

Abstract
Accounts of the role of Broca’s area in sentence comprehension range from specific syntactic operations to domain-general processes. The present study was designed to tease apart these two general accounts by measuring the BOLD response to two syntactically distinct long-distance dependencies that invoke abstractly similar predictive processes: backward anaphora and filler-gap dependencies. Previous research has observed distance effects in Broca’s area for filler-gap dependencies, but not canonical anaphora, which has been interpreted in support of a syntactic movement account. However, filler-gap dependencies engage predictive mechanisms, resulting in active search for the gap, while canonical anaphora do not. Backward anaphora correct for this asymmetry as they engage a predictive mechanism that parallels the active search in filler-gap dependencies. The results revealed a distance effect in the pars triangularis of Broca’s area for the backward anaphora condition, supporting a prediction-based role for this region rather than one for a particular syntactic operation.
Keywords: Syntax, Sentence Processing, fMRI, Left inferior frontal gyrus, Broca’s area, Prediction, Cognitive Control, Syntactic movement, Anaphora, English
1. Introduction
It is a relatively undisputed observation that Broca’s area (left inferior frontal gyrus; LIFG) shows increased response in functional magnetic resonance imaging (fMRI) to sentences with noncanonical word order (Stromswold et al., 1996; Friederici et al., 2006b; Makuuchi et al., 2009; Santi & Grodzinsky, 2007a; Bornkessel et al., 2005; Rogalsky et al., 2008). The critical question, however, is: what role is the area actually playing during the comprehension of sentences? Answers typically come in one of two forms: structural accounts and processing accounts. Structural accounts posit that Broca’s area (or a subregion of it) subserves specific syntactic operations, such as phrase-structure building (Friederici 2006a; Friederici, 2011), syntactic movement (Grodzinsky, 2000; 2008; Santi & Grodzinsky, 2007a) or syntactic unification (Hagoort 2005). Processing accounts posit that Broca’s area is not involved in syntax itself, but in various aspects of sentence comprehension that are sentence-specific, such as the interaction of syntax and semantics (Bornkessel et al., 2005; Bornkessel-Schlesewksy et al., 2009) or syntactic working memory (Fiebach et al., 2005), or domain-general processes such as cognitive control (Novick et al., 2005; January et al., 2009; Bornkessel-Schlesewsky & Schlesewsky, 2013) or verbal working memory (Rogalsky et al., 2008; Rogalsky & Hickok, 2011).
Distinguishing among these alternatives is difficult because sentence contrasts based on structure are often conflated with one or more processing differences. The goal of the experiment reported here is to better tease apart the contribution of syntactic and processing factors to activation in Broca’s area using a novel construction known as backward anaphora (or perhaps more succinctly, cataphora). The current experiment builds on previous studies that manipulated distance between a displaced wh-word and gap location in filler-gap constructions (Fiebach et al., 2005; Santi & Grodzinsky, 2007a; Santi & Grodzinsky, 2010), and one study comparing distance effects in filler-gap constructions to manipulations of the distance between an antecedent and a pronoun in canonical anaphoric dependencies (Santi & Grodzinsky, 2007a). This study revealed activation in an anterior portion of Broca’s area in the vicinity of Brodmann area 45 (BA45) in the pars triangularis for filler-gap dependencies but not for canonical anaphoric dependencies. Because filler-gap dependencies involve syntactic movement while canonical anaphoric dependencies do not, these results have been interpreted as evidence for a syntactic account of Broca’s area (Santi & Grodzinsky, 2007a). However, filler-gap dependencies and canonical anaphoric dependencies also differ along an important dimension of sentence processing that could have been responsible for the difference in activation: filler-gap dependencies trigger an active prediction mechanism that attempts to complete the dependency as incoming material is parsed (Crain & Fodor, 1985; Stowe, 1986; Frazier & Flores d’Arcais, 1989), while canonical anaphoric dependencies do not trigger such a prediction mechanism and instead involve a mechanism that searches backward through the memory encoding of previously parsed material for the antecedent of the pronoun. Backward anaphoric dependencies eliminate this confound by reversing the order of the antecedent and pronoun such that the pronoun precedes its “antecedent”. Backward anaphoric dependencies have been shown to engage an active prediction mechanism similar to the one engaged by filler-gap dependencies (Kazanina et al., 2007a; van Gompel & Liversedge, 2003). In this way, a comparison between filler-gap dependencies and backward anaphoric dependencies better isolates the syntactic difference between filler-gap and anaphoric dependencies (movement versus no-movement) while controlling for processing differences (both involve active prediction mechanisms).
The experiment reported here reveals a syntactic distance effect for backward anaphora in the pars triangularis of Broca’s area. These results suggest that this region is not selectively sensitive to the syntactic operation underlying filler-gap dependencies (e.g., wh-movement), but instead is sensitive to the processing of long-distance dependencies that involve active prediction mechanisms. The results reported here do not isolate exactly which aspects of the prediction mechanisms are responsible for the observed activity (e.g., syntactic working memory mechanisms, cognitive control, or syntax-specific aspects of the prediction such as the application of abstract structural constraints on the locations of gaps and antecedents), but they do serve to further narrow the space of possible roles for the pars triangularis of Broca’s area in sentence comprehension.
1.1. Long-distance dependencies and Broca’s area
Long-distance dependencies are relationships between non-adjacent elements in a sentence. One type of long-distance dependency is filler-gap dependencies: so-called because they consist of an element (the filler) that appears to be displaced from a position (the gap) later in the sentence. Sentence (1) demonstrates one example of a filler-gap dependency, a WH-question. Subscripts indicate the relationship between the filler and the gap:
-
(1)
[Which cat]1 did the dog chase ___1 ?
In order to derive the correct meaning of (1), which cat must be interpreted at the gap location as the object of the verb chase. Filler-gap dependencies are analyzed in generative syntax as an instance of syntactic movement (e.g., Chomsky, 1981; Chomsky, 1995). Movement is a syntactic operation whereby an element at some position in the sentence is displaced to a position that is both higher in the structure and earlier in the linear order of the sentence. Movement results in filler-gap dependencies, as the moved element must be linked to its base-generated position for successful interpretation of the sentence. Brain imaging studies have consistently demonstrated an association between Broca’s area and distance manipulations in syntactic movement (Fiebach et al., 2005; Friederici et al., 2006b; Santi & Grodzinsky, 2007a; Santi & Grodzinsky, 2010). For example, Santi & Grodzinsky (2007a) parametrically increased the distance between a moved noun phrase (NP) and its gap by inserting intervening NPs, showing that an anterior portion of the pars triangularis (BA45) exhibits a linear increase in activity with distance. These imaging results converge with previous evidence in patients with Broca’s aphasia, who appear to have a comprehension deficit selective to sentences with movement (Grodzinsky, 2000). To account for this deficit, Grodzinsky (2000) developed the Trace-Deletion Hypothesis (TDH), according to which the agrammatic deficit in Broca’s aphasia consists of an inability to compute filler-gap dependencies due to damage in Broca’s area. The hypothesis stemming from the combined results of lesion and neuroimaging studies is the syntactic movement hypothesis of Broca’s area (Grodzinsky, 2008), which holds that the region is responsible for the computation of syntactic movement during comprehension. The data supporting an association between movement and Broca’s area are broadly consistent with other accounts of the region’s role in sentence processing, such as the accounts proposed by Friederici and colleagues, which posit that various subregions of Broca’s area support hierarchical syntactic processes (Bahlmann et al., 2008; Friederici 2011; Friederici et al., 2006a; 2006b; Grodzinsky & Friederici, 2006) or ‘syntactic working memory’ (Fiebach et al., 2005), and the accounts proposed by Hagoort and colleagues, which posit that Broca’s area supports syntactic and semantic unification during sentence processing (Hagoort 2005; Snijders et al., 2009).
