

Abstract
Nitrous oxide (N2O) is a potent greenhouse gas and the predominant ozone depleting substance. The only enzyme known to reduce N2O is the nitrous oxide reductase, encoded by the nosZ gene, which is present among bacteria and archaea capable of either complete denitrification or only N2O reduction to di-nitrogen gas. To determine whether the occurrence of nosZ, being a proxy for the trait N2O reduction, differed among taxonomic groups, preferred habitats or organisms having either NirK or NirS nitrite reductases encoded by the nirK and nirS genes, respectively, 652 microbial genomes across 18 phyla were compared. Furthermore, the association of different co-occurrence patterns with enzymes reducing nitric oxide to N2O encoded by nor genes was examined. We observed that co-occurrence patterns of denitrification genes were not randomly distributed across taxa, as specific patterns were found to be more dominant or absent than expected within different taxonomic groups. The nosZ gene had a significantly higher frequency of co-occurrence with nirS than with nirK and the presence or absence of a nor gene largely explained this pattern, as nirS almost always co-occurred with nor. This suggests that nirS type denitrifiers are more likely to be capable of complete denitrification and thus contribute less to N2O emissions than nirK type denitrifiers under favorable environmental conditions. Comparative phylogenetic analysis indicated a greater degree of shared evolutionary history between nosZ and nirS. However 30% of the organisms with nosZ did not possess either nir gene, with several of these also lacking nor, suggesting a potentially important role in N2O reduction. Co-occurrence patterns were also non-randomly distributed amongst preferred habitat categories, with several habitats showing significant differences in the frequencies of nirS and nirK type denitrifiers. These results demonstrate that the denitrification pathway is highly modular, thus underpinning the importance of community structure for N2O emissions.
Introduction
Managing nitrogen (N) is one of the major environmental challenges for the 21st century [1], [2]. Of special concern is the mitigation of nitrous oxide (N2O) due to its status as a potent greenhouse gas and the predominant ozone depleting compound [3], [4]. While there are multiple sources of atmospheric N2O, the majority is produced by microbial transformations of reactive N in fertilized agricultural soil [5]–[7].
The only known sink for N2O in Earth's troposphere is microbial reduction to di-nitrogen (N2), catalyzed by the N2O reductase encoded by the nosZ gene. This unique enzyme is found among a wide range of bacterial and archaeal taxa capable of complete denitrification, an anaerobic respiratory process in which soluble nitrate (NO3 −) or nitrite (NO2 −) is reduced to the gases nitric oxide (NO), N2O and N2 via a series of enzymatic steps. It has been suggested that denitrification is a modular pathway, in that an organism may not always possess the full set of enzymes and thus perform only a subset of steps within the pathway [8]. Bacteria carrying nosZ genes that reduce N2O to N2 yet lack the other denitrification genes have been described [9]–[12], and have recently been shown to be potentially important consumers of N2O produced by denitrification or other N-cycling processes [13]. On the other hand, denitrifiers can also lack the nosZ gene [14], and thus produce N2O as a terminal product. It was recently shown that manipulation of the proportion of denitrifiers with the nosZ gene in soil microcosms significantly affected the ratio of N2O:N2 production, establishing a causal link between denitrifier community composition and potential N2O emissions [15]. Thus, the regulation of N2O emissions ultimately has a genetic basis.
Whether the genetic potential to produce or reduce N2O is more conserved among specific microbial taxa or functional groups of denitrifiers implies that the composition of the microbial community is not trivial from an ecosystem perspective, however this is not known. Denitrifiers can be divided into two functional groups based on whether they possess the copper binding dissimilatory nitrite reductase (NirK) or the cytochrome cd 1 variant (NirS) that catalyze the reduction of NO2 − to NO; the key reaction that defines denitrification [8], [16]. While the two enzymes perform the same function in the denitrification pathway, they are non-homologous and were thought to be mutually exclusive in the genomes of denitrifying organisms [14]. Several studies have shown that communities of denitrifiers with NirK respond differently to environmental gradients than those with NirS [17]–[21], which supports the hypothesis that the two communities occupy different ecological niches [22]. Thus, nosZ co-occurrence patterns in relation to denitrifying organisms with one or the other type of nitrite reductase can provide additional insight on the importance of denitrifier community composition for N2O reduction in different ecosystems.
To understand the ecological implications of whether NO2 − reduction is performed by NirK or NirS denitrifiers, the link to the enzymatic step reducing NO to N2O needs to be considered. This reaction may be performed by a number of different enzymes that vary in structure and evolutionary relationships, including periplasmic associated flavodiiron, flavorubredoxin, and cytochrome-c type proteins, as well as several variants of the membrane-bound heme-copper oxidases [23]. Whereas the periplasmic enzymes are generally associated with response to nitrosative stress [23], [24], respiratory NO reduction to N2O in the denitrification pathway is performed by two variants of the heme-copper oxidase type NO reductases in bacterial and archaeal denitrifiers, encoded by the cnorB or qnorB genes [25]. Homologs of the qnorB gene have also been observed in non-denitrifying pathogenic species as well as anaerobic denitrifying methanotrophs, and are believed to be involved in detoxification [26] and dismutation of NO to N2 and O2 [27], respectively. In fungal denitrifiers, NO reduction is performed by the P450-type nitric oxide reductase (P450nor), however this enzyme is not considered to be involved in energy conservation [28]. Regardless of the cellular role of each enzyme, total N2O production from a given ecosystem could be considered a sum of the activities of each N2O-genic NO reductase, although the contribution of qnorB and cnorB likely outweighs that of other Nor-types due to their prominent role in anaerobic respiration [24].
The aim of our study was to determine whether patterns of co-occurrence of nosZ, being a proxy for the trait N2O reduction, differed among nirK and nirS type denitrifiers, how this relates to taxonomic affiliation, and whether these patterns could be explained by the presence of a nor gene encoding one of the canonical NO reductases or the p450nor variant for fungal denitrifiers. This was investigated by examining the distribution of the genes nirK, nirS, nor and nosZ in 652 publicly available microbial genomes across 18 phyla. The increasing number of genomes from organisms found in a diverse range of environments [29] also allowed for an assessment of the modularity of the denitrification pathway in relation to preferred habitat. Furthermore, we examined how conserved the trait of N2O reduction is from an evolutionary perspective.
Materials and Methods
Data acquisition
A local database was constructed by downloading all 4135 draft and completed microbial genome nucleotide sequences available (November 2012) at the National Center for Biology Information (NCBI, www.ncbi.nlm.nih.gov). To ensure that homology searches were as comprehensive as possible, a two-step procedure was performed for each gene. First, an initial TBLASTN search [30] of the online NCBI microbial genomes database (www.ncbi.nlm.nih.gov/sutils/genom_table.cgi) was performed using translated nirS, nirK, and nosZ gene sequences from either Paracoccus denitrificans PD1222 or Bradyrhizobium japonicum USDA110 as queries. Resulting hits were then translated to amino acid sequences and aligned using SATÉ v2.2.3 [31] with MAFFT [32] as aligner, MUSCLE as merger and RAxML [33] as the tree estimator. Gene identity of the retrieved sequences was confirmed by examining the amino acid alignments in relation to characterized homologs, with emphasis on conserved positions crucial for protein functioning and phylogenetic inference (see below). The resulting amino acid alignments of nirK, nirS and nosZ, with 477, 150 and 282 sequences, respectively, were then used to create Position Specific Score Matrices (PSSM) [34] for conducting a more comprehensive PSI-TBLASTN search of the downloaded database. Truncated sequences and sequences with stop codons were excluded, and redundancy in the data set was reduced by eliminating different strains of the same species with identical nirK, nirS and nosZ amino acid sequences. Strains with identical sequences were kept when a unique co-occurrence pattern of denitrification genes was observed, or when the sequence of another denitrification gene was not identical, resulting in a dataset of 652 organisms (see Table S1 for species name, NCBI taxon ID, project name). We then searched the final set of genomes for homologues of the qnorB and cnorB variants of the NO-reductase. This was performed in a similar manner as described for the nir and nos genes, with the exception that the PSSM was generated by downloading the 10 most diverse representative cNorB and qNorB amino acid sequences from the NCBI conserved domains database (http://www.ncbi.nlm.nih.gov/Structure/cdd/cdd.shtml) to allow for an equal representation of both variants within the initial PSSM. For the eukaryotic species, the amino acid sequence for the P450nor from Fusarium oxysporum [35] was used as a query for TBLASTN searches of each fungal genome, and the resulting hits were aligned to the query sequence to both correctly identify P450nor based on previously described conserved amino acid positions [28], as well as to aid in assembly of exons.
