injection of 3 g/kg L-ornithine. Compared with L-arginine, L-ornithine was even more effective in inducing pancreatitis. It should be noted that large doses of L-arginine produce a toxic effect on the pancreas attributable, at least in part, to the actions of L-ornithine.
PDL
AP may be induced by ligating the distal bile duct at the level of the duodenum, which causes the early development of AP, obstructive jaundice and cholangitis in animals. The duct ligation model was developed in an attempt to resemble clinical conditions including gallstone formation, motility disorders of the sphincter, edema and strictures at the papilla, tumors of the papilla, and parasites impacting the terminal biliopancreatic duct. However, surgical ligation of the pancreatic duct alone usually causes only a mild to moderate degree of pancreatitis and has not been successful in inducing SAP. Instead, most laboratory animals developed chronic lesions in the pancreas characterized by atrophy and apoptosis of acinar and ductal tissue without significant necrosis or inflammation. Human CP is characterized by irreversible fibrosis, whereas pancreatic fibrosis in animal models is reversible (Figure 1C). Miyauchi et al[49] compared CP with fibrosis in three different animal models, the dibutyltin dichloride model, WBN/Kob rats, and PDL rats, and found that an imbalance between the synthesis and degradation of extracellular matrix molecules or the degree of stimulation over a certain period may lead to pancreatic fibrosis.
CDE diet-induced necrotizing pancreatitis
Female albino mice were fed a choline-deficient diet containing 0.5% DL-ethionine which was lethal within 5 d due to the development of an acute hemorrhagic pancreatitis accompanied by massive fat necrosis throughout the peritoneal cavity[50]. Major findings included the accumulation of zymogen granules, vacuolation due to foci of cytoplasmic degradation, and alterations in the morphology of the zymogen granules (Figure 2A). Pancreatitis appeared to be due to the intraparenchymal activation of zymogens resulting from a synergistic action of choline deficiency with the basic toxicity of ethionine toward the acinar cells of the pancreas. Because this experimental model simulated the acute hemorrhagic pancreatitis with fat necrosis that occurs in humans, it may prove useful for exploring the pathogenesis of severe pancreatitis with SIRS (Figure 2B)[51]. The diet model appears to be a good approximation of severe necrotizing human pancreatitis as well as CP with histological and biochemical similarities. Both the gross and histological appearance of the pancreatic and peripancreatic inflammation, as well as the clinical and biochemical course of diet-induced pancreatitis, resembled human disease and should be suitable for evaluation of potential clinically-applicable drugs[52]. For example, our group developed ND-07, a novel drug candidate with potent antioxidative and anti-inflammatory properties, that effectively prevented necrotizing pancreatitis[53].
Figure 2.
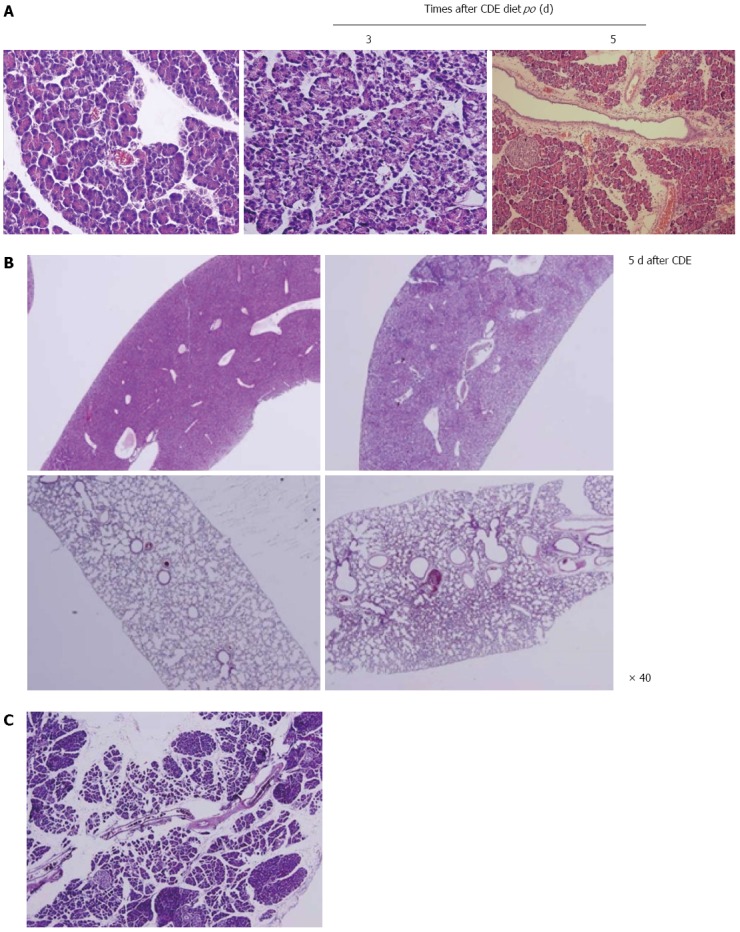
Animal model of choline-deficient, ethionine-supplemented diet-induced necrotizing pancreatitis. A: Choline-deficient, ethionine-supplemented (CDE) diet-induced necrotizing pancreatitis. Massive destruction of pancreatic parenchyma with focal necrotic foci was seen; B: Systemic inflammatory response syndrome hepatic necrosis and pneumonitis was seen; C: Chronic fibrosing pancreatitis was noted 2 mo after CDE diet administration.
Animal models for CP
Since CP is defined as a continuous or recurrent inflammatory disease of the pancreas characterized by progressive and irreversible morphological changes, pancreatitis followed by perilobular and intralobular fibrosis of the parenchyma, calcifications in the parenchyma as well as the formation of pseudocysts[49]. Therefore, animal models of CP are not different from AP models, but need to overcome the acute fatal status according to models, adopting chronic PDL, repetitive overstimulation with cerulean, chronic alcohol feeding, and chronic caring of L-arginine or CDE diet model. However, as seen in Figure 2C, irreversible fibrosis and pancreatic insufficiency following repeated acute attacks of pancreatic necro-inflammation[34], is accompanied.