1.2. Activations in Broca’s area during sentence processing: syntactic operations or domain-general processing?
The syntactic movement hypothesis and related claims share the fundamental assertion that Broca’s area supports a specific syntactic operation. Alternatively, some hypotheses posit that activation in Broca’s area during sentence comprehension reflects domain-general processes. For instance, Kaan & Swaab (2002) reviewed several neuroimaging studies of syntactic processing and concluded that Broca’s area likely contributes to sentence comprehension by contributing additional resources when processing load increases. This conclusion is consistent with the verbal working memory account proposed by Rogalsky, Hickok and colleagues (Rogalsky et al., 2008; Rogalsky & Hickok, 2011), which posits that the posterior portion of Broca’s area in the pars opercularis contributes to the comprehension of complex sentences via its role as the articulatory component of a phonological loop in working memory (Baddeley & Hitch, 1974; Baddeley, 1981). This claim is supported by a study in which the additional activation in the pars opercularis due to a sentence complexity manipulation was eliminated under conditions of articulatory suppression (Rogalsky et al., 2008). Rogalsky et al. interpret this as evidence that verbal working memory resources were saturated by articulatory suppression and unavailable during sentence comprehension. Despite this potential verbal working memory explanation for activation in the pars opercularis, Rogalsky et al. did not find that articulatory rehearsal could account for activations in the anterior portion of Broca’s area in the pars triangularis, so it remains possible that this region could support syntax-specific processes.
Another domain-general processing account of Broca’s area’s role in language is the cognitive control (Novick et al., 2005) hypothesis, which posits that increased activation during sentence processing results from competing representations that must be ruled out and suppressed. This account is supported by data from January et al. (2009), who showed co-localized activation in Broca’s area for a stroop task and a sentence comprehension task involving syntactic ambiguity, suggesting that the region supports a domain-general cognitive control mechanism.
In addition to syntactic operations and domain-general processing, a third possibility is that activations in Broca’s area reflect sentence-specific mechanisms during comprehension and/or production not comprising a purely syntactic operation. For instance, Bornkessel-Schlesewsky, Schlesewsky and colleagues have found that the pars opercularis of Broca’s area shows increased activation with increased difficulty in argument linearization, that is, in assigning thematic roles (e.g., agent, patient) to the participants in a sentence (Bornkessel et al., 2005; Bornkessel-Schlesewsky et al., 2009). Dissociating among these accounts remains a difficult task for clarification of the roles of the different subregions of Broca’s area during sentence processing.
The question of functional specificity in this brain region remains under contention, although evidence has been offered to distinguish among the alternatives. In the study by Santi & Grodzinsky (2007a), the distance effect found for the movement condition did not hold for another type of long-distance dependency, anaphora. An anaphoric dependency is the co-reference between an anaphor (a pronoun or reflexive) and the noun to which it refers (the antecedent). Sentence (2) illustrates such a dependency:
-
(2)
[The boy]1 fell down the stairs and hurt himself1.
Movement and binding constructions are similar in that they both involve a long-distance (non-adjacent) dependency. Because of this, they presumably share some general cognitive demands, such as the working memory required to retrieve the filler/antecedent and integrate it into the context. Despite this similarity, Santi & Grodzinsky (2007a) found that distance in filler-gap dependencies modulated activity in Broca’s area, but distance in anaphoric dependencies did not. This suggests selectivity in the response of this region to the syntactic difference between the two conditions, namely syntactic movement, rather than the domain-general similarities.
However, there is at least one important processing difference between the movement and binding conditions in Santi & Grodzinsky (2007a). In sentences with filler-gap dependencies, the filler always precedes the gap, and serves as a cue to the existence of the long-distance dependency. There is substantial evidence in the sentence processing literature that the parser uses the filler as a cue to engage a prediction mechanism that actively posits a gap location at the first grammatically licensed location that it encounters while parsing the incoming material. This prediction mechanism has been shown to be “active” in the sense that the parser does not appear to wait for unambiguous evidence for the gap location, and can therefore be shown to incorrectly posit gap locations at potential, but not actual, gap locations (Crain & Fodor, 1985; Stowe, 1986; Frazier & Flores d’Arcais, 1989, Garnsey et al. 1989, Traxler and Pickering 1996). In contrast, the linear order of antecedent and pronoun in anaphoric dependencies prevents the possibility of a prediction mechanism. Instead, the pronoun indicates the existence of an anaphoric dependency, and the parser engages in a backward search through memory for the previously encountered antecedent. In this way, the movement sentences in the Santi and Grodzinsky (2007a) experiment involved both syntactic movement AND an active prediction mechanism, while the binding sentences involved no movement and no prediction mechanism. It is possible that this active prediction mechanism may account for the asymmetric activity found in Broca’s area.
1.3. Backward anaphora & the present experiment
The current experiment sought to tease apart the contribution of syntactic movement and the active prediction mechanism to activity in Broca’s area during the processing of long-distance dependencies. To do this we contrasted filler-gap dependencies with backward anaphora. Crucially, in backward anaphora the pronoun precedes the antecedent leading to a configuration in which the pronoun/reflexive can act as a reliable cue to invoke an active prediction mechanism for the antecedent:
-
(3)
Because he1 fell down the stairs, the boy1 went to the hospital.
van Gompel & Liversedge (2003) found that subjects actively predict a coreferential relationship between potential antecedents to the pronoun in these constructions. They presented subjects with sentences like (4a) and (4b):
-
(4a)
When he1 was fed up, the boy1 visited the girl very often.
-
(4b)
When she1 was fed up, the boy visited the girl1 very often.
In (4a), the first noun phrase matches the gender of the first NP; in (4b), it does not. Subjects showed slower reading times to the first NP in (4b) relative to (4a), suggesting that the parser predicted a coreferential relationship with the NP in the subject position of the matrix clause before determining the gender of the noun, and had to reanalyze this relationship after the gender was determined. Using backward anaphora, it is possible to tease apart the contribution of syntactic movement and these prediction mechanisms to activation in Broca’s area. To do this we used a 2×2 design, with factors DISTANCE (short, long) and CONSTRUCTION (filler-gap, backward anaphora). In addition, we added an articulatory rehearsal condition to determine areas involved in speech production in order to account for effects due to verbal working memory. We found a significant main effect of distance in the pars triangularis, with no significant interaction between distance and construction in this region. However, upon investigating effects of distance within each construction separately, we only observed an effect of distance for the backward anaphora condition in the pars triangularis, while failing to replicate previously established distance effects in the filler-gap condition in the this region (see section 4.2 for a discussion). The results suggest that it is unlikely that Broca’s area subserves a specific syntactic process, although it is possible that it supports either sentence-specific or domain-general processing mechanisms depending on which aspect of the active prediction mechanisms are driving the activation (see section 4 for discussion).