Small subunit (SSU) rRNA gene sequences corresponding to the organisms were retrieved from the local genome database using Infernal [36]. In cases where there was more than one SSU rRNA gene sequence in a genome, the longest sequence was chosen. Taxonomic assignment was based on NCBI classification, which was verified by classification of SSU sequences using the SILVA database [37]. In addition, habitat and isolation source was either downloaded from the Genomes online database (GOLD, 2012 November 15, www.genomesonline.org/) [29] or searched for in NCBI using the taxon ID of the respective genome and looking at connected publications when available.
Phylogenetic tree reconstruction
Preliminary amino acid alignments of full-length gene sequences of nirK, nirS, norB, nosZ, and P450nor were created using the MUSCLE alignment algorithm [38] in the Geneious bioinformatics software suite (version 5.6.1, Biomatters, Auckland, New Zealand) with default settings. With these alignments as input, the most suitable substitution models were inferred by ProtTest v3.2 [39], those being LG+Γ for nirK and nosZ, LG+Γ+F for norB and nirS [40], and WAG+Γ+F for P450nor. Subsequently, new alignments were inferred using SATÉ, and after manual inspection of the resulting alignments maximum likelihood (ML) phylogenetic trees were inferred using 30 RAxML tree searches and selecting the best likelihood topology, followed by bootstrap analysis with 250 replicates. Small subunit rRNA sequences were aligned using SINA [41]. After manual improvement of the resulting alignment, the phylogeny was determined using 50 tree searches in RAxML with GTRCAT as the nucleotide substitution model followed by 500 bootstrap replicates. All trees were displayed and annotated using the Interactive Tree of Life (iTOL) online tool [42].
Phylogenetic distribution and conservation of nosZ among nirK- and nirS denitrifiers
To compare the phylogenetic pattern of N2O reduction between denitrifiers with nirK and nirS, the nosZ gene was used as a proxy for the capacity of an organism to reduce N2O and mapped onto the corresponding taxa within the nirK and nirS phylogenies. Quantification of the phylogenetic patterns of nosZ distribution for both genes was performed using Fritz and Purvis' [43] measure of phylogenetic signal strength (D), implemented in the ‘caper’ package within the R statistical programming environment (R foundation for statistical computing, Vienna, Austria). This statistic measures whether a trait is highly clumped (D<0), displays clumping patterns due to Brownian evolutionary processes (D∼0) or random distribution patterns (D = 1), or is highly over-dispersed (D>1) across a given phylogeny. In order to also determine the degree to which the ability to reduce N2O is conserved amongst nirK and nirS denitrifiers, we used the phylogenetic metric ConsenTRAIT [44] to calculate trait depths (τD) of nosZ as a trait on nirK and nirS gene trees in the R environment. Trait depth is a measure of the average distance between organisms exhibiting the trait and their last common ancestor, such that increasing values of τD indicate more conserved traits.
Gene co-occurrence patterns in relation to taxa and habitat
The genomic co-occurrence patterns of nirK, nirS, nor and nosZ were investigated in relation to taxonomic affiliation based on the 16S rRNA gene phylogeny at different taxonomic ranks. In order to determine whether the co-occurrence patterns of nir and nos genes were randomly distributed among taxa, contingency tables of co-occurrence patterns by taxonomic affiliation at the phylum, class, order and family was performed, followed by Chi-squared goodness of fit tests in the R environment. Lower taxonomic ranks were not examined due to low frequencies (<5) in the majority of cells. In addition, the standardized residuals of the resulting cell values were examined to distinguish taxa that differed significantly from the expected values for each pattern of co-occurrence, and presented as a mosaic plot generated in the R environment using the ‘vcd’ package. For each possible combination of nosZ and nir genes, the fraction harboring a nor gene was determined.
The co-occurrence patterns of nirK, nirS and nosZ were also correlated with available information on the organism's lifestyle as determined by habitat preference and for each possible combination, the fraction harboring a nor gene was determined. Information on habitat preference could be retrieved for 626 organisms that were grouped into eight general habitat categories: wastewater treatment plants, extremophilic, marine systems, fresh water systems, soil, plant, animal, and food associated organisms, and those found in multiple habitats. Here, extremophilic represents habitats for thermo-, psychro-, halo-, acido- and alkalophiles as well as oil degrading, radiation resistant and arsenic resistant organisms. The plant and animal associated organisms are specified as being directly associated to a plant or animal host, whereas food associated are those used in food production. Organisms isolated from multiple sources without any specific preferences were lumped into the ‘multiple habitats’ category. The different categories of lifestyle versus the gene occurrence patterns were tested in the same way as for taxa affiliation.
Results
Genomic nirK, nirS, nor and nosZ co-occurrences
In total, 652 organisms from all three domains of life were found to harbor denitrification genes (Fig. 1; for greater detail see Fig. S1 and Table S1). Of these, 458 and 110 possessed a nirK or nirS homolog, respectively, including ten genomes that harbored both nir genes. As stated earlier, we limited our search of NO-reductases to the cnorB and qnorB variants encoding NorB found in the canonical denitrification pathway, as well as the p450nor variant for fungal denitrifiers. This resulted in a total of 431 organisms with a nor gene, whereas 314 genomes were found to possess a nosZ gene. Interestingly, a large part of the nirK organisms (35%) had a truncated pathway lacking a canonical nor and a majority (70%) did not have nosZ. By contrast, only 3.6% of the nirS-type denitrifiers lacked nor and 20% did not have a nosZ gene. Of all organisms harboring nosZ, 30% did not possess any nir genes and about 24% did not have nor. In general, genomes containing both nosZ and nor also had a nir gene, whereas organisms with nosZ that did not have a nor gene also lacked a nir gene (Fig. 2a and d).
Figure 1. Maximum likelihood phylogeny of full-length 16S/18S rRNA sequences from 652 organisms with denitrification genes.

The inner colored ring represents taxonomic affiliation as indicated by the legend. The four outer bar-chart rings show the presence of nirK (orange), nirS (purple), nor (turquoise) and nosZ (magenta). Bar height represents the number of copies (≤4). Bootstrap values >70% are indicated by grey circles, and the scale bar denotes nucleotide substitution rate (GTR+Γ). Classification is based on the SILVA database with denomination according to NCBI taxonomy. For NCBI taxon ID number and project name, see Table S1.
Figure 2. Co-occurrence of nosZ with nirK or nirS in genomes with and without nor genes.
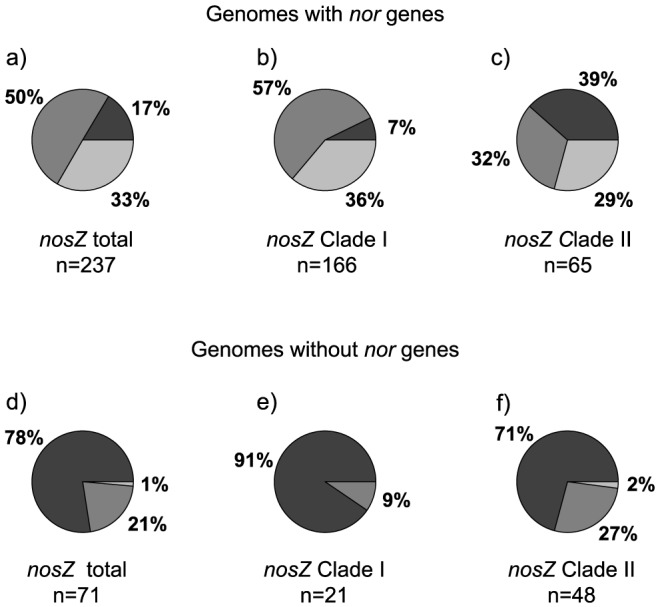
Percentage of genomes with only nosZ (black), nosZ and nirK (dark grey) and nosZ and nirS (light grey) among organisms a) harboring nosZ and nor, b) within nosZ Clade I with nor, c) within nosZ Clade II with nor, d) harboring nosZ without nor, e) within nosZ Clade I without nor, and f) within nosZ Clade II without nor. Six genomes that have both nirS and nirK in addition to nosZ are excluded as well as eight halophilic Archaea that group outside Clade I and II in b) and c).
