

Abstract
Ongoing climate change will lead to more extreme weather events, including severe drought periods and intense drying rewetting cycles. This will directly influence microbial nitrogen (N) turnover rates in soil by changing the water content and the oxygen partial pressure. Therefore, a space for time climate change experiment was conducted by transferring intact beech seedling-soil mesocosms from a northwest (NW) exposed site, representing today's climatic conditions, to a southwest (SW) exposed site, providing a model climate for future conditions with naturally occurring increased soil temperature (+0.8°C in average). In addition, severe drought and intense rainfall was simulated by a rainout shelter at SW and manual rewetting after 39 days drought, respectively. Soil samples were taken in June, at the end of the drought period (August), 24 and 72 hours after rewetting (August) and after a regeneration period of four weeks (September). To follow dynamics of bacterial and archaeal communities involved in N turnover, abundance and activity of nitrifiers, denitrifiers, N2-fixing microbes and N-mineralizers was analyzed based on marker genes and the related transcripts by qPCR from DNA and RNA directly extracted from soil. Abundance of the transcripts was reduced under climate change with most pronounced effects for denitrification. Our results revealed that already a transfer from NW to SW without further treatment resulted in decreased cnor and nosZ transcripts, encoding for nitric oxide reductase and nitrous oxide reductase, respectively, while nirK transcripts, encoding for nitrite reductase, remained unaffected. Severe drought additionally led to reduced nirK and cnor transcripts at SW. After rewetting, nirK transcripts increased rapidly at both sites, while cnor and nosZ transcripts increased only at NW. Our data indicate that the climate change influences activity pattern of microbial communities involved in denitrification processes to a different extend, which may impact emission rates of the greenhouse gas N2O.
Introduction
Beech (Fagus sylvatica L.) dominates the natural forest vegetation from moderate dry to moist areas of sub-mountainous altitude in Central Europe [1]. It often grows on calcareous, limestone derived soils which have low water retention capacity and are poor in bioavailable nitrogen (N) [2], [3]. However, N is an essential component of proteins, nucleotides, coenzymes, photosynthetic pigments, secondary metabolites and other molecules, and is one of the major growth limiting factors for plants [4]. Consequently, productivity of forest ecosystems is strongly dependent on an efficient microbial N turnover characterized by low N losses via denitrification or leaching and rapid recycling of organically bound nutrients by mineralization [5]. Furthermore, in N-limited ecosystems strong competition between microorganisms and plants for N exists, leading to a fragile balance of microbial N mineralization and N immobilization by microbes and plants, which is determined mainly by abiotic factors, including soil temperature, oxygen partial pressure, pH and nitrogen and carbon (C) availability [6], [7]. Consequently, altered environmental conditions induced by global change may lead to a significant disturbance of unfertilized forest ecosystems. Mainly an increase in extreme weather events, including a higher frequency of intense precipitation and thus rapidly changing soil moisture regimes, extreme temperature events, heat waves and an increase in frequency and duration of drought periods [8] might affect the interplay of soil biota and plants in the future.
It is well accepted that drought decreases microbial activity and biomass due to osmotic regulation (accumulation of compatible solutes in cells to avoid dehydration), limited diffusive transport of substrates and extracellular enzymes and decreased microbial motility [9]. Furthermore, root biomass and consequently root exudates are reduced [10], resulting in additional C limitation for soil microorganisms. However, increased oxygen content due to lower soil moisture in combination with reduced competition for N due to reduced plant performance might lead to enhanced nitrification, which is an aerobic process performed by autotrophic bacteria and archaea independent from labile C sources [11]. Increased nitrification will lead to nitrate formation, which may leach to the ground water depending on soil type, reducing the available N pools in soil to a large extend.
Rewetting of dry soils induces additional osmotic stress for microbes, resulting in release of cytoplasmic solutes to avoid bursting [12]. However, microbial activity was shown to increase within minutes [13] or hours [14] after rewetting, mainly due to reconstituting mineralization of both newly exposed organic matter and dead microbial cells accumulated during drought [15]. This nutrient flush in combination with reduced oxygen levels due to high soil moisture leads to enhanced denitrification activities [16], [17] and thus increased N losses from the ecosystem. Nevertheless, the response of microbial processes to environmental stress conditions is not only related to soil moisture and the availability of nutrients and organic matter, but also to changes in microbial community structure [16], [17]. Recent studies on the dynamics of microbes involved in N turnover under different environmental stress factors provide contradictory results. When investigating the influence of temperature and water content on bacterial, archaeal and denitrifying microbial communities, Stres et al. [18] found no significant response of bacterial and denitrifier abundance and community structure, while archaeal communities were strongly influenced by temperature. In contrast, denitrifying communities in a pristine forest soil were highly affected by water content, and both archaeal ammonia oxidizers (AOA) and their bacterial counterpart (AOB) responded to different temperatures and water regimes [19]. These contradicting results indicate that the effects of climate change conditions on microbial communities and their functional traits are still poorly understood and cannot be easily transferred from one ecosystem to another.
Whereas most studies so far investigated either the effects of drought or intensive rainfall on soil microbial communities, data on extreme drought rewetting cycles are still rare, despite their relevance for future climate change scenarios [8]. Thus, in the frame of this project we performed a transplant experiment where intact beech seedling-soil mesocosms from a northwest (NW) exposed site, representing today's climatic conditions, were transferred to a site of southwest (SW) exposure, serving as model for changed climatic conditions. Additionally, a rainout shelter was established at SW to simulate severe drought. After 39 days drought, intense precipitation was simulated by manual rewetting of the mesocosms at both NW and SW site. Soil samples were taken in June (before the rainout shelter was established), at the end of the drought period (August), 24 and 72 hours after rewetting (August) and at the end of the beech vegetation period in September.
In contrast to previous studies investigating climate change effects on soil microflora focusing on shifts in microbial community structure [16], [17] or on effects on single N transformation steps [18], [19], in the present study for the first time consequences for activity pattern of N2-fixers, nitrifiers, denitrifiers and microbes involved in N-mineralization were investigated in addition to shifts in abundance of bacterial and archaeal communities involved in N cycling. Therefore, the abundance of marker genes of prokaryotes encoding for enzymes catalyzing key processes in N turnover as well as their transcripts were quantified using quantitative real-time PCR (qPCR) based on extracted DNA and mRNA from soil (N2 fixation (nifH), nitrification (amoA) and denitrification (nirK, nirS, cnor and nosZ)). The second known gene encoding for nitric oxide reductase, qnor, was not considered, as it was also found in many non-denitrifying microorganisms, suggesting a function in detoxifying NO [20]. From the large pool of genes encoding for N mineralization enzymes alkaline metalloprotease (apr) and chitinase (chiA) were chosen as marker genes in this study. In temperate forest ecosystems, most tree species form ectomycorrhizal fungal (EMF) associations, and especially beech roots are usually 100% colonized by EMF [21], resulting in a high soil chitin concentration. Proteins comprise up to 40% of the total soil N [22], and there is evidence that proteolysis could be the rate-limiting step in soil organic N mineralization [23]. Hence, the hydrolysis of proteins and chitin is particularly important to supply the soil N pool with plant available N. Furthermore, it was shown that the abundance of chiA and apr genes correlates positively with chitinolytic and proteolytic enzyme activities [24] and that both chitinase and protease activity correlates positively with N mineralization rate [23], [25].