2. Materials and methods
2.1. Subjects
Twenty-six right-handed, native speakers of English (age 19–32, 12 males) volunteered for participation. Subjects had normal or corrected-to-normal vision, no hearing impairment, and reported no history of neurological disorder. Subjects were paid $30 an hour for participation. Consent was acquired from each subject before participation and all procedures were approved by the Institutional Review Board of UC Irvine.
2.2. Stimuli and Design
Stimuli were auditory recordings of sentences from a male native English speaker. Filler-gap sentences (WH) consisted of WH-questions in which a matrix clause was modified by an adjunct clause, and the WH-question was formed by displacement of the matrix clause object to the front of the sentence (5a). Backward anaphora sentences (BA) consisted of a fronted causal adjunct clause containing a pronoun (e.g., Because he/she did something…), followed by a brief pause, and the matrix clause with an antecedent for the pronoun in the subject (6a). One hundred matched pairs of sentences (short/long) were created for both constructions. Distance for both dependency types was manipulated with a relative clause: short sentences contained a relative clause at the end of the sentence (5a, 6a); long WH sentences contained a relative clause modifying the subject of the matrix clause (5b); and long BA sentences contained a relative clause modifying the object of the fronted causal adjunct clause (6b):
-
(5a)
WH-SHORT: Which song1 did the band play __1 at the concert [that ended early]?
-
(5b)
WH-LONG: Which song1 did the band [that won the contest] play __1 at the concert?
-
(6a)
BA-SHORT: Because he1 extinguished the flames, the fireman1 saved the resident [that arrived later].
-
(6b)
BA-LONG: Because he1 extinguished the flames [that burned all night long], the fireman1 saved the resident.
Pairs of sentences within each construction were matched for approximate content and number of syllables. Duration of sentences for each condition: WH-SHORT (mean=4.24s, standard deviation =0.40s), WH-LONG (mean=4.24s, standard deviation=0.38s), BA-SHORT (mean=4.37s, standard deviation =0.5s), BA-LONG (mean=4.44s, standard deviation =0.51s). The complete set of materials can be found in the Appendix. In order to avoid familiarity effects, the matched pairs within each construction were divided into two lists, with each list containing fifty sentences from each construction, such that subjects never saw both members of a matched pair. Semantically anomalous sentences (ANOM) were created by generating WH and backward anaphora sentences in the same manner described above, and replacing a noun phrase with one that did not fit context due to animacy violations or selection restrictions (e.g., Which bird did the orchestra that dazzled the audience play expertly and loudly during the musical? Because he vetoed the bill, the plant angered the congress that crafted the legislation). Forty anomalous sentences were generated, distributed nearly equally across both constructions and distances (17 WH, 23 BA). All subjects saw the same set of forty anomalous sentences. Each subject was presented with 50 trials from each of the four conditions, and 40 anomalous trials, for a total of 240 sentence trials. In addition, subjects were asked to subvocally perform articulatory rehearsal of the sequence /pa-ta-ka/ for 50 trials (ART) in order to localize a verbal working memory network, for a combined total of 290 trials.
2.2. Procedure
Subjects were informed that they would be listening to sentences and deciding whether they “made sense” or not. Subjects were instructed to pay close attention to the sentences, understand the meaning of the sentence, and only press a button if they heard an anomalous sentence. During each run, a fixation cross was displayed on a screen. Subjects responded using a button box in the left hand (in order to minimize activations in the left hemisphere) after the offset of the sentence and before the next trial. During ART trials, the fixation cross would flicker red-blue-green at a rate of 2 Hz for 5s, which cued the subjects to articulate the sequence /pa-ta-ka/ without producing sound or opening their mouth while still making movements internal to the vocal tract including tongue movements. Auditory stimuli were delivered with Matlab software (Mathworks, Inc, USA), the Cogent toolbox (http://vislab.ucl.ac.uk/cogent_2000.php), and MR compatible insert headphones. Subjects were given ear covers to attenuate scanner noise. The experiment consisted of twelve runs – one practice run, 10 experimental runs, and one high-resolution anatomical scan. The practice run was intended to familiarize subjects with the task and stimuli. Within experimental runs, five trials of each of the four main conditions, five articulation trials, and four anomalous trials were presented to the subject in random order. The order of trials from each condition was randomized across runs. Each trial lasted a total duration of 10s, with the stimulus jittered from the onset at delays of 0s, 0.5s, 1s, and 1.5s to better capture the peak of the hemodynamic response. Due to a coding error, two subjects did not receive any articulation trials. These subjects were included in the primary analysis, but not the ART analysis. Due to a different coding error, one subject was presented with two identical runs. We consider the impact of repetition effects in this case to be relatively minor (a separate analysis with this subject excluded did not qualitatively change the results), therefore this subject was included in all analyses. The high-resolution anatomical image was collected last. The scanning session lasted about one hour in total.
2.3. fMRI Data Collection and Preprocessing
MR images were obtained in a Philips Achieva 3T (Philips Medical Systems, Andover, MA) fitted with an eight-channel RF receiver head coil at the high field scanning facility at UC Irvine. We first collected a total of 1530 T2*-weighted EPI volumes over 10 runs using Fast Echo EPI in ascending order (TR=2s, TE=25ms, flip angle = 90°, in-plane resolution = 1.95mm × 1.95mm, slice thickness = 3mm with 0.5mm gap). The first four volumes of each run were collected before stimulus presentation and discarded to control for T1 saturation effects. After the functional scans, a high-resolution T1-weighted anatomical image was acquired in the axial plane (TR=8ms, TE=3.7ms, flip angle=8°, size=1mm isotropic).
Slice-timing correction, motion correction, and spatial smoothing were performed using AFNI software (http://afni.nimh.nih.gov/afni). Motion correction was achieved by using a 6-parameter rigid-body transformation, with each functional volume in a run first aligned to a single volume in that run. Functional volumes were aligned to the anatomical image, and subsequently aligned to Talairach space (Talairach and Tournoux, 1988). Functional images were resampled to 2.5mm isotropic voxels, and spatially smoothed using a Gaussian kernel of 6mm FWHM.
