

Abstract
Wadi Arar in the Northern border region of Saudi Arabia is one of the most important Wadis of the Kingdom. The present study provides an analysis of vegetation types, life forms, as well as floristic categories and species distribution. A total of 196 species representing 31 families of vascular plants were recorded. Compositae, Gramineae and Leguminosae were the most common families. Therophytes and chamaephytes are the most frequent life forms, indicating typical desert spectrum vegetation. The distribution of these species in the different sectors of the Wadi as well as the phytochoria for the recorded species is provided. Ninety-one species (46.5%) are typical bi-regional. Furthermore, about 105 species (53.5%) are mono- or pluriregional taxa. The highest number of species (136 or 69.5%) was recorded for annual plants, while the lowest number of species (60% or 30.5%) was recorded for perennial, short perennial or annual to biennial species.
Keywords: Kingdom of Saudi Arabia, Northern border region, Floristic survey, Flora, Wadi Arar, Floristic diversity
1. Introduction
The Kingdom of Saudi Arabia is a huge arid land with an area of about 2,250,000 km2 covering the major part of the Arabian Peninsula, characterized by different ecosystems and diversity of plant species (Abdel Khalik et al., 2013).
The topography of Saudi Arabia, as well as that of Arabian Peninsula, is an ancient massif in which geologic structure developed concurrently with the Alps (Country Profile: Saudi Arabia, 2006). The climate in Saudi Arabia differs greatly between the coast and the interior. High humidity coupled with more moderate temperatures is prevalent along the coast, whereas aridity and extreme temperatures characterize the interior (Country Profile: Saudi Arabia, 2006).
Saudi Arabia has not permanent rivers or lakes; however “Wadis” are frequent along the country. The word “Wadi” in this article will be used to designate a non permanent river, whose runoff is dependent on rain. It frequently happened, but for decades Wadis have been completely inactive. In the dry lands, Wadis represent one of the most prominent desert landforms, which exhibit physiographic irregularities that lead to parallel variations in plant species distribution (Kassas and Girgis, 1964). Wadi “Arar” is considered one of the most important Wadis in the northern region.
The flora of Saudi Arabia is one of the richest biodiversities in the Arabian Peninsula and comprises very important genetic resources of crops and medicinal plants (Atiqur Rahman et al., 2004). The components of this flora are admixture of Asia, Africa and Mediterranean regions’ plants. According to Collenete (1999), this flora comprises 2250 species belonging to 835 genera and about 142 families. 147 species are classified as “endemic”, 721 species as “endangered” and about 22 species are believed as completely extinct (Collenete, 1999).
The number of wild species occurring in a square kilometer area of Saudi Arabia is less significant, especially in areas such as central, eastern and northern regions. Areas along the northwestern and southwestern regions are densely vegetated and contain the highest number of species. Approximately 70% of the country’s floristic species are reported in these areas. Endemism in Saudi Arabia is relatively insignificant in comparison with some of the neighboring countries such as Yemen and Oman.
Saudi Arabia contains 97 trees, 564 shrubs and about 1620 herbs, which cover, respectively, 4.25%, 24.73% and 71.02% of higher plant diversity of the country (http://www.plantdiversityofsaudiarabia.info/index.htm).
Wild plants of Saudi Arabia were studied by many authors (Chaudhary, 1999–2001; Chaudhary and Al-Jowaid, 1999; Collenette, 1999; Migahid, 1996; Miller and Cope, 1996) as general floral studies. Furthermore, many authors have studied more limited areas in the course of floral and taxonomical revision of some genera (Aleem, 1979; Batanouny and Ismail, 1985; Fayed and Zayed, 1989; El Karemy and Zayed, 1992; Hajar et al., 1998; Hegazy et al., 1998; Al Wadie, 2002; Al-Turki and Al-Qlayan, 2003; Atiqur Rahman et al., 2004; Alfarhan et al., 2005; Mosallam, 2007; Alshammari and Sharawy, 2010; El-Ghanem et al., 2010; Al-Sodany et al., 2011; Alatar et al., 2012; Alsherif et al., 2012; Farrag, 2012; Gomaa, 2012; Ihsanullah, 2012; Waly et al., 2012 and Abdel Khalik et al., 2013).
The distribution of life form is closely related to topography and landform (Kassas and Girgis, 1965; Zohary, 1973; Orshan, 1986; Fakhireh et al., 2012). In Wadis, the composition of life forms expresses a typical desert flora, the majority of species being therophytes and chamaephytes. Vegetation of Wadis in general is not constant; it varies from year to year, depending upon the moisture level (Siddiqui and Al-Harbi, 1995). The establishment, growth, regeneration and distribution of the plant communities in the Wadis are controlled by many factors, such as geographical position, physiographic features and human impacts (Shaltout and El-Sheikh, 2003; Kürschner and Neef, 2011; Alatar et al., 2012; Korkmaz and Ozcelik, 2013).
The current study includes survey and identification of the wild plants growing in the Wadi Arar and the surrounding sites in the Northern border region of Saudi Arabia. Moreover, the vegetation types, life forms and floristic categories of the collected species were taken into consideration. For future reference, voucher herbarium specimens of different plant samples were pressed and deposited in the herbarium of College of Sciences in Arar (Northern Border University).
2. The study area
The area of the present study is located in the Northern region of Saudi Arabia (30° 55′ 13″ N, 41° 0′ 3″ E). Wadi Arar (Wadi of the Anizah tribe) is one of the main Wadis which existed in the northeast of Saudi Arabia (Fig. 1). It covers an area of about 9500 km square and extends next to Arar city. According to the Worldwide Bioclimatic Classification System (http://www.globalbioclimatics.org), this region falls within the Mediterranean desertic continental climate. This area is dry zone, characterized by hot, arid, desert climate and an average annual temperature of 21.5 °C (Fig. 2). Precipitation is scanty, with an average annual of 20.2 mm, which usually falls during the winter months, the extreme rainfall occurring in January and May with more than 90 mm (http://www.globalbioclimatics.org).
Figure 1.
Location map of Saudi Arabia showing the Northern border region.
Figure 2.
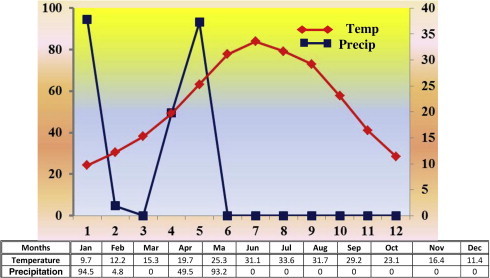
Climate diagram for Arar, Northern border region, Saudi Arabia.
3. Materials and methods
This study was performed in Wadi Arar from November 2011 to April 2013 during active plant growth period, when most species were expected to be present. The wild vegetations were sampled in 25 localities including the different Wadi tributaries; in each locality, the present species were recorded. .
The collected plant specimens were identified and named according to Collenette (1999), Cope (1985), Mighaid (1996), and Chaudhary (1999–2001). They are deposited in the Herbarium of Biology Department, College of Sciences, Northern Border University.