Materials and Methods
Study site description
Permits for performing the experiment were issued by the Landratsamt Tuttlingen, Germany. The experiment was conducted at the Tuttlingen Research Station, a long term ecological beech research forest site in the Swabian Jura, a low mountain range in Southern Germany (8°45′E/47°59′N) at an altitude of 800 m above sea level with an atmospheric N deposition of <10 kg N ha−1 year−1 [26]. The soil was classified as Rendzic Leptosoil (Sceletic) according to the IUSS [27] with the following characteristics: 68% clay, 28% silt, 4% sand, pH 6.0 (measured in 0.01 M CaCl2), 0.5% total nitrogen content, 6.1% organic carbon content, 0.05% inorganic carbon content and a maximum water holding capacity (WHC) of 118% [28]. To simulate climate change, two sites with opposing exposure (NW and SW) of a narrow valley within a distance <1 km of each other were selected as model ecosystems. At both sites, beech was the dominant species and trees showed similar population genetics [29]. The NW site showed a cool-moist climate representing present climatic conditions typical of a beech stand in Central Europe with mean annual air temperature of 6.6°C, mean air temperature during growing season (April to October) of 11.5°C, mean annual precipitation of 810 mm and mean precipitation during growing season of 86 mm [30]. The SW site was characterized by higher radiation, resulting in increased daily maximum of air and soil temperature and thus reduced water availability [31], representing future climatic conditions [8]. During the experimental period, mean soil temperature/volumetric soil moisture at NW and SW were 10.3°C/0.26 m3 m−3 and 11.1°C/0.23 m3 m−3, respectively [28].
Experimental design
This study was part of a transplant experiment investigating N turnover in European beech forests under climate change conditions. The transfer of the intact plant-soil mesocosms was described in detail by Bimueller et al. [28]. In brief, stainless steel cylinders (diameter 16.8 cm, height 15 cm) were used to sample 80 soil cores centered around a beech seedling of 2 mm stem diameter and 30 cm height without disturbing the root architecture. Sampling was done in July 2010 on a preselected sampling area on the NW site of the beech forest, where intensive drilling had revealed similar soil profiles with weathered bedrock occurring at > 15–20 cm depth. Forty randomly selected mesocosms were transferred from the NW to the SW site. The remaining 40 mesocosms were transferred within the NW site at the same elevation, representing in situ controls. After transfer, all plant-soil mesocosms were irrigated with 500 ml water within two hours (corresponding to a typical summer precipitation event of 23.7 l m−2) to avoid drying or death of enclosed beech seedlings after transfer.
After a pre-incubation period of 11 months for acclimatization, eight intact mesocosms were sampled at both NW and SW on 22nd June 2011 (Sampling T1, representing ambient climate change). In order to simulate intensive summer drought, a translucent rain sheltering roof was established at SW from 27th June 2011 to 9th August 2011. Trenches around the 1 m tall rain shelter avoided influence of slope water. After 39 days of enhanced drought at SW, eight mesocosms from both NW and SW were sampled on 2nd August 2011 (Sampling T2, representing ambient/intensive summer drought). For simulation of a severe precipitation event, the remaining plant-soil mesocosms of both sites were irrigated three times with 280 ml water each within two hours on 5th August 2011. On 6th August and 8th August 2011, respectively, eight mesocosms were sampled at both NW and SW (Sampling T3 and T4, representing intensive precipitation). After deconstruction of the rainout shelter at SW on 9th August 2011, the remaining mesocosms were left under ambient climate conditions at NW and SW, respectively, until the final sampling on 27th September 2011 (Sampling T5).
Sampling of the plant-soil mesocosms
For each sampling time, eight plant-soil mesocosms at both NW and SW were taken and treated as independent replicates. The intact mesocosms were carefully excavated by hand and processed within two hours after excavation. The beech seedlings were cut and separated into leaves and stems. The remaining soil-root mesocosms were manually separated into soil, gravel, dead organic material and living roots, which were washed with tap water to remove adhering soil. The plant parts were dried at 60°C for 48 hours for determination of dry biomass, which was significantly reduced at SW compared to NW for both above- and belowground plant biomass (Table S1).
The remaining mesocosm soil was homogenized by manual mixing for 10 min. A soil subsample was immediately frozen on dry ice and stored at −80°C for DNA/RNA extraction. A second soil subsample was freeze-dried for determination of carbon and nitrogen content. A third subsample of soil (100 g fresh weight) was immediately extracted 1:1.5 (soil:solution) with 0.5 M K2SO4 on a rotary shaker for one hour. Afterwards, extracts were vacuum filtered using pumps and glass fibre filters [2] and stored at −20°C for analysis of extractable soil C and N pools.
Total soil carbon and nitrogen content
For analysis of soil C and N content, the freeze-dried soil samples were ball-milled (Retsch MM2, Retsch GmbH, Haan, Germany), weighed into tin capsules (1.5 mg soil per capsule) and analyzed in duplicates using an Elemental Analyzer ‘Euro-EA’ (Eurovector, Milano, Italy).
Extractable soil carbon and nitrogen pools
Extractable soil C and N pools were analyzed as described by Dannenmann et al. [2]. For determination of ammonium and nitrate concentrations, a subsample of the filtered extracts was analyzed colorimetrically by a commercial laboratory (Dr. Janssen, Gillersheim, Germany). Total organic carbon (TOC) and total nitrogen (TN) in the extracts was quantified on an Infrared TOC analyzer with a coupled chemoluminescence-based total N module (DIMATEC GmbH, Germany). TOC concentrations of extracts were referred to as extractable dissolved organic carbon (DOC), while extractable dissolved organic nitrogen (DON) was calculated as the difference between TN and dissolved inorganic N (ammonium plus nitrate).
Nucleic acid extraction
DNA and RNA from 0.4 g homogenized soil were co-extracted using the protocol described by Lueders [32] and the Precellys24 Instrument (PeqLab, Erlangen, Germany). Quality and quantity of the extracted nucleic acids were checked with a spectrophotometer (Nanodrop, PeqLab, Erlangen, Germany) and gel electrophoresis. Afterwards, the extract was divided into two subsamples. One was used for DNA analysis without further treatment. The second subsample was used to prepare RNA by digestion of co-extracted DNA with RNase free DNase I (Promega, Mannheim, Germany) according to manufacturer's instructions and subsequent purification using 3 M RNase free sodium acetate (pH 5.2) and isopropanol. Quality and quantity of the extracted RNA were checked with a spectrophotometer (Nanodrop, PeqLab, Erlangen, Germany) and gel electrophoresis. The absence of DNA was confirmed by performing PCR targeting the 16S rRNA gene using universal primers 968f (5′- AAC GCG AAG AAC CTT AC -3′) and 1401r (5′- CGG TGT GTA CAA GAC CC -3′) and the amplification protocol described by Schreiner et al. [33]. Afterwards, cDNA was synthesized using the ‘High capacity cDNA reverse transcription kit’ (Life Technologies, Darmstadt, Germany) according to manufacturer's instructions. The success of cDNA synthesis was confirmed by performing PCR targeting the 16S rRNA gene as described above. Both DNA and cDNA extracts were stored at −20°C until use.
Real-time PCR assay
Quantitative real-time PCR (qPCR) was performed using an ABI 7300 Cycler (Life Technologies, Darmstadt, Germany) with the following assay reagents: dimethyl sulfoxid (DMSO) and bovine serum albumin (BSA) (Sigma, Germany), primers listed in Table 1 (Metabion, Martinsried, Germany) and 2x Power SYBR Green master mix (Life Technologies, Darmstadt, Germany). The respective reaction mixtures (total volume 25 µl) for quantification of the genes listed in Table 1 consisted of: 12.5 µl SYBR Green master mix, 5 pmol of each primer (for apr gene: 10 pmol of each primer), 0.5 µl 3% BSA and 2 µl DNA template. For the amplification of nirS and nirK genes, 0.5 µl DMSO was added additionally.