First-level analyses were performed on each individual subject’s data using AFNI’s 3dDeconvolve function. The regression analysis was performed to find parameter estimates that best explained variability in the data. Each predictor variable representing the time course of stimulus presentation was entered into a deconvolution analysis that estimated parameters best representing the timecourse of the hemodynamic response function in percent signal change values. The following six regressors of interest were used in the experimental analysis: WH-SHORT, WH-LONG, BA-SHORT, BA-LONG, ANOM, and ART. The six motion parameters were included as regressors of no interest. A second-level analysis was then performed by summing the parameter estimates across each timepoint on the estimated timecourse for each condition for each subject, and entering these values from each subject and condition into AFNI’s 3dANOVA2 function. A cluster-corrected threshold of p < 0.05 (FWE) was used to locate activity for the following contrasts (‘>’ indicates one-tailed tests, ‘-’ indicates two-tailed tests): main effect of distance [LONG > SHORT], main effect of construction [BA - WH], interaction of distance and construction, and the simple effects of BA-distance [BA-LONG > BA-SHORT] and WH-distance [WH-LONG > WH-SHORT]. In order to examine whether distance effects were due to verbal working memory, we ran a separate analysis only including data from the 23 subjects who performed the articulation task, and added the following contrast: [[LONG > SHORT] > ART], in which the activation to articulatory rehearsal was subtracted from the main effect of distance.
3. Results
3.1. Behavioral
Responses to anomalous (ANOM) sentences were categorized according to signal detection theory (hits, misses, false alarms, and correct rejections), such that button presses to anomalous sentences were considered hits, and button presses to non-anomalous (WH and BA) sentences were considered false alarms. One subject was excluded from further analysis due to particularly poor discriminability (d′ = 0.88). The remaining twenty-five subjects had a mean d′ of 2.8 with a standard deviation of .7, indicating successful discriminability of anomalous sentences from normal sentences (by convention, d′=1.0 is considered threshold for discrimination). Table 1 lists the d′ values for each individual condition. These subjects correctly identified anomalous sentences with a rate of 74% (standard deviation 18%), and correctly accepted normal sentences (WH and BA) with a rate of 97%, (standard deviation 3%). These results indicate that the remaining 25 subjects processed the meaning of the sentences during scanning, and importantly, did not identify normal sentences as anomalous with any frequency.
Table 1.
| Condition | BA-SHORT | BA-LONG | WH-SHORT | WH-LONG |
| d′ | 3.04 | 2.82 | 2.96 | 2.31 |
| Stand. dev. | 0.88 | 0.76 | 0.77 | 0.93 |
3.2. fMRI
The main effect of distance revealed one significant cluster of activity in the pars triangularis of Broca’s area (left hemisphere; fig. 1, left). The interaction between distance and construction revealed no activity in this area. At a reduced cluster size threshold, there was a significant interaction in the left pars opercularis and left precentral gyrus, with increased activation for the BA-distance effect in this area (fig. 1, right). The main effect of construction, [BA > WH], revealed activity in bilateral ATL, bilateral angular gyrus, and bilateral precuneus/posterior cingulate (fig. 2, top), while [WH > BA] revealed activity in left precentral gyrus (fig. 2, bottom). As is noted in the discussion, the effect of distance was tightly controlled while the effect of construction was not (namely, the BA sentences consisted of three clauses, one of which was a fronted causal adjunct, while the WH sentences consisted of two clauses, with no causal adjunct), so any interpretation of the main effect of construction will be highly speculative.
Fig. 1.

Group activation maps (n = 25) displayed in Talairach space on a template brain for the main effect of distance (top) and the interaction of distance and construction (bottom).
Activations for the main effect of distance were corrected for multiple comparisons (FWE) at p < 0.05 using an individual t-threshold of p < 0.001 (one-tailed) and a cluster size threshold of 608 mm3. Activations for the interaction are displayed at a reduced cluster size threshold of 200 mm3. Barplots indicate average percent signal change for each condition within selected clusters of activation (error bars indicate standard error of the mean).
Fig. 2.

Group activation maps (n = 25) displayed in Talairach space on a template brain for the main effect of construction. Increased activation for Backward Anaphora sentences is shown on top, and increased activation for WH-questions is shown on bottom. Activations were corrected for multiple comparisons (FWE) at p < 0.05 using an individual t-threshold of p < 0.001 (two-tailed) and a cluster size threshold of 608 mm3. Barplots indicate average percent signal change for each condition within selected clusters of activation (error bars indicate standard error of the mean).
Although the interaction of distance and construction was not significant in the pars triangularis, we planned, a priori, to examine the simple effects of distance separately for each construction, as it is theoretically vital to establish a distance effect in the BA condition on its own. The simple effect of BA-distance [BA-LONG > BA-SHORT] revealed three clusters: one in the left IFG, pars triangularis (fig. 3, left), one in right middle temporal gyrus/superior temporal sulcus (fig. 3, right), and one in the supplementary motor area (bilateral; fig. 3, center). This result confirms our prediction that the novel backward anaphora condition would show a distance effect in the pars triangularis of Broca’s area, given the active prediction mechanism employed in its processing. The simple effect of WH-distance ([WH-LONG-WH-SHORT]) did not reveal any significant activity. While this fails to replicate previous research documenting distance effects for movement constructions in Broca’s area, it is not inconsistent with the observation that distance effects in this region may not be robust. For example, Santi & Grodzinsky found a cluster in the pars triangularis for the linear effect of distance that only had a volume of 128 mm3 at an uncorrected p < 0.005, the smallest cluster revealed by their analysis. In addition, we observed that performance was lowest during the WH-LONG condition (d′ = 2.3); this may have contributed to the lack of a distance effect for the WH-sentences (see section 4.2 for further discussion).
Fig. 3.

Group activation maps (n = 25) displayed in Talairach space on a template brain for the simple effect of distance in the Backward Anaphora condition. Activations were corrected for multiple comparisons (FWE) at p < 0.05 using an individual t-threshold of p < 0.001 (one-tailed) and a cluster size threshold of 608 mm3. Barplots indicate average percent signal change for each condition within selected clusters of activation (error bars indicate standard error of the mean).
Table 2 lists the Talairach coordinates of the center of mass and the cluster size for each of the contrasts listed above (main effect of distance, interaction of distance/construction, main effect of construction, and simple effect of distance in the Backward Anaphora condition).
Table 2.
| Region | Hemisphere | x | y | z | Cluster Size (mm3) |
|---|---|---|---|---|---|
| Main effect of distance | |||||
| Inferior frontal gyrus (pars triangularis) | Left | −50 | 16 | 25 | 656 |
| Interaction of distance & construction | |||||
| Inferior frontal gyrus (pars opercularis)* | Left | −38 | 6 | 19 | 313 |
| Precentral gyrus* | Left | −45 | −6 | 20 | 203 |
| Main effect of construction: BA > WH | |||||
| Anterior temporal lobe | Left | −52 | 0 | −16 | 7,500 |
| Angular gyrus | Left | −47 | −63 | 24 | 5,719 |
| Anterior temporal lobe | Right | 49 | 9 | −19 | 3,547 |
| Posterior cingulate/precuneus | Left | −4 | −55 | 27 | 1,297 |
| Angular gyrus | Right | 51 | −63 | 24 | 891 |
| Main effect of construction: WH > BA | |||||
| Precentral gyrus | Left | −41 | −4 | 38 | 750 |
| BA-LONG > BA-SHORT | |||||
| Inferior frontal gyrus (pars triangularis) | Left | −50 | 17 | 23 | 1000 |
| Superior temporal sulcus | Right | 50 | −27 | −1 | 890 |
| Supplementary motor area | Left/Right | 2 | 12 | 52 | 750 |
n = 25. FWE cluster-corrected p < 0.05, individual voxel threshold p < 0.001, cluster size threshold 608 mm3.
significant at reduced cluster size threshold (200 mm3)
The contrast [[LONG > SHORT] > ART] from the subset of 23 subjects who performed the ART condition revealed two clusters in the vicinity of Broca’s area at a reduced cluster size threshold: one in the left pars triangularis, and a second in the inferior portion of the LIFG in the pars orbitalis. In addition, the contrast revealed effects in bilateral occipital lobe and left superior temporal sulcus. Fig. 4 illustrates these effects, and Table 3 lists the Talairach coordinates of the center of mass and the cluster size for the activations.
