

Abstract
Background:
The neurobiology of neonatal pain processing, especially in preterm infants, differs significantly from older infants, children, adolescence, and adults. Research suggests that strong painful procedures or repeated mild procedures may permanently modify individual pain processing. Acute injuries at critical developmental periods are risk factors for persistent altered neurodevelopment. The purpose of this narrative review is to present the seminal and current literature describing the unique physiological aspects of neonatal pain processing.
Methods:
Articles describing the structures and physiological processes that influence neonatal pain were identified from electronic databases Medline, PubMed, and CINAHL.
Results:
The representation of neonatal pain physiology is described in three processes: Local peripheral nervous system processes, referred to as transduction; spinal cord processing, referred to as transmission and modulation; and supraspinal processing and integration or perception of pain. The consequences of undermanaged pain in preterm infants and neonates are discussed.
Conclusion:
Although the process and pain responses in neonates bear some similarity to processes and pain responses in older infants, children, adolescence, and adults; there are some pain processes and responses that are unique to neonates rendering them at risk for inadequate pain treatment. Moreover, exposure to repeated painful stimuli contributes to adverse long-term physiologic and behavioral sequelae. With the emergence of studies showing that painful experiences are capable of rewiring the adult brain, it is imperative that we treat neonatal pain effectively.
Keywords: Neonate, neurodevelopment, pain, preterm infant, pain processing
INTRODUCTION
Although it has been nearly 27 years since Anand and Hickey's[9] seminal article on neonatal pain established that nociceptive activity constitutes an overwhelming physiologic stress for infants, in some neonatal intensive care units (NICUs), 79.2% of painful procedures are performed without analgesia. Compared with healthy newborns, infants in NICUs experience a median of 75 (range 3-364) painful procedures during hospitalization and 10 (range 0-51) painful procedures per day of hospitalization.[31] Data from preclinical trials and pediatric research demonstrate noxious stimuli during this vulnerable period of neuronal plasticity may trigger unpredicted long-term epigenomic changes that affect the brain, neurodevelopment, and pain reactivity into adulthood.[39,67,68,91,107] This review will present the unique physiological aspects of neonatal pain that influence pain processing in this vulnerable population.
EXPOSURE TO PAIN EARLY IN LIFE
Research suggests that strong painful procedures[90,116] or mild repeated procedures[68] may permanently modify individual pain processing. Acute injury at critical developmental periods are risk factors for persistent altered neurodevelopment.[42] During normal development, infant pain transmission and pain modulation undergo rapid growth beginning at 22 weeks gestation; achieving mature functioning at approximately 2 months of age.[4,120,142] Noxious stimuli during this vulnerable period of neuronal plasticity may trigger unpredicted long-term epigenetic changes, which affect the brain, neurodevelopment, pain modulation, and pain reactivity into adulthood.[67,68,91,94,107]
Extensive preclinical trials have established the physiological impact of early exposure to noxious stimuli on the developing nervous system.[23,97,126,146] Combined with the mounting clinical evidence,[16,144] it is possible that exposure to acute and repetitive pain in infants may have long-term effects contributing to persistent adverse neurological outcomes. The purpose of this narrative review is to present the seminal and current literature describing the unique physiological aspects of neonatal pain processing. The review will answer the following research question: Among neonates, what unique structures and processes influence neonatal pain processing?
ELEMENTS OF PAIN PERCEPTION
Pain is a perception, not a sensation. It involves sensitivity to chemical changes in the tissues and interpretation that such changes are harmful. This perception is real, whether the harm has occurred in the past or is happening in the present. Cognition is involved in the formation of this perception. There are emotional consequences and behavioral responses to the cognitive, emotional and physiological aspects of pain.[104]
Pain is a linguistic description for an expansive variety of experiences and responses. It is an abstract representation of the information that is subsequently reexamined over long periods by the entire somesthetic system.[104] The physiologic mechanisms involved in the phenomenon of pain are known as nociception. The term nociception refers to the ability of a nerve to detect noxious stimuli and transmit the information concerning the stimuli to the brain for interpretation.[101] Unlike the phenomenon pain, nociception does not require self-report. Therefore, the term nociception provides an accurate description of an infant's response to a tissue-damaging stimulus. In the literature, the terms pain and nociception are used interchangeably; in this review, no further distinction between the two terms will be made.
