

Abstract
AIM: To investigate the role of P-selectin, intercellular adhesion molecule-1 (ICAM-1) and dendritic cells (DCs) in liver/kidney of rats with hepatic/renal ischemia-reperfusion injury and the preventive effect of anti-P-selectin lectin-EGF domain monoclonal antibody (anti-PsL-EGFmAb) on the injury.
METHODS: Rat models of hepatic and renal ischemia-reperfusion were established. The rats were then divided into two groups, one group treated with anti-PsL-EGFmAb (n = 20) and control treated with saline (n = 20). Both groups were subdivided into four groups according to reperfusion time (1, 3, 6 and 24 h). The sham-operated group (n = 5) served as a control group. DCs were observed by the microscopic image method, while P-selectin and ICAM-1 were analyzed by immunohistochemistry.
RESULTS: P-selectin increased significantly in hepatic sinusoidal endothelial cells and renal tubular epithelial cells 1 h after ischemia-reperfusion, and the expression of ICAM-1 was up-regulated in hepatic sinusoid and renal vessels after 6 h. CD1a+CD80+DCs gradually increased in hepatic sinusoidal endothelium and renal tubules and interstitium 1 h after ischemia-reperfusion, and there was the most number of DCs in 24-h group. The localization of DCs was associated with rat hepatic/renal function. These changes became less significant in rats treated with anti-PsL-EGFmAb.
CONCLUSION: DCs play an important role in immune pathogenesis of hepatic/renal ischemia-reperfusion injury. Anti-PsL-EGFmAb may regulate and inhibit local DC immigration and accumulation in liver/kidney.
Keywords: Adhesion molecules, Dendritic cells, Hepatic/renal ischemia-reperfusion injury, Anti-P-selectin lectin-EGF domain monoclonal antibody
INTRODUCTION
Hepatic/renal ischemia-reperfusion injury is very common clinically, but the exact mechanism is unknown[1-6]. Recently, it has been reported that cell adhesion molecules play a crucial role in ischemia-reperfusion injury by mediating the interaction of polymorphonuclear neutrophils with endothelium[7-12]. There are evidences that inhibition of the activities of adhesion molecules especially selectins prevented leukocyte adhesion, migration and recruitment[12-15]. In addition, dendritic cells (DCs) and their biological functions have also been implicated in inflammatory diseases, autoimmune diseases, graft rejection and tumors[16-21]. Thus, the roles of DCs in adhesion and migration have attracted great interest because of the observations that their migration into inflamed tissue is mediated by P- or E-selectins and that they play an important initiating and modulating role in immune responses within inflamed tissues[15,17-21]. However, the roles of DCs involved in leukocyte infiltration and immune pathogenesis of hepatic/renal ischemia-reperfusion injury are largely unknown. In view of the previous studies[22,23], we further investigated the roles of P-selectin, intercellular adhesion molecule-1 (ICAM-1) and DCs in rat hepatic/renal ischemia-reperfusion injury and the preventive effect of anti-P-selectin lectin-EGF domain monoclonal antibody (anti-PsL-EGFmAb) on the injury.
MATERIALS AND METHODS
Reagents
The antibodies and reagents used in the experiments included goat anti-mouse CD1a polyclonal antibody (Santa Cruz Biotech Co., California, USA), rabbit anti-mouse CD80 polyclonal antibody (BD Biotech Co., San Diego, CA), anti-rabbit IgG-FITC and anti-goat IgG-RPE (Jackson Co., Baltimore, USA), P-selectin and ICAM-1 immunohistochemistry LSAB kits (Mei Hua Biotech Development Co., Beijing, China), serum alanine aminotransferase (ALT), aspartate aminotransferase (AST), blood urea nitrogen (BUN), serum creatinine (SCr) kits (Shanghai Institute of Medical and Chemistry, Shanghai, China) and anti-PsL-EGFmAb was produced in-house[24].
Animal models
Ninety male Wistar rats (Shanghai Experimental Animal Centers of Chinese Academy of Sciences), weighing 200±10 g were given free access to food and water for three days before the experiments. The rats were anesthetized with 2.5% sodium pentobarbital intraperitoneally and randomly divided into two groups. In group one (n = 40), the ligament linking liver, diaphragm and abdominal wall were separated, then portal vein and liver artery that drain blood to left hepatic lobe were blocked with a microvascular clamp for 60 min. After that the clamp was removed, and reperfusion occurred. In group two (n = 40), the left renal artery was blocked with a microvascular clamp for 60 min, then the clamp was removed and reperfusion started. The right kidney was cut off before the procedure. The two groups were then randomly divided into two subgroups, one treated with anti-PsL-EGFmAb (anti-PsL-EGFmAb-treated group, n = 20) and one treated with normal saline only (saline-treated group, n = 20). Anti-PsL-EGFmAb (2 mg/kg) or normal saline was injected intravenously 5 min before reperfusion. Five rats in each group were sacrificed respectively at 1, 3, 6 and 24 h after reperfusion. Sham-operated rats with anesthesia and opening of celiac cavity but without blocking of hepatic or renal blood flow (n = 10) were used as controls.