Table 1. Thermal profiles and primer used for real-time PCR quantification of different functional genes and transcripts.
| Target gene | Source of standard | Primer | References | Thermal profile | No. of cycles |
| nifH | Azospirillum irakense | nifH-f, nifH-r | [63] | 95°C-45s/55°C-45s/72°C-45s | 40 |
| amoA AOA | Nitrosomonas europaea | amo19F, CrenamoA16r48x | [49], [64] | 94°C-45s/55°C-45s/72°C-45s | 40 |
| amoA AOB | Fosmid clone 54d9 | amoA1F, amoA2R | [65] | 94°C-45s/59°C-45s/72°C-45s | 40 |
| nirS | Pseudomonas stutzeri | cd3aF, R3cd | [66], [67] | 95°C-45s/57°C-45s/72°C-45s | 40 |
| nirK | Azospirillum irakense | nirK876, nirK5R | [68], [69] | 95°C-15s/63-58°C-30s/72°C-30s | 5 a |
| 95°C-15s/58°C-30s/72°C-30s | 40 | ||||
| cnor | Sinorhizobium meliloti | cnorB2f, cnorB6r | [20] | 95°C-15s/60-55°C-30s/72°C-30s | 5 a |
| 95°C-15s/55°C-30s/72°C-30s | 40 | ||||
| nosZ | Pseudomonas stutzeri | nosZ2F, nosZ2R | [70] | 95°C-15s/65-60°C-30s/72°C-30s | 5 a |
| 95°C-15s/60°C-30s/72°C-30s | 40 | ||||
| chiA | Streptomyces griseus | chiF2, chiR | [71] | 95°C-30s/60°C-30s/72°C-60s | 40 |
| apr | Pseudomonas aeruginosa | FPapr1, RPapr2 | [72] | 95°C-20s/53°C-30s/72°C-60s | 40 |
Touchdown: −1°C per cycle.
For quantification, standard curves were calculated using serial dilutions (101 to 106 gene copies μl−1) of plasmid DNA containing PCR products of the respective genes listed in Table 1. PCR detection limit was assessed to 1 gene copy μl−1. In order to prevent PCR inhibition, the optimal dilution for each amplification assay was determined by dilution series of randomly chosen DNA and cDNA extracts in advance (data not shown). The qPCR assays were performed in 96-well plates (Life Technologies, Darmstadt, Germany) for all target genes as described in Table 1. All PCR runs started with a hot start at 95°C for 10 minutes. After each run, the specifity of the SYBR Green-quantified amplicons was checked by melting curve analysis and gel electrophoresis. The amplification efficiency was calculated from the formula Eff = [10(−1/slope)−1] and resulted in the following average efficiencies (standard deviation less than 5% of mean): nifH, 86%, chiA, 88%, apr, 89%, amoA ammonium-oxidizing archaea (AOA), 94%, amoA ammonium-oxidizing bacteria (AOB), 91%, nirK, 98%, nirS, 91%, cnor, 93%, and nosZ, 89%.
Statistical analysis
Statistical analysis was performed using SPSS 11.5 (SPSS, Inc.). Data was evaluated by multivariate analysis of variance (ANOVA) at the significance level P <0.05. The normal distribution of the data was checked by the Kolmogorov-Smirnov test and histograms. If necessary, the data was log-transformed prior to analysis. The homogeneity of the variances was checked by the Levene test. For the pairwise comparison of means with the ANOVA, either the Tukey test or, if the homogeneity of the variances was not given, the Games-Howell test was used. In order to test significant effects between the sites at each time point, Student's T-test for independent samples was performed.
Results
Soil moisture
Mean values of gravimetric soil water content normalized to WHC are presented in Table 2. As expected, transferring plant-soil mesocosms from NW to SW resulted in a decrease in soil moisture from 63.5% WHC to 54.4% WHC after 11 months of acclimatization (T1). This reduced soil water content at SW compared to NW was observed during the whole sampling period and was especially pronounced at T2, where roof-intensified drought at SW reduced soil moisture to 44.5% WHC, while soil moisture at NW was not significantly changed by ambient summer drought (57.2% WHC). Simulated precipitation increased the soil water content to 62.3% WHC (NW, P > 0.05) and 53.0% WHC (SW), respectively. With ongoing season, soil moisture remained constant at both sites (on average 60.6% WHC at NW and 49.8% WHC at SW).
Table 2. Gravimetric soil moisture related to water holding capacity (WHC), total N and C contents as well as extractable N and C pools of soil of soils at NW and SW, sampled in June (T1), after 39 days drought in August (T2), 24 and 72 hours after rewetting in August (T3, T4) and in September (T5) (n = 8, standard deviation of the mean in parentheses).
| NW | SW | P site | P time | |||||||||
| T1 | T2 | T3 | T4 | T5 | T1 | T2 | T3 | T4 | T5 | |||
| Water content | 63.5 (4.0) * | 57.2 (8.6) * | 62.3 (7.0) * | 61.0 (11.0) * | 60.3 (10.4) * | 54.4 (7.0) * | 44.5 (6.4) * | 53.0 (9.1) * | 48.8 (6.5) * | 50.7 (10.6) * | 0.000 | 0.021 |
| % WHC | a | a | a | a | a | a | b | a | ab | ab | ||
| N total | 4.6 (0.3) | 4.3 (0.7) | 4.7 (0.8) | 4.3 (1.0) | 4.7 (1.0) | 4.4 (0.7) | 4.8 (1.0) | 5.0 (0.8) | 4.1 (1.1) | 4.8 (0.9) | 0.643 | 0.216 |
| mg g−1 sdw | a | a | a | a | a | a | a | a | a | a | ||
| NH4+ | 3.8 (1.8) | 3.3 (1.1) | 5.9 (1.0) | 3.9 (0.8) | 3.2 (1.6) | 3.5 (2.2) | 3.6 (1.0) | 6.0 (1.1) | 3.8 (1.5) | 3.1 (1.0) | 0.964 | 0.000 |
| μg g−1 sdw | a | a | b | a | a | a | a | b | a | a | ||
| NO3− | 4.5 (2.5) | 4.2 (2.5) | 3.1 (1.3) | 5.0 (2.8) | 0.3 (0.1) | 4.1 (2.9) | 4.3 (2.3) | 3.1 (0.9) | 5.1 (2.1) | 0.5 (0.3) | 0.976 | 0.000 |
| μg g−1 sdw | a | a | ab | a | b | ab | ab | ab | a | b | ||
| DON | 9.1 (3.5) | 1.0 (1.5) * | 2.3 (2.2) | 3.3 (2.5) | 5.9 (1.3) | 9.8 (3.6) | 5.4 (3.0) * | 5.0 (3.6) | 3.4 (1.8) | 3.8 (2.9) | 0.063 | 0.000 |
| μg g−1 sdw | a | b | bc | bc | c | a | ab | ab | b | b | ||
| C total | 58.6 (4.3) | 57.4 (9.6) | 60.7 (10.7) | 58.5 (15.8) | 64.6 (16.6) | 58.7 (10.8) | 60.7 (13.1) | 66.8 (8.2) | 59.6 (13.5) | 67.2 (11.8) | 0.322 | 0.305 |
| mg g−1 sdw | a | a | a | a | a | a | a | a | a | a | ||
| DOC | 72.2 (11.5) | 20.5 (7.1) | 51.5 (14.3) | 48.9 (17.2) | 52.2 (11.5) | 71.2 (13.8) | 23.6 (5.3) | 55.7 (12.7) | 57.4 (25.9) | 45.2 (20.6) | 0.654 | 0.000 |
| μg g−1 sdw | a | b | c | c | c | a | b | ac | ac | bc |
Asterisks indicate significant differences between NW and SW at the respective sampling times (Student's T test), whereas lower case letters indicate differences among the sampling period for the respective site (multivariate ANOVA). Significant differences between the factors site and sampling time calculated by multivariate ANOVA are indicated by P values <0.05 (bold letters).