Fig. 4.
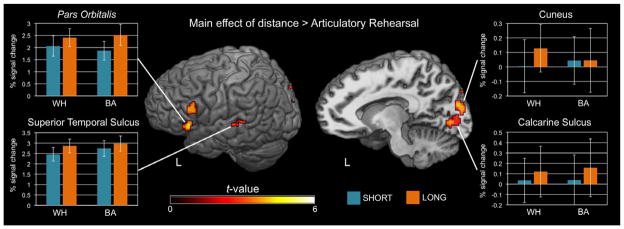
Group activation maps (n = 23) displayed in Talairach space on a template brain for the main effect of distance > articulatory rehearsal. Activations are shown using an individual t-threshold of p < 0.001 (one-tailed) and a cluster size threshold of 300 mm3. Barplots indicate average percent signal change for each condition within selected clusters of activation (error bars indicate standard error of the mean).
Table 3.
| Region | Hemisphere | x | y | z | Cluster Size (mm3) |
|---|---|---|---|---|---|
| Main effect of distance > ART | |||||
| Calcarine sulcus | Left | −5 | −79 | 4 | 1438 |
| Cuneus | Right | 11 | −84 | 23 | 719 |
| Cuneus | Left | −9 | −88 | 16 | 641 |
| Inferior frontal gyrus (pars orbitalis)* | Left | −49 | 22 | 0 | 406 |
| Inferior frontal gyrus (pars triangularis)* | Left | −52 | 18 | 19 | 359 |
| Superior temporal sulcus* | Left | −56 | −33 | 4 | 344 |
n = 23. FWE cluster-corrected p < 0.05, individual voxel threshold p < 0.001, cluster size threshold 610 mm3.
significant at reduced cluster size threshold (300 mm3)
4. Discussion
4.1. Distance effects for Backward Anaphora in Broca’s area
Consistent with our predictions, the distance manipulation in the backward anaphora condition resulted in activity in the pars triangularis of Broca’s area. In combination with previously reported results (Fiebach et al., 2005; Santi & Grodzinsky, 2007a; Santi & Grodzinsky, 2010), this suggests that the contributions of the pars triangularis to sentence processing is not specific to the syntactic operation of movement. Instead, these results suggest that the some aspect of the active prediction mechanisms involved in WH-dependencies and backward anaphoric dependencies is driving activation in Broca’s area.
The question then is which aspect of these prediction mechanisms is driving the effect. The two prediction mechanisms share several functional components, such as access to grammatical knowledge/constraints relevant to licensing the dependencies (e.g., “island” constraints for WH-dependencies (Ross, 1967), and “binding” constraints for anaphoric dependencies (Chomsky, 1982)), cognitive control mechanisms required to manage the resolution of the dependencies in licensed/restricted syntactic contexts, and working memory mechanisms required to retrieve previously encountered material (either syntax-specific or domain-general). The current experiment was designed solely to tease apart the contribution of syntactic movement and active prediction mechanisms; future studies will be required to definitively tease apart the various components of the active prediction mechanisms that could be driving this effect.
Before moving on, it should be noted that two recent studies of activation in Broca’s area are consistent with the hypothesis that Broca’s area supports one or more aspects of prediction mechanisms. First, Pallier et al. (2011) presented sequences of 12 words to participants, and parametrically manipulated the size of the constituent formed by the word sequences from 1 word (i.e., an unrelated list of 12 words) to 12 words (i.e., a full sentence). They observed increased activation in the pars triangularis and pars orbitalis for constituents with real words, and for ‘jabberwocky’ conditions in which the content words in each trial were replaced by nonsense words. Pallier et al. interpret this as evidence that Broca’s area is involved in syntactic structure-building independent of semantic processing. These results, however, are also consistent with our hypothesis that this portion of Broca’s area activates as a result of prediction, as constituents of larger size provide more material from which to form predictions of the upcoming syntactic structure of the constituent, and thereby engaging additional processes such as working memory and cognitive control.
Second, Santi & Grodzinsky (2012) found that WH-questions with one filler and two gap locations did not result in increased activation compared to questions with one filler and one gap location:
-
(7a)
[Which paper]1 did the tired student submit __1 after jogging?
-
(7b)
[Which paper]1 did the tired student submit __1 after reviewing__1?
Crucially, the second gap in these constructions is unpredictable (and in fact is not licensed by itself, leading to the moniker “parasitic gap” for these constructions; Engdahl, 1983). Evidence from online reading times suggest that only the first gap in these constructions engages an active prediction mechanism; the second gap does not appear to be actively predicted by the parser (Wagers & Phillips, 2009). Despite the lack of active prediction in these two-gap constructions, many syntactic analyses derive the two gap locations from two instances of syntactic movement. In this way, the syntactic movement hypothesis should predict increased activation for sentences containing two gaps, whereas a prediction-based hypothesis would predict equal activation between one-gap and two-gap sentences, as only the first gap engages prediction mechanisms in two-gap sentences. The results of Santi & Grodzinsky (2012) and the current experiment together suggest that Broca’s area supports a role in syntactic prediction rather than syntactic movement. Interestingly, Wagers & Phillips (2009) report online reading time evidence that active predictions for two gap locations are in fact made in a different type of double-gap construction known as “across-the-board” movement constructions:
-
(8)
Phil generally dislikes [the poetry]1 that The New Yorker reviews __1 or publishes __1.
Given that both gap locations are actively predicted in across-the-board constructions, the prediction hypothesis suggested here predicts that Broca’s area should be more active in (8) than in (7b). We leave this experiment to future research.