Collection and measurement methods of specimens
Blood, hepatic and renal tissues were collected at different time points. Levels of serum AST, ALT, BUN and SCr were measured with a 747 automatic analyzer (Hitachi Boehringer Mannheim, Mannheim, Germany). Hepatic and renal tissue samples were fixed in 10% formalin and embedded in paraffin. Sections were cut 5 μm thick and stained with hematoxylin and eosin for light microscopic examination. Expression of P-selectin and ICAM-1 in hepatic/renal tissue was detected by an immunohistochemistry method with an LSAB kit.
Distribution of DCs in hepatic and renal tissues
Dual-label immunofluorescence staining for microscopic image analysis was used[25]. Hepatic and renal tissue sections were washed with PBS, and then blocked with 0.3% BSA for 20 min. The sections were incubated with the appropriate concentration of goat anti-mouse CD1a polyclonal antibody and rabbit anti-mouse CD80 polyclonal antibody overnight at 4 °C. Subsequently, the sections were washed with PBS, and added with the appropriate concentration of anti-rabbit IgG-FITC and anti-goat IgG-RPE antibodies respectively. The sections were incubated for 1 h at 37 °C, washed with PBS and then sealed. PBS was used as a negative control. DCs were observed by a microscopic image method. CD1a was positively stained by red-fluorescence and CD80 by green-fluorescence. Cells with dual staining as shown by yellow-fluorescence represented DCs. All data were input into the KS400 imaging process system and software (Vet 3.0), and the area, number and density of DC yellow-fluorescence were analyzed.
Statistical analysis
The data were presented as the mean±SD for each group. All analyses were performed using the SPSS 10.0 program. Correlation analysis and Student’s t test were used. P<0.05 was considered statistically significant.
RESULTS
Histopathologic evaluation
Grossly the left hepatic lobe was more swollen than the right lobe, and both were dark in color 1 h after reperfusion in the saline-treated group. Interstitial congestion and infiltration of inflammatory cells were observed under the light microscope. The renal cortices were pale, and the renal medullae displayed blood stagnation and were dark 1 h after reperfusion in the saline-treated group. Edema, degeneration with different extents and necrosis of renal tubular epithelial cells were observed under the light microscope. Simultaneously, interstitial congestion, edema and infiltration of inflammatory cells were also found. However, the gross appearance of the liver and kidney in the anti-PsL-EGFmAb-treated group was similar to that in the sham-operated group. No degeneration, necrosis, and interstitial changes appeared in hepatic cells and tubular cells except for swelling.
Hepatic and renal function evaluation
The serum levels of ALT (628±91 μ/L) and AST (1608±199 μ/L) 24 h after hepatic reperfusion were much higher in the saline-treated group than those in the sham-operated group (52±11 μ/L and 80±17 μ/L respectively, P<0.01). The group treated with anti-PsL-EGFmAb had significantly low levels of ALT (156±21 μ/L) and AST (325±37 μ/L) compared to the saline-treated group (P<0.01).
The levels of BUN (14.54±0.67 mmol/L) and SCr (102.12±4.67 μmol/L) 24 h after hepatic reperfusion were much higher in the saline-treated group than those in the sham-operated group (7.88±0.57 mmol/L and 39.00±4.47 μmol/L respectively, P<0.01). However, the anti-PsL-EGFmAb-treated group presented with significantly low levels of BUN (8.78±0.46 mmol/L) and SCr (45.62±3.17 μmol/L) compared to the saline-treated group (P<0.01).
Expression of adhesion molecules in hepatic and renal tissues
P-selectin was expressed widely in hepatic and renal tissues 1 h after reperfusion, which was mainly distributed in hepatic sinusoidal endothelium and tubular epithelium as yet maintained low level at 24 h after reperfusion. In addition, it was also expressed on part of hepatic cellular membrane, glomerular mesangium, capillary loops, and interstitium. ICAM-1 was continuously and increasingly up-regulated on hepatic sinusoidal endothelium and renal interstitium. However, treated with anti-PsL-EGFmAb, there were no obvious expressions of P-selectin and ICAM-1 in the hepatic and renal tissues of the rats.
