

Abstract
The risk for Staphylococcus aureus bloodstream infection (BSI) is increased in immunocompromised individuals, including patients with hematologic malignancy and/or chemotherapy. Due to the emergence of antibiotic-resistant strains, designated methicillin-resistant S. aureus (MRSA), staphylococcal BSI in cancer patients is associated with high mortality; however, neither a protective vaccine nor pathogen-specific immunotherapy is currently available. Here, we modeled staphylococcal BSI in leukopenic CD-1 mice that had been treated with cyclophosphamide, a drug for leukemia and lymphoma patients. Cyclophosphamide-treated mice were highly sensitive to S. aureus BSI and developed infectious lesions lacking immune cell infiltrates. Virulence factors of S. aureus that are key for disease establishment in immunocompetent hosts—α-hemolysin (Hla), iron-regulated surface determinants (IsdA and IsdB), coagulase (Coa), and von Willebrand factor binding protein (vWbp)—are dispensable for the pathogenesis of BSI in leukopenic mice. In contrast, sortase A mutants, which cannot assemble surface proteins, display delayed time to death and increased survival in this model. A vaccine with four surface antigens (ClfA, FnBPB, SdrD, and SpAKKAA), which was identified by genetic vaccinology using sortase A mutants, raised antigen-specific immune responses that protected leukopenic mice against staphylococcal BSI.
INTRODUCTION
Staphylococcus aureus is a commensal of the human nares, skin, and gastrointestinal tract that also causes invasive disease, including skin and soft tissue infections (SSTI), bacteremia, sepsis, endocarditis, pneumonia, and osteomyelitis (1, 2). S. aureus causes disease in healthy individuals, which most frequently manifests as purulent SSTIs (3). Invasive disease is associated with bloodstream infection (BSI) that may develop into fulminant sepsis or endocarditis (4, 5). The treatment of S. aureus infections has been complicated by the emergence and spread of antibiotic-resistant strains, designated methicillin-resistant S. aureus (MRSA), that have evolved resistance traits against many therapeutics (6, 7).
Patients with indwelling catheters, endotracheal intubation, medical implantation of foreign bodies (prosthetic joints, implants, and heart valves), trauma, surgical procedures, hemodialysis, peritoneal dialysis, immunosuppressive or cancer therapy, and diabetes, as well as individuals with increased age and low birth weight, are all at elevated risk of S. aureus infection (8, 9). These patient populations can be broadly classified into three groups: individuals with a breach of barrier function, enabling staphylococci to cause invasive infections (trauma, surgery); individuals with implants that serve as a protective niche for staphylococci; and individuals with diminished innate immune defenses, most importantly patients with reduced opsonophagocytic killing of bacteria, which is mediated by neutrophil granulocytes (10, 11). The latter can occur in individuals with primary immunodeficiency disorders, caused in patients with chronic granulomatous disease, for example, by mutations that affect the NADPH oxidase of neutrophil granulocytes to produce oxygen radicals for killing of staphylococci (12, 13). Much more frequently, however, diminished opsonophagocytic killing of staphylococci occurs in individuals with hematologic malignancies and/or anticancer chemotherapy (14), a patient population where S. aureus BSIs cause significant morbidity and mortality (15, 16).
Cyclophosphamide is an alkylating agent that is used for the therapy of malignancies and autoimmune diseases and for bone marrow transplantation (17). Cyclophosphamide therapy induces neutropenia and leukopenia and is associated with increased risk for S. aureus BSI (18, 19). Although cyclophosphamide-induced leukopenia in mice has been used extensively as a preclinical model to assess therapeutic efficacy of antibiotics against bacterial infections, this model has heretofore not been used to analyze the virulence factors for S. aureus BSI in patients with diminished capacity for opsonophagocytic killing (20, 21).
Although there is a need for vaccines and immunotherapies that protect high-risk patients against S. aureus infection, several clinical trials for staphylococcal vaccines failed to meet their study endpoints (22). These trials have focused on single staphylococcal antigens that, during preclinical testing, were demonstrated to act as virulence factors for the pathogenesis of specific disease and, when used as subunit vaccines, elicited immune responses that protect immunocompetent animals (23–27). Clinical trials for S. aureus vaccines included capsular polysaccharides (CP types 5 and 8), α-hemolysin (Hla), coagulase (Coa), and the iron-regulated surface determinant B (IsdB) (28–30).
Here, we use the leukopenic mouse model to analyze S. aureus variants lacking specific protective antigens to characterize targets for a staphylococcal vaccine in cancer patients. Using genetic vaccinology to derive vaccine targets, we report that a cocktail of four surface protein antigens (ClfA, FnBPB, SdrD, and SpAKKAA) provides protection against S. aureus BSI in leukopenic mice.
MATERIALS AND METHODS
Animal care and regulatory compliance.
All experiments involving the care and use of animals followed protocols that were reviewed, approved, and performed under the regulatory supervision of The University of Chicago's Institutional Biosafety Committee and the Institutional Animal Care and Use Committee. Animal care was managed by The University of Chicago Animal Resource Center, accredited by the American Association for Accreditation of Laboratory Animal Care and the Department of Health and Human Services (A3523-01). Animals were maintained in accordance with the applicable portions of the Animal Welfare Act and the Guide for the Care and Use of Laboratory Animals (31). Veterinary care was under the direction of full-time resident veterinarians boarded by the American College of Laboratory Animal Medicine. CD-1 mice (strain code 022) were purchased from Charles River.
Statistical analysis.
Mouse survival was analyzed for significance using the two-tailed log rank test. The bacterial load following S. aureus infection, represented as the log10 CFU/g of organ tissue, was analyzed with the Mann-Whitney test for statistical significance. Quantification of abscess formation was analyzed for statistical significance using the unpaired two-tailed Student t test. Statistical analyses were performed using GraphPad Prism 4 software. All animal experiments were examined for reproducibility using either two or three independent determinations.
Bacterial strains, media, and growth conditions.
Mutants harboring the bursa aurealis mariner transposon in defined genes were transduced with bacteriophage ϕ85 into S. aureus Newman (32). The isdA isdB and coa vwb double mutants have been described (33, 34). S. aureus cultures were grown at 37°C in tryptic soy broth (TSB), and bursa aurealis mutant cultures were supplemented with 10 μg/ml of erythromycin. Escherichia coli strain BL21(DE3) was grown in Luria-Bertani (LB) broth containing 100 μg/ml of ampicillin at 37°C when plasmid was present.
Protein purification.
Recombinant clumping factor A (ClfA), fibrinogen binding protein A (FnBPA), FnBPB, serine-aspartate repeat C (SdrC), SdrD, SdrE, nontoxigenic staphylococcal protein A (SpAKKAA), and staphylococcal immunoglobulin binding protein (SbiKKAA) carrying hexa-histidine tags were produced from E. coli BL21(DE3) carrying plasmids described earlier (35–37). Briefly, overnight cultures of recombinant E. coli strains were diluted 1:100 into fresh medium and grown at 37°C to an absorbance at 600 nm (A600) of 0.5, at which point cultures were induced with 1 mM isopropyl β-d-1-thiogalatopyranoside (IPTG) and grown for an additional 3 h. Bacterial cells were sedimented by centrifugation (3,000 × g for 5 min), washed and suspended in buffer A (50 mM Tris-HCl, pH 7.5, containing 150 mM NaCl and 10 mM imidazole), and disrupted with a French pressure cell at 14,000 lb/in2. Crude lysates were centrifuged (40,000 × g for 40 min), and filtered supernatants were subjected to affinity purification. His-tagged proteins were purified over a nickel-nitrilotriacetic acid (Ni-NTA) agarose affinity column by gravity flow and eluted with an imidazole gradient (10 to 500 mM). Protein purification was ascertained by separating sample aliquots on 12% SDS-PAGE gels and Coomassie staining. Fractions containing the proteins were pooled and subjected to 1% Triton X-114 extraction to remove endotoxin. Following detergent addition, samples were first kept on ice for 10 min and then incubated at 37°C for 10 min followed by centrifugation (16,000 × g for 10 min). Triton X-114 extraction was repeated twice, and the aqueous phase containing the proteins was subjected to gel filtration in endotoxin-free phosphate-buffered saline (PBS; Cellgro) using a Hi-Trap desalting column (GE Healthcare). Protein concentration was assessed with the bicinchoninic acid assay (BCA).
Cyclophosphamide-induced leukopenia and Staphylococcus aureus challenge of animals.
Six-week-old female CD-1 mice were treated every 48 h via intraperitoneal injection with cyclophosphamide monohydrate (CPM; Sigma product no. C0768) dissolved in 200 μl of sterile water at a dose of 150 mg/kg body weight or were mock (water) treated for 7 days prior to the infection and throughout the experiment. To confirm leukopenia, mouse blood from at least three mice per group was drawn via cardiac puncture into collection tubes lined with EDTA (Sarstedt; Microvette, product no. 20.1278.100), and white blood cells were counted with a hematology analyzer (Hemavet 950; Drew Scientific). For infection of animals, overnight cultures of S. aureus were inoculated 1:100 into fresh TSB and grown for 2 h with shaking at 37°C. Staphylococci were sedimented by centrifugation, washed, suspended, and diluted in sterile, endotoxin-free PBS to obtain a starting stock of 1 × 109 CFU/ml. Inocula were determined by CFU enumeration following serial dilution and plating on tryptic soy agar, followed by incubation of plates for 16 h at 37°C. Seven-week-old cyclophosphamide or mock-treated CD-1 mice were anesthetized by intraperitoneal injection with a cocktail of 65 mg ketamine and 2 mg xylazine per kilogram of body weight and infected with 100 μl of bacterial suspension (ranging from 105 to 108 CFU) via intravenous injection into the periorbital venous plexus. Infected animals were monitored for morbidity or recovery over a period of 10 days. Mice were killed by CO2 inhalation, and organs were excised and homogenized in sterile PBS and 0.1% Triton X-100 by using a high-shear lab homogenizer (Omni International). Homogenates were diluted, plated on agar, and incubated for 16 h at 37°C for enumeration of CFU.