Pain responses are integral components of an adaptive biologic system that enables a newborn to function in a dynamic, challenging, and potentially dangerous environment. These responses represent reactions, modulations, and integration by the peripheral nervous system (somatosensory, somatomotor, autonomic), spinal cord, and brain (brain stem, medulla, hypothalamus, thalamus, limbic system, cranial nerves, and the neocortex). Taken together, the responses are concurrent reactions of pain perception; an experience of emotions and autonomic, somatomotor and endocrine responses rather than sequential reactions.[85] This description of concurrent responses to pain and stress is a very important differentiation from past discussions of pain-response physiology. Individuals respond to and communicate pain simultaneously in several different areas, using more than just the ascending and descending pain pathways for communication.[103]
DEVELOPMENTAL NEUROBIOLOGY OF NEONATAL PAIN
Simply expressed, pain is a three-neuron relay that detects sensations in the periphery and conveys the sensations via second and third order neurons through the spinal cord, brainstem, and thalamic relay nuclei to the cerebral cortex.[22] Nociceptive neurons are sensitive to thermal, mechanical, or chemical noxious stimuli. They contain and release neuropeptides and are sensitive to particular growth hormones involved in neurogenic inflammation (e.g. vasodilation, vascular leakage) and neuroimmune regulation. Nociceptive neurons also influence smooth muscle contractions and glandular secretion into the gastrointestinal and urinary tracts.[10]
This representation of neonatal pain physiology is organized in three sections:
Local peripheral nervous system processes or transduction occurs when noxious stimuli are translated into neuronal action potential at the nociceptors, which are the sensory endings of the primary afferent neurons in the periphery
Spinal cord processing, referred to as transmission and modulation, is the propagation of action potentials along ascending pathways from the site of transduction throughout the sensory nervous system to the spinal cord, then centrally to the brain; and activation of descending pathways that exert inhibitory effects on the synaptic transmission of noxious stimuli
Supraspinal processing and integration of pain or perception of pain is the result of neural processing: Recognition, defining and responding to noxious stimuli in the brain.[19,104,105,106]
The discussion is presented sequentially for clarity; however, processes are interactive and occur concurrently.
Maturation of local peripheral nervous system responses or transduction
The peripheral nervous system, part of the somatosensory system, consists of three primary afferent fibers, Aδ (thinly myelinated, mechanosensitive pain receptors), Aβ, and C-polymodal fibers (unmyelinated, mechanical, chemical, and thermal sensitive pain receptors)[21,93] [Table 1]. During the sixth week of gestation, synaptic development between sensory fibers and interneurons in the dorsal horn of the spinal cord begin to appear. By week 7 of gestation, cutaneous sensory receptors appear in the perioral area. By the 11th week, cutaneous receptors have spread to the rest of the face, the palms of the hands, and the soles of the feet, by the 15th week to the trunk and proximal parts of the arms and legs, and by the 20th week to all cutaneous and mucous surfaces.[4] By 24 weeks gestation, the peripheral nervous system is developmentally mature and functional. However, in contrast to adults, neonates have a higher density of high threshold Aδ and low threshold Aβ mechanoreceptors/cm2 that respond with lower firing frequency.[33,66] Tactile and noxious stimuli evoke cutaneous limb withdrawal in neonates as young as 27 weeks gestation.[17,56]
Table 1.
Primary afferent fibers[93]

Tissue injury initiates a cascade of peripheral neuronal responses. Noxious stimuli are transduced to electrical activity at the peripheral terminals of Aδ and C polymodal fibers and are immediately conducted to the dorsal horn of the spinal cord. The cellular and blood vessel damage from the injury along with inflammatory and tumor cells, release biochemical mediators (bradykinin, calcium and potassium ions, substance P, and prostaglandins) that activate or sensitize Aδ and C polymodal afferent nociceptors that transmit pain impulses to the spinal cord and stimulate local inflammatory flare and wheal responses. Concurrently, substance P and prostaglandins increase the local tissue inflammation creating local primary hyperalgesia.[123] With repeated tissue damage, such as repeated heel lance, the associated inflammation and tenderness may extend into adjacent uninjured tissue giving rise to allodynia[121] and a 50% lower cutaneous flexor reflex threshold compared with the intact contralateral heel.[57]
In addition to hyperalgesia, tissue damage in early life causes profound and persistent dendritic sprouting in the local sensory nerve terminals. Compared with the older infant, sprouting is more prominent when tissue damage occurs at birth or shortly thereafter.[50] Behavioral studies show the subsequent lowered mechanical threshold and hyperinnervation of the injured area persists into adulthood.[119]
In the past, lack of myelination was used to support the argument that the premature infant's nervous systems was immature[60,61,139] and therefore the infant was not capable of experiencing pain.[122] However, in the peripheral nerves of adults, nociceptive impulses are transmitted through C-polymodal and A-δ fibers.[102] Incomplete neonatal nerve myelination results in a lengthier conduction velocity, but is fully compensated by the shorter interneuron and neuromuscular distances traveled by the nerve impulse.[125] Spinal and central nervous system (CNS) nociceptive nerve tracts establish myelination during the second and third trimesters of gestation. Ascending nociceptive pathways to the brain stem and thalamus are completely myelinated by 30 weeks; thalamocortical pain fibers in the posterior limb of the internal capsule and corona radiata are myelinated by 37 weeks.[65]
Maturation of spinal cord processing or transmission and modulation
Early in life, the spinal cord of the neonate's immature nervous system functions as an independent unit. Because of the immature descending pathway, the neonatal cortex has little control over pain processes. Biobehavioral pain responses to noxious stimuli are a series of decorticate spinal reflexes. As the cortex assumes control, time, experience, and maturity integrate immature reflexes into sophisticated adult behavior patterns.[48]
The spinal cord level has three important nociceptive functions; (1) local responses, which are often reflexive protective; (2) ascending pain transmission, and (3) modulation of nociceptive impulses through descending pain pathways. A detailed description of the anatomy and physiology of the CNS is not provided. The reader is referred to standard neurobiology text for a detailed review.[21]
Local spinal cord responses
Within the spinal cord, glutamate and tachykinin stimulate N-methyl-D-aspartate (NMDA) and tachykinin receptors mediating nociceptive transmission. NMDA receptors are thought to be responsible for the central sensitization or “wind-up phenomenon” where sensory inputs to the CNS are amplified, resulting in an alteration of the CNS and increased pain.[38] All laminae in the neonatal dorsal horn are uniformly labeled with NMDA sensitive glutamate until day 10-12 after birth, after which higher densities concentrate in the substantia gelatinosa.[49] Spreading to inferior and superior spinal level receptors, the neuronal excitability innervates the area of tissue damage (central sensitization).[50,56] The increased excitability of nociceptive receptors in the dorsal horn (“wind-up”) also causes secondary hyperalgesia in the normal tissue surrounding the site of injury.[112] In addition, nonnociceptive input from the opposite limb produces infant pain.