DC distributive patterns in hepatic and renal tissues
CD1a+CD80+DCs were rarely distributed in hepatic and renal tissues in the sham-operated group, but largely increased in the saline-treated group as demonstrated by the imaging analysis (P<0.01) (Figure 1A-D and Tables 1, 2). Further results found that DCs were mainly distributed in hepatic sinusoids and renal interstitium. CD1a+CD80+DCs gradually increased from 1 h after reperfusion and reached the peak at 24 h (P<0.01) (Table 3). The number of CD1a+CD80+DCs in hepatic and renal tissues were markedly reduced in the anti-PsL-EGFmAb-treated group compared to the saline-treated group (P<0.05) especially at 24 h after reperfusion (P<0.01) (Figure 1E-F, Table 3).
Figure 1.
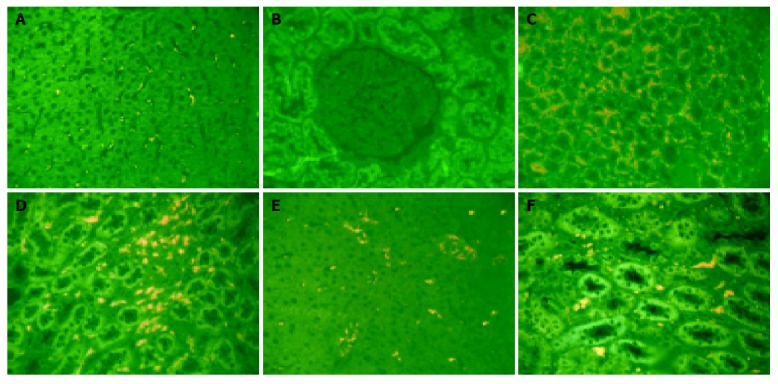
Dendritic cell distribution in each group (×1200). A: DCs were rare in rat hepatic tissue after reperfusion in the sham-operated group; B: DCs were rare in rat renal tissue after reperfusion in the sham-operated group;C: DCs increased in rat hepatic tissue after reperfusion in the saline-treated group; D: DCs increased in rat renal tissue after reperfusion in the saline-treated group; E: The number of DCs in rat hepatic tissue after reperfusion was smaller in the anti-PsL-EGFmAb-treated group than in the saline-treated group; F: The number of DCs in rat renal tissue after reperfusion was smaller in the anti-PsL-EGFmAb-treated group than in the saline-treated group.
Table 1.
Dendritic cell distribution in rat hepatic tissue among the three groups.
| Group | n | Area (mm2) of CD1a+CD80+DCs | Number of CD1a+CD80+ DCs | Density (n/mm2) of CD1a+CD80+ DCs |
| Sham-operated | 5 | 415.8±259.4 | 98.5±56.8 | 626.4±291.5 |
| Saline-treated | 20 | 3 542.6±2 435.8d | 1 035.6±984.2d | 2 654.3±1 025.4d |
| Anti-PsL-EGFmAb-treated | 20 | 1 982.7±1 456.4bf | 625.4±341.3bf | 962.8±659.1bf |
P<0.01,
P<0.001 vs sham-operated group;
P<0.01 vs saline-treated group.
Table 2.
Dendritic cell distribution in rat renal tissue among the three groups.
| Group | n | Area (mm2) of CD1a+CD80+DC | Number of CD1a+CD80+DC | Density (n/mm2) of CD1a+CD80+DC |
| Sham-operated | 5 | 356.3±232.0 | 84.5±63.7 | 575.3±280.1 |
| Saline-treated | 20 | 2 726.9±1 678.4d | 866.9±685.4d | 1647.5±1 376.8d |
| Anti-PsL-EGFmAb-treated | 20 | 1 610.7±1 025.3bf | 383.4±257.6bf | 932.6±754.1bf |
P<0.01,
P<0.001 vs sham-operated group;
P<0.01 vs saline-treated group.
Table 3.
Dendritic cell distribution at different time points after reperfusion between saline-treated and anti-PsL-EGFmAb-treated groups.
| n |
CD1a+CD80+ DCs (n/mm2) in rat hepatic tissue |
CD1a+CD80+ DCs (n/mm2) in rat renal tissue |
|||
| Saline-treated | Anti-PsL-EGFmAb-treated | Saline-treated | Anti-PsL-EGFmAb-treated | ||
| 1 h | 5 | 1239.7±965.2 | 632.7±425.3a | 1076.4±563.9 | 573.5±376.8a |
| 3 h | 5 | 1364.5±1025.4 | 671.9±654.1a | 1132.5±925.7 | 659.8±475.2a |
| 6 h | 5 | 1748.9±1085.1 | 967.2±977.5a | 1435.6±952.8 | 975.3±612.8a |
| 24 h | 5 | 3428.6±1594.5 | 1249.4±872.9b | 2864.3±2041.5d | 1256.2±956.4b |
P<0.05,
P<0.01 vs saline-treated group,
P<0.01 vs 1 h, 3 h or 6 h group.