Life forms of species were determined depending upon the location of the regenerative buds and the shed parts during the unfavorable season (Raunkier, 1934). The floristic categories of the investigated species were made to assign the registered species to world geographical groups according to Wickens (1978) and Zohary (1973).
4. Results
Table 1 shows the plant species recorded in different surveyed sectors of Wadi Arar. The list includes 196 species belonging to 31 families of phanerogams. Their generic representation is quite variable. However, one family is represented by more than 29 genera (Compositae), another by 19 genera (Gramineae); the third, most represented family carries about 14 genera (Cruciferae). Five families are represented by four to eight genera (Caryophyllaceae and Leguminosae with 8 genera; Umbelliferae with 7 genera; and Zygophyllaceae with 4 genera). Furthermore, eight families showed less number of genera (2 to 3 genera) and 15 families are represented by a single genus (Table 1).
Table 1.
A list of the species recorded in the study area with their families, vegetation type, life form and floristic categories.
| Family | Species | Vegetation type | Life form | Floristic categories |
|---|---|---|---|---|
| Aizoaceae | Aizoon canariense L. | Perennial | H | SA-SI + S-Z |
| Amaranthaceae | Aerva javonica (Burm. f.) Juss. ex Scult. | Perennial | Ch | SA-SI + S-Z |
| Amaranthus graecizans L. | Annual | Th | PAL | |
| Amaranthus viridis L. | Annual | Th | COSM | |
| Asclepiadaceae | Calotropis procera (Aiton) W. T. Aiton | Perennial | Ph | SA + S-Z |
| Perennialgularia tomentosa L. | Perennial | Ch | SA-SI + S-Z | |
| Boraginaceae | Arnebia decumbens (Vent.) Coss. & Kralik var. decumbens | Annual | Th | SA-SI + S-Z |
| Arnebia decumbens (Vent.) Coss. & Kralik var. macrocalyx Coss. & Kralik | Annual | Th | SA-SI + S-Z | |
| Arnebia linearifolia A. DC. | Annual | Th | SA-SI + S-Z | |
| Heliotropium bacciferum Forssk. subsp. bacciferum var. bacciferum | Perennial | Ch | SA-SI + S-Z | |
| Capparaceae | Cleome amblyocarpa Barratte & Murb. | Annual | Th | SA-SI + S-Z |
| Caryophyllaceae | Gypsophila capillaries (Forssk.) C. Chr. | Annual | Th | SA |
| Gypsophila linearifolia (Fisch. & C. A. Mey.) Boiss. | Annual | Th | IR-TR | |
| Gypsophila viscosa Murray | Annual | Th | IR-TR | |
| Herniaria hemistemon J. Gay | Perennial | Th | ME + SA-SI | |
| Herniaria hirsuta L. | Annual | Th | ME + SA-SI | |
| Paronychia arabica (L.) DC. | Annual | Th | ME + SA-SI + IR-TR | |
| Polycarpea robbairea (Kuntze) Greuter & Burdet | Perennial | H | SA-SI + S-Z | |
| Polycarpon tetraphyllum (L.) L. | Annual | Th | ME + SA-SI + IR-TR | |
| Pteranthus dichotomus Forssk. | Annual | Th | ME + SA + IR-TR | |
| Silene arabica Boiss. | Annual | Th | SA-SI | |
| Sperennialgularia diandra (Guss.) Boiss. | Annual | Th | ME + SA + IR-TR | |
| Sperennialgularia marina (L.) Bessler. | Annual | Th | ME + IR-TR | |
| Chenopodiaceae | Atriplex leucoclada Boiss. var. turcomania (Moq.) Zohary. | Perennial | Ch | ME + SA-SI |
| Bassia eriophora (Schrad.) Asch. | Annual | Th | SA + IR-TR | |
| Chenopodium ambrosioides L. | Biennial | Th | COSM | |
| Chenopodium murale L. | Annual | Th | COSM | |
| Haloxylon salicornicum (Moq.) Bunge ex Boiss. | Perennial | Ch | S-Z | |
| Salsola villosa Schult. | Perennial | Ch | ME + SA-SI + IR-TR | |
| Salsola volkensii Schweinf. & Asch. | Perennial | Ch | COSM | |
| Suaeda pruinosa Lange | Perennial | Ch | ME + SA | |
| Cistaceae | Helianthemum lippi (L.) Dom. | Perennial | Ch | SA-SI + S-Z |
| Helianthemum vesicarium Boiss. | Perennial | Ch | SA | |
| Cleomaceae | Cleome amblyocarpa Barrtte & Murb. | Annual | Th | SA-SI + S-Z |
| Compositae | Anthemis arvensis L. | Annual | Th | SA |
| Anthemis cotula L. | Annual | Th | SA | |
| Anthemis deserti (Boiss.) Eig. | Annual | Th | ME + SA | |
| Anthemis melampodina Delile. | Annual | Th | ME + SA | |
| Anthemis pseudocotula Boiss. | Annual | Th | ME + SA | |
| Anthemis scrobicularis Yavin. | Annual | Th | ME + SA-SI + IR-TR | |
| Artemisia judaica L. | Perennial | Ch | SA | |
| Artemisia monosperennialma Delile. | Perennial | Ch | ME + SA | |
| Artemisia scoparia Waldst. & Kit. | Perennial | Ch | IR-TR + EU-SI | |
| Atractylis carduus (Forssk.) C. Christ. var. angustifolia Täckh. & Boulos. | Perennial | Th | ME | |
| Calendula arvensis L. | Annual | Th | ME + SA-SI + IR-TR | |
| Calendula tripterocarpa Rupr. | Annual | Th | PAN | |
| Carthamus nitidus Boiss. | Annual | Th | SA + IR-TR | |
| Carthamus oxycantha M. Bieb. | Annual | Th | ME + IR-TR | |
| Centaurea pseudosinaica Czerep. | Annual | Th | IR-TR | |
| Centaurea sinaica DC. | Annual | Th | IR-TR + EU-SI | |
| Chamaemelum mixtum (L.) All. | Annual | Th | ME + SA | |
| Cotula anthemoides L. | Annual | Th | SA | |
| Cotula cinerea Delile. | Annual | Th | SA- SI | |
| Crepis nigricans Viv. | Annual | Th | SA | |
| Filago desertorum Pomel | Annual | Th | SA-SI + IR-TR | |
| Garhadiolus angulosus Jaub. & Spach. | Annual | Th | ME + IR-TR | |
| Glebionis coronaria (L.) Tzvelv. (= Chrysanthemum coronarium L.) | Annual | H | ME + SA-SI | |