Histopathology.
Mouse organs were excised during necropsy and fixed in 10% formalin for 24 h at room temperature. Tissues were embedded in paraffin, thin-sectioned, stained with hematoxylin and eosin (H&E), and examined by light microscopy.
Immunization studies.
For active immunization, 3-week-old CD-1 mice were injected intramuscularly with a mix of 25 μg of each recombinant protein (SpAKKAA, SdrD, FnBPpB, and ClfA) emulsified in complete Freund adjuvant (Difco), 100-μl volumes of each dose, followed by a boost on day 12 with 25 μg of each protein emulsified in incomplete Freund adjuvant (Difco). Cyclophosphamide and mock treatment were started on day 21 and continued throughout the experiment every 48 h. Blood was collected on day 26 via periorbital vein puncture with heparinized microhematocrit capillary tubes (Fisher). Z-gel serum separation microtubes (Sarstedt) were used to collect serum, and specific antibody production was assessed via enzyme-linked immunosorbent assay (ELISA) as described previously (36). On day 27, mice were challenged by intravenous injection with 2 × 106 CFU of S. aureus Newman. Antibody titers in mouse sera were analyzed via ELISA for the vaccine antigens SpAKKAA, SdrD, FnBPB, and ClfA as well as for cross-reactivity toward SbiKKAA, FnBPA, ClfB, SdrC, and SdrE.
RESULTS
Leukopenia following cyclophosphamide treatment of CD-1 mice.
Earlier work used outbred ICR mice and intraperitoneal injection of two doses (150 and 100 mg/kg body weight) of cyclophosphamide, administered 48 h apart (days 1 and 3), to induce leukopenia (<10 neutrophils/mm3 blood, 84% reduction in white blood cells, with 92% and 96% decline in lymphocytes and monocytes, respectively) for 3 days (21). Others used Swiss mice and two injections of 150 mg/kg cyclophosphamide on days 1 and 4 to achieve neutropenia (38). Using female outbred CD-1 mice, we observed that three intraperitoneal injections of 150 mg/kg cyclophosphamide in 48-h intervals (days 1, 3, and 5) were required to cause leukopenia (Fig. 1A and B). At the day 7 interval, neutrophils were >90% depleted relative to the average cell count prior to cyclophosphamide treatment; other leukocyte populations were >80% depleted (Fig. 1A and B). We also observed that leukopenia was not sustained for the 10-day observation period of our experimental plan unless CD-1 mice received continued treatment with cyclophosphamide in 48-h intervals. Cyclophosphamide treatment did not affect the weight of animals compared to the weight of mock-treated animals (data not shown). On the basis of these observations, we developed a protocol whereby CD-1 mice were either mock or cyclophosphamide (150 mg/kg) treated on days 1, 3, and 5 and challenged by intravenous inoculation with S. aureus on day 6. Animals received mock or cyclophosphamide treatment in 48-h intervals (days 7, 9, 11, 13, 15) unless mice succumbed to the infectious challenge.
FIG 1.
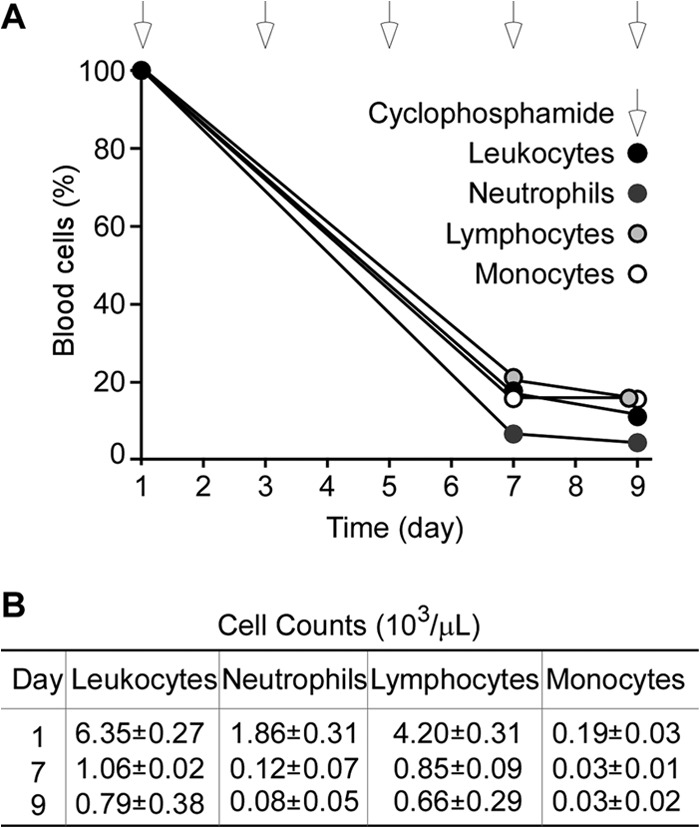
Induction of leukopenia in CD-1 mice by cyclophosphamide treatment. Six-week-old CD-1 mice were treated every 48 h with 150 mg/kg cyclophosphamide via intraperitoneal injection. On days 1, 7, and 9, mice were euthanized, and blood was sampled via cardiac puncture and anticoagulated through the addition of EDTA. (A) The numbers of white blood cells (WBC) in each sample were enumerated by hemocytometry and plotted as a function of time. White arrows above the plot indicate days when animals received cyclophosphamide treatment. (B) The average cell counts (and the standard errors of the means) of leukocytes, neutrophils, lymphocytes, and monocytes were determined on days 1, 7, and 9. On day 7, cyclophosphamide treatment had depleted 83% of leukocytes, 94% or neutrophils, 80% of lymphocytes, and 84% of monocytes compared to those in mock-treated mice.
Staphylococcus aureus bloodstream infection in leukopenic mice.
Cohorts of CD-1 mice (n = 10) were infected via intravenous injection with S. aureus Newman (32), a human clinical isolate that has been used for animal model development (27, 39). Injection of 1 × 108 CFU of S. aureus Newman caused 50% mortality in immunocompetent CD-1 mice over the 10-day observation period, whereas lower inocula (5 × 107 or 1 × 107 CFU) resulted in reduced mortality (Fig. 2A). In contrast, all leukopenic mice that had been treated with cyclophosphamide succumbed to BSI when challenged with 1 × 106 or 1 × 107 CFU S. aureus Newman. Mice that received a higher dose (1 × 107) succumbed with a shortened mean time to death (<24 h) compared to that of animals challenged with 1 × 106 CFU (5 days) (Fig. 2B). Even a challenge dose of 1 × 105 CFU S. aureus Newman caused bloodstream infections with 80% mortality over the 10-day observation period (Fig. 2B).
FIG 2.
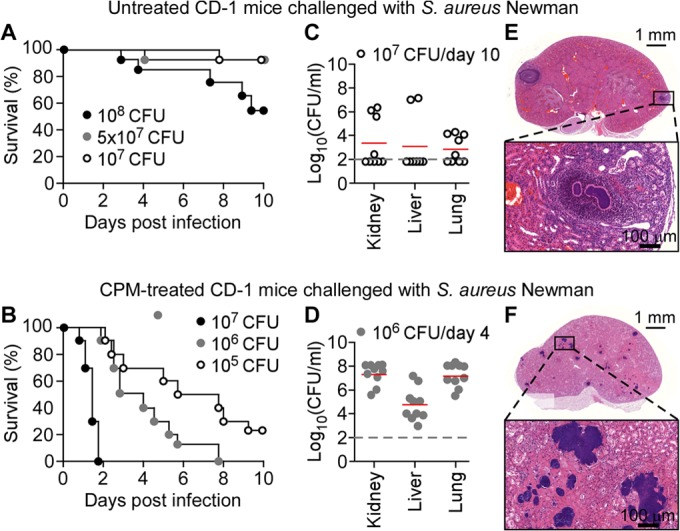
Staphylococcus aureus bloodstream infection in leukopenic mice. (A and B) Cohorts of 7-week-old female CD-1 mice (n = 10) mock treated (A) or treated with cyclophosphamide in 48-h intervals (B) were infected by injection of S. aureus Newman (1 × 105, 1 × 106, 1 × 107, 5 × 107, or 1 × 108 CFU) into the periorbital venous plexus, and animal survival was recorded over 10 days. (C and D) At day 10 (C) or 4 (D) postinfection, kidney, liver, and lung tissues of mock-treated (C) or cyclophosphamide-treated (D) CD-1 mice infected with S. aureus Newman bloodstream infection (1 × 106 or 1 × 107 CFU) were removed during necropsy; homogenized tissues were analyzed for staphylococcal load by plating serially diluted samples on agar plates and enumerating CFU. (E and F) Hematoxylin and eosin-stained thin sections of kidney tissues from mock-treated (E) and cyclophosphamide-treated (F) mice that had been euthanized on day 10 following bloodstream infection with 1 × 107 and 1 × 106 CFU S. aureus Newman, respectively, were viewed by light microscopy and images were acquired. Bars indicate length measurements. See the text for details.