Relationship between DCs in hepatic/renal tissues and hepatic/renal functions
The correlation analysis showed that the number of DCs in rat liver and kidney was associated with ALT, AST, BUN and SCr (r = 0.338, 0.339, 0.412 and 0.526 respectively, all P<0.05).
DISCUSSION
Recent studies have indicated that the interactions of leukocytes with endothelium mediated by cell adhesion molecules, which form the adhesion cascade, are the critical multistep processes of leukocyte firm adhesion, activation and release of inflammatory mediators, which play an important role in hepatic/renal ischemia-reperfusion injury[1-15,21-23]. P-selectin mediates the initial, low-affinity leukocyte-endothelial cell interaction that is manifested as leukocyte rolling. This transient binding results in further leukocyte activation and subsequent firm adhesion and transendothelial migration of leukocytes, both of which are mediated by interactions between ICAM-1 and other specific adhesion molecules[26-29]. It has been demonstrated that blockade of P-selectin or ICAM-1 expression with their ligands attenuated leukocyte adherence and infiltration during ischemia and reperfusion injury[14,30-34]. Recently, DCs have attracted great interest in immune responses or tolerance and diseases as well as their modulating mechanisms to migrate inflamed tissue[16,19-21,35,36]. Pendl et al[19], found that DC migration into inflamed tissue was mainly mediated by P- or E-selectin at very early stage of inflammation, suggesting that DCs also play an important role in the initiation and modulation of the inflammatory response in tissues. However, the roles of DCs in the hepatic/renal ischemia-reperfusion injury and P-selectin in mediating DC migration into the hepatic and renal tissues are not clear so far.
In our study, hepatic and renal tissues appeared with significant pathologic damage after ischemia-reperfusion, including cell swelling, interstitial congestion and inflammatory cell infiltration, with the increased serum levels of ALT and AST, BUN and SCr. We also found that the expressions of P-selectin and ICAM-1 were time-dependent in the hepatic and renal tissues. P-selectin was markedly expressed on hepatic sinusoidal endothelium and renal tubular epithelium in the early stage of hepatic/renal reperfusion, and was maintained at a lower level in the late stage. ICAM-1 was mainly expressed on hepatic sinusoidal endothelium and renal tubular epithelium in the late stage of reperfusion. Moreover, P-selectin and ICAM-1 expressions in hepatic and renal tissues were down-regulated, and paralleled the improvement of hepatic/renal functions and the decrease of pathologic damage when anti-PsL-EGFmAb was injected into rats before reperfusion. It has been reported that leukocyte rolling and recruitment was delayed when P- or E-selectin deficient mice were infected. In addition, ischemia-reperfusion induced by renal injury was markedly attenuated in ICAM-1 deficient mice[20,26,37]. Here we revealed that P-selectin and ICAM-1 played an important role in inflammation initiated by leukocyte rolling, adhesion and recruitment into injured tissue. In addition, this study indicated that blockade of P-selectin and the early events of leukocyte adhesion attenuated inflammatory cell infiltration and pathologic damage, consistent with previous studies that blockade of P-selectin inhibits platelet and its activation and improves hepatic/renal functions after ischemia-reperfusion injury[38,39-42].
DCs are the most potent antigen-presenting cells that have many biological functions involved in many inflammatory processes[15,16,20,21]. In this study, we performed CD1a and CD80 dual-label immunofluorescence staining for the analysis of DC migration into hepatic and renal tissues, and found that CD1a+CD80+DCs were significantly recruited into hepatic and renal tissues of the saline-treated group, but were rare in the sham-operated group. CD1a+CD80+DCs were mainly distributed in the hepatic sinusoids and renal interstitium in agreement with acute immune inflammation of hepatic sinusoids and renal interstitial damage. Our results suggest that DCs may be involved in neutrophil infiltration within the liver and kidney in the early stage of ischemia-reperfusion because their numbers paralleled hepatic and renal damage. Furthermore, we found that CD1a+CD80+DCs started to accumulate within the hepatic and renal tissues 1 h after reperfusion and reached the peak at 24 h after reperfusion, matching with the increasing hepatic and renal damage. CD1a+CD80+DC distribution and populations within hepatic and renal tissues were positively associated with hepatic and renal functions, indicating that DCs play a major role in hepatic/renal reperfusion injury. The potential mechanisms of DCs’ insult on the liver and kidney after the onset of reperfusion might be that they migrate into the injured hepatic sinusoids and renal interstitium, subsequently activate CD4+T cells and allow them to release pro- and anti-inflammatory cytokines, which initiate and maintain the local inflammatory response and tissue injury [2,5,43-46].