LIMITATION OF CURRENT PHARMACOLOGIC TREATMENT OF ACUTE AND CHRONIC PANCREATITIS
AP and SAP
Though AP is a disease of variable severity that can lead to significant morbidity and mortality, current management has remained limited to only supportive measures and the treatment of complications. A myriad of pharmacologic therapies targeting various aspects of the underlying pathophysiology have been evaluated and tried over the last few decades, including anti-secretory agents, protease inhibitors, antioxidants, immunomodulators, non-steroidal anti-inflammatory drugs, and prophylactic antibiotics. Only a few of these therapies have demonstrated promise in significantly altering the progression of this disease, and therefore, further studies are necessary to clearly elucidate these benefits in patients at risk for poor outcomes[54]. Regarding pharmacological prevention and treatment of AP, Bang et al[55] reported that somatostatin and octreotide inhibited the exocrine production of pancreatic enzymes and may therefore be useful as prophylaxis against post ERCP pancreatitis (PEP). Though the protease inhibitor gabexate mesilate has been used routinely as treatment for pancreatitis in some countries, randomized clinical trials and a meta-analysis have not supported this practice. Recently, the NSAIDs indomethacin and diclofenac have showed some potential as prophylaxis against PEP in randomized studies. Antibodies against TNF-α have been suggested as a potential rescue therapy, however, no clinical trials are being conducted at present[56].
Chronic fibrosing pancreatitis
Because exocrine pancreatic insufficiency has been associated with changes in GI intraluminal pH, motility disorders, bacterial overgrowth, and altered pancreatic gland secretions, drug absorption in patients with CP may be affected by the degree of CP severity[57]. Furthermore, the general health condition of CP patients is often quite poor, as most patients with CP limit their food intake due to the pain caused by eating and in some cases food intake may be more or less substituted with alcohol, tobacco and coffee. However, pancreatic fibrosis is a characteristic feature of chronic pancreatic injury, which is a result of the imbalance between the synthesis and degradation of extracellular proteins. As stellate cells are pivotal cells implicated in the TGF-β induction of collagens, our previous studies confirmed that antioxidant or antioxidative phytoceuticals ameliorated the progression of fibrosing pancreatitis through suppressive actions on pancreatic stellate cells.
APPLICATION OF ANTIOXIDATIVE PHYTOCEUTICALS TO AMELIORATE AP AND CP
Resveratrol
Resveratrol, a natural polyphenolic compound, was first discovered in the 1940s. Although initially used for cancer therapy, it has shown beneficial effects against most cardiovascular, cerebrovascular, and several inflammatory diseases[58]. It is found in diverse forms of plant life, notably berry fruits, has positive effects on metabolism, and can increase the lifespans of various organisms. The effects of resveratrol have been attributed to its capacity to interact with multiple molecular targets involved in diverse intracellular pathways. One of the more well-known resveratrol interactions involves the activation of sirtuins, a class of NAD(+)-dependent deacetylases, and subsequent HDAC inhibition that affects multiple transcription factors and other protein targets[59,60]. The intracellular pathways activated are crucial for anti-oxidant defense, regulation of the cell cycle, mitochondrial energy production, vascular tone, oncogene suppression, and many other phenomena. Meng et al[61] investigated whether resveratrol could effectively inhibit the expression of NF-κB activation, alleviate the severity of SAP through its anti-inflammatory effects, and regulate inflammatory mediators. A study by Ma et al[62] found that the beneficial outcomes attributable to resveratrol were closely associated with anti-inflammatory, antioxidant, and chemopreventive effects, as well as the inhibition of platelet aggregation, in SAP. Through these effects, resveratrol was able to down-regulate pro-inflammatory cytokines, improve microcirculation, modulate cell apoptosis, and block calcium overload. Additionally, resveratrol inhibited NF-κB activity and reduced concentrations of TNF-α, IL-6 and IL-1β. It also regulated calcium and scavenged ROS capable of extensive tissue damage on extrapancreatic organs[63]. Furthermore, resveratrol has been shown to ameliorate SIRS by improving underlying lung microcirculation dysfunction through decreasing leukocyte-endothelial interactions, reducing blood viscosity, improving the decrease in blood flow, and stabilizing erythrocytes in SAP rats[61] and inactivated intraperitoneal macrophages[64].
Artemisia extracts
Oxygen free radicals (ORFs) mediate an important step in the initiation of experimental AP. Additionally, several clinical findings have implicated OFRs as possible contributors to the pathogenesis of pancreatic fibrosis. To date, there are no studies reporting potential roles for OFRs in the development of CP with the prevention with antioxidants. Yoo et al[65] conducted a study designed to establish a mouse model of chronic fibrosing pancreatitis and to prove the involvement of OFRs in CP with fibrosis. Repeated intraperitoneal injection of cerulein provoked significant and severe chronic fibrosing pancreatitis after 5 wk. Following treatment with Artemisia extracts, the extent of pancreatic fibrosis was significantly decreased, as was the degree of pancreatic inflammation. Furthermore, the level of NF-κB binding activity, which was increased in CP, was significantly attenuated after Artemisia extract treatment (Figure 3A). The levels of myeloperoxidase and iNOS activities were also significantly decreased in the Artemisia-treated group as compared to the pancreatitis only group. Conversely, cytoprotective proteins such as heat shock protein-70 and metallothionein were significantly increased in the Artemisia-treated group. In addition, Artemisia decreased the expression of alpha-SMA and type I collagen in cultured pancreatic stellate cells.
Figure 3.