Lethal outcome of S. aureus BSI was associated with staphylococcal replication in infected tissues (Fig. 2C to F). Enumeration of S. aureus in organs of mock-treated animals that survived a bloodstream challenge of 1 × 107 CFU revealed small bacterial numbers in kidney (an average of 1.99 × 103 CFU g−1 tissue homogenate), liver (1.26 × 103 CFU g−1), and lung (0.79 × 103 CFU g−1) tissues (Fig. 2C). In contrast, cyclophosphamide-treated animals harbored increased S. aureus loads in infected kidney (6.30 × 107 CFU g−1), liver (1.25 × 105 CFU g−1), and lung (7.94 × 106 CFU g−1) tissues at day 4 postinfection (Fig. 2C and D).
Following infection of immunocompetent mice, S. aureus BSI triggers the development of abscess lesions in many organ systems, which can be visualized in H&E-stained thin-sectioned kidney tissues (39). Renal tissues of mock-treated mice that had been euthanized 10 days following BSI with S. aureus Newman harbored 1 to 4 lesions that presented with characteristic morphology (39): a bacterial nidus (designated the staphylococcal abscess community), surrounded by an eosinophilic pseudocapsule composed of fibrin deposits and large infiltrates of immune cells (34) (Fig. 2E). H&E-stained renal tissues of cyclophosphamide-treated mice harbored >100 small lesions per kidney that were comprised of large numbers of staphylococcal cells and surrounded by eosinophilic deposits, likely comprised of fibrin; however, these lesions did not show immune cell infiltrates (Fig. 2D).
Impact of staphylococcal virulence factors on bloodstream infection in leukopenic mice.
Earlier work analyzed S. aureus Newman mutants with mutational lesions in genes encoding secreted products for defects in the pathogenesis of BSI (39, 40). Other experiments examined purified subunit vaccines, derived from secreted products of S. aureus, for their ability to protect mice against BSI (25, 34, 35). This work established that secreted products required for the pathogenesis of S. aureus BSI can also be used as subunit vaccines and provide disease protection via the formation of antibodies that neutralize staphylococcal virulence determinants (36, 41, 42).
We wondered whether previously characterized S. aureus virulence factors/vaccine determinants might be required for the pathogenesis of bloodstream infections in leukopenic mice and examined three variants with knockout mutations in the hla, coa vwb, and isdA isdB genes. Staphylococcal alpha-toxin (α-hemolysin) is encoded by the hla gene, and the primary translation product functions as a precursor that is cleaved by signal peptidase and secreted into the extracellular medium (43). Association of the soluble, monomeric form of Hla with its ADAM10 host cell receptor triggers oligomerization and membrane pore formation (44), causing injury to vascular endothelial cells, epithelial cells, as well as platelets and cells of the myeloid lineage (45, 46). During bloodstream infection of immunocompetent mice, the hla mutant of S. aureus Newman displayed reduced mortality and increased time to death (47). In contrast, when tested in leukopenic CD-1 mice, no differences in overall mortality or time to death were detected between animals that had been challenged with wild-type or hla mutant S. aureus Newman (Fig. 3A; P = 0.8649). Becker et al. showed that the contributions of S. aureus hla toward lethal pneumonia in mice require the presence of myeloid cells expressing the Hla receptor molecule, ADAM10 (46). In agreement with these results, ablation of white blood cells via cyclophosphamide treatment also eliminates the pathogen's need for Hla production to establish lethal BSI in mice.
FIG 3.
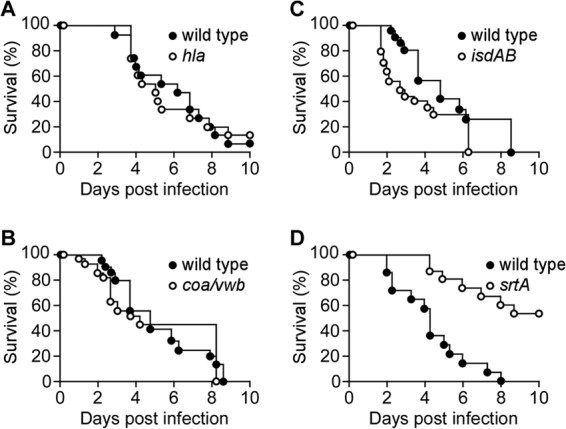
Virulence factor contribution to Staphylococcus aureus bloodstream infection in leukopenic mice. Cohorts of 7-week-old female leukopenic (cyclophosphamide-treated) CD-1 mice (n = 10 to 15) were infected by intravenous injection with 2 × 106 CFU of S. aureus Newman wild type or its isogenic variants with deletions or insertions in hla (A), coa vwb (B), isdAB (C), or srtA (D). Animal survival was recorded over 10 days. Statistical significance in animal survival between two cohorts was analyzed with the two-tailed log rank test; P ≤ 0.05, significant difference.
Coagulase (Coa) and von Willebrand factor binding protein (vWbp) are also secreted into the extracellular medium by S. aureus (48, 49). Both proteins associate with and nonproteolytically activate prothrombin to convert fibrinogen into fibrin clots (50), which promotes escape from phagocytic killing and establishment of abscess lesions in infected tissues via the formation of a fibrin capsule that restricts access of immune cells to staphylococcal abscess communities (34). During bloodstream infection of immunocompetent mice, the coa vwb mutant displayed delayed time to death and increased survival compared to those of wild-type S. aureus Newman (40). In contrast, during bloodstream infection of leukopenic mice, the coa vwb mutant displayed mortality and time to death similar to those of wild-type S. aureus Newman (Fig. 3B; P = 0.5149).
Two surface proteins of S. aureus, IsdA and IsdB, contribute to heme-iron scavenging from host hemoglobin and to passage of the tetrapyrrole moiety across the bacterial envelope (33, 51). S. aureus isdAB mutants display a small delay in time to death during bloodstream infections as well as reduced replication in infected tissues (41, 52). During bloodstream infection in leukopenic mice, the S. aureus isdAB mutant caused mortality similar to that of the wild-type strain (Fig. 3C; P = 0.1356).
Staphylococci require sortase during bloodstream infection in leukopenic mice.
Sortase A is a transpeptidase that cleaves the LPXTG sorting signals of surface protein precursors and links their C-terminal carboxyl group to the peptidoglycan of S. aureus (53, 54). Sortase A mutants (srtA) cannot anchor any one of 18 surface proteins of S. aureus Newman to the bacterial envelope (55) and are unable to form abscess lesions or cause lethal BSI in immunocompetent mice (39, 40). When analyzed during bloodstream infection of leukopenic mice, the srtA mutant displayed a delayed time to death and increased survival compared to those of wild-type S. aureus Newman, indicating that the combined contributions of all surface proteins are required for staphylococcal disease processes in mice that lack most of their immune cells (Fig. 3D; P = 0.0039).
Contribution of surface proteins toward Staphylococcus aureus bloodstream infection in leukopenic mice.
Earlier work had used genetic vaccinology to compare the immune response of immunocompetent mice infected with wild-type S. aureus and sortase A mutants to identify antigens that elicit protective immune responses, as infection with the srtA variant, but not wild-type staphylococci, triggers protective immunity to recurrent infection (37). Using this technology, vaccines (COMBO 1 and COMBO 2) were derived by combining multiple antigens; COMBO 2 with four antigens (ClfA, FnBPB, SdrD, and SpAKKAA) elicited immune responses in immunocompetent mice and protected animals against challenge with wild-type S. aureus (37). Protein A (SpA) binds immunoglobulins via their Fcγ domain and the Fab heavy chains of VH3 clan antibodies, attributes that interfere with opsonophagocytosis of bacteria and with the development of adaptive B cell responses (56, 57). The SpAKKAA variant has lost these functions and, unlike wild-type SpA, elicits neutralizing antibody responses that promote opsonophagocytic killing of staphylococci and development of B cells to establish adaptive immune responses (36). ClfA binds to the C-terminal end of the fibrinogen γ-chain (58, 59), which is also available for staphylococcal binding during fibrin fiber assembly (40). Neutralizing antibodies can block ClfA binding to both fibrinogen and fibrin (40, 60). FnBPB binds to fibronectin, elastin, and fibrinogen, and these activities promote the pathogenesis of S. aureus BSI (61–65). The genome of S. aureus Newman carries a missense near the 3′ end of the genetic determinant for FnBPB, fnbB (66). Nevertheless, secreted fibronectin binding proteins contribute to the pathogenesis of S. aureus Newman infections (39, 67), as sortase-anchored products function not only on the bacterial surface but also when released into the extracellular medium (68). The molecular mechanisms whereby SdrD contributes to S. aureus pathogenesis are currently not known (39). Here, we asked whether any one of these four surface protein genes (clfA, fnbB, sdrD, or spa) contributed to the pathogenesis of S. aureus BSI in leukopenic mice, studying in pairwise comparison the survival or time to death of wild-type and mutant strains. Of note, none of the four surface protein genes (clfA, fnbB, sdrD, or spa) was necessary for S. aureus BSI-associated mortality or was a determinant of time to death in leukopenic mice (Fig. 4; wild type versus clfA, P = 0.1479; wild type versus fnbB, P = 0.1726; wild type versus sdrD, P = 0.0736; wild type versus spa, P = 0.2330).
FIG 4.