It is generally assumed that biological functions of DCs are associated with their homing, which are mediated and modulated by different time-dependent adhesion molecules and chemokines and their interactions[16,19,21,35,47-50]. Our results of P-selectin and ICAM-1 expression at high levels within the liver and kidney were consistent with CD1a+CD80+DC distribution in hepatic sinusoids and renal tubular interstitium and tissue injury. However, we showed that P-selectin and ICAM-1 expressions were inhibited, CD1a+CD80+DC populations were reduced and the injury was attenuated in hepatic and renal tissue in anti-PsL-EGFmAb-treated group. These results support the hypothesis that DC infiltration and accumulation is related to P-selectin and ICAM-1 expression and mediation, and suggest that P-selectin and ICAM-1 adhesion cascade mediate DC migration into hepatic and renal tissues, initiating immune inflammation in hepatic/renal ischemia-reperfusion injury. Furthermore, blockade of P-selectin and ICAM-1 inhibits the migration of the DCs of adhesion cascade to the hepatic and renal tissues, and thus attenuates hepatic and renal pathological damage[48,51,52].
In conclusion, DCs play an important role in immune pathogenesis of hepatic/renal ischemia-reperfusion injury. Anti-PsL-EGFmAb may regulate and inhibit local DC immigration and accumulation in liver/kidney. These findings may be important to further elucidate the mechanisms of hepatic/renal ischemia-reperfusion injury and to explore an efficient approach of anti-adhesive intervention.
Footnotes
Supported by Grants from the National Natural Science Foundation of China, No. 39970340, the Scientific Fund of the Chinese Ministry of Health, 98-2-283 and the Natural Science Foundation of Shanghai, No. 02ZB14041 and 034119916
References
- 1.Farrar CA, Wang Y, Sacks SH, Zhou W. Independent pathways of P-selectin and complement-mediated renal ischemia/reperfusion injury. Am J Pathol. 2004;164:133–141. doi: 10.1016/S0002-9440(10)63104-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Burne MJ, Daniels F, El Ghandour A, Mauiyyedi S, Colvin RB, O'Donnell MP, Rabb H. Identification of the CD4(+) T cell as a major pathogenic factor in ischemic acute renal failure. J Clin Invest. 2001;108:1283–1290. doi: 10.1172/JCI12080. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Burne MJ, Rabb H. Pathophysiological contributions of fucosyltransferases in renal ischemia reperfusion injury. J Immunol. 2002;169:2648–2652. doi: 10.4049/jimmunol.169.5.2648. [DOI] [PubMed] [Google Scholar]
- 4.Burne MJ, Haq M, Matsuse H, Mohapatra S, Rabb H. Genetic susceptibility to renal ischemia reperfusion injury revealed in a murine model. Transplantation. 2000;69:1023–1025. doi: 10.1097/00007890-200003150-00065. [DOI] [PubMed] [Google Scholar]
- 5.Park P, Haas M, Cunningham PN, Bao L, Alexander JJ, Quigg RJ. Injury in renal ischemia-reperfusion is independent from immunoglobulins and T lymphocytes. Am J Physiol Renal Physiol. 2002;282:F352–F357. doi: 10.1152/ajprenal.00160.2001. [DOI] [PubMed] [Google Scholar]
- 6.Carden DL, Granger DN. Pathophysiology of ischaemia-reperfusion injury. J Pathol. 2000;190:255–266. doi: 10.1002/(SICI)1096-9896(200002)190:3<255::AID-PATH526>3.0.CO;2-6. [DOI] [PubMed] [Google Scholar]
- 7.Kojima Y, Suzuki S, Tsuchiya Y, Konno H, Baba S, Nakamura S. Regulation of pro-inflammatory and anti-inflammatory cytokine responses by Kupffer cells in endotoxin-enhanced reperfusion injury after total hepatic ischemia. Transpl Int. 2003;16:231–240. doi: 10.1007/s00147-002-0536-4. [DOI] [PubMed] [Google Scholar]