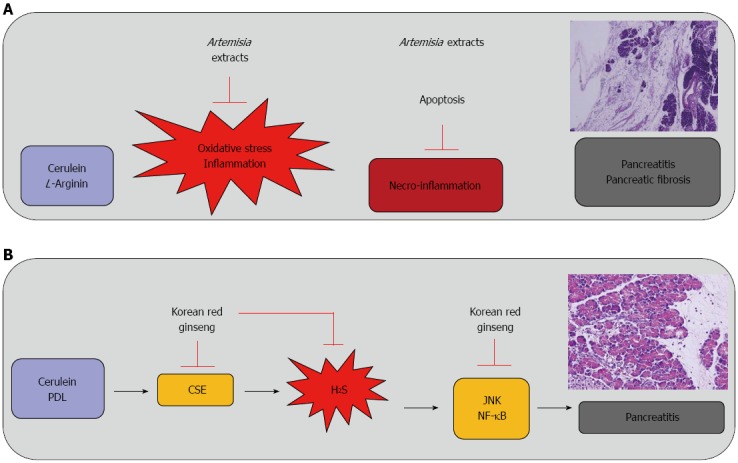
Therapeutic and preventive effect of antioxidative phytoceuticals, Artemisia extract and Korean red ginseng against pancreatitis. A: Therapeutic effect of Artemisia extracts against cerulean or L-arginine-induced pancreatitis and chronic fibrosing pancreatitis; B: Korean red ginseng to ameliorate hydrogen sulfide (H2S)-induced pancreatitis. NF-κB: Nuclear factor kappa B; PDL: Periodontal ligament; CSE: Cystathionine γ-lyase.
Other potential phytoceuticals from nature
There have been published reports describing successful trials demonstrating the beneficial preventive or therapeutic effects of phytoceuticals in diverse animal models of pancreatitis. As examples, rhubarb has been shown to significantly attenuate SAP by inhibiting activation of MAPKs and the expression of inflammatory mediators in taurocholate-induced pancreatitis[66], Nardostachys jatamansi has been implicated as potentially protective in cerulean-induced pancreatitis via the induction of HO-1 expression[67], and Curcuma longa has also been implicated as potentially protective against cerulein-induced AP and pancreatitis-associated lung injury via significant attenuation of inflammatory mediators such as IL-1β and TNF-α[68]. Additional examples include the anti-inflammatory roles observed for cannabidiol and O-1602, the ligands of G protein-coupled receptor 55, in cerulein-induced AP in mice[69] and the protective effects of Scolopendra subspinipes mutilans water extract in cerulean-induced pancreatitis via the deactivation of c-Jun NH2-terminal kinase, p38, and NF-κB and subsequent inhibition of high-mobility group box protein-1[70], Furthermore, attenuation of cerulein-induced AP by apamin, a component of bee venom, or α-pinene, has been observed and attributed to JNK inhibition[71,72] and amelioration of AP by Dachengqi decoction has been observed and attributed to regulation of the necrosis-apoptosis switch in the pancreatic acinar cell and rat models[73,74]. Protective effects of three Chinese herbal medicines containing ligustrazine, kakonein, and Panax notoginsenosides have been demonstrated on multiple organs in rats with SAP[75] and protective effects of baicalin and octreotide have also been demonstrated on multiple organ injury in SAP[76]. Beneficial pancreatic repair effects have been shown following the use of Emblica officinalis, a medicinal plant native to India, or melatonin in L-arginine-induced AP in rats[77,78]. An improving effect of pentoxifylline and/or alpha lipoic acid on L-arginine-induced SAP has also been described and attributed to antioxidant and anti-inflammatory actions[79]. Other research has shown effects of Korean red ginseng on superoxide dismutase inhibitor-induced pancreatitis in rats through inhibition of NF-κB[80] and the efficacy of Salvia miltiorrhizae injection in the treatment of rats to promote Bax-mediated apoptosis in SAP[81].
Antioxidants in the treatment of pancreatitis
Oxidative stress plays an important role in the pathogenesis of both AP and CP. Although its impact has been well documented and has been studied clinically in CP, it is less well defined in SAP. In their study of the pathophysiological aspects of oxidative stress in AP, Hackert and Werner[82] showed that ROS not only participated in the inflammatory cascade, but also mediated inflammatory cell adhesion and consecutive tissue damage. Furthermore, ROS are known to be involved in the generation of pain, an additional important clinical feature of patients suffering from AP. Mechanistically, oxidative stress activates NF-κB, resulting in up-regulation of inflammatory cytokines in pancreatic acinar cells[83]. This mechanism suggests that small-molecule antioxidants may be clinically useful anti-inflammatory agents via inhibition of oxidant-induced cytokine production[84]. Similarily, the antioxidant pyrrolidine dithiocarbamate significantly attenuated SAP through inhibition of HMGB1[85] and raxofelast, an inhibitor of lipid peroxidation, significantly reduced NF-κB activation and attenuated cerulean-induced pancreatitis[86]. The potent antioxidant and anti-inflammatory functions of melatonin have also been demonstrated through their ability to ameliorate cerulean-induced pancreatitis by modulating the actions of Nrf2 and NF-κB[87].
KOREAN RED GINSENG TO AMELIORATE PANCREATITIS VIA SUPPRESSION OF H2S
Korean red ginseng (KRG) has been reported to reduce the risk of inflammation in diverse organs. In our previous studies[88], we demonstrated significant inhibitory actions of KRG on Helicobacter pylori-induced H2S synthesis and the pathogenic connections between H2S synthesis and development of pancreatitis. Therefore, KRG may be a good example of a natural antioxidative phytoceutical for use in ameliorating AP through the inhibition of H2S synthesis. In one of our recent studies that tested the hypothesis that KRG prevents pancreatitis by mitigating H2S generation and pancreatic inflammation, we performed in vitro experiments to document the inhibitory effects of KRG on H2S-associated inflammation in pancreatic cells and in vivo experiments to document the therapeutic effect of KRG on cerulean-induced and PDL-induced AP. KRG was administered at a dose of 200 mg/kg 16 h and 1 h before the first cerulein injection and at a dose of 500 mg/kg 2 h and 4 h after the first cerulein injection by oral gavage. In the mice treated with KRG, pancreatic injuries as evidenced by pancreatic wet weight, histological examinations, serum levels of amylase and lipase, myeloperoxidase activities, serum and pancreatic levels of IL-6, immunohistochemical staining of F4/80 for infiltrating macrophages, and H2S synthesis, were all significantly ameliorated (Figure 3B). The novel finding that KRG decreased PDL-induced hyperamylasemia encouraged us to explore the possibility that KRG pretreatment may prevent ERCP-induced hyperamylasemia. These experiments are ongoing in our clinic.