Soil biochemical parameters
As expected, neither total soil N nor C contents differed between NW and SW or during sampling times and were 4.6 mg N g−1 soil dry weight (sdw) and 61.3 mg C g−1 sdw in average. Contrary, the measured labile pools (ammonium, nitrate, DON and DOC) were highly influenced by drought and rewetting with similar intensities and trends at NW and SW (except for DON). Therefore, data of NW at the respective time points were described as example in this result section (except for DON). Ammonium contents increased from 3.8 µg N g−1 sdw (NW, T1) to 5.9 µg N g−1 sdw (NW, T3) and declined again to 3.2 µg N g−1 sdw (NW, T5). Nitrate contents decreased from 4.5 µg N g−1 sdw (NW, T1) to 3.1 µg N g−1 sdw (NW, T3, P > 0.05), increased to 5.0 µg g−1 sdw (NW, T4, P > 0.05) and decreased to 0.3 µg N g−1 sdw (NW, T5). DON contents were similar between NW and SW at T1 (9.1 µg N g−1 sdw respective 9.8 µg N g−1 sdw), but while DON at SW declined continuously during sampling time to 3.8 µg g−1 sdw (T5), at NW the lowest DON value was observed at T2 (1.0 µg g−1 sdw) followed by a continuously increase to 5.9 µg g−1 sdw (T5). For DOC contents no difference between NW and SW was observed: DOC decreased from 72.2 µg C g−1 sdw (NW, T1) to 20.5 µg C g−1 sdw (NW, T2), increased after rewetting 51.5 µg C g−1 sdw (NW, T3) and remained constant hereafter. All data regarding soil biochemical parameters is summarized in Table 2.
Abundance of functional transcripts involved in nitrogen cycling
In general, the genetic potential (based on the abundance levels of the investigated functional groups) was not or not consistently affected by the investigated climate change scenario (Fig. S1). In contrast, transcription pattern showed a significant site effect for all quantified genes with lower transcript numbers at SW compared to NW in average (Fig. 1). Transcripts of nifH, amoA AOB and nirS genes were undetectable in all samples.
Figure 1. Transcript abundance of functional genes involved in the nitrogen cycle (chiA, apr, amoA AOA, nirK, cnor and nosZ) are shown for NW (black bar) and SW (grey bar) in June (T1), after 39 days drought in August (T2), 24 and 72 hours after rewetting in August (T3, T4) and in September (T5) (n = 8, error bars represent standard deviation of the mean).

Asterisks indicate significant differences between NW and SW at the respective sampling times (Student's T test), whereas lower case letters indicate differences among the sampling period for the respective site (multivariate ANOVA). Significant differences between the factors site and sampling time calculated by multivariate ANOVA are indicated by P values <0.05 (bold letters).
Concerning mineralization, transcript numbers of chiA were within the range of 105 transcripts g−1 sdw in average and thus 50 times lower compared to apr transcript abundance. Independent from the site, both chiA and apr transcript numbers were highest 24 hours after the simulated intense precipitation event (T3), with 2.3×105 transcripts g−1 sdw (NW) and 1.4×105 transcripts g−1 sdw (SW) for chiA respective 8.7×106 transcripts g−1 sdw (NW) and 7.3×106 transcripts g−1 sdw (SW) for apr. However, due to high variations among the replicates the increase was not significant.
In terms of ammonia oxidation, amoA transcripts of AOA dominated clearly over AOB, which were undetectable in all samples. On NW, AOA transcript numbers remained constant over time (7.9×104 transcripts g−1 sdw in average), while at SW 72 hours after rewetting (T4) a significant but temporarily decrease from 3.1×104 to 8.6×103 transcripts g−1 sdw was observed, followed by an increase to 2.6×104 transcripts g−1 sdw (T5).
Regarding denitrification, transcripts of the functional genes nirK, nirS, cnor and nosZ were quantified, targeting the stepwise reduction of nitrite (NO2 −) to dinitrogen via nitric oxide (NO) and nitrous oxide (N2O). While nirS transcripts were undetectable in all samples, functionally redundant nirK transcript numbers ranged from 3.8×102 to 5.1×105 transcripts g−1 sdw and were highly affected by site, drought and simulated precipitation. At NW, nirK transcripts were highest 24 hours after rewetting (5.1×105 transcripts g−1 sdw at T3) and lowest at T5 with 6.9×103 transcripts g−1 sdw. In contrast, nirK transcript numbers at SW were severely decreased by roof-intensified drought at T2 (3.8×102 transcripts g−1 sdw). Although rewetting led to an temporarily increase in nirK abundance up to 8.0×103 transcripts g−1 sdw (T3, P > 0.05), transcripts decreased afterwards and remained lower hereafter compared to T1 at SW. However, transcript numbers of nirK were significantly reduced at SW compared to NW at the drought and rewetting treatment (T2–T4). In contrast to nirK, cnor transcripts at NW were more affected by drought and rewetting than at SW. At NW, a decrease of cnor transcript numbers from 1.5×106 to 1.8×105 transcripts g−1 sdw at T2 (drought) was observed (P > 0.05), followed by a temporarily increase to 1.9×106 transcripts g−1 sdw after rewetting (P > 0.05). At SW, cnor decreased from 6.6×104 transcripts g−1 sdw (T1) to 3.8×104 transcripts g−1 sdw (T2, P > 0.05), remained constant during rewetting and increased to 8.9×104 transcripts g−1 sdw at T5 (P > 0.05). Although the time-dependent effects were not significant, cnor transcript numbers were significantly lower at SW compared to NW for T1-T4. For nosZ transcripts, similar time patterns as for cnor were observed. Transcription levels of nosZ at NW decreased during drought to 8.1×103 transcripts g−1 sdw (P > 0.05), increased significantly after rewetting (3.9×106 transcripts g−1 sdw at T3) and declined again to 6.4×104 transcripts g−1 sdw at T5 (P > 0.05). At SW, nosZ transcripts were highest at T1 (4.1×104 transcripts g−1 sdw), declined to 4.6×103 transcripts g−1 sdw after rewetting (T3) and increased again to 7.6×103 transcripts g−1 sdw at T5 (P > 0.05). However, also nosZ transcripts were lower at SW compared to NW, except after drought (T2) and in September (T5).
Discussion
Biological nitrogen fixation is negligible in the investigated beech forest ecosystem
Despite general N limitation of forest ecosystems and the presence of nifH genes in all of our samples (2×107 to 5×107 copies g−1 sdw, Fig S1), no nifH transcripts were detected, indicating a negligible role of biological nitrogen fixation (BNF) in the investigated ecosystem. This is in contrast to other studies showing abundant BNF in temperate and boreal forests [34], [35], [36]. However, it might be possible that atmospheric N deposition in this study was sufficient to outbalance ecosystem N losses by gaseous emissions and leaching, thus providing no advantage in the highly energy demanding N2 fixation [37].
Activity of bacteria involved in N mineralization
In terrestrial ecosystems, the majority of soil N is present in organic macromolecules like chitin, proteins and nucleic acids [22], [38], which cannot be assimilated by plants directly [39]. Therefore, abundances of apr and chiA transcripts were used as proxy for soil N mineralizing microbes [24], [40], [41]. For both apr and chiA transcript numbers a significant site effect was observed with lower transcripts under climate change conditions (increased soil temperature and decreased water availability) in average. This was surprising, as soil warming was shown to enhance biological processes like decomposition and N mineralization in general [42], [43]. However, reduced soil moisture may result in decreased microbial activity due to osmotic regulation (accumulation of compatible solutes in microbial cells), limited diffusive transport of substrates and extracellular enzymes and lower microbe motility [9]. In the investigated ecosystem, the negative effect of low water availability seamed to dominate over temperature-induced higher activity of microorganisms. This is in line with previous studies showing that net N mineralization rate was more limited by low soil moisture than promoted by increased temperature [44], [45]. Surprisingly, transcript levels for both apr and chiA remained unaffected by drought and rewetting at both NW and SW, which is in contrast to other studies showing rapidly increasing N mineralization after rewetting of dry soil due to accumulated plant and microbial necromass and microbial cell lysis caused by osmotic stress [15], [46].