- 8.Ke B, Shen XD, Lassman CR, Gao F, Katori M, Busuttil RW, Kupiec-Weglinski JW. Interleukin-13 gene transfer protects rat livers from antigen-independent injury induced by ischemia and reperfusion. Transplantation. 2003;75:1118–1123. doi: 10.1097/01.TP.0000062861.80771.D5. [DOI] [PubMed] [Google Scholar]
- 9.Krieglstein CF, Granger DN. Adhesion molecules and their role in vascular disease. Am J Hypertens. 2001;14:44S–54S. doi: 10.1016/s0895-7061(01)02069-6. [DOI] [PubMed] [Google Scholar]
- 10.Tailor A, Granger DN. Role of adhesion molecules in vascular regulation and damage. Curr Hypertens Rep. 2000;2:78–83. doi: 10.1007/s11906-000-0063-6. [DOI] [PubMed] [Google Scholar]
- 11.Lee HT, Xu H, Nasr SH, Schnermann J, Emala CW. A1 adenosine receptor knockout mice exhibit increased renal injury following ischemia and reperfusion. Am J Physiol Renal Physiol. 2004;286:F298–F306. doi: 10.1152/ajprenal.00185.2003. [DOI] [PubMed] [Google Scholar]
- 12.Zhou T. Adhesion molecules and multiorgan injury. Shanghai Mianyixue Zazhi. 1999;19:196–198. [Google Scholar]
- 13.Kokura S, Wolf RE, Yoshikawa T, Ichikawa H, Granger DN, Aw TY. Endothelial cells exposed to anoxia/reoxygenation are hyperadhesive to T-lymphocytes: kinetics and molecular mechanisms. Microcirculation. 2000;7:13–23. [PubMed] [Google Scholar]
- 14.Wang F, Song W, Zhou T. Cell adhesion and the evolvement of anti-adhesive treatment. Xibao Yu Fenzi Mianyixue Zazhi. 2001;17:624–625. [Google Scholar]
- 15.Park KM, Kramers C, Vayssier-Taussat M, Chen A, Bonventre JV. Prevention of kidney ischemia/reperfusion-induced functional injury, MAPK and MAPK kinase activation, and inflammation by remote transient ureteral obstruction. J Biol Chem. 2002;277:2040–2049. doi: 10.1074/jbc.M107525200. [DOI] [PubMed] [Google Scholar]
- 16.Sun GZ, Wu KY, Zhou T, Zhang DQ. Dendritic cells and kidney. Shanghai Mianyixue Zazhi. 2003;23:424–426. [Google Scholar]
- 17.Heimberger AB, Crotty LE, Archer GE, McLendon RE, Friedman A, Dranoff G, Bigner DD, Sampson JH. Bone marrow-derived dendritic cells pulsed with tumor homogenate induce immunity against syngeneic intracerebral glioma. J Neuroimmunol. 2000;103:16–25. doi: 10.1016/s0165-5728(99)00172-1. [DOI] [PubMed] [Google Scholar]
- 18.Ni HT, Spellman SR, Jean WC, Hall WA, Low WC. Immunization with dendritic cells pulsed with tumor extract increases survival of mice bearing intracranial gliomas. J Neurooncol. 2001;51:1–9. doi: 10.1023/a:1006452726391. [DOI] [PubMed] [Google Scholar]
- 19.Pendl GG, Robert C, Steinert M, Thanos R, Eytner R, Borges E, Wild MK, Lowe JB, Fuhlbrigge RC, Kupper TS, et al. Immature mouse dendritic cells enter inflamed tissue, a process that requires E- and P-selectin, but not P-selectin glycoprotein ligand 1. Blood. 2002;99:946–956. doi: 10.1182/blood.v99.3.946. [DOI] [PubMed] [Google Scholar]
- 20.Bondanza A, Zimmermann VS, Dell'Antonio G, Dal Cin E, Capobianco A, Sabbadini MG, Manfredi AA, Rovere-Querini P. Cutting edge: dissociation between autoimmune response and clinical disease after vaccination with dendritic cells. J Immunol. 2003;170:24–27. doi: 10.4049/jimmunol.170.1.24. [DOI] [PubMed] [Google Scholar]
- 21.Liu W, Zhou T, Shi H, Sun GZ, Zhang DQ, Chen N. Cell adhesion and the mechanisms of dendritic cells immigration. Shengming Kexue. 2002;14:379–382. [Google Scholar]
- 22.Zhou T, Li X, Wu P, Zhang D, Zhang M, Chen N, Dong D. Effect of anti-P-selectin monoclonal antibody on renal ischemia/reperfusion injury in rats. Chin Med J (Engl) 2000;113:790–793. [PubMed] [Google Scholar]
- 23.Wu P, Li X, Zhou T, Zhang MJ, Chen JL, Wang WM, Chen N, Dong DC. Role of P-selectin and anti-P-selectin monoclonal antibody in apoptosis during hepatic/renal ischemia reperfusion injury. World J Gastroenterol. 2000;6:244–247. doi: 10.3748/wjg.v6.i2.244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Zhou T, Song W, Wang F, Ni PH, Chen N, Zhang DQ, Yu QW. Cloning, expression of the lectin-EGF domain of P-selectin, and preparation of its monoclonal antibody. Shengwu Huaxue Yu Shengwu Wuli Xuebao (Shanghai) 2003;35:172–176. [PubMed] [Google Scholar]