Footnotes
Supported by The 2013 grant from the Korean Society of Ginseng funded by Korean Ginseng Cooperation
P- Reviewer: Masamune A, Wronski M S- Editor: Gou SX L- Editor: A E- Editor: Ma S
References
- 1.Bhatia M, Wong FL, Cao Y, Lau HY, Huang J, Puneet P, Chevali L. Pathophysiology of acute pancreatitis. Pancreatology. 2005;5:132–144. doi: 10.1159/000085265. [DOI] [PubMed] [Google Scholar]
- 2.Sah RP, Garg P, Saluja AK. Pathogenic mechanisms of acute pancreatitis. Curr Opin Gastroenterol. 2012;28:507–515. doi: 10.1097/MOG.0b013e3283567f52. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Sah RP, Dawra RK, Saluja AK. New insights into the pathogenesis of pancreatitis. Curr Opin Gastroenterol. 2013;29:523–530. doi: 10.1097/MOG.0b013e328363e399. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Thrower E, Husain S, Gorelick F. Molecular basis for pancreatitis. Curr Opin Gastroenterol. 2008;24:580–585. doi: 10.1097/MOG.0b013e32830b10e6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Vonlaufen A, Wilson JS, Apte MV. Molecular mechanisms of pancreatitis: current opinion. J Gastroenterol Hepatol. 2008;23:1339–1348. doi: 10.1111/j.1440-1746.2008.05520.x. [DOI] [PubMed] [Google Scholar]
- 6.Sah RP, Dudeja V, Dawra RK, Saluja AK. Cerulein-induced chronic pancreatitis does not require intra-acinar activation of trypsinogen in mice. Gastroenterology. 2013;144:1076–1085.e2. doi: 10.1053/j.gastro.2013.01.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Bhatia M, Brady M, Shokuhi S, Christmas S, Neoptolemos JP, Slavin J. Inflammatory mediators in acute pancreatitis. J Pathol. 2000;190:117–125. doi: 10.1002/(SICI)1096-9896(200002)190:2<117::AID-PATH494>3.0.CO;2-K. [DOI] [PubMed] [Google Scholar]
- 8.Bhatia M. Acute pancreatitis as a model of SIRS. Front Biosci (Landmark Ed) 2009;14:2042–2050. doi: 10.2741/3362. [DOI] [PubMed] [Google Scholar]
- 9.Bhatia M, Moochhala S. Role of inflammatory mediators in the pathophysiology of acute respiratory distress syndrome. J Pathol. 2004;202:145–156. doi: 10.1002/path.1491. [DOI] [PubMed] [Google Scholar]
- 10.Browne GW, Pitchumoni CS. Pathophysiology of pulmonary complications of acute pancreatitis. World J Gastroenterol. 2006;12:7087–7096. doi: 10.3748/wjg.v12.i44.7087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Gukovskaya AS, Gukovsky I. Autophagy and pancreatitis. Am J Physiol Gastrointest Liver Physiol. 2012;303:G993–G1003. doi: 10.1152/ajpgi.00122.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Fortunato F, Bürgers H, Bergmann F, Rieger P, Büchler MW, Kroemer G, Werner J. Impaired autolysosome formation correlates with Lamp-2 depletion: role of apoptosis, autophagy, and necrosis in pancreatitis. Gastroenterology. 2009;137:350–60, 360.e1-5. doi: 10.1053/j.gastro.2009.04.003. [DOI] [PubMed] [Google Scholar]
- 13.Gukovsky I, Pandol SJ, Mareninova OA, Shalbueva N, Jia W, Gukovskaya AS. Impaired autophagy and organellar dysfunction in pancreatitis. J Gastroenterol Hepatol. 2012;27 Suppl 2:27–32. doi: 10.1111/j.1440-1746.2011.07004.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Rakonczay Z, Hegyi P, Takács T, McCarroll J, Saluja AK. The role of NF-kappaB activation in the pathogenesis of acute pancreatitis. Gut. 2008;57:259–267. doi: 10.1136/gut.2007.124115. [DOI] [PubMed] [Google Scholar]
- 15.Shi C, Zhao X, Wang X, Andersson R. Role of nuclear factor-kappaB, reactive oxygen species and cellular signaling in the early phase of acute pancreatitis. Scand J Gastroenterol. 2005;40:103–108. doi: 10.1080/00365520410009555. [DOI] [PubMed] [Google Scholar]
- 16.Hahm KB, Im YH, Lee C, Parks WT, Bang YJ, Green JE, Kim SJ. Loss of TGF-beta signaling contributes to autoimmune pancreatitis. J Clin Invest. 2000;105:1057–1065. doi: 10.1172/JCI8337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Vaquero E, Gukovsky I, Zaninovic V, Gukovskaya AS, Pandol SJ. Localized pancreatic NF-kappaB activation and inflammatory response in taurocholate-induced pancreatitis. Am J Physiol Gastrointest Liver Physiol. 2001;280:G1197–G1208. doi: 10.1152/ajpgi.2001.280.6.G1197. [DOI] [PubMed] [Google Scholar]
- 18.Rakonczay Z, Jármay K, Kaszaki J, Mándi Y, Duda E, Hegyi P, Boros I, Lonovics J, Takács T. NF-kappaB activation is detrimental in arginine-induced acute pancreatitis. Free Radic Biol Med. 2003;34:696–709. doi: 10.1016/s0891-5849(02)01373-4. [DOI] [PubMed] [Google Scholar]