It was recently established that the N2O reductase can be divided into two phylogenetic clades (I and II; Fig. S5), with clade II being a previously unaccounted, yet environmentally abundant lineage [45], [46]. Of the genomes with nosZ, 187 and 113 harbored lineages from clade I and II, respectively and 89 and 56% of these also had a nor gene. Among those with nor, organisms that also possessed nirK dominated in clade I, whereas in clade II the co-occurrence patterns were evenly distributed (Fig. 2b and c, respectively). By contrast, nosZ-harboring genomes lacking nor most often also lacked nir genes (Fig. 2d–f).
Analysis of denitrification gene phylogeny and nosZ conservation
Phylogenetic analysis of amino acid alignments resulted in well-supported tree topologies, with 48%, 63%, 62% and 66% of nodes having bootstrap support greater than 70% for nirK, nirS, norB, and nosZ phylogenies, respectively (Figs. S2–S5). Similar to previous reports, the nirK phylogeny (Fig. S2) could be divided into two major clusters [47], each consisting of sequences from a highly diverse range of phyla with no clear correspondence with organismal phylogeny [14]. While the occurrence of organisms with nosZ was dispersed throughout the overall nirK phylogeny, the Group 1 cluster had a noticeably higher number of organisms with nosZ than the Group 2 cluster, which was dominated by sequences from genomes that lack the nosZ gene. Similarly, nirK from the eukaryotic genomes formed a well-supported monophyletic clade that also corresponded with the lack of nosZ genes. This pattern of a more clumped occurrence of nosZ across the nirK phylogeny was confirmed by the calculated phylogenetic signal strength (D) of 0.335, which indicates a moderately clumped distribution that was significantly different from both random (p<0.001) and Brownian distributions (p = 0.002) of phylogenetic trait patterns.
The norB phylogeny could be clearly divided into cnorB and qnorB variants, with a diverse range of bacterial and archaeal phyla found within each clade (Fig. S4). However, a greater proportion of organisms with nirS were found within the cnorB clade, whereas those with nirK and nosZ were dispersed more equally across both clades.
In contrast to nirK and norB, the nirS and nosZ phylogenies (Figs. S2 and S3) were more congruent to taxonomic affiliation on the phylum level, with some exceptions observed within each tree. While the few nirS genomes lacking nosZ appeared to be randomly distributed throughout the nirS phylogeny, sequences from genomes lacking nosZ within the phyla Deinococcus-Thermus and Chloroflexi each formed monophyletic clades within the nirS phylogeny. This was reflected in the resulting phylogenetic signal strength (D = 0.431) that was also significantly non-random (p<0.001) and did not correspond to a Brownian process (p = 0.02), yet was slightly less clumped than the pattern of nosZ distribution observed for the nirK phylogeny. Additional analysis using the ConsenTRAIT metric (τD), performed using nosZ occurrence as a proxy for the trait N2O reduction on both the nirS and nirK phylogenies, indicated a higher degree of evolutionary conservation of nosZ with nirS (τD = 0.089) than nirK (τD = 0.043).
Gene co-occurrence patterns in relation to taxa
The SSU tree showing the distribution of nir, nor and nos genes (Fig. 1) indicated that patterns of gene co-occurrence were not randomly distributed across different taxonomic groups. Chi-squared tests of taxonomy-based contingency tables confirmed that the patterns nirK only (K-type), nirK+nosZ (KZ-type), nirK+nirS (KS-type), nirK+nirS+nosZ (KSZ-type), nirS only (S-type), nirS+nosZ (SZ-type) and only nosZ (Z-type) were indeed not randomly distributed amongst groupings at either the phylum, class, or order levels (P<0.001; Tables 1, S2 and S3). Calculation of standardized χ2 residuals highlighted several taxonomic groups that significantly contributed to the resulting chi-squared values. Within the largest represented phylum, the proteobacteria, nearly half of all organisms had a complete denitrification pathway including nir, nor and nosZ and the majority had nir and nor (Table 1). The K and KZ patterns dominated among the Alphaproteobacteria, largely due to the overrepresentation of these patterns within the order Rhizobiales (Table S3). However, the negative χ2 residual values observed for the abundance of K-only patterns indicates that proteobacteria in general are less likely to have a partial pathway, particularly Beta- and Gammaproteobacteria (Tables 1 and S2). This is further supported by the abundances of SZ and KZ patterns, which were higher than expected by chance within the proteobacteria, as well as the high percentage of organisms harboring nor genes for each pattern. The SZ pattern was twice as frequent as the KZ or Z patterns among the Gammaproteobacteria, which could be attributed to the Pseudomonadales. Within the Betaproteobacteria, the SZ- and KZ-types were equally represented, while Z-types were significantly underrepresented. SZ-type organisms within the Betaproteobacteria were mainly represented by the orders Burkholderiales and Rhodocyclales, whereas the KZ-type Betaproteobacteria were in turn primarily associated with the orders Neisseriales and Burkholderiales. The K-and Z-type Alpha- and Gammaproteobacteria lacked norB to a significantly greater extent than those within the Beta- and Deltaproteobacteria.The Ascomycota and Actinobacteria phyla were exclusively K-type organisms, whereas Z-type and KZ-type organisms were both overrepresented within the Bacteroidetes phylum (Table 1). A third of all KZ-type Bacteroidetes did not have a norB gene, which can be attributed to the Flavobacteriales (Tables 1 and S3).
Table 1. Frequency table of nirK (K), nirS (S) and nosZ (Z) co-occurrence types and taxonomic affiliation at phylum level as well as the percentage of organisms within each phylum that also harbor a nor gene.
| Phylum | K | %nor | KS | %nor | KSZ | %nor | KZ | %nor | S | %nor | SZ | %nor | Z | %nor |
| Actinobacteria | 102↑ | 42 | 0 | - | 0 | - | 0↓ | - | 0 | - | 0↓ | - | 0↓ | - |
| Aquificae | 0 | - | 0 | - | 0 | - | 0 | - | 0 | - | 2↑ | 100 | 0 | - |
| Ascomycota | 27↑ | 67 | 0 | - | 0 | - | 0↓ | - | 0 | - | 0 | - | 0↓ | - |
| Bacteroidetes | 8↓ | 100 | 0 | - | 1 | 0 | 24↑ | 67 | 0 | - | 0↓ | - | 24↑ | 17 |
| Chlorobi | 0 | - | 0 | - | 0 | - | 0 | - | 0 | - | 0 | - | 2↑ | 0 |
| Chloroflexi | 5 | 0 | 0 | - | 0 | - | 1 | 0 | 2↑ | 0 | 0 | - | 2 | 0 |
| Crenarchaeota | 0 | - | 0 | - | 0 | - | 0 | - | 2↑ | 100 | 2↑ | 100 | 0 | - |
| Deferribacteres | 0 | - | 0 | - | 0 | - | 0 | - | 0 | - | 0 | - | 1↑ | 0 |
| Deinococcus-Thermus | 0↓ | - | 2↑ | 100 | 0 | - | 0 | - | 3↑ | 100 | 0 | - | 0 | - |
| Euryarchaeota | 5 | 60 | 0 | - | 0 | - | 4 | 100 | 0 | - | 0 | - | 5↑ | 0 |
| Firmicutes | 16 | 25 | 0 | - | 0 | - | 4 | 100 | 0 | - | 1 | 100 | 8↑ | 12 |
| Gemmatimonadetes | 0 | - | 0 | - | 0 | - | 1 | 0 | 0 | - | 0 | - | 0 | |
| Nitrospirae | 1 | 0 | 0 | - | 0 | - | 0 | - | 1↑ | 100 | 0 | - | 0 | |
| Planctomycetes | 1 | 0 | 0 | - | 0 | - | 0 | - | 0 | - | 0 | - | 0 | |
| Proteobacteria | 135↓ | 66 | 2 | 100 | 5 | 100 | 96↑ | 96 | 12 | 92 | 74↑ | 99 | 47 | 51 |
| Spirochaetes | 2 | 50 | 0 | - | 0 | - | 3 | 33 | 0 | - | 1 | 100 | 3 | 67 |
| Thaumarchaeota | 9↑ | 0 | 0 | - | 0 | - | 0 | - | 0 | - | 0 | - | 0 | - |
| Verrucomicrobia | 3 | 67 | 0 | - | 0 | - | 1 | 0 | 0 | - | 0 | - | 2 | 0 |
Residuals are according to Pearson Chi-squared test (P<0.001).