The NMDA receptor fields of dorsal horn cells in infants are larger than adult fields until 42 weeks gestation, then decline to adult size by 43-44 weeks gestation.[49] This increased expression of NMDA receptors in the dorsal horn of the spinal cord accentuates the low pain threshold of preterm infants and is thought to be associated with the increased vulnerability of excitotoxic damage in the newborn brain[92] resulting in stronger and more prolonged pain for the infant.[20,45] NMDA dependent C fiber-evoked depolarization of spinal cord cells and “wind-up” of cells to repeated C fiber stimulation has been demonstrated in the young (8-14 day) spinal cord in vitro and observed in both preterm and term neonates exposed to sequential painful procedures.[130]
In adults, γ-aminobutyric acid (GABA) inhibits the excitatory activity of glutamate, but in infants, GABA induces depolarization dependant, in part, on the intracellular concentration of chloride.[25] GABA is over expressed in infants until 44 weeks.[40] The enlarged NMDA receptor fields and the levels of immature spinal GABA signaling contribute to the nociceptive hypersensitivity in infants. The result-pain responses are elicited with less invasive stimuli.[55]
These local spinal cord responses have a profound effect on a neonate's biobehavioral response to stimulation. Compared with full-term infants, children, adolescence, and adults, preterm infants have a lower threshold and a more pronounced reflex response to touch.[46] The decreased pain threshold makes the infant more sensitive to noxious stimuli such that touch stimulation near the injured area causes intense pain for days or weeks.[56] With repeated exposure to noxious stimuli or touch, the lowered threshold declines even further due to the influence of NMDA and GABA on the excitability of the sensory neurons of the spinal cord. The significant variability in pain response observed in neonates is due to the continually decreasing touch threshold, the increasing neuronal sensitization and the preceding painful/stressful interventions in the last hour,[81] 24 h,[71] or cumulatively since birth.[70,87] The clinical implication for tiny neonates is that compared with adults, the behavioral response to routine care will be the same as the behavioral response to an invasive procedure. In addition, a diaper change can elicit pain-like behaviors and physiologic responses if preceded by a heel lance 30 min before.[82] Depending on the gestational age of the infant, the number of prior painful experience, the infant's behavioral state, or acuity of illness, a single stimulus may elicit a pain response that lasts several minutes or cause no reaction at all.[48,53,114]
The affinity of the NMDA receptor decreases with postnatal age. NMDA-evoked calcium influx in rat substantia gelatinosa neurons is very high in the first postnatal week then declines to adult levels by 6-8 weeks postnatally.[83] The amount of current associated with immature NMDA receptors is initially much greater in neonates compared with adults and declines with age and synaptic activity. This is due to a developmental switch in the subunit composition of the NMDA receptor.[62] Glutamatergic synaptic currents undergo a characteristic pattern of maturation and development. This pattern involves changes in the kinetic of NMDA receptor currents and the formation of “silent synapses” that initially only express NMDA currents and are only later made functional by the addition of AMPA receptor currents. This allows functional networks to be adaptively modified by experience.[44]
Ascending transmission
There are a considerable number of seminal rigorously executed studies[4,51,72] and recent reviews[68,129,133] demonstrating even the smallest infants are capable of mounting a response to noxious stimuli. In neonates, the ascending nociceptive pathways are developmentally mature by 20 weeks gestation.[4,48] By 30 weeks gestation, the ascending pathway of an neonate is capable of the same functionality as an adult.[47]
Research has established that facial expressions and body movements represent evidence-based acute pain behavioral variables in infants. Brow bulge, eye squeeze, and a deepening of the nasolabial furrow have been exhibited in infants as young as 26 weeks’ gestation in response to a heel lance procedures.[37,89] The same expression is associated with pain in adults, although in infants younger than 30 weeks’ gestation, the response is not as robust or universal. Heart rate, heart rate variability (HRV), and oxygen saturation are physiologic variables associated with acute pain in infants.[78] These protective autonomic responses and facial responses triggered by the ascending pain fiber connecting with the reticular activating system and the periaqueductal gray (PAG) area are not dependent on cortical input.[37,89]
Descending transmission, modulation of pain
Descending inhibitory controls are immature at birth.[45] Descending inhibitory pathways progress downward from the brainstem via the dorsolateral funiculus of the spinal cord to the dorsal horn in fetal life. Once noxious transmission and pain perception has occurred, fibers in the spinothalamic tract stimulate regions of the midbrain that send descending projections to the dorsal horn to modulate pain impulses. However, these inhibitory pathways do not extend collateral branches into the dorsal horn for some time and do not become functionally effective until P10 in rats.[54] This delay may be due to deferred expression of serotonin and noradrenaline or the immaturity of crucial interneurons. Interneuron maturation in the substantia gelatinosa occurs largely in the postnatal period and appears to be particularly important in the modulation of nociceptive stimuli.[27] Since the immature endogenous analgesic system cannot diminish noxious inputs as they enter the CNS noxious inputs are more profound in infants compared with adults. This mechanism also explains why the stimulus-produced analgesia from the PAG is not effective until P21 in rats.[141]