- 19.Yang S, Bing M, Chen F, Sun Y, Chen H, Chen W. Autophagy regulation by the nuclear factor κB signal axis in acute pancreatitis. Pancreas. 2012;41:367–373. doi: 10.1097/MPA.0b013e31822a9b05. [DOI] [PubMed] [Google Scholar]
- 20.Hahm KB, Kim JH, You BM, Kim YS, Cho SW, Yim H, Ahn BO, Kim WB. Induction of apoptosis with an extract of Artemisia asiatica attenuates the severity of cerulein-induced pancreatitis in rats. Pancreas. 1998;17:153–157. doi: 10.1097/00006676-199808000-00007. [DOI] [PubMed] [Google Scholar]
- 21.Bhatia M. Apoptosis of pancreatic acinar cells in acute pancreatitis: is it good or bad? J Cell Mol Med. 2004;8:402–409. doi: 10.1111/j.1582-4934.2004.tb00330.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Bhatia M. Apoptosis versus necrosis in acute pancreatitis. Am J Physiol Gastrointest Liver Physiol. 2004;286:G189–G196. doi: 10.1152/ajpgi.00304.2003. [DOI] [PubMed] [Google Scholar]
- 23.Weng TI, Wu HY, Chen BL, Liu SH. Honokiol attenuates the severity of acute pancreatitis and associated lung injury via acceleration of acinar cell apoptosis. Shock. 2012;37:478–484. doi: 10.1097/SHK.0b013e31824653be. [DOI] [PubMed] [Google Scholar]
- 24.Yoo BM, Yeo M, Oh TY, Choi JH, Kim WW, Kim JH, Cho SW, Kim SJ, Hahm KB. Amelioration of pancreatic fibrosis in mice with defective TGF-beta signaling. Pancreas. 2005;30:e71–e79. doi: 10.1097/01.mpa.0000157388.54016.0a. [DOI] [PubMed] [Google Scholar]
- 25.Bhatia M, Wong FL, Fu D, Lau HY, Moochhala SM, Moore PK. Role of hydrogen sulfide in acute pancreatitis and associated lung injury. FASEB J. 2005;19:623–625. doi: 10.1096/fj.04-3023fje. [DOI] [PubMed] [Google Scholar]
- 26.Bhatia M. Hydrogen sulfide and substance P in inflammation. Antioxid Redox Signal. 2010;12:1191–1202. doi: 10.1089/ars.2009.2927. [DOI] [PubMed] [Google Scholar]
- 27.Bhatia M, Zhi L, Zhang H, Ng SW, Moore PK. Role of substance P in hydrogen sulfide-induced pulmonary inflammation in mice. Am J Physiol Lung Cell Mol Physiol. 2006;291:L896–L904. doi: 10.1152/ajplung.00053.2006. [DOI] [PubMed] [Google Scholar]
- 28.Bhatia M, Sidhapuriwala JN, Ng SW, Tamizhselvi R, Moochhala SM. Pro-inflammatory effects of hydrogen sulphide on substance P in caerulein-induced acute pancreatitis. J Cell Mol Med. 2008;12:580–590. doi: 10.1111/j.1582-4934.2007.00131.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Witt H, Beer S, Rosendahl J, Chen JM, Chandak GR, Masamune A, Bence M, Szmola R, Oracz G, Macek M, et al. Variants in CPA1 are strongly associated with early onset chronic pancreatitis. Nat Genet. 2013;45:1216–1220. doi: 10.1038/ng.2730. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Pinho AV, Rooman I, Reichert M, De Medts N, Bouwens L, Rustgi AK, Real FX. Adult pancreatic acinar cells dedifferentiate to an embryonic progenitor phenotype with concomitant activation of a senescence programme that is present in chronic pancreatitis. Gut. 2011;60:958–966. doi: 10.1136/gut.2010.225920. [DOI] [PubMed] [Google Scholar]
- 31.Wood NJ. Genetics: Global role for CPA1 variants in the pathogenesis of chronic pancreatitis. Nat Rev Gastroenterol Hepatol. 2013;10:567. doi: 10.1038/nrgastro.2013.172. [DOI] [PubMed] [Google Scholar]
- 32.Chen JM, Férec C. Chronic pancreatitis: genetics and pathogenesis. Annu Rev Genomics Hum Genet. 2009;10:63–87. doi: 10.1146/annurev-genom-082908-150009. [DOI] [PubMed] [Google Scholar]
- 33.Brock C, Nielsen LM, Lelic D, Drewes AM. Pathophysiology of chronic pancreatitis. World J Gastroenterol. 2013;19:7231–7240. doi: 10.3748/wjg.v19.i42.7231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Witt H, Apte MV, Keim V, Wilson JS. Chronic pancreatitis: challenges and advances in pathogenesis, genetics, diagnosis, and therapy. Gastroenterology. 2007;132:1557–1573. doi: 10.1053/j.gastro.2007.03.001. [DOI] [PubMed] [Google Scholar]
- 35.Braganza JM, Lee SH, McCloy RF, McMahon MJ. Chronic pancreatitis. Lancet. 2011;377:1184–1197. doi: 10.1016/S0140-6736(10)61852-1. [DOI] [PubMed] [Google Scholar]
- 36.Su KH, Cuthbertson C, Christophi C. Review of experimental animal models of acute pancreatitis. HPB (Oxford) 2006;8:264–286. doi: 10.1080/13651820500467358. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Foster JR. A review of animal models of nonneoplastic pancreatic diseases. Toxicol Pathol. 2014;42:243–259. doi: 10.1177/0192623313508479. [DOI] [PubMed] [Google Scholar]