Combinations with Pearson residuals >2.
Combinations with Pearson residuals <−2.
The lack of a nir gene was particularly notable among species within the Bacteroidetes (42%), but also within the Deltaproteobacteria (71.4%), Firmicutes (27.5%), and Euryarchaeota (35.7%), with the frequencies of Z-type organisms being significantly higher than expected within these groups (Tables 1 and S2). The occurrence of S-type organisms was higher than expected within the Deinococcus-Thermus phylum. Ten genomes carried both a nirK and a nirS gene (Fig. 1; Table S1), including Gammaproteobacterium HdN1, Rhodothermus marinus and four strains of Pseudomonas stutzeri that also possessed a nosZ gene. Interestingly, the Bacteroidete Rhodothermus marinus did not have a nor gene although the other three genes were present.
Gene co-occurrence patterns in relation to habitat preference
When comparing data relating to lifestyle in terms of habitat preference with nir, nor and nosZ gene co-occurrences, a significantly non-random pattern of distribution was observed across the different nir and nos categories (χ2 = 136, P<0.001; Fig. 3). Examination of standardized residuals revealed that K-type organisms were significantly overrepresented among animal host associated lifestyles, whereas the frequencies of both S- and SZ-types were significantly lower than expected in this category. By contrast, S-types were overrepresented among soil and extreme habitats, while K-types did not deviate from the expected frequency of occurrence or were underrepresented in each category, respectively. Contrasting patterns of K- and SZ-types were also observed in wastewater and multiple habitat categories, which had higher than expected frequencies of SZ-type organisms. Less than half (47%) of the K-type genomes did not have a nor gene and the percentage was especially low among organisms isolated from soil. However, in fresh water systems and among plant hosts 67 and 76%, respectively harbored nor. Interestingly, in wastewater the occurrence of organisms with both nir genes (KS) was significantly higher than expected, as well as organisms with a KSZ pattern. The frequency of organisms with only nosZ was overrepresented in marine ecosystems, yet lower than expected in the category of plant host-associated lifestyles. The Z-type organisms exhibited a low occurrence of nor in marine (12%) and host associated environments, whereas the opposite was observed in fresh water, soil and wastewater.
Figure 3. Gene co-occurrence types and habitat preference.

a) Mosaic plot of nirK (K), nirS (S) and nosZ (Z) co-occurrence types across different habitat categories where tile size reflects the number of occurrences and patterns indicate significant overrepresentation (>2) or underrepresentation (<−2) of co-occurrence patterns, as determined by standardized Pearson residuals from χ2 test results (P<0.001). Circles indicate non-occurring combinations. b) The percentage of organisms harboring a nor gene for each combination across the habitat categories. No value indicates non-occurring combinations.
Discussion
The genome data-set analyzed in this study demonstrated that the denitrification pathway is modular with no less than seven different co-occurrence types of the genes coding for the reductases NirK, NirS and NosZ. Interestingly, we found ten organisms that possess both a nirK and a nirS gene in the same genome. Although that is still rare, it contradicts the previous assumption that the two nitrite reductases are mutually exclusive [14]. The presence of homologues for both nir genes was previously reported in the genome of a Methylomonas strain [48]. This may be significant in light of recent findings by [49], who demonstrated the co-occurence of both nir genes in co-existing subpopulations affiliated to Pseudovibrio detected in the metagenome of a marine denitrifying community. Nevertheless, it has not yet been demonstrated if the two types of nitrite reductases are functional when present in the same genome, although it is not unlikely considering that two divergent copies of nirS in a Thauera sp. isolate were expressed under different conditions [50].
The nosZ gene was used as a proxy for the capacity of an organism to reduce N2O and we found that this gene occurred much more frequently in the genomes of organisms with nirS than in those with nirK. This was previously suggested [14], although the limited number of observations at that time did not support a significant pattern. The presence or absence of a nor gene partly explained this pattern, as nirS almost always co-occurred with nor, although there were exceptions. The substantial difference in co-occurrence of nosZ with nirS vs. nirK in the genome data-set is likely governed by an underlying ecological or evolutionary mechanism that constrains the loss of nor and nosZ in organisms with nirS. This is also supported by the higher degree of phylogenetic conservation observed for nosZ presence when mapped onto the nirS phylogeny than onto that of nirK. Previous studies have shown that loss of the nos regulon may occur through either short-term adaptive processes, such as phase variation in nirK Azospirillum soil isolates [51], or more long-term gene loss events as observed for nirK Neisseria species [52]. It is tempting to attribute the higher frequency of nirS, nor and nosZ co-occurrence to regulatory mechanisms that may be shared between the three enzyme complexes, but not with NirK, which is fully functional without accessory proteins. The expression of all three enzymes is controlled by oxygen and NO levels within the cell [11], [53] and initiated by the transcription factors NNR, NnrR or DNR [54], however no specific link between nirS, nor and nosZ expression has been described to date. Based on the gene co-occurrence patterns, the majority of nirS organisms are more likely to perform complete denitrification to N2 than the nirK types under favorable environmental conditions and we suggest that the presence of nor in the genome of an organism also harboring nir or nosZ serve as an indicator of whether the organism is a denitrifier sensu stricto or merely a nitrite or nitrous oxide reducer.
In the context of different co-occurrence patterns with nosZ, it is interesting that a range of studies have shown evidence of niche differentiation between nirS and nirK denitrifying communities [17]–[19], [21], [55]. If the co-occurrence patterns found in our genome comparisons are reflected in nature, an ecosystem in which the denitrifier community is dominated by nirK bearing organisms would be more likely to emit more N2O than one dominated by denitrifiers with nirS under the same environmental conditions. Indeed, the ratio of nirS/nirK type denitrifiers was recently demonstrated to have a significant and negative relationship with the capacity of a soil to act as an N2O sink when modeling the relative influence of abiotic and denitrifier community factors on the potential for soil N2O reduction [13]. This is also supported by previous findings. Clark et al. [56] reported a positive correlation between nirK abundance and N2O emissions from arable and woodland soils, and based on qPCR measurements estimated that only 10–30% of nirK type denitrifiers also carry the nosZ gene, depending on soil type. Similarly, Cuhel et al. [57] described a negative relationship between the ratio of N2O emissions to total denitrification activity (i.e. N2 + N2O) and the relative abundance of nirS-type denitrifiers in pastureland soils.