| Gymnarrhena micrantha Desf. | Annual | Th | ME + SA-SI | |
| Helichrysum conglobatum (Viv.) Steud. | Perennial | Th | ME | |
| Homognaphalium pulvinatum (Delile.) Fayed & Zareh. | Annual | Th | SA–SI + IR–TR | |
| Ifloga spicata (Forssk.) Sch. Bip. subsp. albescens Chrtek. | Annual | Th | ME + SA-SI | |
| Koelpinia linearis Pall. | Annual | Th | ME + SA + IR-TR | |
| Lactuca serriola L. | Annual or biennial | Th | IR-TR | |
| Lasiopogon muscoides (Desf.) DC. | Annual | Th | ME + SA-SI | |
| Launaea capitata (Spreng.) Dandy | Annual | Th | ME + SA–SI | |
| Launaea fragilis (Asso) Pau subsp. fragilis (= L. tenuiloba (Boiss.) Kuntze) | Annual | Th | ME + SA–SI | |
| Launaea mucronata (Forssk.) Muschl. subsp. mucronata | Short-Perennial | Th | SA–SI + IR–TR | |
| Launaea nudicaulis (L.) Hook. fil. | Perennial | Th | SA–SI + IR–TR + S–Z | |
| Leontodon hispidulus (Delile.) Boiss. | Annual | Th | ME + SA-SI | |
| Leontodon laciniatus (Bertol.) Widder. | Annual | Th | ME + SA-SI | |
| Matricaria aurea (Loefl.) Sch. Bip. | Annual | Th | SA-SI | |
| Nauplius graveolens (Forssk.) Wiklund. (= Asteriscus graveolens (Forssk.) Less.) | Perennial | Ch | ME + SA-SI | |
| Picris altissima Delile. | Annual | Th | ME + SA | |
| Picris asplenioides L. | Annual | Th | ME + IR-TR | |
| Picris cyanocarpa Boiss. | Annual | Th | SA | |
| Pseudognaphalium luteoalbum (L.) Hilliard & B. L. Burtt. | Annual | Th | ME + SA + IR-TR | |
| Reichardia tingitana (L.) Roth | Perennial | Cr | ME + SA-SI + IR-TR | |
| Senecio belbeysius Delile. | Annual | Th | ME + IR-TR + EU-SI | |
| Senecio glaucus L. subsp. coronopifolius (Maire) C. Alexander (= S. desfontainei Druce) | Annual | Ch | SA-SI + IR-TR | |
| Sonchus oleraceus L. | Annual | Th | COSM | |
| Sonchus tenerrimus L. | Annual | Th | COSM | |
| Urosperennialmum picroides (L.) F.W. Schmidt | Annual | Th | ME + IR-TR | |
| Convolvulaceae | Convolvulus arvensis L. | Perennial | H | PAL |
| Cuscuta campestris Yunck. | Perennial | P | SA-SI + S-Z | |
| Cruciferae | Arabidopsis kneuckeri (Bornm.) O. E. Schulz | Annual | Th | SA-SI |
| Carrichtera annualua (L.) DC. | Annual | Th | ME + SA-SI | |
| Coronopus didymus (L.) Sm. | Annual or biennial | Th | SA | |
| Diplotaxis acris (Forssk.) Boiss. | Annual | Th | ME + IR-TR | |
| Enarthrocarpus strangulatus Boiss. | Annual | Th | SA-SI | |
| Eruca sativa Mill. | Annual | Th | ME + SA + IR-TR + EU-SI | |
| Erucaria crassifolia (Forssk.) Delile | Annual | Th | ME + SA-SI | |
| Erucaria hispanica (L.) Druce | Annual | Th | ME + SA-SI | |
| Farsetia aegyptia Turra | Perennial | Ch | SA-SI + S-Z | |
| Farsetia stylosa R. Br. | Annual or short-Perennial | Th | SA-SI | |
| Horwoodia dicksoniae (Viv.) Turril | Annual | Th | SA-SI | |
| Matthiola arabica Boiss. | Annual | Th | SA | |
| Matthiola longipetala (Vent.) DC. | Annual | Th | ME + IR-TR | |
| Savignya parviflora (Delile) Web. | Annual | Th | SA-SI + IR-TR | |
| Schimperenniala arabica Hochst. & Steud. Ex Endl | Annual | Th | SA-SI | |
| Sisymbrium irio L. | Annual | Th | ME + SA-SI + IR-TR + EU-SI | |
| Zilla spinosa (L.) Prantl subsp. spinosa | Perennial | Ch | ME + SA-SI + IR-TR + EU-SI | |
| Cucurbitaceae | Citrullus colocynthis (L.) Schrad. | Perennial | H | ME + SA-SI + IR-TR + S-Z |
| Euphorbiaceae | Euphorbia peplus L. | Annual | Th | COSM |
| Euphorbia prostrata Aiton, Hort. | Annual | Th | ME + SA-SI + IR-TR | |
| Euphorbia serpens Kunth. | Annual | Th | SA-SI + IR-TR | |
| Fumariaceae | Hypecoum pendulum L. | Annual | Th | ME + IR-TR |
| Geraniaceae | Erodium crassifolium L’ Hér. (= E. hirtum (Forssk.) Willd.) | Perennial | Th | SA-SI |
| Gramineae | Agropyron cristatum (L.) Gaertn. | Perennial | H | IR-TR |
| Bromus scoparius L. | Annual | Th | ME + SA-SI + IR-TR | |
| Bromus tectorum L. | Annual | Th | COSM | |
| Cutandia memphitica (Spreng.) Benth. | Annual | Th | ME + IR-TR | |
| Cynodon dactylon (L.) Perennials. | Perennial | G | PAN | |
| Enneapogon lophotrichus Chiov. Ex H. Scholz & P. Köng | Annual | Th | IR-TR | |
| Eragrostis aegyptiaca (Willd) Delile. | Annual | Th | S-Z | |
| Eremopyrum bonaepartis (Spreng.) Nevski | Annual | Th | IR-TR | |
| Eremopyrum confusum Melderis | Annual | Th | IR-TR | |
| Hordeum murinum L. subsp. glaucum (Steud.) Tzvelv | Annual | Th | ME + IR-TR | |
| Hordeum murinum L. subsp. leporinum (Link.) Arcang. | Annual | Th | ME + IR-TR | |
| Lamarckia aurea (L.) Moench | Annual | Th | ME + IR-TR + S-Z | |
| Lolium multiflorum Lam. | Annual | Th | ME + IR-TR + EU-SI | |
| Lolium perennialenne L. | Annual | Th | COSM | |
| Lolium rigidum Gaudin | Annual | Th | ME + IR –TR | |
| Parapholis incurva (L.) C. E. | Annual | Th | ME + IR-TR + EU-SI | |
| Phalaris paradoxa L. | Annual | Th | ME + IR-TR | |
| Poa annualua L. | Annual or short-Perennial | Th | ME + IR-TR + EU-SI | |
| Polypogon maritimus Willd. | Annual | G | COSM | |
| Polypogon monspeliensis (L.) Desf. | Annual | Th | ME + SA-SI + IR-TR | |
| Rostraria pumila (Desf.) Tzvelev. | Annual | Th | ME + SA-SI | |
| Schismus arabicus Nees. | Annual | Th | ME + SA + IR-TR | |