It has to be taken into account that transcript abundances of chiA and apr must not necessarily reflect in situ mineralization rates because of post-transcriptional and –translational modifications of the underlying mRNA and/or enzymes [47]. Moreover, besides chitinases and proteases a lot of other enzymes contribute to N mineralization in soil. When relating total mineralization rates to abundance and activity of microbes the role of fungal proteases has to be considered especially in forest soils. However, here the molecular basis is fairly unknown so far, as sequenced fungal isolates are rare.
Activity pattern of ammonia oxidizers
Nitrification, the stepwise oxidation of ammonia to nitrate via nitrite, was quantified via expression of archaeal and bacterial amoA genes. Abundance of AOA ranged from 4×107 to 2×108 copies g−1 sdw and was thus 100-times higher than AOB abundance (Fig S1). Consequently, on transcript level AOB was undetectable. This numerical dominance of AOA over AOB was found in several studies [19], [48], [49]. Although nitrification as aerobic process could be expected to be positively influenced by climate change (increased soil temperature and reduced soil moisture and thus higher oxygen content) as reported previously [50], [51], our results revealed lower AOA transcript numbers at SW compared to NW in average. It is likely that this was due to a soil moisture threshold for optimal nitrification, which is in the range of 60–65% of WHC at the investigated study site [3]. While water availability at NW was within this range, soil moisture at SW was reduced far beyond this optimum. Surprisingly, although AOA were thought to be highly responsive to changing environmental conditions and more dynamic than AOB [19], no drought or rewetting effect on AOA transcript abundance was observed in this study. This was also reflected in soil ammonium concentrations which remained constant during the sampling period, with the exception of temporarily increased ammonium levels after rewetting. As archaeal amoA transcripts were not reduced, this might be due to decreased expression of bacterial amoA genes because of lower oxygen availability (which could not be measured as AOB were below detection limit) and/or increased mineralization as described above. However, ammonium concentrations declined within 72 hours after rewetting, indicating high ammonium immobilization by heterotrophs and/or rapid plant uptake.
Transcript abundances of different genes involved in denitrification related processes
The nitrate produced during nitrification may serve as substrate for denitrification, resulting in the stepwise reduction to gaseous compounds (NO, N2O, N2) and consequently in a loss of N from the ecosystem. In the present study, the last three steps of denitrification (the reduction of NO2 −, NO and N2O) were investigated by quantifying the transcripts of nirK/nirS, cnor and nosZ genes, respectively. Denitrification is closely linked to labile C and nitrate availability as well as oxygen partial pressure [52] and thus can be assumed to be highly sensitive to climate change with increased temperature and fluctuating water regimes.
The reduction of NO2 − is catalyzed by two functionally redundant enzymes encoded by nirK and nirS genes which do never co-occur in one organism [53]. The numerical dominance of nirK over nirS gene abundance observed in this study (Fig S1) was reported previously in soil ecosystems [41], [54], [55]. However, the low nirS gene abundance in our samples resulted in undetectable transcript numbers of nirS. Interestingly, transfer from NW to SW representing ambient climate change did not decrease nirK transcripts. This might be explained by enhanced microbial activity because of increased temperature which outbalanced the negative effect of higher soil oxygen content [3] and/or the presence of anaerobic microsites due to elevated soil respiration and clayey texture promoting denitrification [56], [57]. This would be in agreement with the observation that nirK was not decreased during ambient summer drought at NW. In contrast, roof-intensified drought at SW resulted in a dramatic decline of nirK transcripts, indicating that soil moisture and consequently substrate diffusion and presence of anaerobic microsites were decreased to an extent that could not be compensated by higher temperature. However, rewetting led to a rapid increase of nirK transcript numbers on both sites, which resulted consequently in reduced soil nitrate concentrations. This rapid recovery from drought stress after rewetting was reported previously for nirK harbouring microorganisms [19].
In contrast to nirK, cnor and nosZ transcripts were already influenced by ambient climate change conditions, resulting in decreased transcript levels of both genes at SW compared to NW at T1. Denitrifiers are heterotrophic organisms and among others dependent on labile C sources [52]. As plant biomass was significantly reduced at SW, it is likely that also root exudation and consequently labile C compounds were lowered [2], resulting in decreased denitrification activity. On the other hand, the different response might be due to higher oxygen sensitivity of cnor and nosZ compared to nirK, which at least is known for nosZ [58], [59]. Accordingly, after ambient summer drought at NW as well as after roof-intensified drought at SW both cnor and nosZ transcripts showed decreasing trends, but with different intensities. Interestingly, after rewetting a relative increase of cnor and nosZ was only observed at NW, although nitrate concentrations and DOC availability were similar at both sites. It might be speculated that roof-intensified drought at SW reduced soil moisture far beyond the optimum threshold for NO2 − and N2O reduction, thus causing severe environmental stress [60]. Consequently cnor and nosZ harboring microorganisms need time for recovery when environmental conditions become more favorable again. Such resilience according to changing moisture systems was observed previously for nosZ harboring microorganisms.[61] On the other hand, rewetting of dry soil might have caused increased cell lysis due to high osmotic stress [16], which would also explain the lack of immediate response.
Synthesis
In conclusion, climate change conditions including higher temperature and lower soil moisture resulted in significant site effects with decreased expression of investigated functional genes at SW while the genetic potential was not or not consistently affected. Our data revealed that particularly transcripts related to denitrification were affected by climate change. Already transfer from NW to SW without further treatment resulted in significant reduction of cnor and nosZ transcripts while nirK remained unaffected (Fig. 2A), suggesting the possibility of higher NO losses at moderately increased soil temperatures and decreased water availability. Besides its potential as greenhouse gas, NO is an important signaling molecule for plants participating in a variety of physiological processes including seed germination, root formation, programmed cell death as well as defense and stress responses [62]. Therefore, a possible increase in NO production under climate change may also directly affect plant performance in near future. Severe drought decreased additionally nirK transcripts (Fig. 2B), which could indicate N2 as main gaseous denitrification product at SW. This is in contrast to ambient summer drought at NW, where decreasing trends for cnor and nosZ transcripts were observed while nirK remained unaffected. This is similar to the activity pattern at SW in June (T1) and could suggest higher gaseous NO losses at NW after natural summer drought. Although all investigated denitrification transcripts were significantly lower at SW compared to NW after rewetting (Fig. 2C), nirK increased, cnor remained constant and nosZ decreased relative to the abundance at severe drought, suggesting possible higher gaseous NO and N2O losses. In contrast, rewetting resulted in an increase of all denitrification transcripts at NW. Taken all together, our data suggest that climate change could increase the emission of greenhouse gases (NO, N2O), but this effect might be only temporarily as in September no difference between the denitrification transcript numbers at NW and SW was observed.
Figure 2. Scheme of the microbial nitrogen cycle under different climate change conditions.