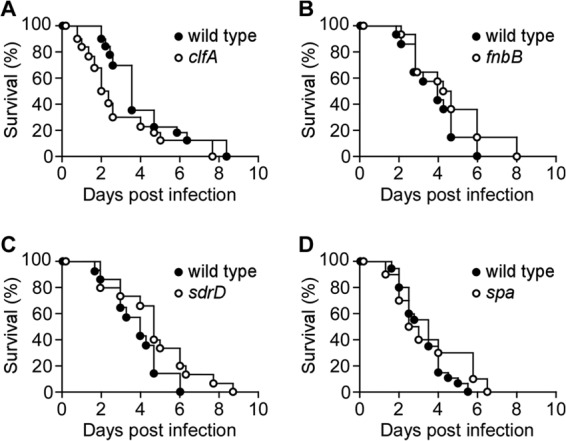
Contributions of individual surface proteins to Staphylococcus aureus bloodstream infection in leukopenic mice. Cohorts of 7-week-old female leukopenic (cyclophosphamide-treated) CD-1 mice (n = 10 to 20) were infected by intravenous injection with 2 × 106 CFU of S. aureus Newman wild type or its isogenic variants with deletions or insertions in clfA (A), fnbB (B), sdrD (C), or spa (D). Animal survival was recorded over 10 days. Statistical significance in animal survival between two cohorts was analyzed with the two-tailed log rank test; P ≤ 0.05, significant difference.
Surface protein vaccine protects leukopenic mice against S. aureus bloodstream infection.
To address the question of whether the surface protein vaccine derived via genetic vaccinology may protect leukopenic mice against S. aureus BSI, we first immunized immunocompetent CD-1 mice with a cocktail of purified recombinant proteins emulsified in adjuvant, including ClfA, FnBPB, SdrD, and SpAKKAA (COMBO 2). Animals received a booster immunization on day 12 and were bled to determine the concentration of serum IgG against each antigen. Compared to mock (adjuvant alone)-immunized mice, animals that had received the surface protein vaccine (COMBO 2) developed specific antibodies against SpAKKAA, FnBPB, ClfA, SdrC, SdrD, and SdrE but not against FnBPA, ClfB, and SbiKKAA (Fig. 5A). These findings can be explained by the high degree of sequence identity (76%) between the A domain of the serine-aspartate repeat protein family (SdrCDE), which is lower between the A domains of FnBPA and FnBPB (60%) as well as ClfA and ClfB (50%). The immunoglobulin binding domains (IgBDs) of SpA display sequence homology to the N-terminal two IgBDs of Sbi (staphylococcal binder of immunoglobulin; 35% identity), which was also not sufficient to elicit a strong cross-reactive immune response. As expected, mock-immunized animals did not generate antibodies against staphylococcal surface proteins (Fig. 5A).
FIG 5.
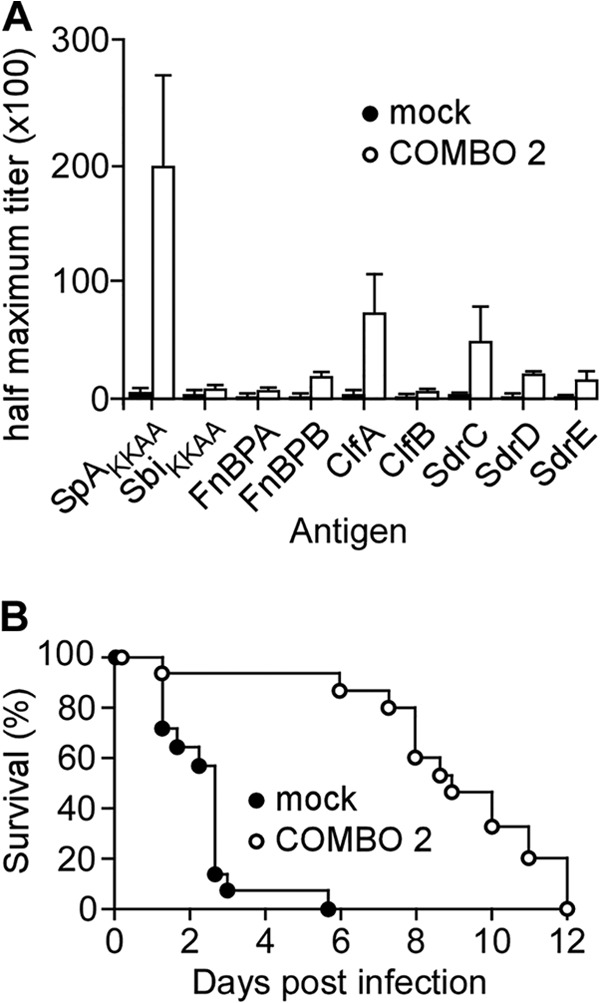
Surface protein vaccine prolongs the time to death of leukopenic mice with Staphylococcus aureus bloodstream infection. Three-week-old CD-1 mice were immunized with adjuvant alone (mock) or with a cocktail of 25 μg each of four recombinant purified proteins emulsified in adjuvant: ClfA, FnBPB, SdrD, and SpAKKAA. Mock or booster immunizations occurred on day 12. Animals were treated with cyclophosphamide on day 21 and in 48-h intervals thereafter for the duration of the experiment. Animals (n = 5) were bled on day 26, and mouse serum half-maximal antibody titers against purified S. aureus surface proteins (SpAKKAA, SbiKKAA, FnBPA, FnBPB, ClfA, ClfB, SdrC, SdrD, and SdrE) were determined by ELISA. Brackets denote standard errors of the means. (B) Leukopenic (cyclophosphamide-treated) CD-1 mice (n = 10) that had been mock immunized or immunized with the surface protein vaccine (vaccinated) were challenged by intravenous injection with 2 × 106 CFU S. aureus Newman on day 27 following the first immunization. Animal survival was recorded over 10 days. Statistical significance in animal survival between two cohorts was analyzed with the two-tailed log rank test; P ≤ 0.05, significant difference.
Vaccinated mice were treated with cyclophosphamide starting on day 21, and once animals were found to be leukopenic, they were challenged on day 27 with 2 × 106 CFU S. aureus Newman. Mock-immunized animals succumbed to challenge within 1 to 5 days, with a mean time to death of 2.5 days. Leukopenic animals that had received the surface protein vaccine succumbed more slowly to BSI, with a mean time to death of 8.5 days (mock versus vaccine, P < 0.0001) (Fig. 5B). To monitor the ability of the surface protein vaccine to reduce staphylococcal replication, cohorts of 5 animals (mock or vaccinated) were euthanized 3 h, 6 h, 1 day, and 3 days postchallenge, and the bacterial load in kidney, liver, and lung tissues was quantified via CFU enumeration. This experiment revealed that staphylococci disseminated within 3 h into organ tissues (Fig. 6). While the load of S. aureus in liver and lung tissues increased slowly in mock-immunized animals, mice that had received the surface protein vaccine did not increase their bacterial load in both organ tissues (Fig. 6D; P = 0.0119 in liver and P = 0.0195 in lung on day 3 postinfection). S. aureus replicated very quickly in renal tissues, and by day 3, an average bacterial load of 2.51 × 107 CFU g−1 was enumerated in mock-immunized animals (Fig. 6D). Leukopenic animals that had received the surface protein vaccine harbored a reduced bacterial load (3.98 × 104 CFU g−1) (Fig. 6D; mock versus COMBO 2, P = 0.0556). Thus, immunization with the surface protein vaccine not only extended the time to death of leukopenic animals with S. aureus BSI but also limited staphylococcal replication in infected tissues.
FIG 6.
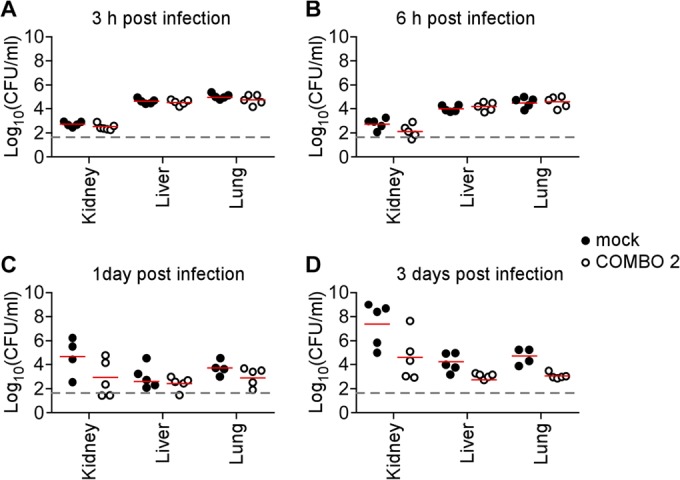
Surface protein vaccine inhibits bacterial replication in leukopenic mice with Staphylococcus aureus bloodstream infection. Mock-immunized (closed circles) or vaccinated (ClfA, FnBPB, SdrD, and SpAKKAA) (open circles) leukopenic (cyclophosphamide-treated) CD-1 mice were infected with 2 × 106 CFU of S. aureus Newman as described in the legend to Fig. 5. Animals were euthanized 3 h (A), 6 h (B), 1 day (C), or 3 days (D) postchallenge. Kidney, lung, and liver tissues were removed during necropsy and analyzed for staphylococcal load by plating serially diluted samples on agar plates and enumerating CFU. Statistical significance of differences in bacterial loads between mock-treated and vaccinated leukopenic mice was analyzed with the Mann-Whitney test; P ≤ 0.05, significant difference.