A highly truncated version of the denitrification pathway was observed among the substantial fraction of organisms that possessed the nosZ gene without having nir or a putatively N2O producing nor gene, and these were predominantly found within nosZ clade II. Several representatives from this clade, including the host-associated bacteria Wolinella succinogenes and Campylobacter fetus, as well as the thermophile Geobacillus thermodenitrificans and the soil bacterium Anaeromyxobacter dehalogenans, have been demonstrated to grow with N2O as the single electron acceptor [10], [12], [58]–[60]. An alternative, dissimilatory function for the nitrous oxide reductase has been recently proposed [61], but from an ecosystem perspective this detoxifying instead of energy conserving reaction would still result in N2O reduction. Net negative N2O fluxes in field measurements have been reported [62], [63] and recently it was demonstrated that both the abundance and phylogenetic diversity of nosZ clade II largely explained the soil N2O sink capacity [13]. The authors also showed that groups that were identified as significant indicators of N2O reduction were predominant in clade II communities, especially those associated with nosZ II lineages from organisms lacking nir. Because clade II has been shown to be both diverse and abundant in different ecosystems [45], [46], these organisms in particular can potentially be significant N2O consumers in the environment.
Although denitrification as a trait is widely distributed across different phyla, the observed nir/nos co-occurrence patterns were not randomly distributed across taxa, from phylum down to order level, with significant patterns related to certain taxonomic groups. The data set was too small to detect any patterns at lower taxonomic ranks. Accordingly, closely related organisms do not necessarily share this trait, and even if they do the denitrification genes may be distantly related or they may not have the same set of genes [8], [14], [64]–[66]. Organisms harboring a nosZ gene without possessing any nir gene were mainly found amongst the Bacteroidetes, and Firmicutes belonging to nosZ clade II, and predominantly lacked a nor gene. Thus, environments in which these taxa are highly abundant may have substantially lower N2O emission rates than those dominated by organisms that only have nir and nor genes, such as several representatives among the Actinobacteria and Ascomycota. This coincides with the results of Philippot et al [67], [68],who investigated the abundance of different taxa at high taxonomic ranks, as well as the abundance of different denitrifier genes and potential N2O production and denitrification rates on a field that was subjected to different cattle grazing regimes. Interestingly, the percentage of N2O to total denitrification activity (N2 + N2O) was lowest in the region of the field with the highest relative abundance of Bacteroidetes and lowest abundance of Actinobacteria. Furthermore, regions of the field in which nirS and nosZ clade I were most abundant also had the highest relative abundance of Betaproteobacteria, corresponding to our finding that the co-occurrence of nirS and nosZ is the predominant pattern of denitrification genes within this class. Thus, general patterns of gene co-occurrences in conjunction with taxonomic information can potentially aid in our interpretation of experimental data and predictions of genetic N2O emission potential.
Comparisons of gene co-occurrence patterns to preferred habitat categories showed differential associations of denitrifier gene combinations with specific habitats or lifestyles. Most notable was the prevalence of K-types over S-types amongst animal host associated organisms, suggesting that nirK might be involved to a greater extent during animal host adaptation and pathogenicity. Previous studies examining the relationship between nitrite reduction and pathogenicity of Neisseria and Brucella species have indicated that nirK is expressed primarily to help the bacterium cope with low oxygen levels within the host [69], [70]. However, nirS has also been shown to be important for virulence in Pseudomonas aeruginosa, albeit by regulation of virulence factors through maintaining a steady state of NO within the host [71]. It is known that prokaryotes undergo a process of genome shrinkage and gene loss during adaptation to animal hosts [72]. Since the nirK gene alone is sufficient to encode a functional protein while nirS requires several accessory genes [73], it may be more advantageous for host associated organisms to possess nirK rather than nirS. Interestingly, both the KS- and KSZ- types were significantly overrepresented in wastewater. One could speculate that carrying both types of nir genes could imply an adaptive advantage in an environment supporting denitrification activity, provided that both genes are transcribed to functional nitrite reductases under different conditions. The SZ type was more often found among genomes isolated from wastewater and freshwater than expected by chance. In agreement, it has been suggested that nirS denitrifiers are better adapted to stable, high water content conditions where oxygen availability is expected to be consistently low, while nirK communities tend to dominate under more recurrently changing conditions [74]. Other studies have also indicated that nirS communities are more frequently detected, more diverse and more phylogenetically clustered in marine systems than nirK communities [22], suggesting that nirS denitrifiers dominate in this habitat and that nirS denitrifying communities are shaped by habitat filtering more so than nirK communities. However, our analysis based on habitat data recovered from the GOLD database does not provide enough data or detail to confirm these hypotheses.
In conclusion, our results show that the co-occurrences of denitrification genes are not randomly distributed among taxonomic groups, preferred habitats or nirK and nirS denitrifiers. Although N2O emissions are subject to environmental factors and inherent cellular regulatory mechanisms, the ultimate control over whether N2O is emitted during denitrification is the presence of nosZ in the genomes of organisms that comprise the denitrifying community. Thus, the substantial difference between taxa as well as nirS, nirK and nor co-occurrence with nosZ likely plays a significant role in determining the genetic N2O emission potential from a given ecosystem, thereby underpinning the significance of microbial community structure for N2O emissions. Further research examining the underlying physiological or evolutionary mechanisms that result in the non-random patterns of nir/nor/nos gene occurrence observed here may provide additional predictive value for future N2O mitigation strategies.
Supporting Information
Maximum likelihood phylogeny of full-length 16S and 18S rRNA sequences from 652 organisms harboring denitrification genes. The inner colored ring represents taxonomic affiliation as indicated by the legend. The four outer bar-chart rings show the presence of nirK (orange), nirS (purple), nor (turquoise) and nosZ (magenta). Bar height represents the number of copies (≤4). Bootstrap values >70% are indicated by grey circles, and the scale bar denotes nucleotide substitution rate (GTR+Γ).
(EPS)
Maximum likelihood phylogeny of 458 full-length nirK amino acid sequences, rooted at midpoint. Co-occurrences of nirS, nor and nosZ genes in the genomes are indicated with purple, turquoise and magenta bars, respectively. Bar height depicts the number of co-occurring gene copies (≤3) and the scale bar denotes nucleotide substitution rate (LG+Γ). Strain names are colored to indicate taxonomic affiliation according to the legend, and bootstrap values >70% are designated by circles.
(EPS)
Maximum likelihood phylogeny of 110 full-length nirS amino acid sequences, rooted at midpoint. Co-occurrences of nirK, nor and nosZ genes in the genomes are indicated with orange, turquoise and magenta bars, respectively. Bar height depicts the number of co-occurring gene copies (≤2) and the scale bar denotes nucleotide substitution rate (LG+Γ+F). Strain names are colored to indicate taxonomic affiliation according to the legend, and bootstrap values >70% are designated by circles.
(EPS)
Maximum likelihood phylogenies of 413 prokaryotic and 18 fungal (inserted graph) full-length nor amino acid sequences. Both phylogenies are rooted at midpoint. Co-occurrences of nirK, nirS and nosZ genes in the genomes are indicated with orange, purple and magenta bars, respectively. Bar height depicts the number of co-occurring gene copies (≤4) and the scale bar denotes nucleotide substitution rate (LG+Γ+F and WAG+Γ+F respectively). cnorB sequences are shaded in orange and qnorB sequences in green. Strain names are colored to indicate taxonomic affiliation according to the legend, and bootstrap values >70% are designated by circles.
(EPS)
Maximum likelihood phylogeny of 314 full-length nosZ amino acid sequences, rooted at midpoint. Co-occurrences of nirK, nirS and nor genes in the genomes are indicated with orange, purple and turquoise bars, respectively. Bar height depicts the number of co-occurring gene copies (≤4) and the scale bar denotes nucleotide substitution rate (LG+Γ). Clade I is shaded in orange, clade II in blue and the halophilic Archaea in turquoise. Strain names are colored to indicate taxonomic affiliation according to the legend, and bootstrap values >70% are designated by circles.
(EPS)
Taxon identification number and project name according to NCBI of 652 organisms harboring denitrification genes. Copy numbers of nirK , nirS , nor and nosZ are indicated.
(PDF)
Frequency table of gene co-occurrence patterns and taxonomic affiliation at class level. The nirK (K), nirS (S) and nosZ (Z) co-occurrence types and percentage of organisms within each class that also harbor a nor gene are indicated.