| Schismus barbatus (L.) Thell. | Annual | Th | ME + SA-SI + IR-TR | |
| Setaria pumila (Poir.) Roem. & Schutt. | Annual | Th | ME + IR-TR + S-Z | |
| Stipagrostis ciliata (Desf.) De Winter | Perennial | H | SA + IR-TR + SU | |
| Triticum aestivum L. | Annual | Th | SA + IR-TR + SU | |
| Labiatae | Phlomis brachyodon (Boiss.) Zoh. | Perennial | H | IR-TR |
| Salvia aegyptiaca L. | Perennial | Ch | SA-SI + S-Z | |
| Salvia lanigera Poir. | Perennial | Ch | ME + SA-SI | |
| Salvia spinosa L. | Perennial | H | ME + IR-TR | |
| Teucrium oliverianum Ging. ex. Benth. | Perennial | Ch | COSM | |
| Teucrium polium L. | Perennial | Ch | ME + IR-TR | |
| Leguminosae | Alhagi graecorum Boiss. | Perennial | Ch | ME + SA + IR-TR + SU |
| Astragalus bombycinus Boiss. | Annual | H | SA-SI + IR-TR | |
| Astragalus corrugatus Bertol. subsp. corrugatus | Annual | Th | IR-TR | |
| Astragalus hauarensis Boiss. | Annual | H | IR-TR | |
| Astragulus schemperenniali Boiss. var. subsessilis | Annual | Th | ME | |
| Astragalus sieberi DC. | Perennial | Ch | SA + IR-TR | |
| Astragulus spinosus (Forssk.) Muschl. | Perennial | Ch | SA-SI + IR-TR | |
| Astragalus tribuloides Delile var. tribuloides | Annual | Th | SA-SI + IR-TR | |
| Hippocrepis areolata Desv. | Annual | Th | ME + S-Z | |
| Hippocrepis cyclocarpa Murb. | Annual | Th | ME | |
| Hippocrepis multisiliquosa L. | Annual | Th | IR-TR | |
| Medicago laciniata (L.) Mill. var. brachyacantha Boiss. | Annual | Th | SA-SI | |
| Medicago laciniata (L.) Mill. var. laciniata | Annual | Th | SA-SI | |
| Medicago monospeliaca (L.) Trautv. | Annual | Th | ME + IR-TR | |
| Medicago polymorpha L. | Annual | Th | ME + IR-TR + EU-SI | |
| Melilotus albus Medik. | Annual | Th | ME + SA + IR-TR | |
| Melilotus indicus (L.) All. | Annual | Th | PAL | |
| Onobrychis ptolemaica (Delile.) DC. | Perennial | H | IR-TR | |
| Tephrosia purpurea (L.) Perennials. subsp. Leptostachya (DC.) Brummitt, Bol. | Perennial | Ch | SA-SI + S-Z | |
| Trigonella hamosa L. | Annual | Th | ME + SA-SI | |
| Trigonella laciniata L. | Annual | Th | ME + SA-SI + IR-TR | |
| Trigonella stellata Forssk. | Annual | Th | SA-SI + IR-TR | |
| Malvaceae | Malva parviflora L. | Annual | Th | PAN |
| Orobanchaceae | Cistanche phelypaea (L.) Cout. | Perennial | P | ME + SA-SI + IR-TR + S-Z |
| Orobanche tinctoria Forssk | Perennial | P | SA-SI + IR-TR | |
| Plantaginaceae | Plantago amplexicaulis Cav. subsp. Bauphula (Edgew.) Rech. f. | Annual | Th | SA-SI + IR-TR |
| Plantago ciliata Desf. | Annual | Th | ME + SA-SI + IR-TR | |
| Plantago crassifolia Forssk. | Annual | H | ME | |
| Plantago lagopus L. | Annual | Th | ME + IR-TR | |
| Plantago lanceolata L. | Annual | Th | ME + SA-SI + IR-TR | |
| Plantago ovata Forssk. | Annual | Th | SA + IR-TR | |
| Plantago psammophila Agnew & Chalibi-Ka, bi. | Annual | Th | SA-SI + IR-TR | |
| Polygonaceae | Emex spinosa (L.) Campd. | Annual | Th | PAN |
| Polygonum equisetiforme Sm. | Annual | Ch | M + IR-TR | |
| Rumex vesicarius L. | Perennial | Th | ME + SA-SI + S-Z | |
| Portulacaceae | Portulaca oleracea L. | Annual | Th | ME + SA-SI |
| Primulaceae | Anagallis arvensis L. var. arvensis | Annual | Th | ME + IR-TR + EU-SI |
| Resedaceae | Oligomeris linifolia (Vahl ex Hornem.) J. F. Macbr. (= Reseda linifolia Vahl ex Hornem.) | Annual | Th | ME + SA + S-Z |
| Reseda muricata C. Presl. | Annual | Th | SA-SI + IR-TR | |
| Scrophulariaceae | Veronica polita Fr. (= V. didyma sensu auct) | Annual | Th | M + IR-TR |
| Solanaceae | Hyoscyamus muticus L. | Perennial | Ch | SA-SI + IR-TR |
| Solanum nigrum L. | Perennial | Ch | COSM | |
| Umbelliferae | Ammi majus L. | Annual | Th | ME |
| Anisosciadium isociadum Bornm | Annual | Th | SA + IR-TR | |
| Anisosciadium lanatum Boiss. | Annual | Th | SA + IR-TR | |
| Deverra tortuosa (Desf.) DC. | Perennial | Ch | SA–SI | |
| Ducrosia anethifolia | Perennial | Th | IR-TR | |
| Ferula ovina | Perennial | Ch | IR-TR | |
| Pteroselinum crispum (Mill.) A. W. Hill. = P. Sativum (L.) P. Hoffm. | Biennial | Th | PAL | |
| Tordylium aegyptiacum (L.) Poir. | Annual | Th | ME + SA | |
| Urticaceae | Forsskaolea tenacissima L. | Annual | H | SA-SI + S-Z |
| Zygophyllaceae | Fagonia bruguieri DC. | Perennial | H | SA-SI + IR-TR |
| Fagonia glutinosa Delile. | Perennial | H | SA-SI | |
| Peganum harmala L. | Perennial | H | ME + SA–SI + IR–TR | |
| Tribulus macropterus Boiss. | Annual | Ch | SA-SI + IR-TR | |
| Tribulus megistopterus Kralik. | Annual | Ch | SA-SI + IR-TR | |
| Zygophyllum simplex L. | Annual | Th | SA-SI + S-Z |
Vegetation type: Annual = annualual; Bi = biennial; Perennial = perennialennial; s-Perennial = short perennialennial.
Life form:-Ch = chamaephytes; Cr = cryptophytes; G = geophytes; H = hemicryptophytes; P = parasites; Ph = phanerophytes; Th = therophytes.
Floristic categories (the Chorotypes): COSM = Cosmopolitan; EU-SI = Euro-Siberian; IR-TR = Irano-Turanian; ME = Mediterranean; PAL = Palaeotropical; PAN = Pantropical; S-Z = Sudano-Zambezian; SA = SaharoArabian; SA-SI = Saharo-Sindian; SU = Sudanian; TR = Tropical.