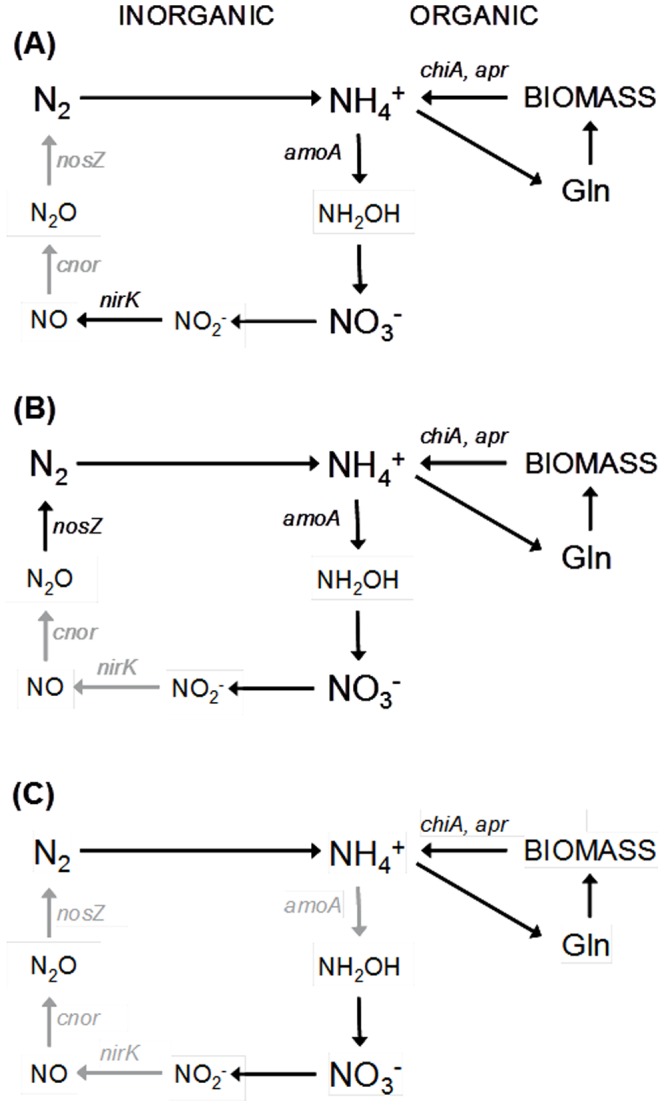
(A) comparison between NW and SW at ambient climate change (T1), (B) comparison between NW and SW at ambient/roof-intensified summer drought (T2) and (C) comparison between NW and SW after rewetting (T3, T4). Decreased N turnover processes under climate change indicated by significantly lower transcripts at SW compared to NW are shown in grey (P <0.05).
As amoA AOA transcripts were reduced after rewetting at SW but chiA and apr transcripts were not affected (Fig. 2C), this suggests that climate change triggered a truncation of soil microbial N cycling in this ecosystem type with decreasing importance of microbial nitrate production and consumption, whereas the effect on N mineralization was not thus pronounced.
Overall, the observed shifts in N turnover appeared to be more related to altered transcription patterns due to environmental factors than to changes in the genetic potential of the microbial community. Accordingly, transcript levels recovered at the end of the growing season under climate change conditions, indicating resilience of the respective microorganisms.
Although our study provides evidence that analysis of N cycle transcript abundances is a useful tool to reconstruct the microbial N cycle and to analyze the response of N cycle processes to climate change at the level of genes encoding for single enzymatic steps, it has to be taken into account that transcript levels of functional genes may allow prediction but must not reflect actual turnover rates. Therefore, more long-term studies measuring gross/net N fluxes additionally to functional genes and their transcripts are required, which would offer the possibility to link biogeochemical quantification of gross N turnover with gene expression for single enzymatic steps. With respect to functional redundancy within microbial populations, also community structure should be investigated to further sharpen our understanding of climate change effects on resilience and vulnerability of N cycle processes.
Supporting Information
Copy numbers of functional genes involved in the nitrogen cycle ( nifH , chiA , apr , amoA AOA, amoA AOB, nirK , nirS , cnor and nosZ ) are shown for NW (black bar) and SW (grey bar) in June (T1), after 39 days drought in August (T2), 24 and 72 hours after rewetting in August (T3, T4) and in September (T5) (n = 8, error bars represent standard deviation of the mean). Asterisks indicate significant differences between NW and SW at the respective sampling times, whereas lower case letters indicate differences among the sampling period for the respective site.
(PDF)
Dry plant biomass at NW and SW, sampled in June (T1), after 39 days drought in August (T2), 24 and 72 hours after rewetting in August (T3, T4) and in September (T5) (n = 8, standard deviation of the mean in parentheses). Asterisks indicate significant differences between NW and SW at the respective sampling times (Student's T test), whereas lower case letters indicate differences among the sampling period for the respective site (multivariate ANOVA). Significant differences between the factors site and sampling time calculated by multivariate ANOVA are indicated by P values <0.05 (bold letters).
(PDF)
Acknowledgments
We also thank Conny Galonska for nucleic acid extraction and all student helpers for their great support during the sampling campaigns.
Data Availability
The authors confirm that all data underlying the findings are fully available without restriction. The basic soil data is owned by a third party, and requests for this data may be sent to schloter@helmholtz-muenchen.de.
Funding Statement
This work has been supported by the German Science Foundation (DFG) under the contract numbers SCHL 447/11-1, DA 1217/2-1, and KO 1035/41-1. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Ellenberg H (1996) Vegetation Mitteleuropas mit den Alpen. Stuttgart: Ulmer.
- 2. Dannenmann M, Simon J, Gasche R, Holst J, Naumann PS, et al. (2009) Tree girdling provides insight on the role of labile carbon in nitrogen partitioning between soil microorganisms and adult European beech. Soil Biol Biochem 41:1622–1631. [Google Scholar]
- 3. Dannenmann M, Gasche R, Ledebuhr A, Papen H (2006) Effects of forest management on soil N cycling in beech forests stocking on calcareous soils. Plant Soil 287:279–300. [Google Scholar]
- 4. Rennenberg H, Dannenmann M, Gessler A, Kreuzwieser J, Simon J, et al. (2009) Nitrogen balance in forest soils: nutritional limitation of plants under climate change stresses. Plant Biol 11:4–23. [DOI] [PubMed] [Google Scholar]
- 5. Schimel JP, Bennett J (2004) Nitrogen mineralization: Challenges of a changing paradigm. Ecology 85:591–602. [Google Scholar]
- 6. Huang S, Pant HK, Lu J (2007) Effects of water regimes on nitrous oxide emission from soils. Ecological Engineering 31:9–15. [Google Scholar]
- 7. Hwang SJ, Hanaki K (2000) Effects of oxygen concentration and moisture content of refuse on nitrification, denitrification and nitrous oxide production. Bioresource Technology 71:159–165. [Google Scholar]
- 8.IPCC (2007) Intergovernmental Panel on Climate Change, Climate change 2007: Synthesis Report, A contribution of Working groups I, II and III to the Fourth Assessment Report of the IPCC.
- 9.Voroney RP (2007) The soil habitat. In: Paul EAeditor. Soil Microbiology, Ecology and Biochemistry. Oxford: Elsevier. pp. 25–49.