Neurotransmitters are essential components of adult and neonatal pain transmission. Adult and neonatal pain transmission is mediated at the level of the spinal cord by the neurotransmitters substance P, somatostatin, calcitonin gene-related peptide, vasoactive intestinal polypeptide, and glutamate. Modulation of pain transmission occurs through the local release of endogenous opioids, enkephalin or serotonin, norepinephrine, acetylcholine, neurotensin, and GABA, glycine, and dopamine from the PAG area.
GABA has a crucial role in preventing the spread of excitatory glutamatergic activity. In the adult spinal cord, GABA is an inhibitory amino acid transmitter that produces membrane hyperpolarization through the activation of postsynaptic GABAA and GABAB receptors and depresses transmitter release acting through presynaptic GABAB receptors. However in neonates, GABA is transiently overexpressed in the developing spinal cord. In 90% of embryonic dorsal horn neurons cultured for more than a week, both GABA and glycine induced an increased calcium and cellular depolarization.[117] This effect decreased with age in culture so that by 30 days it was absent and these agents caused hyperpolarization.[147] During the first two postnatal weeks, the expression of the GABA-synthesizing enzyme, glutamate decarboxylase (GAD), indicates 50% of neurons are GABA-positive compared with 20% GABA-positive neurons in the third postnatal week.[138] This phenomenon where GABA mediates most of the excitatory drive in the immature brain also occurs in the supraspinal area of the postnatal rat brain.[26]
In the preterm infant, dopamine and norepinephrine are not available to modulate nociceptive activity before 36-40 weeks’ gestation. Moreover, the inhibitory fibers extending from the PAG area and other areas in the brainstem do not release serotonin until approximately 6-8 weeks after birth.[8,50,100] Because the afferent excitatory pain neurotransmitters, which are plentiful at birth, are not balanced by descending inhibitory neurotransmitters, preterm infants have limited ability to modulate pain. The immaturity of the descending pathway exposes neonates to greater pain sensitivity and intensity before 48 weeks gestation compared with older infants and adults.[6,59]
The maturation of C fiber synaptic connections in the dorsal horn, interneuronal development in the substantia gelatinosa, and the functional development of descending inhibitory systems from the supraspinal centers occur postnatally in the rat. Modulatory mechanisms reach maturation later than the basic excitatory mechanism from low-threshold inputs; thus, a newborn infant can mount a response to painful stimuli. This response, however, may not always be predictable or well organized. Lack of inhibition contributes to exaggerated and generalized responses to all low and high thresholds of sensory inputs, whereas specific pain responses may require convergent afferent inputs building up over time to become clinically apparent.[49] The onset of the inhibitory processes is a crucial determinant of the neuron's firing activity and ultimately signals the emergence of maturing pain responses in infants.
Supraspinal processes and integration
At 8 weeks gestation, the fetal neocortex begins to development and by 20 weeks, each cortex has a full complement of 109 neurons.[99] Afferent neurons in the thalamus produce axons that arrive in the cerebrum before mid-gestation. These fibers “linger” just below the neocortex until migration and dendritic arborization of cortical neurons are complete and the establishment of intracortical synaptic connections between 20 weeks of gestation is complete.[113] By 24-26 weeks, incoming thalamocortical fibers and synaptic connections are completed.[4] Somatosensory-evoked potentials are slow and simple before 29 weeks’ gestation; however, by 40 weeks, the pattern is complex and latency is short. The cerebral cortex is functionally mature (including the sensorimotor cortex, limbic system, diencephalon, thalamus, midbrain brainstem regions) by 22 weeks’ gestation and bilaterally synchronous by 27 weeks.[4,110,132]
Cortical cell migration from the germinal lining of the ventricles in which they originate to specific locations in the cortical plate is complete at approximately 24 weeks’ gestation.[2] The support structure of the germinal matrix is still highly vascular after the completion of cell migration until 28 weeks’ gestation, making the support structure extremely vulnerable to hemorrhage.[143] During the process of migration and differentiation, apoptosis or programmed cell death eliminates a large number of neurons from diverse areas of the cerebral cortex.[15] The number of cortical neurons reaches the maximum at 28 weeks’ gestation and then declines by approximately 70% before birth.[111]
At 20 weeks gestation, intermittent nonspecific electroencephalographic bursts are seen in both cerebral hemispheres. They become sustained at 22 weeks and bilaterally synchronous at 26-27 weeks.[131] These nonspecific neuronal bursts are present during neuronal development. Their disappearance signals the emergence of specific potentials and the maturation of functional brain circuitry.[28,34] Fabrizi et al.[43] demonstrated a critical crossover period in brain response following tactile and noxious stimulation from nonspecific, evenly dispersed neuronal bursts to modality-specific, localized, evoked potentials. Their results suggest that specific neural circuits necessary for discrimination between touch and nociception emerge from 35 to 37 weeks gestation in the human. Table 2 summarizes the clinical implications of the maturation of pain pathways.