4.2 The lack of distance effects for WH-dependencies in Broca’s Area
Although we found a main effect of distance and no interaction in the pars triangularis (suggesting distance effects for both constructions), closer inspection of the data revealed that the WH distance manipulation did not in fact reveal a significant cluster in this region. The fact that the WH distance manipulation did not result in activation in Broca’s area was unexpected, given previous results documenting increased activation for distance effects in movement constructions (Santi & Grodzinsky, 2007a; Fiebach et al., 2005; Friederici et al., 2006b). However, it is worth noting that Fiebach et al. (2005) found a distance effect only for object-extraction and not for subject-extraction in German, suggesting that there may be more complexity to distance effects than previously thought. The materials in the current experiment differed from the Santi & Grodzinsky 2007a materials in several ways: (i) S&G manipulated distance by number of intervening noun phrases (with a combination of relative clauses and conjunction), whereas we manipulated distance with a relative clause, (ii) S&G tested three levels of the distance manipulation, whereas we tested only two, and (iii) S&G used embedded WH-questions (such that the full sentence was actually a declarative), whereas as we used matrix WH-questions (i.e., the full sentence was a question). These differences suggest that what counts as distance for the prediction mechanisms, and how strong those effects are, may be an interesting area of investigation, especially given the apparent asymmetry between WH-dependencies and BA- dependencies in the current experiment. As mentioned in the results section, their distance effect was statistically tenuous as well. In addition, our behavioral data indicate that the WH-LONG condition was more difficult for subjects to process, and this may have impacted our ability to detect a distance effect in the WH condition. At any rate, the crucial test of the prediction hypothesis resided in the novel backward anaphora condition, which did not include a manipulation related to syntactic movement, but nonetheless resulted in a distance effect in the area of interest.
4.3. Effects of articulatory rehearsal
Activity in the ART condition did not account for the distance effect in the pars triangularis. This suggests that these distance effects are not accounted for by a verbal working memory account. We did find activations to the ART condition in the more posterior and inferior parts of the LIFG (pars opercularis, frontal operculum) that are also reported for syntactic complexity effects (Rogalsky et al., 2008; Fiebach et al., 2005; Friederici et al., 2006b) and we agree with the conclusions of Rogalsky et al. (2008) and Rogalsky & Hickok (2011) that activations in those studies reflect verbal working memory rather than sentence processing or syntactic operations. However, as one reviewer pointed out, given that some distance manipulations find activations in the pars opercularis (Fiebach et al., 2005) while others find activations in the pars triangularis (Santi & Grodzinsky, 2007a; the present study) this does leave open the question of why certain manipulations obtain effects in the pars opercularis, and why other manipulations obtain effects in the pars triangularis. We address this issue by positing differential demands on maintenance (i.e., working memory) vs. selection/suppression (e.g., cognitive control, argument mapping).
In a study that did not manipulate distance, Santi & Grodzinsky (2007b) found that WH-questions activated the pars opercularis while canonical (forward) anaphoric dependencies activated the pars triangularis. Given that anaphora engage the pars triangularis and WH-sentences engage the pars opercularis, we suggest that anaphora rely more on selection/suppression to determine potential antecedents given the representational salience of argument representations and less on verbal working memory, because the anaphor is relatively simple phonologically (e.g., he or she), therefore less demanding to maintain in working memory. We suggest that movement relies more on verbal working memory in maintaining the filler (given the phonological complexity of the WH-phrase compared to a simple pronoun), but less on selection/suppression because structural positions of the possible gaps are less semantically salient representations than arguments. Therefore, distance manipulations for the backward anaphora are likely to show increased engagement of pars triangularis, while distance manipulations for movement are likely to show increased engagement of the pars opercularis. This does contradict the results of Santi & Grodzinsky (2007a) and the lack of a movement distance effect in our results, but we discussed above the tenuous nature of significance of their results, reasons for our lack of significant distance effects in our movement condition, and point to more robust distance effects for movement in other studies (e.g., Fiebach et al., 2005). In addition, this conjecture is compatible with the results of Caplan et al. (2008), who found that sentences with arguments that were thematically unconstrained (e.g., the fireman who called the deputy saved the sailor) resulted in greater activation than those that were thematically constrained (e.g., the policeman who arrested the thief watched the driver) in the anterior portions of the LIFG. However, we cannot make any strong conclusions from this study beyond the fact that the distance effects found in the pars triangularis cannot be attributed to increased demands on verbal working memory as tested in our study.
4.4. Activations in the posterior temporal lobe
The distance manipulation in the backward anaphora condition also revealed activations in right posterior STS/MTG, which have been found in previous studies of syntactic distance and complexity (Bornkessel et al., 2005; Fiebach et al., 2005; Pallier et al., 2011), although we did not observe effects for these regions in the left hemisphere that are typically reported. Grodzinsky & Friederici (2006) and Friederici (2002) suggest that these areas are involved in either lexical-syntactic integration or syntactic repair. This is indirectly supported by results in the P600 event-related potential (ERP) literature. The P600 response is elicited in situations involving syntactic reanalysis and repair such as garden-path sentences and syntactic violations, as well as sentences that are syntactically well-formed but cause difficulties in syntactic integration (Friederici et al., 1996; Kaan et al., 2000; Osterhout & Holcomb, 1992). The distribution of the response on the scalp is typically centro-posterior, and may have a source in the posterior temporal lobe (Friederici et al., 2003). Other researchers have posited the superior/middle temporal lobe as part of phonological and lexical processing networks (Hickok & Poeppel, 2004; 2007; Wise et al., 2001). It is difficult to ascertain whether increased activation in these areas due to the syntactic distance manipulation reflects increased load on phonological and lexical processing, or in lexical-syntactic integration itself. Further research is needed to clarify the role of these regions during sentence processing to disambiguate these two proposals, and is beyond the scope of the present study.
4.5. Main effects of construction
While the effects from the contrasts [BA > WH] and [WH > BA] invite speculation, the experiment was not designed to control for differences between these conditions, which are many (backward anaphora sentences contained an additional clause in general, BA sentences contain a causal adjunct clause, the two constructions result in different prosodies, the constructions differ in lexical content, etc.). Therefore we will not attempt to draw any conclusions beyond noting that the ATL has been previously implicated in sentence processing (Humphries et al., 2005; Mazoyer et al., 1993; Pallier et al., 2011; Rogalsky & Hickok, 2009) and the angular gyrus in semantic processing (Binder et al., 2009), suggesting that the backward anaphora condition required more of both syntactic and semantic processing, while the frontal areas revealed may reflect increased reliance on working memory and/or cognitive control during the WH condition.
5. Conclusions
While a definitive answer to the question of the specific role of Broca’s area will require additional studies, the results of the present study suggest Broca’s area is more likely to support the mechanisms deployed during the active prediction of gap locations in WH-questions and antecedents in backward anaphora (or a related general cognitive process), and less likely to support specific syntactic operations like movement. This hypothesis is consistent both with the current result that both filler-gap and anaphoric long-distance dependencies result in activation in the pars triangularis, but only if the dependencies involve prediction mechanisms, and with recent studies investigating the role of Broca’s area in syntactic processing (Pallier et al, 2011; Santi & Grodzinsky, 2012). Determining exactly which components of the prediction mechanisms are driving this activation, and whether the components are language-specific or domain-general will require future studies.