- 10. Gorissen A, Tietema A, Joosten NN, Estiarte M, Penuelas J, et al. (2004) Climate change affects carbon allocation to the soil in shrublands. Ecosystems 7:650–661. [Google Scholar]
- 11. Kowalchuk GA, Stephen JR (2001) Ammonia-oxidizing bacteria: A model for molecular microbial ecology. Annu Rev Microbiol 55:485–529. [DOI] [PubMed] [Google Scholar]
- 12. Halverson LJ, Jones TM, Firestone MK (2000) Release of intracellular solutes by four soil bacteria exposed to dilution stress. Soil Sci Soc Am J 64:1630–1637. [Google Scholar]
- 13. Borken W, Davidson EA, Savage K, Gaudinski J, Trumbore SE (2003) Drying and wetting effects on carbon dioxide release from organic horizons. Soil Sci Soc Am J 67:1888–1896. [Google Scholar]
- 14. Prieme A, Christensen S (2001) Natural perturbations, drying-wetting and freezing-thawing cycles, and the emission of nitrous oxide, carbon dioxide and methane from farmed organic soils. Soil Biol Biochem 33:2083–2091. [Google Scholar]
- 15. Borken W, Matzner E (2009) Reappraisal of drying and wetting effects on C and N mineralization and fluxes in soils. Glob Chang Biol 15:808–824. [Google Scholar]
- 16. Fierer N, Schimel JP (2003) A proposed mechanism for the pulse in carbon dioxide production commonly observed following the rapid rewetting of a dry soil. Soil Sci Soc Am J 67:798–805. [Google Scholar]
- 17. Pesaro M, Nicollier G, Zeyer J, Widmer F (2004) Impact of soil drying-rewetting stress microbial communities and activities and on degradation of two crop protection products. Appl Environ Microbiol 70:2577–2587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Stres B, Danevcic T, Pal L, Fuka MM, Resman L, et al. (2008) Influence of temperature and soil water content on bacterial, archaeal and denitrifying microbial communities in drained fen grassland soil microcosms. FEMS Microbiol Ecol 66:110–122. [DOI] [PubMed] [Google Scholar]
- 19. Szukics U, Abell GCJ, Hödl V, Mitter B, Sessitsch A, et al. (2010) Nitrifiers and denitrifiers respond rapidly to changed moisture and increasing temperature in a pristine forest soil. FEMS Microbiol Ecol 72:395–406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Braker G, Tiedje JM (2003) Nitric oxide reductase (norB) genes from pure cultures and environmental samples. Appl Environ Microbiol 69:3476–3483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21. Pena R, Simon J, Rennenberg H, Polle A (2013) Ectomycorrhiza affect architecture and nitrogen partitioning of beech (Fagus sylvatica L.) seedlings under shade and drought. Environ Exp Bot 87:207–217. [Google Scholar]
- 22. Schulten HR, Schnitzer M (1997) The chemistry of soil organic nitrogen: a review. Biol Fertil Soils 26:1–15. [Google Scholar]
- 23. Asmar F, Eiland F, Nielsen NE (1994) Effect of extracellular-enzyme activities on solubilization rate of soil organic nitrogen. Biol Fertil Soils 17:32–38. [Google Scholar]
- 24. Brankatschk R, Töwe S, Kleineidam K, Schloter M, Zeyer J (2011) Abundances and potential activities of nitrogen cycling microbial communities along a chronosequence of a glacier forefield. ISME J 5:1025–1037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Ekenler M, Tabatabai MA (2002) beta-glucosaminidase activity of soils: effect of cropping systems and its relationship to nitrogen mineralization. Biol Fertil Soils 36:367–376. [Google Scholar]
- 26. Dannenmann M, Butterbach-Bahl K, Gasche R, Willibald G, Papen H (2008) Dinitrogen emissions and the N2:N2O emission ratio of a Rendzic Leptosol as influenced by pH and forest thinning. Soil Biol Biochem 40:2317–2323. [Google Scholar]
- 27.IUSS (2007) Working Group WRB, World Reference Base for Soil Resources 2006, first update 2007. Rome: FAO.
- 28. Bimueller C, Dannenmann M, Tejedor J, von Luetzow M, Buegger F, et al. (2014) Prolonged summer droughts retard soil N processing and stabilization in organo-mineral fractions. Soil Biol Biochem 68:241–251. [Google Scholar]
- 29. Bilela S, Dounavi A, Fussi B, Konnert M, Holst J, et al. (2012) Natural regeneration of Fagus sylvatica L. adapts with maturation to warmer and drier microclimatic conditions. Forest Ecol Manag 275:60–67. [Google Scholar]
- 30. Gessler A, Schrempp S, Matzarakis A, Mayer H, Rennenberg H, et al. (2001) Radiation modifies the effect of water availability on the carbon isotope composition of beach (Fagus sylvatica). New Phytol 150:653–664. [Google Scholar]
- 31. Gessler A, Keitel C, Nahm M, Rennenberg H (2004) Water shortage affects the water and nitrogen balance in central European beech forests. Plant Biol 6:289–298. [DOI] [PubMed] [Google Scholar]
- 32. Lueders T, Manefield M, Friedrich MW (2004) Enhanced sensitivity of DNA- and rRNA-based stable isotope probing by fractionation and quantitative analysis of isopycnic centrifugation gradients. Environ Microbiol 6:73–78. [DOI] [PubMed] [Google Scholar]
- 33. Schreiner K, Hagn A, Kyselková M, Moënne-Loccoz Y, Welzl G, et al. (2010) Comparison of barley succession and Take-All disease as environmental factors shaping the rhizobacterial community during Take-All decline. Appl Environ Microbiol 76:4703–4712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Cole D (1995) Soil nutrient supply in natural and managed forests. Plant Soil 168-169:43–53. [Google Scholar]
- 35. Cleveland CC, Townsend AR, Schimel DS, Fisher H, Howarth RW, et al. (1999) Global patterns of terrestrial biological nitrogen (N2) fixation in natural ecosystems. Glob Biogeochem Cycle 13:623–645. [Google Scholar]
- 36. van der Heijden MGA, Bardgett RD, van Straalen NM (2008) The unseen majority: soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol Lett 11:296–310. [DOI] [PubMed] [Google Scholar]
- 37. Zehr JP, Jenkins BD, Short SM, Steward GF (2003) Nitrogenase gene diversity and microbial community structure: a cross-system comparison. Environ Microbiol 5:539–554. [DOI] [PubMed] [Google Scholar]
- 38. Lipson D, Näsholm T (2001) The unexpected versatility of plants: organic nitrogen use and availability in terrestrial ecosystems. Oecologia 128:305–316. [DOI] [PubMed] [Google Scholar]
- 39.Paul EA, Clark FE (1996) Ammonification and nitrification. In: Paul EA, Clark FEeditors. Soil Microbiol Biochem. 2 ed. San Diego: Academic Press. pp. 182–183.