(PDF)
Frequency table of gene co-occurrence patterns and taxonomic affiliation at order level. The nirK (K), nirS (S) and nosZ (Z) co-occurrence types and percentage of organisms within each order that also harbor a nor gene are indicated.
(PDF)
Data Availability
The authors confirm that all data underlying the findings are fully available without restriction. All data are available from the NCBI database (www.ncbi.nlm.nih.gov/) as indicated by the taxon ID number and genome project name presented within the paper and its Supporting Information files.
Funding Statement
This work was supported by the Oscar and Lili Lamm foundation (www.stiftelsenlamm.a.se/) through a grant (contract 2010-11-13) to SH, strategic funding from the Faculty of Natural Resources and Agricultural Sciences at the Swedish University of Agricultural Sciences to SH, and a research grant (contract 2011-4421) from the Swedish Research Council (www.vr.se) to SH. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Rockström J, Steffen W, Noone K, Persson Å, Chapin FS III, et al. (2009) Planetary Boundaries: Exploring the Safe Operating Space for Humanity. Ecol Soc 4:32. [Google Scholar]
- 2.Sutton MA, Howard CM, Erisman JW, Billen G, Bleeker A, et al. (2011) The European Nitrogen Assessment: Sources, Effects and Policy Perspectives. Cambridge: Cambridge University Press. 612 p.
- 3.Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Stocker TF, D. Qin G-K, Plattner M, Tignor SK, Allen Jet al., editors. Cambridge: Cambridge University Press. 1535 p.
- 4. Ravishankara AR, Daniel JS, Portmann RW (2009) Nitrous oxide (N2O): the dominant ozone-depleting substance emitted in the 21st century. Science 326:123–125 10.1126/science.1176985 [DOI] [PubMed] [Google Scholar]
- 5. Firestone MK, Firestone RB, Tiedje JM (1980) Nitrous Oxide from Soil Denitrification: Factors Controlling its Biological Production. Am Assoc Adv Sci 208:749–751 10.1126/science.208.4445.749 [DOI] [PubMed] [Google Scholar]
- 6. Conrad R (1996) Soil microorganisms as controllers of atmospheric trace gases (H2, CO, CH4, OCS, N2O, and NO). Microbiol Rev 60:609–640. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Kool DM, Dolfing J, Wrage N, Van Groenigen JW (2011) Nitrifier denitrification as a distinct and significant source of nitrous oxide from soil. Soil Biol Biochem 43:174–178 10.1016/j.soilbio.2010.09.030 [DOI] [Google Scholar]
- 8. Zumft WG (1997) Cell biology and molecular basis of denitrification. Microbiol Mol Biol Rev 61:533–616. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Sanford RA, Cole JR, Tiedje JM (2002) Characterization and Description of Anaeromyxobacter dehalogenans gen. nov., sp. nov., an Aryl-Halorespiring Facultative Anaerobic Myxobacterium. Appl Environ Microbiol 68:893–900 10.1128/AEM.68.2.893 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Simon J, Einsle O, Kroneck PMH, Zumft WG (2004) The unprecedented nos gene cluster of Wolinella succinogenes encodes a novel respiratory electron transfer pathway to cytochrome c nitrous oxide reductase. FEBS Lett 569:7–12 10.1016/j.febslet.2004.05.060 [DOI] [PubMed] [Google Scholar]
- 11. Zumft WG, Kroneck PMH (2007) Respiratory transformation of nitrous oxide (N2O) to dinitrogen by Bacteria and Archaea. Adv Microb Physiol 52:107–227 10.1016/S0065-2911(06)52003-X [DOI] [PubMed] [Google Scholar]
- 12. Sanford R, Wagner DD, Wu Q, Chee-Sanford JC, Thomas SH, et al. (2012) Unexpected nondenitrifier nitrous oxide reductase gene diversity and abundance in soils. Proc Natl Acad Sci U S A 109:19709–19714 10.1073/pnas.1211238109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Jones CM, Spor A, Brennan FP, Breuil M, Bru D, et al. (2014) Recently identified microbial guild mediates soil N2O sink capacity. Nat Clim Change 4:801–805 10.1038/nclimate2301 [DOI] [Google Scholar]
- 14. Jones CM, Stres B, Rosenquist M, Hallin S (2008) Phylogenetic analysis of nitrite, nitric oxide, and nitrous oxide respiratory enzymes reveal a complex evolutionary history for denitrification. Mol Biol Evol 25:1955–1966 10.1093/molbev/msn146 [DOI] [PubMed] [Google Scholar]
- 15. Philippot L, Andert J, Jones CM, Bru D, Hallin S (2011) Importance of denitrifiers lacking the genes encoding the nitrous oxide reductase for N2O emissions from soil. Glob Change Biol 17:1497–1504 10.1111/j.1365-2486.2010.02334.x [DOI] [Google Scholar]
- 16.Shapleigh JP (2006).The Denitrifying Prokaryotes. In:The Prokaryotes. Dworkin M, Falkow S, Rosenberg E, Schleifer K-H, Stackebrandt Eeditors. New York: Springer New York. pp. 769–792. doi:10.1007/0-387-30742-7.
- 17. Santoro AE, Boehm AB, Francis CA (2006) Denitrifier community composition along a nitrate and salinity gradient in a coastal aquifer. Appl Environ Microbiol 72:2102–2109 10.1128/AEM.72.3.2102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Desnues C, Michotey VD, Wieland A, Zhizang C, Fourçans A, et al. (2007) Seasonal and diel distributions of denitrifying and bacterial communities in a hypersaline microbial mat (Camargue, France). Water Res 41:3407–3419 10.1016/j.watres.2007.04.018 [DOI] [PubMed] [Google Scholar]
- 19. Smith JM, Ogram A (2008) Genetic and functional variation in denitrifier populations along a short-term restoration chronosequence. Appl Environ Microbiol 74:5615–5620 10.1128/AEM.00349-08 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Enwall K, Throbäck IN, Stenberg M, Söderström M, Hallin S (2010) Soil resources influence spatial patterns of denitrifying communities at scales compatible with land management. Appl Environ Microbiol 76:2243–2250 10.1128/AEM.02197-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Yuan Q, Liu P, Lu Y (2012) Differential responses of nirK- and nirS-carrying bacteria to denitrifying conditions in the anoxic rice field soil. Environ Microbiol Rep 4:113–122 10.1111/j.1758-2229.2011.00311.x [DOI] [PubMed] [Google Scholar]
- 22. Jones CM, Hallin S (2010) Ecological and evolutionary factors underlying global and local assembly of denitrifier communities. ISME J 4:633–641 10.1038/ismej.2009.152 [DOI] [PubMed] [Google Scholar]
- 23. Simon J, Klotz MG (2013) Diversity and evolution of bioenergetic systems involved in microbial nitrogen compound transformations. Biochim Biophys Acta 1827:114–135 10.1016/j.bbabio.2012.07.005 [DOI] [PubMed] [Google Scholar]
- 24. Stein LY, Klotz MG (2011) Nitrifying and denitrifying pathways of methanotrophic bacteria. Biochem Soc Trans 39:1826–1831 10.1042/BST20110712 [DOI] [PubMed] [Google Scholar]
- 25. Zumft WG (2005) Nitric oxide reductases of prokaryotes with emphasis on the respiratory, heme-copper oxidase type. J Inorg Biochem 99:194–215 10.1016/j.jinorgbio.2004.09.024 [DOI] [PubMed] [Google Scholar]
- 26. Philippot L (2005) Denitrification in pathogenic bacteria: for better or worst? Trends Microbiol 13:191–192 10.1016/j.tim.2005.03.001 [DOI] [PubMed] [Google Scholar]
- 27. Ettwig KF, Butler MK, Le Paslier D, Pelletier E, Mangenot S, et al. (2010) Nitrite-driven anaerobic methane oxidation by oxygenic bacteria. Nature 464:543–548 10.1038/nature08883 [DOI] [PubMed] [Google Scholar]
- 28. Daiber A, Shoun H, Ullrich V (2005) Nitric oxide reductase (P450nor) from Fusarium oxysporum. J Inorg Biochem 99:185–193 10.1016/j.jinorgbio.2004.09.018 [DOI] [PubMed] [Google Scholar]