From the standpoint of species richness within the described families of the region, only one family exceeded 30 species (Compositae: 48), one family was represented by 26 species (Gramineae), the third family contained 22 species (Leguminosae), two families had more than 10 species (Caryophyllaceae: 12 and Cruciferae: 17). Moreover, seven families showed eight to five species (Boraginaceae: 5, Labiatae and Zygophyllaceae: 6, Plantaginaceae: 7, Umbelliferae and Chenopodiaceae: 8 genera). As well, two families were represented by three species, six possessed two species only. Finally 12 families were represented by a single species (Table 1).
With regard to the growth type, the majority of the recorded species in this study were annual plants (136 species or 69.5% of the total recorded species).The second most recorded growth type was the perennials, which were represented by 53 species (27.0% of the total species). Biennial, annual or short-perennial growth types in the investigated area do not exceeded two species and ultimately only one species was recorded as a short-perennial (Launaea mucronata sp. mucronata) (Table 2, Fig. 3).
Table 2.
Tubular summary showing the total number of families, genera and species, growth types and life forms of collected plants.
| Total number of |
Growth type |
Life forms |
||||||
|---|---|---|---|---|---|---|---|---|
| Families | Genera | Species | Type | Number of species | Percentage (%) | Form | Number of species | Percentage (%) |
| 31 | 130 | 196 | Annual | 136 | 69.5 | H | 16 | 8.2 |
| Perennial | 53 | 27 | Ch | 33 | 16.8 | |||
| Biennial | 2 | 1 | Ph | 1 | 0.5 | |||
| Annual or short-Perennial | 2 | 1 | Th | 140 | 71.5 | |||
| P | 3 | 1.5 | ||||||
| Annual or Biennial | 2 | 1 | G | 2 | 1 | |||
| Cr | 1 | 0.5 | ||||||
| Short-Perennial | 1 | 0.5 | ||||||
| Total Number | 196 | 100% | Total Number | 196 | 100% | |||
Figure 3.
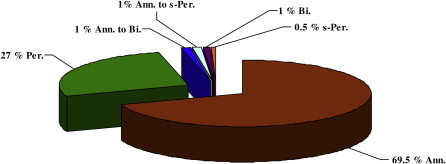
Vegetation type relative spectrum of Wadi Arar vegetarian: Ann = annual; Bi = biennial; perennial; s-per = short perennial.
Table 2 showed the life form spectra of the recorded species according to the classification of Raunkiaer (1937). Therophytes were the most frequent life form plants constituted by 140 species, followed by chamaephytes with 33 species and hemicryptophytes with 16 species, with a percentage of 71.5%, 16.8% and 8.2% of the total registered species respectively. Parasite plants were represented by three species from three different genera (Cuscuta campestris, Cistanche phelypaea and Orobanche tinctoria). Two species were geophytes (Cynodon dactylon and Polypogon maritimus) and both cryptophytes and phanerophytes were represented only by single species (Reichardia tingitana and Calotropis procera respectively) (Table 2, Fig. 4).
Figure 4.
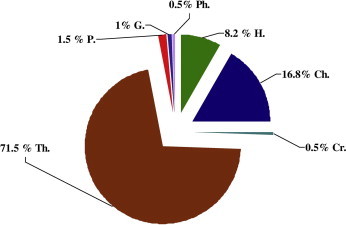
Life form relative spectrum of Wadi Arar vegetarion: Ch = Chamaephytes; Cr = Crptophytes; G = geophytes; H = hemicrptophytes; P = Parasites; Ph = phanerophytes; Th = therophytes.
From a phytogeographical point of view, the recorded species in the different sectors of Wadi Arar may be classified as monoregional, biregional or pluriregional taxa. A total of 46 species representing 23.5% of the total number of recorded species were monoregional taxa of different affinities. The recorded monoregional species fall under three main phytochoria: Irano-Turanian taxa (15 species forming 7.7% of recorded species), Saharo-Sindian taxa (13 species forming 6.6% of recorded species) and Saharo-Arabian taxa (10 species forming 5.1% of recorded species). Among these, six species were typical Mediterranean species (Atractylis carduus, Helichrysum conglobatum, Astragulus schemperenniali, Hippocrepis cyclocarpa, Plantago crassifolia and Ammi majus). Mediterranean species were rarely represented in different sectors of the Wadi. The last phytochoria (Sudano-Zambezian) was rarely represented in the different sectors of the Wadi with only two species (Haloxylon salicornicum and Eragrostis aegyptiaca) forming 1.0% of the total number of recorded species (see Table 3, Figs. 5 and 6).
Table 3.
The number of recorded species belonging to the main floristic categories and their relevant percent (%) abbreviations.
| Phytochoria | Number of species | Percentage (%) |
|---|---|---|
| Monoregional | ||
| ME | 06 | 3.1 |
| SA-SI | 13 | 6.6 |
| IR-TR | 15 | 7.7 |
| SA | 10 | 5.1 |
| S-Z | 02 | 1.0 |
| Total | 46 | 23.5 |
| Biregionals | ||
| SA-SI + S-Z | 17 | 8.7 |
| SA + S-Z | 01 | 0.5 |
| ME + SA-SI | 19 | 9.7 |
| ME + IR-TR | 19 | 9.7 |
| SA + IR-TR | 06 | 3.1 |
| ME + SA | 08 | 4.1 |
| IR-TR + EU-SI | 02 | 1.0 |
| SA-SI + IR-TR | 18 | 9.2 |
| ME + S-Z | 01 | 0.5 |
| Total | 91 | 46.5 |
| Pleuriregionals | ||
| ME + SA-SI + IR-TR | 14 | 7.1 |
| ME + SA + IR-TR | 06 | 3.1 |
| ME + IR-TR + EU-SI | 06 | 3.1 |
| ME + IR-TR + S-Z | 02 | 1.0 |
| ME + SA-SI + IR-TR + EU-SI | 02 | 1.0 |
| ME + SA-SI + IR-TR + S-Z | 02 | 1.0 |
| ME + SA + IR-TR + EU-SI | 01 | 0.5 |
| ME + SA + IR-TR + SU | 01 | 0.5 |
| ME + SA + S-Z | 01 | 0.5 |
| ME + SA-SI + S-Z | 01 | 0.5 |
| SA–SI + IR–TR + S–Z | 01 | 0.5 |
| SA + IR-TR + SU | 02 | 1.0 |
| Total | 39 | 19.8 |
| Cosm | 12 | 6.12 |
| Pal | 04 | 2.04 |
| Pan | 04 | 2.04 |
| Total | 20 | 10.2 |
Figure 5.
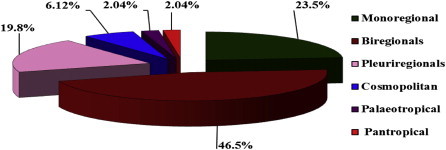
Floristic category spectrum of Wady Arar according to number of vegetarian regions.
Figure 6.