DISCUSSION
Chemotherapy of tumors, premature birth, human immunodeficiency virus (HIV) infection, leukemia, chronic granulomatous disease (CGD), and other inheritable diseases cause functional depletion or annihilation of phagocytic cells (69). Unless diagnosed early and treated with antimicrobial prophylaxis, such immunodeficiency is associated with significant morbidity and mortality because of bacterial infection (69, 70). S. aureus is a frequent cause of morbidity and mortality in individuals with immunodeficiency, owing to its status as a commensal of humans and to the dissemination of MRSA strains (4, 71). For example, individuals infected with HIV experience a 6-fold-higher risk of S. aureus SSTI than healthy individuals (72).
Cyclophosphamide treatment of mice has been used extensively to induce leukopenia and examine the preclinical efficacy of antibiotics in immunocompromised hosts (20, 21, 73). Cyclophosphamide-induced leukopenia in mice has also been used for the study of ESKAPE pathogens (Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter species), antibiotic-resistant bacteria most frequently associated with nosocomial infection and therapy failure (74). While most of the work in leukopenic mice focused thus far on studying the efficacy of antibiotics (75, 76), progress was also documented for the development of vaccine and immunotherapeutics to prevent P. aeruginosa BSI (77–79). Nevertheless, the development of vaccines that prevent S. aureus BSI in leukopenic mice was heretofore not investigated. Such study may be useful in designing vaccines for clinical use in combating S. aureus BSI in individuals that can be immunized prior to the onset of immunodeficiency, i.e., the chemotherapy of malignant tumors. Alternatively, immunocompromised individuals could receive prophylactic therapy with antibodies that neutralize key virulence determinants of staphylococcal BSI.
Work on staphylococcal vaccines commenced more than a century ago (80). Preclinical work over several decades indicates that immunization of immunocompetent mice with purified Hla, Coa, ClfA, or IsdB can elicit antigen-specific immune responses that protect animals from S. aureus BSI-associated mortality (24, 25, 34, 40, 47, 81). However, clinical trials with whole-cell killed (82) or subunit vaccines formulated from secreted virulence factors, α-hemolysin (Hla) and coagulase, did not protect against recurrent SSTI (29). Immunotherapy with antibodies neutralizing Hla (30, 83) or ClfA, the fibrinogen binding surface protein and agglutinin (40, 58), also did not protect against S. aureus infection (84, 85). Conjugates of S. aureus type 5/8 capsular polysaccharide (CP5/CP8) with Pseudomonas exotoxin A raise opsonophagocytic antibodies (23, 86); however, the vaccine did not protect hemodialysis patients from S. aureus infection (28). Finally, the clinical trial for V710, the recombinant IsdB vaccine (24, 87), was recently terminated; multiorgan dysfunction and mortality following S. aureus infection occurred more frequently in individuals receiving V710 than in control cohorts, and V710 immunization did not show clinical benefit (88). A key concept explaining recent failures of human vaccine trials with single staphylococcal antigens is based on the observation that, at least in the mouse model, S. aureus requires many different secreted products to cause disease (35, 39). If so, vaccine-induced neutralization of several different virulence factors may be required for the establishment of protective immunity (89). Researchers focusing on rabbits as models for S. aureus BSI or pneumonia identified secreted T cell superantigens as key virulence determinants and protective antigens (90). Clinical development of the corresponding superantigen vaccines has not yet advanced to a stage where this approach can be evaluated for efficacy (91). Further, T cell superantigen vaccines have not been evaluated for efficacy against S. aureus disease in leukopenic rabbits. Finally, clinical trials that examine vaccine or immunotherapeutic efficacy focus on health care-related S. aureus infections in patients at high risk for staphylococcal BSI, which includes, for example, end-stage renal disease with hemodialysis (28) or very-low-birth-weight neonates (92). These populations display at least partial defects in innate and adaptive immune responses; however, animal models that examine preclinical efficacy against S. aureus disease in an immunocompromised state have heretofore not been developed (93, 94).
Here, we used cyclophosphamide-induced leukopenia in mice to answer whether specific virulence factors/protective antigens contribute to the pathogenesis of S. aureus BSI. Our results demonstrate that the structural genes for several protective antigens (clfA, coa vwb, fnbB, hla, isdAB, sdrD, and spa) are not required for the pathogenesis of staphylococcal BSI in leukopenic mice. If so, vaccines designed to raise neutralizing antibodies against any one of these secreted products would not be expected to provide protection against S. aureus BSI in leukopenic patients. Nevertheless, an S. aureus sortase A (srtA) mutant, which cannot assemble any one of 18 LPXTG surface proteins into the bacterial envelope (including ClfA, FnBPB, IsdAB, SdrD, and SpA), displayed a significant decrease in virulence when tested in leukopenic mice, suggesting that a combinatorial vaccine or immunoprophylaxis targeting multiple surface proteins may be protective for staphylococcal BSI in patients with functional leukopenia.
Earlier work demonstrated that S. aureus infection in mice does not elicit protective immune responses that prevent subsequent infections with this pathogen (95). This phenotype requires S. aureus expression of SpA, the B cell superantigen, which inhibits the development of protective antibody responses (57). Mice that have been infected with spa mutants lacking the immunoglobulin binding attributes of SpA elicit antibody responses against many different antigens and provide protection against subsequent disease (57). A similar phenotype is observed in mice that have been infected with srtA mutants (37). Linear regression analysis was used to identify the antibody responses mediating protective immunity against S. aureus (37). This approach, designated genetic vaccinology, identified four surface protein antigens—ClfA, FnBPB, SdrD, and SpA—as protective antigens (37, 96). Indeed, when used as a combination vaccine in immunocompetent mice, ClfA, FnBPB, SdrD, and SpAKKAA (COMBO 2) elicit protective immunity against S. aureus BSI (37).
Here, we show that the surface protein vaccine (containing ClfA, FnBPB, SdrD, and SpAKKAA) elicits immune responses that protect leukopenic mice against S. aureus BSI. Of note, vaccine protection is limited to a delay in time to death in these severely immunocompromised mice. Although increases in survival were not recorded, we believe the observed protection may have clinical relevance for the following two reasons. First, chemotherapy of hematologic malignancy rarely diminishes the blood concentration of neutrophils and macrophages to the same degree as was achieved here with continued cyclophosphamide treatment of mice. If so, the residual opsonophagocytic capacities of chemotherapy or leukemia patients may elevate vaccine-induced protection in this patient population. Second, even the delayed-time-to-death protection affords clinical opportunities of initiating antibiotic therapies against S. aureus, which may enable recipients of the surface protein vaccine or specific immunotherapies (antibodies against surface proteins) to survive BSI.
Clearance of S. aureus from infected host tissues is dependent on opsonophagocytic killing (OPK) of the pathogen; however, staphylococci have evolved virulence strategies to resist OPK (97). A hallmark of S. aureus is the pathogen's ability to coagulate fibrinogen and to deploy surface proteins for its agglutination with fibrin fibrils as a means to resist OPK (40). Further, SpA blocks the effector functions of immunoglobulin specific for S. aureus, thereby interfering with staphylococcal OPK (98). Antibodies raised via the COMBO 2 vaccine target both of these virulence strategies by neutralizing fibrinogen binding proteins (clumping factors and fibronectin binding proteins) and SpA and may therefore promote OPK even in leukopenic mice whose abilities for phagocytic killing of staphylococci have been severely impaired (37).
ACKNOWLEDGMENTS
We thank members of our laboratory for helpful discussions.
This study was supported by grants from the National Institute of Allergy and Infectious Diseases, Infectious Diseases Branch (AI052474, AI075258, and AI110937 to O.S. and D.M.), and by Novartis Vaccines and Diagnostics (Siena, Italy). P.G. acknowledges support from the Molecular Cell Biology training grant (T32 GM007183). O.S. and D.M. acknowledge membership within and support from the Region V Great Lakes Regional Center of Excellence in Biodefense and Emerging Infectious Diseases Consortium (NIH award 1-U54-AI-057153).
H.K.K., O.S., and D.M. declare a conflict of interest as inventors of patent applications that are related to the development of Staphylococcus aureus vaccines and are currently under commercial license.