- 29. Pagani I, Liolios K, Jansson J, Chen I-M a, Smirnova T, et al. (2011) The Genomes OnLine Database (GOLD) v.4: status of genomic and metagenomic projects and their associated metadata. Nucleic Acids Res 40:D571–D579 10.1093/nar/gkr1100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Altschul SF, Gish W (1990) Basic Local Alignment Search Tool. J Mol Biol 215:403–410 10.1016/S0022-2836(05)80360-2 [DOI] [PubMed] [Google Scholar]
- 31. Liu K, Warnow TJ, Holder MT, Nelesen SM, Yu J, et al. (2012) SATe-II: very fast and accurate simultaneous estimation of multiple sequence alignments and phylogenetic trees. Syst Biol 61:90–106 10.1093/sysbio/syr095 [DOI] [PubMed] [Google Scholar]
- 32. Katoh K, Misawa K, Kuma K, Miyata T (2002) MAFFT: a novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res 30:3059–3066 10.1093/nar/gkf436 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Stamatakis A (2006) RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 22:2688–2690 10.1093/bioinformatics/btl446 [DOI] [PubMed] [Google Scholar]
- 34. Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, et al. (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 25:3389–3402 10.1093/nar/25.17.3389 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35. Kizawa H, Tomura D, Oda M, Fukamizu a, Hoshino T, et al. (1991) Nucleotide sequence of the unique nitrate/nitrite-inducible cytochrome P-450 cDNA from Fusarium oxysporum. J Biol Chem 266:10632–10637. [PubMed] [Google Scholar]
- 36. Nawrocki EP, Kolbe DL, Eddy SR (2009) Infernal 1.0: inference of RNA alignments. Bioinformatics 25:1335–1337 10.1093/bioinformatics/btp157 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Quast C, Pruesse E, Yilmaz P, Gerken J, Schweer T, et al. (2013) The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res 41:D590–D596 10.1093/nar/gks1219 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38. Edgar RC, Drive RM, Valley M (2004) MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res 32:1792–1797 10.1093/nar/gkh340 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39. Abascal F, Zardoya R, Posada D (2005) ProtTest: selection of best-fit models of protein evolution. Bioinformatics 21:2104–2105 10.1093/bioinformatics/bti263 [DOI] [PubMed] [Google Scholar]
- 40. Le SQ, Gascuel O (2008) An improved general amino acid replacement matrix. Mol Biol Evol 25:1307–1320 10.1093/molbev/msn067 [DOI] [PubMed] [Google Scholar]
- 41. Pruesse E, Peplies J, Glöckner FO (2012) SINA: accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 28:1823–1829 10.1093/bioinformatics/bts252 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Letunic I, Bork P (2007) Interactive Tree Of Life (iTOL): an online tool for phylogenetic tree display and annotation. Bioinformatics 23:127–128 10.1093/bioinformatics/btl529 [DOI] [PubMed] [Google Scholar]
- 43. Fritz SA, Purvis A (2010) Selectivity in mammalian extinction risk and threat types: a new measure of phylogenetic signal strength in binary traits. Conserv Biol 24:1042–1051 10.1111/j.1523-1739.2010.01455.x [DOI] [PubMed] [Google Scholar]
- 44. Martiny AC, Treseder K, Pusch G (2013) Phylogenetic conservatism of functional traits in microorganisms. ISME J 7:830–838 10.1038/ismej.2012.160 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45. Jones CM, Graf DR, Bru D, Philippot L, Hallin S (2013) The unaccounted yet abundant nitrous oxide-reducing microbial community: a potential nitrous oxide sink. ISME J 7:417–426 10.1038/ismej.2012.125 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46. Orellana LH, Rodriguez-R LM, Higgins S, Chee-Sanford JC, Sanford R a, et al. (2014) Detecting nitrous oxide reductase (NosZ) genes in soil metagenomes: method development and implications for the nitrogen cycle. mBio 5:e01193–14 10.1128/mBio.01193-14 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47. Bartossek R, Nicol GW, Lanzen A, Klenk H-P, Schleper C (2010) Homologues of nitrite reductases in ammonia-oxidizing archaea: diversity and genomic context. Environ Microbiol 12:1075–1088 10.1111/j.1462-2920.2010.02153.x [DOI] [PubMed] [Google Scholar]
- 48. Campbell M, Nyerges G, Kozlowski J, Poret-Peterson AT, Stein LY, et al. (2011) Model of the molecular basis for hydroxylamine oxidation and nitrous oxide production in methanotrophic bacteria. FEMS Microbiol Lett 322:82–89 10.1111/j.1574-6968.2011.02340.x [DOI] [PubMed] [Google Scholar]
- 49. Kraft B, Tegetmeyer HE, Meier D, Geelhoed JS, Strous M (2014) Rapid succession of uncultured marine bacterial and archaeal populations in a denitrifying continuous culture. Environ Microbiol 16:3275–3286 10.1111/1462-2920.12552 [DOI] [PubMed] [Google Scholar]
- 50. Etchebehere C, Tiedje J (2005) Presence of Two Different Active nirS Nitrite Reductase Genes in a Denitrifying Thauera sp. from a High-Nitrate-Removal-Rate Reactor. Appl Environ Microbiol 71:5642 10.1128/AEM.71.9.5642 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51. Vial L, Lavire C, Mavingui P, Blaha D, Haurat J, et al. (2006) Phase variation and genomic architecture changes in Azospirillum. J Bacteriol 188:5364–5373 10.1128/JB.00521-06 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52. Barth KR, Isabella VM, Clark VL (2009) Biochemical and genomic analysis of the denitrification pathway within the genus Neisseria. Microbiology 155:4093–4103 10.1099/mic.0.032961-0 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53. Körner H, Sofia HJ, Zumft WG (2003) Phylogeny of the bacterial superfamily of Crp-Fnr transcription regulators: exploiting the metabolic spectrum by controlling alternative gene programs. FEMS Microbiol Rev 27:559–592 10.1016/S0168-6445(03)00066-4 [DOI] [PubMed] [Google Scholar]
- 54. Spiro S (2012) Nitrous oxide production and consumption: regulation of gene expression by gas-sensitive transcription factors. Philos Trans R Soc Lond B Biol Sci 367:1213–1225 10.1098/rstb.2011.0309 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Bañeras L, Ruiz-Rueda O, López-Flores R, Quintana XD, Hallin S (2012) The role of plant type and salinity in the selection for the denitrifying community structure in the rhizosphere of wetland vegetation. Int Microbiol 15:89–99 10.2436/20.1501.01.162 [DOI] [PubMed] [Google Scholar]
- 56. Clark IM, Buchkina N, Jhurreea D, Goulding KWT, Hirsch PR (2012) Impacts of nitrogen application rates on the activity and diversity of denitrifying bacteria in the Broadbalk Wheat Experiment. Philos Trans R Soc Lond B Biol Sci 367:1235–1244 10.1098/rstb.2011.0314 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57. Cuhel J, Simek M, Laughlin RJ, Bru D, Chèneby D, et al. (2010) Insights into the effect of soil pH on N2O and N2 emissions and denitrifier community size and activity. Appl Environ Microbiol 76:1870–1878 10.1128/AEM.02484-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Payne WJ, Grant M a, Shapleigh J, Hoffman P (1982) Nitrogen oxide reduction in Wolinella succinogenes and Campylobacter species. J Bacteriol 152:915–918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59. Liu X, Gao C, Zhang A, Jin P, Wang L, et al. (2008) The nos gene cluster from gram-positive bacterium Geobacillus thermodenitrificans NG80-2 and functional characterization of the recombinant NosZ. FEMS Microbiol Lett 289:46–52 10.1111/j.1574-6968.2008.01362.x [DOI] [PubMed] [Google Scholar]
- 60. Kern M, Simon J (2009) Electron transport chains and bioenergetics of respiratory nitrogen metabolism in Wolinella succinogenes and other Epsilonproteobacteria. Biochim Biophys Acta 1787:646–656 10.1016/j.bbabio.2008.12.010 [DOI] [PubMed] [Google Scholar]