- 38.Aghdassi AA, Mayerle J, Christochowitz S, Weiss FU, Sendler M, Lerch MM. Animal models for investigating chronic pancreatitis. Fibrogenesis Tissue Repair. 2011;4:26. doi: 10.1186/1755-1536-4-26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Willemer S, Elsässer HP, Adler G. Hormone-induced pancreatitis. Eur Surg Res. 1992;24 Suppl 1:29–39. doi: 10.1159/000129237. [DOI] [PubMed] [Google Scholar]
- 40.Dabrowski A, Konturek SJ, Konturek JW, Gabryelewicz A. Role of oxidative stress in the pathogenesis of caerulein-induced acute pancreatitis. Eur J Pharmacol. 1999;377:1–11. doi: 10.1016/s0014-2999(99)00421-5. [DOI] [PubMed] [Google Scholar]
- 41.Yamasaki M, Takeyama Y, Shinkai M, Ohyanagi H. Pancreatic and bile duct obstruction exacerbates rat caerulein-induced pancreatitis: a new experimental model of acute hemorrhagic pancreatitis. J Gastroenterol. 2006;41:352–360. doi: 10.1007/s00535-005-1767-9. [DOI] [PubMed] [Google Scholar]
- 42.Kim H. Cerulein pancreatitis: oxidative stress, inflammation, and apoptosis. Gut Liver. 2008;2:74–80. doi: 10.5009/gnl.2008.2.2.74. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Paran H, Mayo A, Kidron D, Sivak G, Reshef T, Vider T, Ziv O, Freund U. Experimental acute necrotising pancreatitis: evaluation and characterisation of a model of intraparenchymal injection of sodium taurocholate in rats. Eur J Surg. 2000;166:894–898. doi: 10.1080/110241500447308. [DOI] [PubMed] [Google Scholar]
- 44.Kudari A, Wig JD, Vaiphei K, Kochhar R, Majumdar S, Gupta R, Yadav TD, Doley RP. Histopathological sequential changes in sodium taurocholate-induced acute pancreatitis. JOP. 2007;8:564–572. [PubMed] [Google Scholar]
- 45.Lichtenstein A, Milani R, Fernezlian SM, Leme AS, Capelozzi VL, Martins MA. Acute lung injury in two experimental models of acute pancreatitis: infusion of saline or sodium taurocholate into the pancreatic duct. Crit Care Med. 2000;28:1497–1502. doi: 10.1097/00003246-200005000-00040. [DOI] [PubMed] [Google Scholar]
- 46.Wittel UA, Wiech T, Chakraborty S, Boss B, Lauch R, Batra SK, Hopt UT. Taurocholate-induced pancreatitis: a model of severe necrotizing pancreatitis in mice. Pancreas. 2008;36:e9–21. doi: 10.1097/MPA.0b013e3181575103. [DOI] [PubMed] [Google Scholar]
- 47.Hegyi P, Rakonczay-Jr Z, Sari R, Czako L, Farkas N, Gog C, Nemeth J, Lonovics J, Takacs T. Insulin is necessary for the hypertrophic effect of cholecystokinin-octapeptide following acute necrotizing experimental pancreatitis. World J Gastroenterol. 2004;10:2275–2277. doi: 10.3748/wjg.v10.i15.2275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Rakonczay Z, Hegyi P, Dósa S, Iványi B, Jármay K, Biczó G, Hracskó Z, Varga IS, Karg E, Kaszaki J, et al. A new severe acute necrotizing pancreatitis model induced by L-ornithine in rats. Crit Care Med. 2008;36:2117–2127. doi: 10.1097/CCM.0b013e31817d7f5c. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Miyauchi M, Suda K, Kuwayama C, Abe H, Kakinuma C. Role of fibrosis-related genes and pancreatic duct obstruction in rat pancreatitis models: implications for chronic pancreatitis. Histol Histopathol. 2007;22:1119–1127. doi: 10.14670/HH-22.1119. [DOI] [PubMed] [Google Scholar]
- 50.Lombardi B, Estes LW, Longnecker DS. Acute hemorrhagic pancreatitis (massive necrosis) with fat necrosis induced in mice by DL-ethionine fed with a choline-deficient diet. Am J Pathol. 1975;79:465–480. [PMC free article] [PubMed] [Google Scholar]
- 51.Niederau C, Lüthen R, Niederau MC, Grendell JH, Ferrell LD. Acute experimental hemorrhagic-necrotizing pancreatitis induced by feeding a choline-deficient, ethionine-supplemented diet. Methodology and standards. Eur Surg Res. 1992;24 Suppl 1:40–54. doi: 10.1159/000129238. [DOI] [PubMed] [Google Scholar]
- 52.Ida S, Ohmuraya M, Hirota M, Ozaki N, Hiramatsu S, Uehara H, Takamori H, Araki K, Baba H, Yamamura K. Chronic pancreatitis in mice by treatment with choline-deficient ethionine-supplemented diet. Exp Anim. 2010;59:421–429. doi: 10.1538/expanim.59.421. [DOI] [PubMed] [Google Scholar]
- 53.Lee JH, An CS, Yun BS, Kang KS, Lee YA, Won SM, Gwag BJ, Cho SI, Hahm KB. Prevention effects of ND-07, a novel drug candidate with a potent antioxidative action and anti-inflammatory action, in animal models of severe acute pancreatitis. Eur J Pharmacol. 2012;687:28–38. doi: 10.1016/j.ejphar.2012.04.048. [DOI] [PubMed] [Google Scholar]
- 54.Easler JJ, Mounzer R, Papachristou GI. Pharmacological therapy for acute pancreatitis: where are we now? where are we going? Minerva Gastroenterol Dietol. 2012;58:365–376. [PubMed] [Google Scholar]