Footnotes
Published ahead of print 2 september 2014
REFERENCES
- 1.Lowy FD. 1998. Staphylococcus aureus infections. N. Engl. J. Med. 339:520–532. 10.1056/NEJM199808203390806. [DOI] [PubMed] [Google Scholar]
- 2.Kuehnert MJ, Kruszon-Moran D, Hill HA, McQuillan G, McAllister SK, Fosheim G, McDougal LK, Chaitram J, Jensen B, Fridkin SK, Killgore G, Tenover FC. 2006. Prevalence of Staphylococcus aureus nasal colonization in the United States, 2001-2002. J. Infect. Dis. 193:172–179. 10.1086/499632. [DOI] [PubMed] [Google Scholar]
- 3.Klevens RM, Morrison MA, Nadle J, Petit S, Gershman K, Ray S, Harrison LH, Lynfield R, Dumyati G, Townes JM, Craig AS, Zell ER, Fosheim GE, McDougal LK, Carey RB, Fridkin SK. 2007. Invasive methicillin-resistant Staphylococcus aureus infections in the United States. JAMA 298:1763–1771. 10.1001/jama.298.15.1763. [DOI] [PubMed] [Google Scholar]
- 4.DeLeo FR, Otto M, Kreiswirth BN, Chambers HF. 2010. Community-associated meticillin-resistant Staphylococcus aureus. Lancet 375:1557–1568. 10.1016/S0140-6736(09)61999-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Lessa FC, Mu Y, Davies J, Murray M, Lillie M, Pearson A, Fridkin SK. 2010. Comparison of incidence of bloodstream infection with methicillin-resistant Staphylococcus aureus between England and United States, 2006-2007. Clin. Infect. Dis. 51:925–928. 10.1086/656414. [DOI] [PubMed] [Google Scholar]
- 6.David MZ, Daum RS. 2010. Community-associated methicillin-resistant Staphylococcus aureus: epidemiology and clinical consequences of an emerging epidemic. Clin. Microbiol. Rev. 23:616–687. 10.1128/CMR.00081-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.DeLeo FR, Chambers HF. 2009. Waves of resistance: Staphylococcus aureus in the antibiotic era. Nat. Rev. Microbiol. 7:629–641. 10.1038/nrmicro2200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Kallen AJ, Mu Y, Bulens S, Reingold A, Petit S, Gershman K, Ray SM, Harrison LH, Lynfield R, Dumyati G, Townes JM, Schaffner W, Patel P, Fridkin SK. 2010. Health care-associated invasive MRSA infections, 2005-2008. JAMA 304:641–648. 10.1001/jama.2010.1115. [DOI] [PubMed] [Google Scholar]
- 9.Spellberg B, Daum RS. 2012. Development of a vaccine against Staphylococcus aureus. Semin. Immunopathol. 34:335–348. 10.1007/s00281-011-0293-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Puel A, Cypowyj S, Bustamante J, Wright JF, Liu L, Lim HK, Migaud M, Israel L, Chrabieh M, Audry M, Gumbleton M, Toulon A, Bodemer C, El-Baghdadi J, Whitters M, Paradis T, Brooks J, Collins M, Wolfman NM, Al-Muhsen S, Galicchio M, Abel L, Picard C, Casanova JL. 2011. Chronic mucocutaneous candidiasis in humans with inborn errors of interleukin-17 immunity. Science 332:65–68. 10.1126/science.1200439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Casanova JL, Holland SM, Notarangelo LD. 2012. Inborn errors of human JAKs and STATs. Immunity 36:515–528. 10.1016/j.immuni.2012.03.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Curnutte JT, Whitten DM, Babior BM. 1974. Defective superoxide production by granulocytes from patients with chronic granulomatous disease. N. Engl. J. Med. 290:593–597. 10.1056/NEJM197403142901104. [DOI] [PubMed] [Google Scholar]
- 13.Babior BM. 2004. NADPH oxidase. Curr. Opin. Immunol. 16:42–47. 10.1016/j.coi.2003.12.001. [DOI] [PubMed] [Google Scholar]
- 14.Chemaly RF, Hachem RY, Husni RN, Bahna B, Abou Rjaili G, Waked A, Graviss L, Nebiyou Bekele B, Shah JN, Raad II. 2010. Characteristics and outcomes of methicillin-resistant Staphylococcus aureus surgical-site infections in patients with cancer: a case-control study. Ann. Surg. Oncol. 17:1499–1506. 10.1245/s10434-010-0923-5. [DOI] [PubMed] [Google Scholar]
- 15.Skov R, Gottschau A, Skinhøj P, Frimodt-Møller N, Rosdahl VT, Espersen F. 1995. Staphylococcus aureus bacteremia: a 14-year nationwide study in hematological patients with malignant disease or agranulocytosis. Scand. J. Infect. Dis. 27:563–568. 10.3109/00365549509047068. [DOI] [PubMed] [Google Scholar]
- 16.González-Barca E, Carratalà J, Mykietiuk A, Fernández-Sevilla A, Gudiol F. 2001. Predisposing factors and outcome of Staphylococcus aureus bacteremia in neutropenic patients with cancer. Eur. J. Clin. Microbiol. Infect. Dis. 20:117–119. 10.1007/s100960000431. [DOI] [PubMed] [Google Scholar]
- 17.Emadi A, Jones RJ, Brodsky RA. 2009. Cyclophosphamide and cancer: golden anniversary. Nat. Rev. Clin. Oncol. 6:638–647. 10.1038/nrclinonc.2009.146. [DOI] [PubMed] [Google Scholar]
- 18.Bishop JF, Schimpff SC, Diggs CH, Wiernik PH. 1981. Infections during intensive chemotherapy for non-Hodgkin's lymphoma. Ann. Intern. Med. 95:549–555. 10.7326/0003-4819-95-5-549. [DOI] [PubMed] [Google Scholar]
- 19.Mahajan SN, Shah JN, Hachem R, Tverdek F, Adachi JA, Mulanovich V, Rolston KV, Raad II, Chemaly RF. 2012. Characteristics and outcomes of methicillin-resistant Staphylococcus aureus bloodstream infections in patients with cancer treated with vancomycin: 9-year experience at a comprehensive cancer center. Oncologist 17:1329–1336. 10.1634/theoncologist.2012-0029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Craig WA. 1998. Pharmacokinetic/pharmacodynamic parameters: rationale for antibacterial dosing of mice and men. Clin. Infect. Dis. 26:1–10. 10.1086/516284. [DOI] [PubMed] [Google Scholar]
- 21.Zuluaga AF, Salazar BE, Rodriguez CA, Zapata AX, Agudelo M, Vesga O. 2006. Neutropenia in outbred mice by a simplified low-dose cyclophosphamide regimen: characterization and applicability to diverse experimental models of infectious diseases. BMC Infect. Dis. 6:55. 10.1186/1471-2334-6-55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Daum RS, Spellberg B. 2012. Progress toward a Staphylococcus aureus vaccine. Clin. Infect. Dis. 54:560–567. 10.1093/cid/cir828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Fattom AI, Horwith G, Fuller S, Propst M, Naso R. 2004. Development of StaphVAX, a polysaccharide conjugate vaccine against S. aureus infection: from the lab bench to phase III clinical trials. Vaccine 22:880–887. 10.1016/j.vaccine.2003.11.034. [DOI] [PubMed] [Google Scholar]
- 24.Kuklin NA, Clark DJ, Secore S, Cook J, Cope LD, McNeely T, Noble L, Brown MJ, Zorman JK, Wang XM, Pancari G, Fan H, Isett K, Burgess B, Bryan J, Brownlow M, George H, Meinz M, Liddell ME, Kelly R, Schultz L, Montgomery D, Onishi J, Losada M, Martin M, Ebert T, Tan CY, Schofield TL, Nagy E, Meineke A, Joyce JG, Kurtz MB, Caulfield MJ, Jansen KU, McClements W, Anderson AS. 2006. A novel Staphylococcus aureus vaccine: iron surface determinant B induces rapid antibody responses in rhesus macaques and specific increased survival in a murine S. aureus sepsis model. Infect. Immun. 74:2215–2223. 10.1128/IAI.74.4.2215-2223.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Bubeck Wardenburg J, Schneewind O. 2008. Vaccine protection against Staphylococcus aureus pneumonia. J. Exp. Med. 205:287–294. 10.1084/jem.20072208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Menzies BE, Kernodle DS. 1996. Passive immunization with antiserum to a nontoxic alpha-toxin mutant from Staphylococcus aureus is protective in a murine model. Infect. Immun. 64:1839–1841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Duthie ES, Lorenz LL. 1952. Staphylococcal coagulase: mode of action and antigenicity. J. Gen. Microbiol. 6:95–107. 10.1099/00221287-6-1-2-95. [DOI] [PubMed] [Google Scholar]
- 28.Shinefield H, Black S, Fattom A, Horwith G, Rasgon S, Ordonez J, Yeoh H, Law D, Robbins JB, Schneerson R, Muenz L, Fuller S, Johnson J, Fireman B, Alcorn H, Naso R. 2002. Use of a Staphylococcus aureus conjugate vaccine in patients receiving hemodialysis. N. Engl. J. Med. 346:491–496. 10.1056/NEJMoa011297. [DOI] [PubMed] [Google Scholar]
- 29.Harrison KJ. 1963. Clinical trial of coagulase and alpha-hemolysin toxoids in chronic furunculosis. Br. Med. J. 2:149–152. 10.1136/bmj.2.5350.149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kernodle DS. 2011. Expectations regarding vaccines and immune therapies directed against Staphylococcus aureus alpha-hemolysin. J. Infect. Dis. 203:1692–1693. 10.1093/infdis/jir141. [DOI] [PubMed] [Google Scholar]
- 31.National Research Council. 2011. Guide for the care and use of laboratory animals, 8th ed. National Academies Press, Washington, DC. [Google Scholar]
- 32.Baba T, Bae T, Schneewind O, Takeuchi F, Hiramatsu K. 2007. Genome sequence of Staphylococcus aureus strain Newman and comparative analysis of staphylococcal genomes. J. Bacteriol. 190:300–310. 10.1128/JB.01000-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Mazmanian SK, Skaar EP, Gaspar AH, Humayun M, Gornicki P, Jelenska J, Joachmiak A, Missiakas DM, Schneewind O. 2003. Passage of heme-iron across the envelope of Staphylococcus aureus. Science 299:906–909. 10.1126/science.1081147. [DOI] [PubMed] [Google Scholar]