- 40. Gschwendtner S, Reichmann M, Müller M, Radl V, Munch J, et al. (2010) Abundance of bacterial genes encoding for proteases and chitinases in the rhizosphere of three different potato cultivars. Biol Fertil Soils 46:649–652. [Google Scholar]
- 41. Töwe S, Albert A, Kleineidam K, Brankatschk R, Dümig A, et al. (2010) Abundance of microbes involved in nitrogen transformation in the rhizosphere of Leucanthemopsis alpina (L.) Heywood grown in soils from different sites of the Damma glacier forefield. Microb Ecol 60:762–770. [DOI] [PubMed] [Google Scholar]
- 42. Shaw MR, Harte J (2001) Control of litter decomposition in a subalpine meadow-sagebrush steppe ecotone under climate change. Ecol Appl 11:1206–1223. [Google Scholar]
- 43. Domisch T, Finer L, Laine J, Laiho R (2006) Decomposition and nitrogen dynamics of litter in peat soils from two climatic regions under different temperature regimes. Eur J Soil Biol 42:74–81. [Google Scholar]
- 44. Emmett BA, Beier C, Estiarte M, Tietema A, Kristensen HL, et al. (2004) The response of soil processes to climate change: Results from manipulation studies of shrublands across an environmental gradient. Ecosystems 7:625–637. [Google Scholar]
- 45. Beier C, Emmett BA, Penuelas J, Schmidt IK, Tietema A, et al. (2008) Carbon and nitrogen cycles in European ecosystems respond differently to global warming. Sci Total Environ 407:692–697. [DOI] [PubMed] [Google Scholar]
- 46. Mikha MM, Rice CW, Milliken GA (2005) Carbon and nitrogen mineralization as affected by drying and wetting cycles. Soil Biol Biochem 37:339–347. [Google Scholar]
- 47. Röling WFM (2007) Do microbial numbers count? Quantifying the regulation of biogeochemical fluxes by population size and cellular activity. FEMS Microbiol Ecol 62:202–210. [DOI] [PubMed] [Google Scholar]
- 48. Jung J, Yeom J, Han J, Kim J, Park W (2012) Seasonal changes in nitrogen-cycle gene abundances and in bacterial communities in acidic forest soils. Journal of Microbiology 50:365–373. [DOI] [PubMed] [Google Scholar]
- 49. Leininger S, Urich T, Schloter M, Schwark L, Qi J, et al. (2006) Archaea predominate among ammonia-oxidizing prokaryotes in soils. Nat 442:806–809. [DOI] [PubMed] [Google Scholar]
- 50. Hart SC (2006) Potential impacts of climate change on nitrogen transformations and greenhouse gas fluxes in forests: a soil transfer study. Glob Chang Biol 12:1032–1046. [Google Scholar]
- 51. Tourna M, Freitag TE, Nicol GW, Prosser JI (2008) Growth, activity and temperature responses of ammonia-oxidizing archaea and bacteria in soil microcosms. Environ Microbiol 10:1357–1364. [DOI] [PubMed] [Google Scholar]
- 52.Barnard R, Leadley PW, Hungate BA (2005) Global change, nitrification, and denitrification: A review. Glob Biogeochem Cycle 19.
- 53. Jones CM, Hallin S (2010) Ecological and evolutionary factors underlying global and local assembly of denitrifier communities. ISME J 4:633–641. [DOI] [PubMed] [Google Scholar]
- 54. Enwall K, Throback IN, Stenberg M, Soderstrom M, Hallin S (2010) Soil resources influence spatial patterns of denitrifying communities at scales compatible with land management. Appl Environ Microbiol 76:2243–2250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55. Sharma S, Aneja MK, Mayer J, Munch JC, Schloter M (2005) Diversity of transcripts of nitrite reductase genes (nirK and nirS) in rhizospheres of grain legumes. Appl Environ Microbiol 71:2001–2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56. Renault P, Sierra J (1994) Modeling oxygen diffusion in aggregated soils: II. Anaerobiosis in topsoil layers. Soil Sci Soc Am J 58:1023–1030. [Google Scholar]
- 57. Smith KA (1980) A model of the extent of anaerobic zones in aggregated soils, and its potential application to estimates of denitrification. Journal of Soil Science 31:263–277. [Google Scholar]
- 58. Morley N, Baggs EM, Dorsch P, Bakken L (2008) Production of NO, N2O and N2 by extracted soil bacteria, regulation by NO2 − and O2 concentrations. FEMS Microbiol Ecol 65:102–112. [DOI] [PubMed] [Google Scholar]
- 59. Otte S, Grobben NG, Robertson LA, Jetten MSM, Kuenen JG (1996) Nitrous oxide production by Alcaligenes faecalis under transient and dynamic aerobic and anaerobic conditions. Appl Environ Microbiol 62:2421–2426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60. Stark JM, Firestone MK (1995) Mechanisms for soil moisture effects on activity of nitrifying bacteria. Appl Environ Microbiol 61:218–221. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61. Yang L, Kent AD, Wang X, Funk TL, Gates RS, et al. (2014) Moisture effects on gas-phase biofilter ammonia removal efficiency, nitrous oxide generation, and microbial communities. J Hazard Mat 271:292–301. [DOI] [PubMed] [Google Scholar]
- 62. He H, He L, Gu M (2014) The diversity of nitric oxide function in plant responses to metal stress. BioMetals 27:219–228. [DOI] [PubMed] [Google Scholar]
- 63. Rösch C, Mergel A, Bothe H (2002) Biodiversity of denitrifying and dinitrogen-fixing bacteria in an acid forest soil. Appl Environ Microbiol 68:3818–3829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64. Schauss K, Focks A, Leininger S, Kotzerke A, Heuer H, et al. (2009) Dynamics and functional relevance of ammonia-oxidizing archaea in two agricultural soils. Environ Microbiol 11:446–456. [DOI] [PubMed] [Google Scholar]
- 65. Rotthauwe JH, Witzel KP, Liesack W (1997) The ammonia monooxygenase structural gene amoA as a functional marker: molecular fine-scale analysis of natural ammonia-oxidizing populations. Appl Environ Microbiol 63:4704–4712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66. Michotey V, Mejean V, Bonin P (2000) Comparison of methods for quantification of cytochrome cd 1-denitrifying bacteria in environmental marine samples. Appl Environ Microbiol 66:1564–1571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67. Throbäck IN, Enwall K, Jarvis Å, Hallin S (2004) Reassessing PCR primers targeting nirS, nirK and nosZ genes for community surveys of denitrifying bacteria with DGGE. FEMS Microbiol Ecol 49:401–417. [DOI] [PubMed] [Google Scholar]
- 68. Braker G, Fesefeldt A, Witzel K-P (1998) Development of PCR primer systems for amplification of nitrite reductase genes (nirK and nirS) to detect denitrifying bacteria in environmental samples. Appl Environ Microbiol 64:3769–3775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69. Henry S, Baudoin E, Lopez-Gutierrez JC, Martin-Laurent F, Brauman A, et al. (2004) Quantification of denitrifying bacteria in soils by nirK gene targeted real-time PCR. J Microbiol Methods 59:327–335. [DOI] [PubMed] [Google Scholar]
- 70. Henry S, Bru D, Stres B, Hallet S, Philippot L (2006) Quantitative detection of the nosZ gene, encoding nitrous oxide reductase, and comparison of the abundances of 16S rRNA, narG, nirK, and nosZ genes in soils. Appl Environ Microbiol 72:5181–5189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71. Xiao X, Yin X, Lin J, Sun L, You Z, et al. (2005) Chitinase genes in lake sediments of Ardley Island, Antarctica. Appl Environ Microbiol 71:7904–7909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72. Bach HJ, Hartmann A, Schloter M, Munch JC (2001) PCR primers and functional probes for amplification and detection of bacterial genes for extracellular peptidases in single strains and in soil. J Microbiol Methods 44:173–182. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Copy numbers of functional genes involved in the nitrogen cycle ( nifH , chiA , apr , amoA AOA, amoA AOB, nirK , nirS , cnor and nosZ ) are shown for NW (black bar) and SW (grey bar) in June (T1), after 39 days drought in August (T2), 24 and 72 hours after rewetting in August (T3, T4) and in September (T5) (n = 8, error bars represent standard deviation of the mean). Asterisks indicate significant differences between NW and SW at the respective sampling times, whereas lower case letters indicate differences among the sampling period for the respective site.
(PDF)
Dry plant biomass at NW and SW, sampled in June (T1), after 39 days drought in August (T2), 24 and 72 hours after rewetting in August (T3, T4) and in September (T5) (n = 8, standard deviation of the mean in parentheses). Asterisks indicate significant differences between NW and SW at the respective sampling times (Student's T test), whereas lower case letters indicate differences among the sampling period for the respective site (multivariate ANOVA). Significant differences between the factors site and sampling time calculated by multivariate ANOVA are indicated by P values <0.05 (bold letters).
(PDF)
Data Availability Statement
The authors confirm that all data underlying the findings are fully available without restriction. The basic soil data is owned by a third party, and requests for this data may be sent to schloter@helmholtz-muenchen.de.