Table 2.
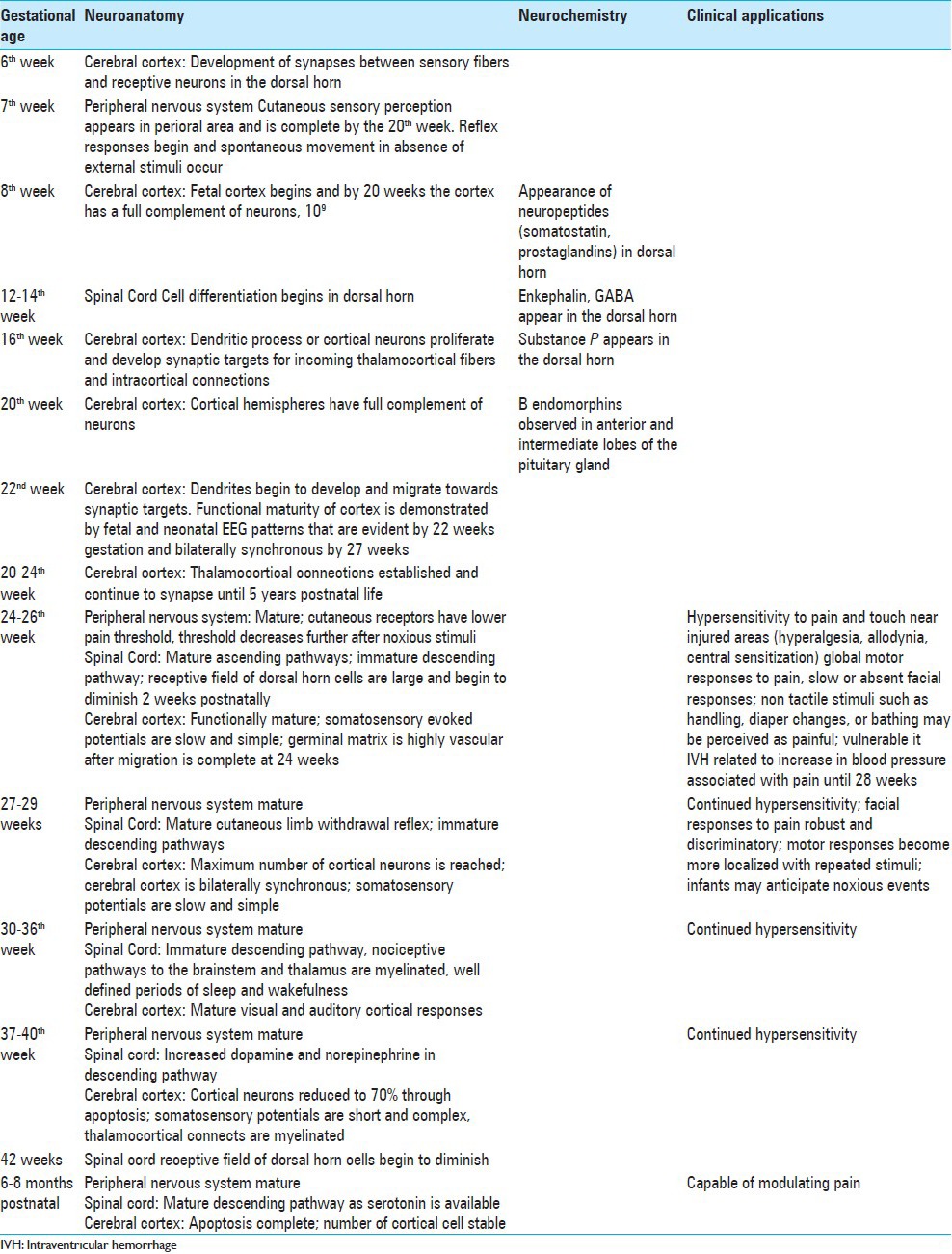
CONSEQUENCE OF UNDERMANAGED PAIN
Multiple lines of evidence suggest that repeated and prolonged pain exposure early in life alters an infant's subsequent pain processing, long-term development, later pain sensitivity and may contribute to the transition from acute to chronic pain.[29,68,79,128] Indeed, undermanaged acute pain early in life is considered to be the greatest risk factor for the development of chronic pain in children and adults.[98,124]
Undermanaged pain in neonates leads to significant short- and long-term adverse consequences.[4,14,58] Short-term consequences of painful procedures consist of decreased oxygen saturation,[140,148] significantly lower partial pressures of oxygen,[115] increased heart rate,[63,64] increased HRV,[18] rapid fluctuations in intracranial pressure,[135] and increased levels of plasma cortisol, aldosterone, growth hormone, catecholamines, and glucagon.[4,9] Physiologic responses pose a problem for preterm neonates because these responses deplete the inadequate energy stores of the preterm infant increasing the infant's chance of morbidity and mortality.[4,9,88]
Physical and psychological stress increases the opportunity for infection through generalized depression of the immune system.[5] A compromised immune system is a neonatal challenge because the immaturity of the neonatal immune system, the lack of immunoglobins, decreased energy reserves and limited to exposure to infectious agents[5,30,118,134] are associated with an increased incidence of sepsis. Emerging research has focused on the interaction between the nociceptive pathway and immune function suggesting effective analgesic strategies may be utilized to modulate immune function and decrease infection.[32]
It is hypothesized that the long-term consequences of undermanaged pain in neonates are linked to the plasticity of the neonate's nervous system. Changes in the neonate's peripheral and CNS are innate and critical for normal fetal and neonatal development.[29] Data demonstrate that acute injury at critical developmental periods are risk factors for altered neurodevelopment.[42] During normal development, infant pain transmission and pain modulation undergo rapid growth beginning at 22 weeks gestation achieving mature functioning at approximately 2 months of age.[4,120,142] Noxious stimuli during this vulnerable period of neuronal plasticity may trigger unpredicted long-term epigenetic changes, which affects brain, neurodevelopment, pain modulation, and pain reactivity into adulthood.[67,68,91,94,107]
Findings from cohort and crosssectional studies explain how repeated painful procedures are associated with long-term adverse events such as enhanced perceptual sensitization,[80] blunting of the hypothalamic-pituitary-adrenal (HPA) axis response,[86] greater distress during subsequent surgery requiring higher fentanyl dosing,[108] lower thresholds for withdrawal responses that persist for at least the first year of life,[1] lower cognitive and motor development at 8 and 18 months,[69] increased sensitivity to childhood injuries[74] and higher incidence of somatic complaints.[73,75,76] Other evidence suggests that routine morphine administration in ventilated neonates has no beneficial effects on pain expression, may result in adverse neurological outcomes,[13,24] and contributes to negative effects on cognitive functioning at 5 years of age.[41]