- 61. Sullivan MJ, Gates AJ, Appia-Ayme C, Rowley G, Richardson DJ (2013) Copper control of bacterial nitrous oxide emission and its impact on vitamin B12-dependent metabolism. Proc Natl Acad Sci U S A 110:19926–19931 10.1073/pnas.1314529110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62. Chapuis-Lardy L, Wrage N, Metay A, Chotte J-L, Bernoux M (2007) Soils, a sink for N2O? A review. Glob Chang Biol 13:1–17 10.1111/j.1365-2486.2006.01280.x [DOI] [Google Scholar]
- 63. Wu D, Dong W, Oenema O, Wang Y, Trebs I, et al. (2013) N2O consumption by low-nitrogen soil and its regulation by water and oxygen. Soil Biol Biochem 60:165–172 10.1016/j.soilbio.2013.01.028 [DOI] [Google Scholar]
- 64. Philippot L (2002) Denitrifying genes in bacterial and Archaeal genomes. Biochim Biophys Acta 1577:355–376 10.1016/S0167-4781(02)00420-7 [DOI] [PubMed] [Google Scholar]
- 65. Jones CM, Welsh A, Throbäck IN, Dörsch P, Bakken LR, et al. (2011) Phenotypic and genotypic heterogeneity among closely related soil-borne N2 - and N2O-producing Bacillus isolates harboring the nosZ gene. FEMS Microbiol Ecol 76:541–552 10.1111/j.1574-6941.2011.01071.x [DOI] [PubMed] [Google Scholar]
- 66. Liu B, Mao Y, Bergaust L, Bakken LR, Frostegård Å (2013) Strains in the genus Thauera exhibit remarkably different denitrification regulatory phenotypes. Environ Microbiol 15:2816–2828 10.1111/1462-2920.12142 [DOI] [PubMed] [Google Scholar]
- 67. Philippot L, Cuhel J, Saby NP a, Chèneby D, Chronáková A, et al. (2009) Mapping field-scale spatial patterns of size and activity of the denitrifier community. Environ Microbiol 11:1518–1526 10.1111/j.1462-2920.2009.01879.x [DOI] [PubMed] [Google Scholar]
- 68. Philippot L, Bru D, Saby NP a, Cuhel J, Arrouays D, et al. (2009) Spatial patterns of bacterial taxa in nature reflect ecological traits of deep branches of the 16S rRNA bacterial tree. Environ Microbiol 11:3096–3104 10.1111/j.1462-2920.2009.02014.x [DOI] [PubMed] [Google Scholar]
- 69. Baek S, Rajashekara G, Splitter GA, Shapleigh JP (2004) Denitrification genes regulate Brucella virulence in mice. J Bacteriol 186:6025–6031 10.1128/JB.186.18.6025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70. Roop RM, Gaines JM, Anderson ES, Caswell CC, Martin DW (2009) Survival of the fittest: how Brucella strains adapt to their intracellular niche in the host. Med Microbiol Immunol 198:221–238 10.1007/s00430-009-0123-8 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Van Alst NE, Wellington M, Clark VL, Haidaris CG, Iglewski BH (2009) Nitrite reductase NirS is required for type III secretion system expression and virulence in the human monocyte cell line THP-1 by Pseudomonas aeruginosa. Infect Immun 77:4446–4454 10.1128/IAI.00822-09 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Toft C, Andersson SGE (2010) Evolutionary microbial genomics: insights into bacterial host adaptation. Nat Rev Genet 11:465–475 10.1038/nrg2798 [DOI] [PubMed] [Google Scholar]
- 73. Philippot L, Piutti S, Martin-laurent F, Hallet S, Germon JC (2002) Molecular Analysis of the nitrate-reducing community from unplanted and maize-planted soils. Appl Env Microbiol 68:6121–6128 10.1128/AEM.68.12.6121 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74. Petersen DG, Blazewicz SJ, Firestone M, Herman DJ, Turetsky M, et al. (2012) Abundance of microbial genes associated with nitrogen cycling as indices of biogeochemical process rates across a vegetation gradient in Alaska. Environ Microbiol 14:993–1008 10.1111/j.1462-2920.2011.02679.x [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Maximum likelihood phylogeny of full-length 16S and 18S rRNA sequences from 652 organisms harboring denitrification genes. The inner colored ring represents taxonomic affiliation as indicated by the legend. The four outer bar-chart rings show the presence of nirK (orange), nirS (purple), nor (turquoise) and nosZ (magenta). Bar height represents the number of copies (≤4). Bootstrap values >70% are indicated by grey circles, and the scale bar denotes nucleotide substitution rate (GTR+Γ).
(EPS)
Maximum likelihood phylogeny of 458 full-length nirK amino acid sequences, rooted at midpoint. Co-occurrences of nirS, nor and nosZ genes in the genomes are indicated with purple, turquoise and magenta bars, respectively. Bar height depicts the number of co-occurring gene copies (≤3) and the scale bar denotes nucleotide substitution rate (LG+Γ). Strain names are colored to indicate taxonomic affiliation according to the legend, and bootstrap values >70% are designated by circles.
(EPS)
Maximum likelihood phylogeny of 110 full-length nirS amino acid sequences, rooted at midpoint. Co-occurrences of nirK, nor and nosZ genes in the genomes are indicated with orange, turquoise and magenta bars, respectively. Bar height depicts the number of co-occurring gene copies (≤2) and the scale bar denotes nucleotide substitution rate (LG+Γ+F). Strain names are colored to indicate taxonomic affiliation according to the legend, and bootstrap values >70% are designated by circles.
(EPS)
Maximum likelihood phylogenies of 413 prokaryotic and 18 fungal (inserted graph) full-length nor amino acid sequences. Both phylogenies are rooted at midpoint. Co-occurrences of nirK, nirS and nosZ genes in the genomes are indicated with orange, purple and magenta bars, respectively. Bar height depicts the number of co-occurring gene copies (≤4) and the scale bar denotes nucleotide substitution rate (LG+Γ+F and WAG+Γ+F respectively). cnorB sequences are shaded in orange and qnorB sequences in green. Strain names are colored to indicate taxonomic affiliation according to the legend, and bootstrap values >70% are designated by circles.
(EPS)
Maximum likelihood phylogeny of 314 full-length nosZ amino acid sequences, rooted at midpoint. Co-occurrences of nirK, nirS and nor genes in the genomes are indicated with orange, purple and turquoise bars, respectively. Bar height depicts the number of co-occurring gene copies (≤4) and the scale bar denotes nucleotide substitution rate (LG+Γ). Clade I is shaded in orange, clade II in blue and the halophilic Archaea in turquoise. Strain names are colored to indicate taxonomic affiliation according to the legend, and bootstrap values >70% are designated by circles.
(EPS)
Taxon identification number and project name according to NCBI of 652 organisms harboring denitrification genes. Copy numbers of nirK , nirS , nor and nosZ are indicated.
(PDF)
Frequency table of gene co-occurrence patterns and taxonomic affiliation at class level. The nirK (K), nirS (S) and nosZ (Z) co-occurrence types and percentage of organisms within each class that also harbor a nor gene are indicated.
(PDF)
Frequency table of gene co-occurrence patterns and taxonomic affiliation at order level. The nirK (K), nirS (S) and nosZ (Z) co-occurrence types and percentage of organisms within each order that also harbor a nor gene are indicated.
(PDF)
Data Availability Statement
The authors confirm that all data underlying the findings are fully available without restriction. All data are available from the NCBI database (www.ncbi.nlm.nih.gov/) as indicated by the taxon ID number and genome project name presented within the paper and its Supporting Information files.