- 25.Zhou T, Li X, Sun GZ, Wu KY, Hu QS, Chen YY, Zhang DQ, Chen N. Localization of dendritic cells in human kidney with nephritis. Zhongguo Weixunhuan. 2003;7:208 210. [Google Scholar]
- 26.Maroszyńska I, Fiedor P. Leukocytes and endothelium interaction as rate limiting step in the inflammatory response and a key factor in the ischemia-reperfusion injury. Ann Transplant. 2000;5:5–11. [PubMed] [Google Scholar]
- 27.Tailor A, Granger DN. Hypercholesterolemia promotes P-selectin-dependent platelet-endothelial cell adhesion in postcapillary venules. Arterioscler Thromb Vasc Biol. 2003;23:675–680. doi: 10.1161/01.ATV.0000056742.97580.79. [DOI] [PubMed] [Google Scholar]
- 28.Young CS, Palma JM, Mosher BD, Harkema J, Naylor DF, Dean RE, Crockett E. Hepatic ischemia/reperfusion injury in P-selectin and intercellular adhesion molecule-1 double-mutant mice. Am Surg. 2001;67:737–744. [PubMed] [Google Scholar]
- 29.Zhou T, Wang F, Sun GZ, Zhang YM. The glycobiological evolvement of adhesion molecules and cell adhesion. Zhongguo Weixunhuan. 2003;7:187–190. [Google Scholar]
- 30.Singbartl K, Green SA, Ley K. Blocking P-selectin protects from ischemia/reperfusion-induced acute renal failure. FASEB J. 2000;14:48–54. doi: 10.1096/fasebj.14.1.48. [DOI] [PubMed] [Google Scholar]
- 31.Cutrn JC, Perrelli MG, Cavalieri B, Peralta C, Rosell Catafau J, Poli G. Microvascular dysfunction induced by reperfusion injury and protective effect of ischemic preconditioning. Free Radic Biol Med. 2002;33:1200–1208. doi: 10.1016/s0891-5849(02)01017-1. [DOI] [PubMed] [Google Scholar]
- 32.Laude K, Beauchamp P, Thuillez C, Richard V. Endothelial protective effects of preconditioning. Cardiovasc Res. 2002;55:466–473. doi: 10.1016/s0008-6363(02)00277-8. [DOI] [PubMed] [Google Scholar]
- 33.Lee HT, Emala CW. Protective effects of renal ischemic preconditioning and adenosine pretreatment: role of A(1) and A(3) receptors. Am J Physiol Renal Physiol. 2000;278:F380–F387. doi: 10.1152/ajprenal.2000.278.3.F380. [DOI] [PubMed] [Google Scholar]
- 34.Forlow SB, Ley K. Selectin-independent leukocyte rolling and adhesion in mice deficient in E-, P-, and L-selectin and ICAM-1. Am J Physiol Heart Circ Physiol. 2001;280:H634–H641. doi: 10.1152/ajpheart.2001.280.2.H634. [DOI] [PubMed] [Google Scholar]
- 35.van den Broeke LT, Daschbach E, Thomas EK, Andringa G, Berzofsky JA. Dendritic cell-induced activation of adaptive and innate antitumor immunity. J Immunol. 2003;171:5842–5852. doi: 10.4049/jimmunol.171.11.5842. [DOI] [PubMed] [Google Scholar]
- 36.Burne-Taney MJ, Kofler J, Yokota N, Weisfeldt M, Traystman RJ, Rabb H. Acute renal failure after whole body ischemia is characterized by inflammation and T cell-mediated injury. Am J Physiol Renal Physiol. 2003;285:F87–F94. doi: 10.1152/ajprenal.00026.2003. [DOI] [PubMed] [Google Scholar]
- 37.Nishiwaki A, Ueda T, Ugawa S, Shimada S, Ogura Y. Upregulation of P-selectin and intercellular adhesion molecule-1 after retinal ischemia-reperfusion injury. Invest Ophthalmol Vis Sci. 2003;44:4931–4935. doi: 10.1167/iovs.02-1324. [DOI] [PubMed] [Google Scholar]
- 38.Khandoga A, Biberthaler P, Enders G, Teupser D, Axmann S, Luchting B, Hutter J, Messmer K, Krombach F. P-selectin mediates platelet-endothelial cell interactions and reperfusion injury in the mouse liver in vivo. Shock. 2002;18:529–535. doi: 10.1097/00024382-200212000-00008. [DOI] [PubMed] [Google Scholar]