A common feature of these studies is that the tissue injury occurred at a critical period of development and the adverse effects of the injury outlasted the injury itself. The long-term effects of the injury were adolescents and adults with altered pain sensitivity compared with controls. In addition, the area surrounding the injury retained the increased sensitivity to pain so that a new injury in that area resulted in enhance hyperalgesia, which is greater in magnitude and significantly more prolonged than controls. The altered sensory and pain sensitivity outcomes reflected in these studies demonstrate that early painful injuries result in long-term local and global alteration in sensory and pain processing.[126]
Epigenetic modifications
Epigenetic changes in the neonate's peripheral and CNS are innate and critical for normal fetal and neonatal development.[29] During gestation, neuronal tissues acquire specific patterns of DNA methylation to regulate genome function.[96] Increased methylation inhibits gene expression, decreased methylation increases gene expression. The timing of the injury is significant as epigenetic changes are more pronounced during rapid changes in an infant's environment.[127]
In contrast to the genetic code, covalent modification of DNA by methylation is influenced by injury.[127,136] Acute injury at critical developmental periods are risk factors for chronic pain.[42] An individual's gene expression profile changes rapidly after an injury.[95] Post injury, over 1000 genes are activated within the dorsal root ganglion (DRG).[77] Noxious stimuli during this vulnerable period of neuronal plasticity may trigger unpredicted long-term epigenetic changes. Since environmental influences on pain severity are linked to early-life stress,[67,91,98,107] the importance of mitigating abnormal stimulation during the newborn period cannot be overemphasized.
CONCLUSION
Although pain responses in neonates bear some similarity to pain responses in older infants, children, adolescence, and adults; there are some pain responses that are unique to neonates rendering them at risk for inadequate pain treatment.[11,17,35,36,49,52,89,145] Neonates have mature and functional ascending pain pathways capable of transmitting noxious impulses by 24 weeks gestation. However, the neurotransmitters that modulate the ascending impulse do not appear in the descending pathway until approximately 48 weeks gestation.[50,100,109] The immature descending pain pathway exposes preterm infants and neonates to a greater intensity of pain for a prolonged period of time. This increased magnitude and protracted duration of pain is associated with increased peripheral and DRG in sprouting,[119] hyperinnervation of the area adjacent to the tissue injury,[119] extensive escalations in spinal cord neuronal excitability resulting in secondary hyperalgesia of uninjured tissue,[5,138] and a 30-50% lower pain threshold in the presence of repeated noxious stimulation compared with adults.[57]
Exposure to repeated painful stimuli contributes to adverse long-term physiologic and behavioral sequelae. The short-term consequences of undermanaged pain in neonates are behavioral and physiologic responses to noxious stimuli. The long-term consequences of repeated painful procedures vary with gestational age[119,137] and include permanent structural changes in the neonate's nervous system,[3,7,12,15,109] decreased sensitivity childhood injuries[74] and higher incidence of somatic complaints.[73,75,76] Painful procedures should be treated with behavioral and environmental as well as pharmacological interventions to reduce the risk of long-term adverse events. With the emergence of studies showing that painful experiences are capable of rewiring the adult brain, it is imperative we treat neonatal pain effectively.