- 34.Cheng AG, McAdow M, Kim HK, Bae T, Missiakas DM, Schneewind O. 2010. Contribution of coagulases towards Staphylococcus aureus disease and protective immunity. PLoS Pathog. 6:e1001036. 10.1371/journal.ppat.1001036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Stranger-Jones YK, Bae T, Schneewind O. 2006. Vaccine assembly from surface proteins of Staphylococcus aureus. Proc. Nat. Acad. Sci. U. S. A. 103:16942–16947. 10.1073/pnas.0606863103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Kim HK, Cheng AG, Kim H-Y, Missiakas DM, Schneewind O. 2010. Non-toxigenic protein A vaccine for methicillin-resistant Staphylococcus aureus infections. J. Exp. Med. 207:1863–1870. 10.1084/jem.20092514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Kim HK, Kim HY, Schneewind O, Missiakas DM. 2011. Identifying protective antigens of Staphylococcus aureus, a pathogen that suppresses host immune responses. FASEB J. 25:3605–3612. 10.1096/fj.11-187963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Griffith DC, Rodriguez D, Corcoran E, Dudley MN. 2008. Pharmacodynamics of RWJ-54428 against Staphylococcus aureus, Streptococcus pneumoniae, and Enterococcus faecalis in a neutropenic mouse thigh infection model. Antimicrob. Agents Chemother. 52:244–247. 10.1128/AAC.00776-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Cheng AG, Kim HK, Burts ML, Krausz T, Schneewind O, Missiakas DM. 2009. Genetic requirements for Staphylococcus aureus abscess formation and persistence in host tissues. FASEB J. 23:3393–3404. 10.1096/fj.09-135467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.McAdow M, Kim HK, DeDenta AC, Hendrickx APA, Schneewind O, Missiakas DM. 2011. Preventing Staphylococcus aureus sepsis through the inhibition of its agglutination in blood. PLoS Pathog. 7:e1002307. 10.1371/journal.ppat.1002307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Kim HK, DeDent A, Cheng AG, McAdow M, Bagnoli F, Missiakas DM, Schneewind O. 2010. IsdA and IsdB antibodies protect mice against Staphylococcus aureus abscess formation and lethal challenge. Vaccine 28:6382–6392. 10.1016/j.vaccine.2010.02.097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.McAdow M, DeDent AC, Emolo C, Cheng AG, Kreiswirth BN, Missiakas DM, Schneewind O. 2012. Coagulases as determinants of protective immune responses against Staphylococcus aureus. Infect. Immun. 80:3389–3398. 10.1128/IAI.00562-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Gray GS, Kehoe M. 1984. Primary sequence of the alpha-toxin gene from Staphylococcus aureus Wood 46. Infect. Immun. 46:615–618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Wilke GA, Bubeck Wardenburg J. 2010. Role of a disintegrin and metalloprotease 10 in Staphylococcus aureus alpha-hemolysin-mediated cellular injury. Proc. Natl. Acad. Sci. U. S. A. 107:13473–13478. 10.1073/pnas.1001815107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Inoshima I, Inoshima N, Wilke GA, Powers ME, Frank KM, Wang Y-T, Bubeck-Wardenburg J. 2011. A Staphylococcus aureus pore-forming toxin subverts the activity of ADAM10 to cause lethal infection in mice. Nat. Med. 17:1310–1314. 10.1038/nm.2451. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Becker RE, Berube BJ, Sampedro GR, Dedent AC, Bubeck Wardenburg J. 2014. Tissue-specific patterning of host innate immune responses by Staphylococcus aureus α-toxin. J. Innate Immun. 6:619–631. 10.1159/000360006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Powers ME, Kim HK, Wang Y-T, Bubeck-Wardenburg J. 2012. ADAM10 mediates vascular injury induced by Staphylococcus aureus α-hemolysin. J. Infect. Dis. 206:352–356. 10.1093/infdis/jis192. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Much H. 1908. Über eine vorstufe des fibrinfermentes in kulturen von Staphylokokkus aureus. Biochem. Z. 14:143–155. [Google Scholar]
- 49.Bjerketorp J, Nilsson M, Ljungh A, Flock JI, Jacobsson K, Frykberg L. 2002. A novel von Willebrand factor binding protein expressed by Staphylococcus aureus. Microbiology 148:2037–2044. [DOI] [PubMed] [Google Scholar]
- 50.Friedrich R, Panizzi P, Fuentes-Prior P, Richter K, Verhamme I, Anderson PJ, Kawabata S, Huber R, Bode W, Bock PE. 2003. Staphylocoagulase is a prototype for the mechanism of cofactor-induced zymogen activation. Nature 425:535–539. 10.1038/nature01962. [DOI] [PubMed] [Google Scholar]
- 51.Haley KP, Skaar EP. 2012. A battle for iron: host sequestration and Staphylococcus aureus acquisition. Microbes Infect. 14:217–227. 10.1016/j.micinf.2011.11.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Torres VJ, Pishchany G, Humayun M, Schneewind O, Skaar EP. 2006. Staphylococcus aureus IsdB is a hemoglobin receptor required for heme-iron utilization. J. Bacteriol. 188:8421–8429. 10.1128/JB.01335-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Mazmanian SK, Liu G, Ton-That H, Schneewind O. 1999. Staphylococcus aureus sortase, an enzyme that anchors surface proteins to the cell wall. Science 285:760–763. 10.1126/science.285.5428.760. [DOI] [PubMed] [Google Scholar]
- 54.Ton-That H, Liu G, Mazmanian SK, Faull KF, Schneewind O. 1999. Purification and characterization of sortase, the transpeptidase that cleaves surface proteins of Staphylococcus aureus at the LPXTG motif. Proc. Natl. Acad. Sci. U. S. A. 96:12424–12429. 10.1073/pnas.96.22.12424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Mazmanian SK, Liu G, Jensen ER, Lenoy E, Schneewind O. 2000. Staphylococcus aureus mutants defective in the display of surface proteins and in the pathogenesis of animal infections. Proc. Natl. Acad. Sci. U. S. A. 97:5510–5515. 10.1073/pnas.080520697. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Goodyear CS, Silverman GJ. 2003. Death by a B cell superantigen: in vivo VH-targeted apoptotic supraclonal B cell deletion by a staphylococcal toxin. J. Exp. Med. 197:1125–1139. 10.1084/jem.20020552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Falugi F, Kim HK, Missiakas DM, Schneewind O. 2013. Role of protein A in the evasion of host adaptive immune responses by Staphylococcus aureus. mBio 4(5):e00575-13. 10.1128/mBio.00575-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.McDevitt D, Francois P, Vaudaux P, Foster TJ. 1994. Molecular characterization of the clumping factor (fibrinogen receptor) of Staphylococcus aureus. Mol. Microbiol. 11:237–248. 10.1111/j.1365-2958.1994.tb00304.x. [DOI] [PubMed] [Google Scholar]
- 59.Ganesh VK, Rivera JJ, Smeds E, Ko Y-P, Bowden MG, Wann ER, Gurusidappa S, Fitzgerald JR, Höök M. 2008. A structural model of the Staphylococcus aureus ClfA-fibrinogen interaction opens new avenues for the design of anti-staphylococcal therapeutics. PLoS Pathog. 4:e1000226. 10.1371/journal.ppat.1000226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Patti JM. 2004. A humanized monoclonal antibody targeting Staphylococcus aureus. Vaccine 22:S39–S43. 10.1016/j.vaccine.2004.08.015. [DOI] [PubMed] [Google Scholar]
- 61.Jönsson K, Signäs C, Müller HP, Lindberg M. 1991. Two different genes encode fibronectin binding proteins in Staphylococcus aureus. The complete nucleotide sequence and characterization of the second gene. Eur. J. Biochem. 202:1041–1048. [DOI] [PubMed] [Google Scholar]
- 62.Roche FM, Downer R, Keane F, Speziale P, Park PW, Foster TJ. 2004. The N-terminal A domain of fibronectin-binding proteins A and B promotes adhesion of Staphylococcus aureus to elastin. J. Biol. Chem. 279:38433–38440. 10.1074/jbc.M402122200. [DOI] [PubMed] [Google Scholar]
- 63.Wann ER, Gurusiddappa S, Höök M. 2000. The fibronectin-binding MSCRAMM FnbpA of Staphylococcus aureus is a bifunctional protein that also binds to fibrinogen. J. Biol. Chem. 275:13863–13871. 10.1074/jbc.275.18.13863. [DOI] [PubMed] [Google Scholar]
- 64.Burke FM, Di Poto A, Speziale P, Foster TJ. 2011. The A domain of fibronectin-binding protein B of Staphylococcus aureus contains a novel fibronectin binding site. FEBS J. 278:2359–2371. 10.1111/j.1742-4658.2011.08159.x. [DOI] [PubMed] [Google Scholar]