An additional avenue to pursue is suggested by Bornkessel-Schlesewsky & Schlesewsky (2013) in a recent review. They suggest that Broca’s area is not involved in sentence comprehension directly, but that it is a mediator between non-sequential processing in the ventral stream and sequential processing in the dorsal stream, and a provider of top-down information to each stream. This is consistent with the claims of Novick et al. (2005) that the function of Broca’s area during sentence comprehension is cognitive control. Within the context of the present experimental findings and distance effects more generally, demands on cognitive control may increase while holding the filler or anaphor in memory and processing additional syntactic/semantic content from the ongoing sentence during the longer conditions. Whether or not different subregions of Broca’s area contribute to sentence processing beyond cognitive control and verbal working memory remains to be investigated. In addition, further research into the contribution of other areas potentially involved in sentence-level processing (e.g., anterior temporal lobe, Mazoyer et al., 1993; Humphries et al., 2001; Rogalsky & Hickok, 2009, Pallier et al., 2011) and the interaction of these networks will be critical to clarify the neural bases of syntax and sentence processing.
Supplementary Material
Highlights.
Measured effects of distance in long distance dependencies: syntactic movement and backward anaphora
Found distance effect for sentences with backward anaphora in anterior Broca’s area
Distance effects for sentences in anterior Broca’s area not specific to syntactic movement
Contradicts previous interpretations that distance effects reflect syntactic movement
Distance effects contingent on ‘active prediction’ during processing
Acknowledgments
This research was supported by NIH grant # DC03681 awarded to Gregory Hickok. We would like to thank Jonathon Venezia for useful suggestions concerning fMRI analysis, and two anonymous reviewers for their comments.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Bahlmann J, Schubotz RI, Friederici AD. Hierarchical artificial grammar processing engages Broca’s area. Neuroimage. 2008;42(2):525–534. doi: 10.1016/j.neuroimage.2008.04.249. [DOI] [PubMed] [Google Scholar]
- Binder JR, Desai RH, Graves WW, Conant LL. Where is the semantic system? A critical review and meta-analysis of 120 functional neuroimaging studies. Cerebral Cortex. 2009;19(12):2767–2796. doi: 10.1093/cercor/bhp055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bornkessel I, Zysset S, Friederici AD, von Cramon DY, Schlesewsky M. Who did what to whom? The neural basis of argument hierarchies during language comprehension. Neuroimage. 2005;26(1):221–233. doi: 10.1016/j.neuroimage.2005.01.032. [DOI] [PubMed] [Google Scholar]
- Bornkessel-Schlesewsky I, Grewe T, Schlesewsky M. Prominence vs. aboutness in sequencing: a functional distinction within the left inferior frontal gyrus. Brain and Language. 2012;120(2):96–107. doi: 10.1016/j.bandl.2010.06.004. [DOI] [PubMed] [Google Scholar]
- Bornkessel-Schlesewsky I, Schlesewsky M. Reconciling time, space and function: A new dorsal-ventral stream model of sentence comprehension. Brain and Language. 2013;125(1):60–76. doi: 10.1016/j.bandl.2013.01.010. [DOI] [PubMed] [Google Scholar]
- Bornkessel-Schlesewsky I, Schlesewsky M, von Cramon DY. Word order and Broca’s region: evidence for a supra-syntactic perspective. Brain and Language. 2009;111(3):125–139. doi: 10.1016/j.bandl.2009.09.004. [DOI] [PubMed] [Google Scholar]
- Buchsbaum BR, Hickok G, Humphries C. Role of left posterior superior temporal gyrus in phonological processing for speech perception and production. Cognitive Science. 2001;25(5):663–678. [Google Scholar]
- Caplan D, Stanczak L, Waters G. Syntactic and thematic constraint effects on blood oxygenation level dependent signal correlates of comprehension of relative clauses. Journal of Cognitive Neuroscience. 2008;20(4):643–656. doi: 10.1162/jocn.2008.20044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chomsky N. Some concepts and consequences of the theory of government and binding. Vol. 6. MIT press; 1982. [Google Scholar]
- Chomsky N. The minimalist program. Vol. 28. Cambridge, MA: MIT press; 1995. [Google Scholar]
- Crain S, Fodor JD. How can grammars help parsers. In: Dowty D, Kartunnen D, Zwicky AM, editors. Natural language parsing: Psycholinguistics, computational, and theoretical perspectives. Cambridge University Press; 1985. pp. 94–129. [Google Scholar]
- Engdahl E. Parasitic gaps. Linguistics and Philosophy. 1983;6(1):5–34. [Google Scholar]
- Embick D, Marantz A, Miyashita Y, O’Neil W, Sakai KL. A syntactic specialization for Broca’s area. Proceedings of the National Academy of Sciences USA. 2000;97(11):6150–6154. doi: 10.1073/pnas.100098897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fiebach CJ, Schlesewsky M, Lohmann G, Von Cramon DY, Friederici AD. Revisiting the role of Broca’s area in sentence processing: syntactic integration versus syntactic working memory. Human brain mapping. 2005;24(2):79–91. doi: 10.1002/hbm.20070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Frazier L, Flores D’Arcais GB. Filler driven parsing: A study of gap filling in Dutch. Journal of memory and language. 1989;28(3):331–344. [Google Scholar]
- Friederici AD. Towards a neural basis of auditory sentence processing. Trends in cognitive sciences. 2002;6(2):78–84. doi: 10.1016/s1364-6613(00)01839-8. [DOI] [PubMed] [Google Scholar]
- Friederici AD, Bahlmann J, Heim S, Schubotz RI, Anwander A. The brain differentiates human and non-human grammars: functional localization and structural connectivity. Procedings of the National Academy of Sciences USA. 2006a;103(7):2458–2463. doi: 10.1073/pnas.0509389103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Friederici AD, Fiebach CJ, Schlesewsky M, Bornkessel ID, von Cramon DY. Processing linguistic complexity and grammaticality in the left frontal cortex. Cerebral Cortex. 2006b;16(12):1709–1717. doi: 10.1093/cercor/bhj106. [DOI] [PubMed] [Google Scholar]
- Friederici AD, Hahne A, Mecklinger A. Temporal structure of syntactic parsing: early and late event-related brain potential effects. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1996;22(5):1219. doi: 10.1037//0278-7393.22.5.1219. [DOI] [PubMed] [Google Scholar]
- Friederici AD, Rüschemeyer SA, Hahne A, Fiebach CJ. The role of left inferior frontal and superior temporal cortex in sentence comprehension: localizing syntactic and semantic processes. Cerebral cortex. 2003;13(2):170–177. doi: 10.1093/cercor/13.2.170. [DOI] [PubMed] [Google Scholar]
- Grodzinsky Y. The neurology of syntax: Language use without Broca’s area. Behavioral and brain sciences. 2000;23(01):1–21. doi: 10.1017/s0140525x00002399. [DOI] [PubMed] [Google Scholar]