Footnotes
Available FREE in open access from: http://www.surgicalneurologyint.com/text.asp?2014/5/14/479/144630
REFERENCES
- 1.Abdulkader HM, Freer Y, Garry EM, Fleetwood-Walker SM, McIntosh N. Prematurity and neonatal noxious events exert lasting effects on infant pain behaviour. Early Hum Dev. 2008;84:351–5. doi: 10.1016/j.earlhumdev.2007.09.018. [DOI] [PubMed] [Google Scholar]
- 2.Als H. Reading the premature infant. In: Goldson E, editor. Nurturing the Premature Infant: Developmental Interventions in the Neonatal Intensive Care Nursery. New York, NY: Oxford University Press; 1999. pp. 18–85. [Google Scholar]
- 3.Anand KJ, Coskun V, Thrivikraman KV, Nemeroff CB, Plotsky PM. Long-term behavioral effects of repetitive pain in neonatal rat pups. Physiol Behav. 1999;66:627–37. doi: 10.1016/s0031-9384(98)00338-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Anand KJ, Hickey PR. Pain and its effects in the human neonate and fetus. N Engl J Med. 1987;317:1321–9. doi: 10.1056/NEJM198711193172105. [DOI] [PubMed] [Google Scholar]
- 5.Anand KJ. The applied physiology of pain. In: Anand K, McGrath P, editors. Pain in Neonates. Amsterdam, The Netherlands: Elsevier Science; 1993. pp. 39–66. [Google Scholar]
- 6.Anand KJ. Clinical importance of pain and stress in preterm neonates. Biol Neonate. 1998;73:1–9. doi: 10.1159/000013953. [DOI] [PubMed] [Google Scholar]
- 7.Anand KJ. Effects of perinatal pain and stress. Prog Brain Res. 2000;122:117–29. doi: 10.1016/s0079-6123(08)62134-2. [DOI] [PubMed] [Google Scholar]
- 8.Anand KJ, editor. Developmental Interventions in Neonatal Care Contemporary Forums. Chicago, IL: 1995. Nov 8-11, Immediate and long-term effects of pain on neonatal brain development. [Google Scholar]
- 9.Anand KJ, Brown MJ, Causon RC, Christofides ND, Bloom SR, Aynsley-Green A. Can the human neonate mount an endocrine and metabolic response to surgery? J Pediatr Surg. 1985;20:41–8. doi: 10.1016/s0022-3468(85)80390-0. [DOI] [PubMed] [Google Scholar]
- 10.Anand KJ, Carr D. The neuroanatomy, neurophysiology, and neurochemistry of pain, stress and analgesia in newborns and children. Pediatr Clin North Am. 1989;36:795–822. doi: 10.1016/s0031-3955(16)36722-0. [DOI] [PubMed] [Google Scholar]
- 11.Anand KJ, Coskun V, Thrivikraman KV, Nemeroff CB, Poltsky PM. Long-term behavioral effects of repetitive pain in neonatal rat pups. Physiol Behav. 1999;66:627–37. doi: 10.1016/s0031-9384(98)00338-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Anand KJ, Grunau RE, Oberlander TF. Developmental character and long-term consequences of pain in infants and children. Child Adolesc Psychiatr Clin N Am. 1997;6:703–23. [Google Scholar]
- 13.Anand KJ, Hall RW, Desai N, Shephard B, Bergqvist LL, Young TE, et al. Effects of morphine analgesia in ventilated preterm neonates: Primary outcomes from the NEOPAIN randomised trial. Lancet. 2004;363:1673–82. doi: 10.1016/S0140-6736(04)16251-X. [DOI] [PubMed] [Google Scholar]
- 14.Anand KJ, Hickey PR. Halothane-morphine compared with high dose sufentanil for anesthesia and postoperative analgesia in neonatal cardiac surgery. N Engl J Med. 1992;326:1–9. doi: 10.1056/NEJM199201023260101. [DOI] [PubMed] [Google Scholar]
- 15.Anand KJ, Scalzo FM. Can adverse neonatal experiences alter brain development and subsequent behavior? Biol Neonate. 2000;77:69–82. doi: 10.1159/000014197. [DOI] [PubMed] [Google Scholar]
- 16.Andreae MH, Andreae DA. Regional anaesthesia to prevent chronic pain after surgery: A Cochrane systematic review and meta-analysis. Br J Anaesth. 2013;111:711–20. doi: 10.1093/bja/aet213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Andrews K, Fitzgerald M. The cutaneous withdrawal reflex in human neonates: Sensitization, receptive fields and effects of contra lateral stimulation. Pain. 1994;56:95–101. doi: 10.1016/0304-3959(94)90154-6. [DOI] [PubMed] [Google Scholar]