- 39.Khandoga A, Biberthaler P, Messmer K, Krombach F. Platelet-endothelial cell interactions during hepatic ischemia-reperfusion in vivo: a systematic analysis. Microvasc Res. 2003;65:71–77. doi: 10.1016/s0026-2862(02)00018-3. [DOI] [PubMed] [Google Scholar]
- 40.Khandoga A, Biberthaler P, Enders G, Axmann S, Hutter J, Messmer K, Krombach F. Platelet adhesion mediated by fibrinogen-intercelllular adhesion molecule-1 binding induces tissue injury in the postischemic liver in vivo. Transplantation. 2002;74:681–688. doi: 10.1097/00007890-200209150-00016. [DOI] [PubMed] [Google Scholar]
- 41.Ishikawa M, Cooper D, Russell J, Salter JW, Zhang JH, Nanda A, Granger DN. Molecular determinants of the prothrombogenic and inflammatory phenotype assumed by the postischemic cerebral microcirculation. Stroke. 2003;34:1777–1782. doi: 10.1161/01.STR.0000074921.17767.F2. [DOI] [PubMed] [Google Scholar]
- 42.Thurman JM, Ljubanovic D, Edelstein CL, Gilkeson GS, Holers VM. Lack of a functional alternative complement pathway ameliorates ischemic acute renal failure in mice. J Immunol. 2003;170:1517–1523. doi: 10.4049/jimmunol.170.3.1517. [DOI] [PubMed] [Google Scholar]
- 43.Rabb H, Daniels F, O'Donnell M, Haq M, Saba SR, Keane W, Tang WW. Pathophysiological role of T lymphocytes in renal ischemia-reperfusion injury in mice. Am J Physiol Renal Physiol. 2000;279:F525–F531. doi: 10.1152/ajprenal.2000.279.3.F525. [DOI] [PubMed] [Google Scholar]
- 44.Yokota N, Burne-Taney M, Racusen L, Rabb H. Contrasting roles for STAT4 and STAT6 signal transduction pathways in murine renal ischemia-reperfusion injury. Am J Physiol Renal Physiol. 2003;285:F319–F325. doi: 10.1152/ajprenal.00432.2002. [DOI] [PubMed] [Google Scholar]
- 45.Burne MJ, Elghandour A, Haq M, Saba SR, Norman J, Condon T, Bennett F, Rabb H. IL-1 and TNF independent pathways mediate ICAM-1/VCAM-1 up-regulation in ischemia reperfusion injury. J Leukoc Biol. 2001;70:192–198. [PubMed] [Google Scholar]
- 46.Le Moine O, Louis H, Demols A, Desalle F, Demoor F, Quertinmont E, Goldman M, Devière J. Cold liver ischemia-reperfusion injury critically depends on liver T cells and is improved by donor pretreatment with interleukin 10 in mice. Hepatology. 2000;31:1266–1274. doi: 10.1053/jhep.2000.7881. [DOI] [PubMed] [Google Scholar]
- 47.Hirata T, Furie BC, Furie B. P-, E-, and L-selectin mediate migration of activated CD8+ T lymphocytes into inflamed skin. J Immunol. 2002;169:4307–4313. doi: 10.4049/jimmunol.169.8.4307. [DOI] [PubMed] [Google Scholar]
- 48.Yanaba K, Kaburagi Y, Takehara K, Steeber DA, Tedder TF, Sato S. Relative contributions of selectins and intercellular adhesion molecule-1 to tissue injury induced by immune complex deposition. Am J Pathol. 2003;162:1463–1473. doi: 10.1016/S0002-9440(10)64279-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Martinez-Mier G, Toledo-Pereyra LH, McDuffie JE, Warner RL, Ward PA. P-selectin and chemokine response after liver ischemia and reperfusion. J Am Coll Surg. 2000;191:395–402. doi: 10.1016/s1072-7515(00)00360-4. [DOI] [PubMed] [Google Scholar]
- 50.Briaud SA, Ding ZM, Michael LH, Entman ML, Daniel S, Ballantyne CM. Leukocyte trafficking and myocardial reperfusion injury in ICAM-1/P-selectin-knockout mice. Am J Physiol Heart Circ Physiol. 2001;280:H60–H67. doi: 10.1152/ajpheart.2001.280.1.H60. [DOI] [PubMed] [Google Scholar]
- 51.Lu YT, Chen PG, Liu SF. Time course of lung ischemia-reperfusion-induced ICAM-1 expression and its role in ischemia-reperfusion lung injury. J Appl Physiol (1985) 2002;93:620–628. doi: 10.1152/japplphysiol.01200.2001. [DOI] [PubMed] [Google Scholar]
- 52.Bertuglia S, Giusti A. Blockade of P-selectin does not affect reperfusion injury in hamsters subjected to glutathione inhibition. Microvasc Res. 2002;64:56–64. doi: 10.1006/mvre.2002.2400. [DOI] [PubMed] [Google Scholar]