- 55.Bang UC, Semb S, Nojgaard C, Bendtsen F. Pharmacological approach to acute pancreatitis. World J Gastroenterol. 2008;14:2968–2976. doi: 10.3748/wjg.14.2968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Pande H, Thuluvath P. Pharmacological prevention of post-endoscopic retrograde cholangiopancreatography pancreatitis. Drugs. 2003;63:1799–1812. doi: 10.2165/00003495-200363170-00003. [DOI] [PubMed] [Google Scholar]
- 57.Olesen AE, Brokjaer A, Fisher IW, Larsen IM. Pharmacological challenges in chronic pancreatitis. World J Gastroenterol. 2013;19:7302–7307. doi: 10.3748/wjg.v19.i42.7302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Carrizzo A, Forte M, Damato A, Trimarco V, Salzano F, Bartolo M, Maciag A, Puca AA, Vecchione C. Antioxidant effects of resveratrol in cardiovascular, cerebral and metabolic diseases. Food Chem Toxicol. 2013;61:215–226. doi: 10.1016/j.fct.2013.07.021. [DOI] [PubMed] [Google Scholar]
- 59.Markus MA, Morris BJ. Resveratrol in prevention and treatment of common clinical conditions of aging. Clin Interv Aging. 2008;3:331–339. [PMC free article] [PubMed] [Google Scholar]
- 60.Marques FZ, Markus MA, Morris BJ. Resveratrol: cellular actions of a potent natural chemical that confers a diversity of health benefits. Int J Biochem Cell Biol. 2009;41:2125–2128. doi: 10.1016/j.biocel.2009.06.003. [DOI] [PubMed] [Google Scholar]
- 61.Meng Y, Ma QY, Kou XP, Xu J. Effect of resveratrol on activation of nuclear factor kappa-B and inflammatory factors in rat model of acute pancreatitis. World J Gastroenterol. 2005;11:525–528. doi: 10.3748/wjg.v11.i4.525. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Ma Q, Zhang M, Wang Z, Ma Z, Sha H. The beneficial effect of resveratrol on severe acute pancreatitis. Ann N Y Acad Sci. 2011;1215:96–102. doi: 10.1111/j.1749-6632.2010.05847.x. [DOI] [PubMed] [Google Scholar]
- 63.Jha RK, Ma Q, Sha H, Palikhe M. Emerging role of resveratrol in the treatment of severe acute pancreatitis. Front Biosci (Schol Ed) 2010;2:168–175. doi: 10.2741/s54. [DOI] [PubMed] [Google Scholar]
- 64.Ma ZH, Ma QY, Wang LC, Sha HC, Wu SL, Zhang M. Effect of resveratrol on peritoneal macrophages in rats with severe acute pancreatitis. Inflamm Res. 2005;54:522–527. doi: 10.1007/s00011-005-1388-z. [DOI] [PubMed] [Google Scholar]
- 65.Yoo BM, Oh TY, Kim YB, Yeo M, Lee JS, Surh YJ, Ahn BO, Kim WH, Sohn S, Kim JH, et al. Novel antioxidant ameliorates the fibrosis and inflammation of cerulein-induced chronic pancreatitis in a mouse model. Pancreatology. 2005;5:165–176. doi: 10.1159/000085268. [DOI] [PubMed] [Google Scholar]
- 66.Feng Z, Fei J, Wenjian X, Jiachen J, Beina J, Zhonghua C, Xiangyi Y, Shaoying W. Rhubarb attenuates the severity of acute necrotizing pancreatitis by inhibiting MAPKs in rats. Immunotherapy. 2012;4:1817–1821. doi: 10.2217/imt.12.131. [DOI] [PubMed] [Google Scholar]
- 67.Bae GS, Kim MS, Park KC, Koo BS, Jo IJ, Choi SB, Lee DS, Kim YC, Kim TH, Seo SW, et al. Effect of biologically active fraction of Nardostachys jatamansi on cerulein-induced acute pancreatitis. World J Gastroenterol. 2012;18:3223–3234. doi: 10.3748/wjg.v18.i25.3223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Seo SW, Bae GS, Kim SG, Yun SW, Kim MS, Yun KJ, Park RK, Song HJ, Park SJ. Protective effects of Curcuma longa against cerulein-induced acute pancreatitis and pancreatitis-associated lung injury. Int J Mol Med. 2011;27:53–61. doi: 10.3892/ijmm.2010.548. [DOI] [PubMed] [Google Scholar]
- 69.Li K, Feng JY, Li YY, Yuece B, Lin XH, Yu LY, Li YN, Feng YJ, Storr M. Anti-inflammatory role of cannabidiol and O-1602 in cerulein-induced acute pancreatitis in mice. Pancreas. 2013;42:123–129. doi: 10.1097/MPA.0b013e318259f6f0. [DOI] [PubMed] [Google Scholar]
- 70.Jo IJ, Bae GS, Park KC, Choi SB, Jung WS, Jung SY, Cho JH, Choi MO, Song HJ, Park SJ. Scolopendra subspinipes mutilans protected the cerulein-induced acute pancreatitis by inhibiting high-mobility group box protein-1. World J Gastroenterol. 2013;19:1551–1562. doi: 10.3748/wjg.v19.i10.1551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Bae GS, Heo KH, Park KC, Choi SB, Jo IJ, Seo SH, Kim DG, Shin JY, Kang DG, Lee HS, et al. Apamin attenuated cerulein-induced acute pancreatitis by inhibition of JNK pathway in mice. Dig Dis Sci. 2013;58:2908–2917. doi: 10.1007/s10620-013-2800-0. [DOI] [PubMed] [Google Scholar]
- 72.Bae GS, Park KC, Choi SB, Jo IJ, Choi MO, Hong SH, Song K, Song HJ, Park SJ. Protective effects of alpha-pinene in mice with cerulein-induced acute pancreatitis. Life Sci. 2012;91:866–871. doi: 10.1016/j.lfs.2012.08.035. [DOI] [PubMed] [Google Scholar]