- Grodzinsky Y, Friederici AD. Neuroimaging of syntax and syntactic processing. Current Opinions in Neurobiology. 2006;16(2):240–246. doi: 10.1016/j.conb.2006.03.007. [DOI] [PubMed] [Google Scholar]
- Grodzinsky Y, Santi A. The battle for Broca’s region. Trends in Cognitive Sciences. 2008;12(12):474–480. doi: 10.1016/j.tics.2008.09.001. [DOI] [PubMed] [Google Scholar]
- Hagoort P. On Broca, brain, and binding: a new framework. Trends in Cognitive Sciences. 2005;9(9):416–423. doi: 10.1016/j.tics.2005.07.004. [DOI] [PubMed] [Google Scholar]
- Hickok G, Buchsbaum B, Humphries C, Muftuler T. Auditory–motor interaction revealed by fMRI: speech, music, and working memory in area Spt. Journal of Cognitive Neuroscience. 2003;15(5):673–682. doi: 10.1162/089892903322307393. [DOI] [PubMed] [Google Scholar]
- Hickok G, Poeppel D. Dorsal and ventral streams: a framework for understanding aspects of the functional anatomy of language. Cognition. 2004;92(1):67–99. doi: 10.1016/j.cognition.2003.10.011. [DOI] [PubMed] [Google Scholar]
- Hickok G, Poeppel D. The cortical organization of speech processing. Nature Reviews Neuroscience. 2007;8(5):393–402. doi: 10.1038/nrn2113. [DOI] [PubMed] [Google Scholar]
- Humphries C, Willard K, Buchsbaum B, Hickok G. Role of anterior temporal cortex in auditory sentence comprehension: an fMRI study. Neuroreport. 2001;12(8):1749–1752. doi: 10.1097/00001756-200106130-00046. [DOI] [PubMed] [Google Scholar]
- January D, Trueswell JC, Thompson-Schill SL. Co-localization of Stroop and syntactic ambiguity resolution in Broca’s area: Implications for the neural basis of sentence processing. Journal of Cognitive Neuroscience. 2009;21(12):2434–2444. doi: 10.1162/jocn.2008.21179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaan E, Harris A, Gibson E, Holcomb P. The P600 as an index of syntactic integration difficulty. Language and cognitive processes. 2000;15(2):159–201. [Google Scholar]
- Kaan E, Swaab TY. The brain circuitry of syntactic comprehension. Trends in cognitive sciences. 2002;6(8):350–356. doi: 10.1016/s1364-6613(02)01947-2. [DOI] [PubMed] [Google Scholar]
- Kazanina N, Lau EF, Lieberman M, Yoshida M, Phillips C. The effect of syntactic constraints on the processing of backwards anaphora. Journal of Memory and Language. 2007;56(3):384–409. [Google Scholar]
- Makuuchi M, Bahlmann J, Anwander A, Friederici AD. Segregating the core computational faculty of human language from working memory. Proceedings of the National Academy of Sciences USA. 2009;106(20):8362–8367. doi: 10.1073/pnas.0810928106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mazoyer BM, Tzourio N, Frak V, Syrota A, Murayama N, Levrier O, Mehler J. The cortical representation of speech. Journal of Cognitive Neuroscience. 1993;5(4):467–479. doi: 10.1162/jocn.1993.5.4.467. [DOI] [PubMed] [Google Scholar]
- Novick JM, Trueswell JC, Thompson-Schill SL. Cognitive control and parsing: Reexamining the role of Broca’s area in sentence comprehension. Cognitive, Affective, & Behavioral Neuroscience. 2005;5(3):263–281. doi: 10.3758/cabn.5.3.263. [DOI] [PubMed] [Google Scholar]
- Osterhout L, Holcomb PJ. Event-related brain potentials elicited by syntactic anomaly. Journal of memory and language. 1992;31(6):785–806. [Google Scholar]
- Pallier C, Devauchelle AD, Dehaene S. Cortical representation of the constituent structure of sentences. Proceedings of the National Academy of Sciences. 2011;108(6):2522–2527. doi: 10.1073/pnas.1018711108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogalsky C, Hickok G. Selective attention to semantic and syntactic features modulates sentence processing networks in anterior temporal cortex. Cerebral Cortex. 2009;19(4):786–796. doi: 10.1093/cercor/bhn126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogalsky C, Hickok G. The role of Broca’s area in sentence comprehension. Journal of Cognitive Neuroscience. 2011;23(7):1664–1680. doi: 10.1162/jocn.2010.21530. [DOI] [PubMed] [Google Scholar]
- Rogalsky C, Matchin W, Hickok G. Broca’s area, sentence comprehension, and working memory: an fMRI Study. Frontiers in Human Neuroscience. 2008;2:14. doi: 10.3389/neuro.09.014.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ross JR. Constraints on variables in syntax. Bloomington, IN: Indiana University Linguistics Club; 1967. [Google Scholar]
- Santi A, Grodzinsky Y. Working memory and syntax interact in Broca’s area. Neuroimage. 2007a;37(1):8–17. doi: 10.1016/j.neuroimage.2007.04.047. [DOI] [PubMed] [Google Scholar]
- Santi A, Grodzinsky Y. Taxing working memory with syntax: Bihemispheric modulations. Human brain mapping. 2007b;28(11):1089–1097. doi: 10.1002/hbm.20329. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santi A, Grodzinsky Y. fMRI adaptation dissociates syntactic complexity dimensions. Neuroimage. 2010;51(4):1285–1293. doi: 10.1016/j.neuroimage.2010.03.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santi A, Grodzinsky Y. Broca’s area and sentence comprehension: a relationship parasitic on dependency, displacement or predictability? Neuropsychologia. 2012;50(5):821–832. doi: 10.1016/j.neuropsychologia.2012.01.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Snijders TM, Vosse T, Kempen G, Van Berkum JJ, Petersson KM, Hagoort P. Retrieval and unification of syntactic structure in sentence comprehension: an FMRI study using word-category ambiguity. Cerebral Cortex. 2009;19(7):1493–1503. doi: 10.1093/cercor/bhn187. [DOI] [PubMed] [Google Scholar]
- Stowe LA. Parsing WH-constructions: Evidence for on-line gap location. Language and cognitive processes. 1986;1(3):227–245. [Google Scholar]
- Stromswold K, Caplan D, Alpert N, Rauch S. Localization of syntactic comprehension by positron emission tomography. Brain and language. 1996;52(3):452–473. doi: 10.1006/brln.1996.0024. [DOI] [PubMed] [Google Scholar]
- Talairach J, Tournoux P. Co-planar stereotaxic atlas of the human brain. 3-Dimensional proportional system: an approach to cerebral imaging 1988 [Google Scholar]
- Traxler MJ, Pickering MJ. Plausibility and the processing of unbounded dependencies: An eye-tracking study. Journal of Memory and Language. 1996;35(3):454–475. [Google Scholar]
- van Gompel RP, Liversedge SP. The influence of morphological information on cataphoric pronoun assignment. Journal of Experimental Psychology: Learning, Memory, and Cognition. 2003;29(1):128. doi: 10.1037//0278-7393.29.1.128. [DOI] [PubMed] [Google Scholar]
- Wagers M, Phillips C. Multiple dependencies and the role of the grammar in real-time comprehension. Journal of Linguistics. 2009;45(2):395–433. [Google Scholar]
- Wise RJ, Scott SK, Blank SC, Mummery CJ, Murphy K, Warburton EA. Separate neural subsystems within “Wernicke’s area”. Brain. 2001;124(1):83–95. doi: 10.1093/brain/124.1.83. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.