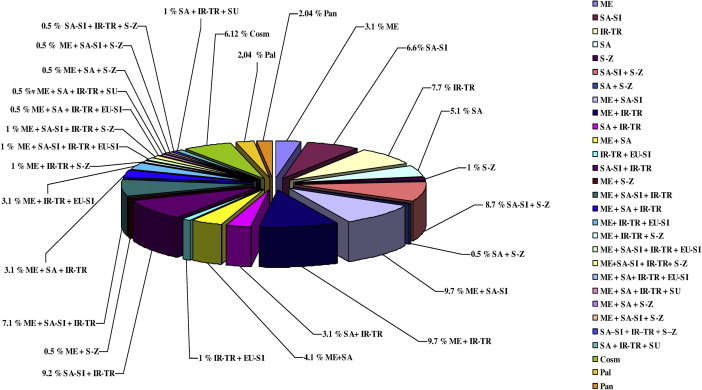
Floristic category of Wadi Arar: COSM = Cosmopolitian; EU-SI = Euro-Siberian; IR-TR = Irano-Turanian; SA-SI = Saharo-Sindian; SU = Sudanian; TR = Tropical.
The other 91 species (46.5%) were biregional taxa. Both Mediterranean-Saharo-Sindian and Mediterranean-Irano-Turanian phytochorial regions were represented by 19 species (9.7%). Saharo-Sindian-Irano-Turanian region was represented by 18 species (9.2%). Saharo-Sindian-Irano-Turanian region was represented by 17 species (8.7%). Both Mediterranean-Saharo-Arabian and Saharo-Arabian-Irano-Turanian regions were represented by eight species (4.1%) and six species (3.1%) respectively. Irano-Turanian-Euro-Siberian region was represented by two species (Artemisia scoparia and Centaurea sinaica). Only one species was registered in both Saharo-Arabian-Sudano-Zambezian region (Calotropis procera) and Mediterranean-Sudano-Zambezian region (Hippocrepis areolata) (Table 3, Figs. 5 and 6).
39 species representing 19.8% of the recorded species were pluriregional taxa of different affinities. The recorded pluriregional species fell also under three main phytochoria: Mediterranean-Saharo-Sindian-Irano-Turanian phytochorion (14 species forming 7.1% of recorded species) and both Mediterranean-Saharo-Arabian-Irano-Turanian and Mediterranean-Irano-Turanian-Euro-Siberian phytochoria (six species for each phytochorion forming 3.1% of recorded species). Mediterranean-Irano-Turanian-Sudano-Zambezian, Mediterranean-Saharo-Sindian-Irano-Turanian-Euro-Siberian, Mediterranean-Saharo-Sindian-Irano-Turanian-Sudano-Zambezian and SaharoArabian-Irano-Turanian-Sudanian regions were represented by two species (1.0%) which were Lamarckia aurea, Setaria pumila; Sisymbrium irio, Zilla spinosa; Citrullus colocynthis, Cistanche phelypaea and Stipagrostis ciliata, Triticum aestivum respectively. Only one species (or 0.5% of the registered species) occurred in the following regions: Mediterranean-SaharoArabian-Irano-Turanian-Euro-Siberian (Eruca sativa), Mediterranean-SaharoArabian-Irano-Turanian-Sudanian (Alhagi graecorum), Mediterranean-SaharoArabian-Sudano-Zambezian (Oligomeris linifolia), Mediterranean-Saharo-Sindian-Sudano-Zambezian (Rumex vesicarius) and Saharo-Sindian-Irano-Turanian-Sudano-Zambezian (Launaea nudicaulis). The remaining twenty species were distributed as follows: 12 species are cosmopolitan, followed by four palaeotropical species and four pantropical species (Table 3, Figs. 5 and 6).
5. Discussion and conclusions
By the end of the present study, which focused on flora of Wadi Arar area, five of collected families (Compositae, Gramineae, Leguminosae, Caryophyllaceae and Cruciferae) contributed nearly to two thirds of the total flora. The most representative families of this region were corresponded to a typical Mediterranean-North African floristic structure and this agrees with the results of Quézel (1978) study.
According to Al-Hassan (2006), about 458 species which belong to these floristic structures were represented in the northern sector of the Kingdom of Saudi Arabia. In Wadi Arar area, the present survey recorded 196 species, and this represented over one third of the checklist recorded before in the whole northern region of the Kingdom. The total number of identified genera was 130, indicating a high generic index of 1.5 (196/130). These results were conforming to those of Pielou (1975) and Magurran (1988) studies, where the taxonomic diversity is higher in an area in which the species were divided between many genera. Taxonomic diversity in Wadi Arar is 1.5 for species/genera ratio and 6.3 for genera/families. This indicates a higher taxonomic diversity in the studied region than in other areas (Tables 1 and 2). This high diversity was due to the abundance of water sources, soil fertility and the diversity of relief (open shallow valleys, mountains and sand dunes) as mentioned by Al-Shammeri (2010) and Al-Shammari and Sharawy (2010).
The wild vegetation in this studied area comprised 196 plant species, including 136 annuals (69.5%) and 53 perennials (27.0%). The high contribution of annuals can be attributed to their short life cycle that enables them to resist the instability of the ecosystem (Gomaa, 2012). Moreover, they were generally characterized by a high allocation of resources to the reproductive organs (Harper, 1977) and the production of flowers early in their lifespan to ensure some seed production even in a year when the growing season is cut short (Sans and Masalles, 1995). These results corroborated previous studies focusing on Hail province (Al-Turki and Al-Qlayan, 2003; Sharawy and Alshammari, 2009 and Al-Shammari and Sharawy, 2010). Moreover, members of the family Compositae dominated the wild plants of Wadi Arar region (48 species), followed by Gramineae (26 species), Leguminosae (22 species), Cruciferae (17 species) and Caryophyllaceae (17 species), (Table 2, Fig. 3).
The vegetation of northern region does not had appreciable contribution to the flora of Saudi Arabia with regard to other parts of the country (Al-Turki and Al-Qlayan, 2003). Vegetation of the northern part of the country is also dominated by annual species. However, perennial components such as Salsola sp., Haloxylon salicornicum, Artemisia sp., Zilla spinosa, Hyoscyamus muticus, Fagonia glutinosa etc. are forming large communities in all range lands. (http://www.plantdiversityofsaudiarabia.info/biodiversity-saudi-arabia/flora/Flora.htm). The studied area was widely regarded as a transition zone between the floristic regions of Mediterranean, Irano-Turanian and Saharo-Sindian, Saharo-Arabian or Sudano-Zambezian regions. The influences of these phytogeographical zones were very much reflected in the flora of Northern region. The other floristic elements such as cosmopolitan, pantropical, palaeotropical, Euro-Siberian and Sudanian elements were represented by a varying number of species reflecting their differential capability to penetrate the region.
Among the recorded species from this region, some were extremely rare and poorly known and their results reveal that some of these plants had been collected from a single locality (Calotropis procera, Gypsophila sp., Cistanche phelypaea and Orobanche tinctoria), while others have been collected in the studied area only once or twice (Aerva javanica, Anthemis sp., Koelpinia linearis, Solanum nigrum and Anagallis arvensis), (Al-Turki and Al-Qlayan, 2003).