- 73.Wang J, Chen G, Gong H, Huang W, Long D, Tang W. Amelioration of experimental acute pancreatitis with Dachengqi Decoction via regulation of necrosis-apoptosis switch in the pancreatic acinar cell. PLoS One. 2012;7:e40160. doi: 10.1371/journal.pone.0040160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Zhao J, Chen J, Tang W, Wan L, Xiong W, Zhou L. Effect of Da-Cheng-Qi decoction on pancreatitis-associated lung injury in patients and anti-inflammatory responses in rat models. Pharm Biol. 2011;49:1058–1064. doi: 10.3109/13880209.2011.565059. [DOI] [PubMed] [Google Scholar]
- 75.Zhang XP, Wang C, Wu DJ, Ma ML, Ou JM. Protective effects of ligustrazine, kakonein and Panax notoginsenosides on multiple organs in rats with severe acute pancreatitis. Methods Find Exp Clin Pharmacol. 2010;32:631–644. doi: 10.1358/mf.2010.32.9.1444768. [DOI] [PubMed] [Google Scholar]
- 76.Zhang XP, Zhang L, Yang P, Zhang RP, Cheng QH. Protective effects of baicalin and octreotide on multiple organ injury in severe acute pancreatitis. Dig Dis Sci. 2008;53:581–591. doi: 10.1007/s10620-007-9868-3. [DOI] [PubMed] [Google Scholar]
- 77.Sidhu S, Pandhi P, Malhotra S, Vaiphei K, Khanduja KL. Beneficial effects of Emblica officinalis in L-arginine-induced acute pancreatitis in rats. J Med Food. 2011;14:147–155. doi: 10.1089/jmf.2010.1108. [DOI] [PubMed] [Google Scholar]
- 78.Sidhu S, Pandhi P, Malhotra S, Vaiphei K, Khanduja KL. Melatonin treatment is beneficial in pancreatic repair process after experimental acute pancreatitis. Eur J Pharmacol. 2010;628:282–289. doi: 10.1016/j.ejphar.2009.11.058. [DOI] [PubMed] [Google Scholar]
- 79.Abdin AA, El-Hamid MA, El-Seoud SH, Balaha MF. Effect of pentoxifylline and/or alpha lipoic acid on experimentally induced acute pancreatitis. Eur J Pharmacol. 2010;643:289–296. doi: 10.1016/j.ejphar.2010.06.020. [DOI] [PubMed] [Google Scholar]
- 80.Joo KR, Shin HP, Cha JM, Nam S, Huh Y. Effect of Korean red ginseng on superoxide dismutase inhibitor-induced pancreatitis in rats: a histopathologic and immunohistochemical study. Pancreas. 2009;38:661–666. doi: 10.1097/MPA.0b013e3181a9eb85. [DOI] [PubMed] [Google Scholar]
- 81.Zhang R, Zhang X, Zhang J, Wu J, Ye Q, Xu R, Ye J, Fang X, Jin L, He J, et al. Efficacy and mechanism of Salvia miltiorrhizae injection in the treatment of rats with severe acute pancreatitis. Inflammation. 2009;32:109–119. doi: 10.1007/s10753-009-9109-3. [DOI] [PubMed] [Google Scholar]
- 82.Hackert T, Werner J. Antioxidant therapy in acute pancreatitis: experimental and clinical evidence. Antioxid Redox Signal. 2011;15:2767–2777. doi: 10.1089/ars.2011.4076. [DOI] [PubMed] [Google Scholar]
- 83.Seo JY, Kim H, Seo JT, Kim KH. Oxidative stress induced cytokine production in isolated rat pancreatic acinar cells: effects of small-molecule antioxidants. Pharmacology. 2002;64:63–70. doi: 10.1159/000056152. [DOI] [PubMed] [Google Scholar]
- 84.Yu JH, Lim JW, Namkung W, Kim H, Kim KH. Suppression of cerulein-induced cytokine expression by antioxidants in pancreatic acinar cells. Lab Invest. 2002;82:1359–1368. doi: 10.1097/01.lab.0000032377.09626.c7. [DOI] [PubMed] [Google Scholar]
- 85.Zhang ZW, Zhang QY, Zhou MT, Liu NX, Chen TK, Zhu YF, Wu L. Antioxidant inhibits HMGB1 expression and reduces pancreas injury in rats with severe acute pancreatitis. Dig Dis Sci. 2010;55:2529–2536. doi: 10.1007/s10620-009-1073-0. [DOI] [PubMed] [Google Scholar]
- 86.Altavilla D, Famulari C, Passaniti M, Campo GM, Macrì A, Seminara P, Marini H, Calò M, Santamaria LB, Bono D, et al. Lipid peroxidation inhibition reduces NF-kappaB activation and attenuates cerulein-induced pancreatitis. Free Radic Res. 2003;37:425–435. doi: 10.1080/1071576031000070093. [DOI] [PubMed] [Google Scholar]
- 87.Jung KH, Hong SW, Zheng HM, Lee HS, Lee H, Lee DH, Lee SY, Hong SS. Melatonin ameliorates cerulein-induced pancreatitis by the modulation of nuclear erythroid 2-related factor 2 and nuclear factor-kappaB in rats. J Pineal Res. 2010;48:239–250. doi: 10.1111/j.1600-079X.2010.00748.x. [DOI] [PubMed] [Google Scholar]
- 88.Choi KS, Song H, Kim EH, Choi JH, Hong H, Han YM, Hahm KB. Inhibition of Hydrogen Sulfide-induced Angiogenesis and Inflammation in Vascular Endothelial Cells: Potential Mechanisms of Gastric Cancer Prevention by Korean Red Ginseng. J Ginseng Res. 2012;36:135–145. doi: 10.5142/jgr.2012.36.2.135. [DOI] [PMC free article] [PubMed] [Google Scholar]