- 65.Shinji H, Yosizawa Y, Tajima A, Iwase T, Sugimoto S, Seki K, Mizunoe Y. 2011. Role of fibronectin-binding proteins A and B in in vitro cellular infections and in vivo septic infections by Staphylococcus aureus. Infect. Immun. 79:2215–2223. 10.1128/IAI.00133-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Grundmeier M, Hussain M, Becker P, Heilmann C, Peters G, Sinha B. 2004. Truncation of fibronectin-binding proteins in Staphylococcus aureus strain Newman leads to deficient adherence and host cell invasion due to loss of the cell wall anchor function. Infect. Immun. 72:7155–7163. 10.1128/IAI.72.12.7155-7163.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Xiong YQ, Bayer AS, Yeaman MR, Van Wamel W, Manna AC, Cheung AL. 2004. Impacts of sarA and agr in Staphylococcus aureus strain Newman on fibronectin-binding protein A gene expression and fibronectin adherence capacity in vitro and in experimental infective endocarditis. Infect. Immun. 72:1832–1836. 10.1128/IAI.72.3.1832-1836.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Becker S, Frankel MB, Schneewind O, Missiakas DM. 2014. Release of protein A from the cell wall envelope of Staphylococcus aureus. Proc. Natl. Acad. Sci. U. S. A. 111:1574–1579. 10.1073/pnas.1317181111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Freeman AF, Holland SM. 2009. Antimicrobial prophylaxis for primary immunodeficiencies. Curr. Opin. Allergy Clin. Immunol. 9:525–530. 10.1097/ACI.0b013e328332be33. [DOI] [PubMed] [Google Scholar]
- 70.Marín M, Gudiol C, Garcia-Vidal C, Ardanuy C, Carratalà J. 2014. Bloodstream infections in patients with solid tumors: epidemiology, antibiotic therapy, and outcomes in 528 episodes in a single cancer center. Medicine 93:143–149. 10.1097/MD.0000000000000026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Chambers HF. 2005. Community-associated MRSA—resistance and virulence converge. N. Engl. J. Med. 352:1485–1487. 10.1056/NEJMe058023. [DOI] [PubMed] [Google Scholar]
- 72.Popovich KJ, Weinstein RA, Aroutcheva A, Rice T, Hota B. 2010. Community-associated methicillin-resistant Staphylococcus aureus and HIV: intersecting epidemics. Clin. Infect. Dis. 50:979–987. 10.1086/651076. [DOI] [PubMed] [Google Scholar]
- 73.Craig WA, Andes DR. 2008. In vivo pharmacodynamics of ceftobiprole against multiple bacterial pathogens in murine thigh and lung infection models. Antimicrob. Agents Chemother. 52:3492–3496. 10.1128/AAC.01273-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Rice LB. 2009. Federal funding for the study of antimicrobial resistance in nosocomial pathogens: no ESKAPE. J. Infect. Dis. 197:1079–1081. 10.1086/533452. [DOI] [PubMed] [Google Scholar]
- 75.Guo B, Abdelraouf K, Ledesma KR, Chang KT, Nikolaou M, Tam VH. 2011. Quantitative impact of neutrophils on bacterial clearance in a murine pneumonia model. Antimicrob. Agents Chemother. 55:4601–4605. 10.1128/AAC.00508-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Thompson MG, Black CC, Pavlicek RLRL, Honnold CLCL, Wise MCMC, Alamneh YA, Moon JK, Kessler JL, Si Y, Williams R, Yildirim S, Kirkup BC, Jr, Green RK, Hall ER, Palys TJ, Zurawski DV. 2014. Validation of a novel murine wound model of Acinetobacter baumannii infection. Antimicrob. Agents Chemother. 58:1332–1342. 10.1128/AAC.01944-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Cryz SJ, Jr, Furer E, Germanier R. 1983. Passive protection against Pseudomonas aeruginosa infection in an experimental leukopenic mouse model. Infect. Immun. 40:659–664. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.von Specht BU, Knapp B, Muth G, Broker M, Hungerer KD, Diehl KD, Massarrat K, Seemann A, Domdey H. 1995. Protection of immunocompromised mice against lethal infection with Pseudomonas aeruginosa by active or passive immunization with recombinant P. aeruginosa outer membrane protein F and outer membrane protein I fusion proteins. Infect. Immun. 63:1855–1862. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Scarff JM, Goldberg JB. 2008. Vaccination against Pseudomonas aeruginosa pneumonia in immunocompromised mice. Clin. Vaccine Immunol. 15:367–375. 10.1128/CVI.00419-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Meakins J. 1910. An experimental study of opsonic immunity to Staphylococcus aureus. J. Exp. Med. 12:67–81. 10.1084/jem.12.1.67. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Palmqvist N, Patti JM, Tarkowski A, Josefsson E. 2004. Expression of staphylococcal clumping factor A impedes macrophage phagocytosis. Microb. Infect. 6:188–195. 10.1016/j.micinf.2003.11.005. [DOI] [PubMed] [Google Scholar]
- 82.Rogers DE, Melly MA. 1965. Speculation on the immunology of staphylococcal infections. Ann. N. Y. Acad. Sci. 128:274–284. [DOI] [PubMed] [Google Scholar]
- 83.Parish HJ, Cannon DA. 1960. Staphylococcal infection: antitoxic immunity. Br. Med. J. 1:743–747. 10.1136/bmj.1.5175.743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Weems JJ, Jr, Steinberg JP, Filler S, Baddley JW, Corey GR, Sampathkumar P, Winston L, John JF, Kubin CJ, Talwani R, Moore T, Patti JM, Hetherington S, Texter M, Wenzel E, Kelley VA, Fowler VG., Jr 2006. Phase II, randomized, double-blind, multicenter study comparing the safety and pharmacokinetics of tefibazumab to placebo for treatment of Staphylococcus aureus bacteremia. Antimicrob. Agents Chemother. 50:2751–2755. 10.1128/AAC.00096-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Projan SJ, Nesin M, Dunman PM. 2006. Staphylococcal vaccines and immunotherapy: to dream the impossible dream? Curr. Opin. Pharmacol. 6:473–479. 10.1016/j.coph.2006.04.005. [DOI] [PubMed] [Google Scholar]
- 86.Fattom A, Schneerson R, Szu SC, Vann WF, Shiloach J, Karakawa WW, Robbins JB. 1990. Synthesis and immunologic properties in mice of vaccines composed of Staphylococcus aureus type 5 and type 8 capsular polysaccharides conjugated to Pseudomonas aeruginosa exotoxin A. Infect. Immun. 58:2367–2374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Moustafa M, Aronoff GR, Chandran C, Hartzel JS, Smugar SS, Galphin CM, Mailloux LU, Brown E, Dinubile MJ, Kartsonis NA, Guris D. 2012. Phase IIa study of the immunogenicity and safety of the novel Staphylococcus aureus vaccine V710 in adults with end-stage renal disease receiving hemodialysis. Clin. Vaccine Immunol. 19:1509–1516. 10.1128/CVI.00034-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Fowler VG, Allen KB, Moreira ED, Moustafa M, Isgro F, Boucher HW, Corey GR, Carmeli Y, Betts R, Hartzel JS, Chan IS, McNeely TB, Kartsonis NA, Guris D, Onorato MT, Smugar SS, DiNubile MJ, Sobanjo-ter Meulen A. 2013. Effect of an investigational vaccine for preventing Staphylococcus aureus infections after cardiothoracic surgery: a randomized trial. JAMA 309:1368–1378. 10.1001/jama.2013.3010. [DOI] [PubMed] [Google Scholar]
- 89.DeDent A, Kim HK, Missiakas DM, Schneewind O. 2012. Exploring Staphylococcus aureus pathways to disease for vaccine development. Semin. Immunopathol. 34:317–333. 10.1007/s00281-011-0299-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Buonpane RA, Churchill HR, Moza B, Sundberg EJ, Peterson ML, Schlievert PM, Kranz DM. 2007. Neutralization of staphylococcal enterotoxin B by soluble, high-affinity receptor antagonists. Nat. Med. 13:725–729. 10.1038/nm1584. [DOI] [PubMed] [Google Scholar]
- 91.Salgado-Pabón W, Schlievert PM. 2014. Models matter: the search for an effective Staphylococcus aureus vaccine. Nat. Rev. Microbiol. 12:585–591. 10.1038/nrmicro3308. [DOI] [PubMed] [Google Scholar]
- 92.Weisman LE. 2007. Antibody for the prevention of neonatal nosocomial staphylococcal infection: a review of the literature. Arch. Ped. 14:S31–S34. 10.1016/S0929-693X(07)80008-X. [DOI] [PubMed] [Google Scholar]
- 93.Weisman LE, Thackray HM, Steinhorn RH, Walsh WF, Lassiter HA, Dhanireddy R, Brozanski BS, Palmer KG, Trautman MS, Escobedo M, Meissner HC, Sasidharan P, Fretz J, Kokai-Kun JF, Kramer WG, Fischer GW, Mond JJ. 2011. A randomized study of a monoclonal antibody (pagibaximab) to prevent staphylococcal sepsis. Pediatrics 128:271–279. 10.1542/peds.2010-3081. [DOI] [PubMed] [Google Scholar]
- 94.Shinefield HR, Black S. 2005. Prevention of Staphylococcus aureus infections: advances in vaccine development. Expert Rev. Vaccines 4:669–676. 10.1586/14760584.4.5.669. [DOI] [PubMed] [Google Scholar]
- 95.Burts ML, Williams WA, DeBord K, Missiakas DM. 2004. EsxA and EsxB are secreted by an ESAT-6-like system that is required for the pathogenesis of Staphylococcus aureus infections. Proc. Nat. Acad. Sci. U. S. A. 102:1169–1174. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Kim HK, Thammavongsa V, Schneewind O, Missiakas D. 2012. Recurrent infections and immune evasion strategies of Staphylococcus aureus. Curr. Opin. Microbiol. 15:92–99. 10.1016/j.mib.2011.10.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Cheng AG, DeDent AC, Schneewind O, Missiakas DM. 2011. A play in four acts: Staphylococcus aureus abscess formation. Trends Microbiol. 19:225–232. 10.1016/j.tim.2011.01.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Forsgren A, Nordström K. 1974. Protein A from Staphylococcus aureus: the biological significance of its interaction with IgG. Ann. N. Y. Acad. Sci. 236:252–266. 10.1111/j.1749-6632.1974.tb41496.x. [DOI] [PubMed] [Google Scholar]