Regarding the life form spectra, therophytes had the highest contribution, followed by chamaephytes, hemicryptophytes, parasites, geophytes and finally both phanerophytes and cryptophytes (Tables 1 and 2 and Fig. 4). The life form of desert plants is closely linked with rainfall, topography and land form (Kassas and Girgis, 1965; Zohary, 1973; Orshan, 1986). Therophytes (which had a lifespan as short as few weeks) are characteristic of desert regions (Gomaa, 2012).
Considering the floristic categories, in the studied area, 46.5% of the recorded species were belonging to the biregional category: (Mediterranean-Saharo-Sindian and Mediterranean-Irano-Turanian phytochoria had 19 species, Saharo-Sindian-Irano-Turanian phytochoria possessed 18 species, Saharo-Sindian-Sudano-Zambezian phytochoria include 17 species, Mediterranean-Saharo-arabian phytochoria contained eight species, Saharo-arabian-Irano-Turanian phytochoria comprised six species, Irano-Turanian-Euro-Siberian phytochoria had two species (Artemisia scoparia and Centaurea sinaica) and only one species is present in both Saharo-arabian-Sudano-Zambezian phytochoria (Calotropis procera) and Mediterranean-Sudano-Zambezian phytochoria (Hippocrepis areolata).
A monoregional category was the second one with 23.5% of the total recorded species: (15 species in Irano-Turanian region, 13 species in Saharo-Sindian region, 10 species in Saharo-arabian region, six species in Mediterranean region and two species in Sudano-Zambezian region (Haloxylon salicornicum and Eragrostis aegyptiaca). Pluriregional category was the third one with 19.8% of the total recorded species in this area (14 species in Mediterranean-Saharo-Sindian-Irano-Turanian region, six species in both Mediterranean-Saharo-arabian-Irano-Turanian and Mediterranean-Irano-Turanian-Euro-Siberian regions, two species in Mediterranean-Irano-Turanian-Sudano-Zambezian region (Lamarckia aurea and Setaria pumila), Mediterranean-Saharo-Sindian-Irano-Turanian-Euro-Siberian region (Sisymbrium irio and Zilla spinosa), Mediterranean-Saharo-Sindian-Irano-Turanian-Sudano-Zambezian region (Citrullus colocynthis and Cistanche phelypaea) and Saharo-arabian-Irano-Turanian-Sudanian region (Stipagrostis ciliata and Triticum aestivum) and only one species in Mediterranean-Saharo-arabian-Irano-Turanian-Euro-Siberian region (Eruca sativa), Mediterranean-Saharo-arabian-Irano-Turanian-Sudanian region (Alhagi graecorum), Mediterranean-Saharo-arabian-Sudano-Zambezian region (Oligomeris linifolia), Mediterranean-Saharo-Sindian-Sudano-Zambezian region (Rumex vesicarius) and Saharo-Sindian-Irano-Turanian-Sudano-Zambezian region (Launaea nudicaulis). Cosmopolitan category has 6.12% and eventually both Palaeotropical and Pantropical categories had 2.04%, (Table 3 and Figs. 5 and 6).
The results indicate that Irano-Turanian elements predominate the studied area (97 species, 49.5%), and were represented as mono-regional (15 species, 7.7%), bi-regional under the influence of Mediterranean, SaharoArabian, Euro-Siberian and Saharo-Sindian regions (45 species, 23%) and Pleuriregionals under the influence of Mediterranean, Saharo-Sindian, SaharoArabian, Euro-Siberian, Sudano-Zambezian and Sudanian regions (37 species, 18.8%). Mediterranean region was represented by 89 species (or 45.4%), followed by Saharo-Sindian region (87 species, 44.4%), followed by Sudano-Zambezian region (45 species, 23%) and at last SaharoArabian region (36 species, 18.4%). Taxa of wide geographical range, like Cosmopolitan (12 species, 6.12%), Palaeotropical (4 species, 2.04%) and Pantropical (4 species, 2.04%), have apparently lower presence value (Table 3).
Acknowledgment
The authors are grateful to Deanship of Scientific Research, Northern Border University, for supporting this research. They also thank Dr. Arabi M. Guetat, Biology Department, College of science, Northern Border University for continuous help throughout the present study.
Footnotes
Peer review under responsibility of King Saud University.

Contributor Information
Ahmed K. Osman, Email: ahmosman2000@yahoo.com, osman_a_2000@yahoo.com.
Faraj Al-Ghamdi, Email: falghamdi@nbu.edu.sa, faalghamdi@kau.edu.sa.
Abdulhakim Bawadekji, Email: hakimbawadekji@gmail.com.
References
- Abdel Khalik K., El-Sheikh M., El-Aidarous A. Floristic diversity and vegetation analysis of Wadi Al-Noman, Holy Mecca, Saudi Arabia. Turk. J Botany. 2013;37:894–907. [Google Scholar]
- Alatar A., El-Sheikh M.A., Thomas J. Vegetation analysis of Wadi Al-Jufair, a hyper-arid region in Najd, Saudi Arabia. Saudi J. Biol. Sci. 2012;19:357–368. doi: 10.1016/j.sjbs.2012.04.003. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- Aleem A.A. A contribution to the study of sea grasses along the Red Sea coast of Saudi Arabia. Aquat. Botany. 1979;7:71–78. [Google Scholar]
- Alfarhan, A.H., Al-Turki, T.A., Basahy, A.Y., 2005. Flora of Jizan Region. Vol. 1, King Abdulaziz City for Science and Technology (KACST).
- Al-Hassan H.O. Camel and Range Research Center; Al-Jouf, Saudi Arabia: 2006. Wild plants of Northern Region of the Kingdom of Saudi Arabia (Field guide with photography). Ministry of Agriculture. [Google Scholar]
- Al-Shammeri M.A. Hail University; Hail, Saudia Arabia: 2010. Historical Hema Faid. [Google Scholar]
- Al-Shammari A.M., Sharawy S.M. Wild plants diversity of the Hema Faid Region (Hail Province, Saudi Arabia) Asian J. Plant Sci. 2010;9(8):447–454. [Google Scholar]
- Alsherif E.A., Ayesh A.M., Allogmani A.S., Rawi S.M. Exploration of wild plants wealth with economic importance tolerant to difficult conditions in Khulais Governorate Saudi Arabia. Sci. Res. Essays. 2012;7(45):3903–3913. [Google Scholar]
- Al-Sodany Y.M., Mosallam H.A., Bazaid S.A. Vegetation analysis of Mahazat Al-Sayd protected area: the second largest fenced nature reserve in the world. World Appl. Sci. J. 2011;15(8):1144–1156. [Google Scholar]
- Al-Turki T.A., Al-Qlayan H.A. Contribution to the flora of Saudi Arabia: Hail region. Saudi J. Biol. Sci. 2003;10:190–222. [Google Scholar]
- Al Wadie H. Floristic composition and vegetation of Wadi Talha, Asser Mountains, South West Saudi Arabia. J. Biol. Sci. 2002;2:285–288. [Google Scholar]
- Atiqur Rahman M., Mossa J.S., Al-Said M.S., Al-Yahya M.A. Medicinal plant diversity in the flora of Saudi Arabia 1: a report on seven plant families. Fitoterapia. 2004;75:149–161. doi: 10.1016/j.fitote.2003.12.012. [DOI] [PubMed] [Google Scholar]
- Batanouny K., Ismail A. Plant communities along Medina-Badr road across the Hejaz Mountains, Saudi Arabia. Vegetatio. 1985;53:33–43. [Google Scholar]
- Chaudhary S.A. vol. 1-3. Ministry of Agriculture and Water press; Riyadh: 1999. 2001. (Flora of the Kingdom of the Saudi Arabia). [Google Scholar]
- Chaudhary S.A., Al-Jowaid A.A. National Agriculture and Water Research Center. Ministry of Agriculture and Water; Riyadh, Saudi Arabia: 1999. Vegetation of the Kingdom of Saudi Arabia. [Google Scholar]
- Collenette, S., 1999. Wild Flowers of Saudi Arabia, pp. 799, Riyadh: National Commission for Wild Life Conservation and Development (VCWCD).
- Cope, T., 1985. A key to the grasses of Arabian Peninsula. Arabian Journal Science Research, Special Publication.
- Country Profile: Saudi Arabia, September (2006) Library of Congress – Federal research division (http://lcweb2.loc.gov/frd/cs/profiles/Saudi_Arabia.pdf).
- El-Ghanem W.A., Hassan L.M., Galal T.M., Badr A. Floristic composition and vegetation analysis in Hail region north of central Saudi Arabia. Saudi J. Biol. Sci. 2010;17:119–128. doi: 10.1016/j.sjbs.2010.02.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- El Karemy Z., Zayed K. Distribution of Plant Communities across Al Abna Escarpment, SW Saudi Arabia. Phyton (Horn, Austria) 1992;32:79–101. [Google Scholar]
- Fakhireh A., Ajorlo M., Shahryari A., Mansouri S., Nouri S., Pahlavanravi A. The autecological characteristics of Desmostachya bipinnata in hyper-arid regions. Turk J Botany. 2012;36:690–696. [Google Scholar]
- Farrag H.F. Floristic composition and vegetation-soil relationships in Wadi Al-Argy of Taif region, Saudi Arabia. Int. Res. J. Plant Sci. 2012;3(8):147–157. [Google Scholar]
- Fayed A., Zayed K. Vegetation along Makkah-Taif road (Saudi Arabia) Arabian Gulf J. Sci. Res. 1989;7:97–117. [Google Scholar]
- Gomaa N.H. Composition and diversity of weed communities in Al-Jouf province, northern Saudi Arabia. Saudi J. Biol. Sci. 2012;19:369–376. doi: 10.1016/j.sjbs.2012.05.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hajar A., Yousef M., Baeshin N. Studies on the plant ecology and phytosociology of Al-Bahah region, Saudi Arabia: 1. Area along Al-Bahah – Al-Qonfodah road. Bull. Faculty Sci. Assuit Univ. 1998;27:53–84. [Google Scholar]
- Harper J.L. Academic Press; London: 1977. Population Biology of Plants. [Google Scholar]
- Hegazy A.K., El-Demerdash M.A., Hosni H.A. Vegetation, species diversity and floristic relations along an altitudinal gradient in south-west Saudi Arabia. J. Arid Environ. 1998;38:3–13. [Google Scholar]
- Ihsanullah Daur. Plant flora in the rangeland of western Saudi Arabia. Pak. J. Bot. 2012;44:23–26. [Google Scholar]
- Kassas M., Girgis W.A. Habitat and plant communities in the Egyptian desert. V. The limestone plateau. J. Ecol. 1964;52:107–119. [Google Scholar]
- Kassas M., Girgis W.A. Habitat and plant communities in the Egyptian desert. VI. The units of a desert ecosystem. J. Ecol. 1965;53:715–728. [Google Scholar]
- Korkmaz M., Ozcelik H. Soil-plant relations in the annual Gypsophila (Caryophyllaceae) taxa of Turkey. Turk. J. Botany. 2013;37:85–98. [Google Scholar]
- Kürschner H., Neef R. A first synthesis of the flora and vegetation of the Tayma oasis and surroundings (Saudi Arabia) Plant Div. Evol. 2011;129:27–58. [Google Scholar]
- Magurran A.E. Princeton University Press; Pinceton, New Jersy: 1988. Ecological diversity and it, s Measurement Princeton. [Google Scholar]
- Migahid A.M. vol. I–III. King Abdul Aziz University Press; Jeddah: 1996. (Flora of Saudi Arabia). [Google Scholar]
- Miller, A.G., Cope, T.A., 1996. Flora of Arabian Peninsula and Socatra. Edinburgh University Press in association with Royal Botanical Gardens Edinburgh and Royal Botanical Gardens Kew. England.
- Mosallam H.A. Comparative study on the vegetation of protected and non-protected areas, Sudera, Taif, Sauidi Arabia. Int. J. Agric. Biol. 2007;9:202–214. [Google Scholar]
- Orshan G. The desert of the Middle East. In: Evenari M., Noy-Meir I., Goodall D.W., editors. Ecosystems of the World, 12 B, Hot Deserts and Arid Shrublands. Elsevier; Amsterdam: 1986. [Google Scholar]
- Pielou E.C. 1st edn. Wiley Interscience; New York: 1975. Ecological diversity. [Google Scholar]
- Quézel P. Analysis of the flora of Mediterranean and Saharan Africa. Ann. Missouri Botan. Garden. 1978;65:479–534. [Google Scholar]
- Sans F.X., Masalles R.M. Phenological patterns in an arable land weed community related to disturbance. Weed Res. 1995;35:321–332. [Google Scholar]
- Shaltout K.H., El-Sheikh M.A. Vegetation of the urban habitats in the Nile Delta region. Egypt. Urban Ecosystems. 2003;6:205–221. [Google Scholar]
- Sharawy S.M., Alshammari A.M. Checklist of poisonous plants and animals in Aja Mountain, Hail region, Saudi Arabia. Aust. J. Basic Appl. Sci. 2009;3:2217–2225. [Google Scholar]
- Siddiqui A.Q., Al-Harbi A.H. A preliminary study of the ecology of Wadi Hanifah stream with reference to animal communities. Arab Gulf J. Sci. Res. 1995;13:695–717. [Google Scholar]
- Waly N.M., Al-Zahrani H.S., Felemban W.F. Taxonomical studies of some acacia seeds growing in Saudi Arabia. J. Am. Sci. 2012;8(3):264–275. [Google Scholar]
- Wickens G.E. The flora of Jebel Marra (Sudan Republic) and its geographical affinities. Kew Bull. Additional Ser. 1978:5–385. [Google Scholar]
- Zohary, M., 1973. Geobotanical foundations of the Middle East. Stuttgart: Gustav Fischer –Verlag.