- 18.Appelhans BM, Luecken LJ. Heart rate variability and pain: Associations of two interrelated homeostatic processes. Biol Psychol. 2008;77:174–82. doi: 10.1016/j.biopsycho.2007.10.004. [DOI] [PubMed] [Google Scholar]
- 19.Banasik J. Pain. In: Copstead LC, Banasik JL, editors. Pathophysiology: Biological and Behavioral Perspectives. 2nd ed. Philadelphia, PA: W. B. Saunders; 2000. pp. 1080–92. [Google Scholar]
- 20.Basbaum A. Memories of pain. Sci Med. 1996;3:22. [Google Scholar]
- 21.Basbaum A, Jessell T. Pain. In: Kandel ER, Schwartz JH, Jessell TM, Siegelbaum SA, Hudspeth AJ, editors. Principles of Neural Science. 5th ed. New York, NY: McGraw-Hill Professional; 2012. pp. 530–55. [Google Scholar]
- 22.Basbaum AI, Fields HL. Endogenous pain control systems: Brainstem spinal pathways and endorphin circuitry. Annu Rev Neurosci. 1984;7:309–38. doi: 10.1146/annurev.ne.07.030184.001521. [DOI] [PubMed] [Google Scholar]
- 23.Beggs S, Currie G, Salter MW, Fitzgerald M, Walker SM. Priming of adult pain responses by neonatal pain experience: Maintenance by central neuroimmune activity. Brain. 2012;135:404–17. doi: 10.1093/brain/awr288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Bellu R, de Waal K, Zanini R. Opioids for neonates receiving mechanical ventilation: A systematic review and meta-analysis. Arch Dis Child Fetal Neonatal Ed. 2010;95:F241–51. doi: 10.1136/adc.2008.150318. [DOI] [PubMed] [Google Scholar]
- 25.Ben-Ari Y, Khalilov I, Kahle KT, Cherubini E. The GABA Excitatory/Inhibitory Shift in Brain Maturation and Neurological Disorders. Neuroscientist. 2012;18:467–86. doi: 10.1177/1073858412438697. [DOI] [PubMed] [Google Scholar]
- 26.Ben-Ari Y, Khazipov R, Leinekugel X, Caillard O, Gaiarsa JL. GABAA, NMDA, and AMDA receptors: A developmentally regulated ‘ménage a trois’. Trends Neurosci. 1997;20:523–9. doi: 10.1016/s0166-2236(97)01147-8. [DOI] [PubMed] [Google Scholar]
- 27.Bicknell HR, Beal JA. Axonal and dendrite development of substantia gelatinosa neurons in the lumbosacral spinal cord of the rat. J Comp Neurol. 1984;226:508–22. doi: 10.1002/cne.902260406. [DOI] [PubMed] [Google Scholar]
- 28.Blankenship AG, Feller MB. Mechanisms underlying spontaneous patterned activity in developing neural circuits. Nat Rev Neurosci. 2010;11:18–29. doi: 10.1038/nrn2759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Buchheit T, Van de Ven T, Shaw A. Epigenetics and the Transition from Acute to Chronic Pain. Pain Med. 2012;13:1474–90. doi: 10.1111/j.1526-4637.2012.01488.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Calvo M, Dawes JM, Bennett DL. The role of the immune system in the generation of neuropathic pain. Lancet Neurol. 2012;11:629–42. doi: 10.1016/S1474-4422(12)70134-5. [DOI] [PubMed] [Google Scholar]
- 31.Carbajal R, Rousset A, Danan C, Coquery S, Nolent P, Ducrocq S, et al. Epidemiology and treatment of painful procedures in neonates in intensive care units. JAMA. 2008;300:60–70. doi: 10.1001/jama.300.1.60. [DOI] [PubMed] [Google Scholar]
- 32.Chiu IM, Heesters BA, Ghasemlou N, Von Hehn CA, Zhao F, Tran J, et al. Bacteria activate sensory neurons that modulate pain and inflammation. Nature. 2013;501:52–7. doi: 10.1038/nature12479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Choonara I. Management of pain in newborn infants. Semin Perinatol. 1992;16:32–40. [PubMed] [Google Scholar]
- 34.Colonnese MT, Kaminska A, Minlebaev M, Milh M, Bloem B, Lescure S, et al. A conserved switch in sensory processing prepares developing neocortex for vision. Neuron. 2010;67:480–98. doi: 10.1016/j.neuron.2010.07.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Coskun V, Anand KJ. Development of supraspinal pain processing. In: Anand KJS, McGrath PJ, editors. Pain in Neonates. 2nd ed. Amsterdam, The Netherlands: Elsevier Science; 2000. pp. 23–54. [Google Scholar]
- 36.Craig K, Grunau RV. Neonatal pain perception and behavioral measurement. In: Anand KJ, Stevens BJ, McGrath PJ, editors. Pain in Neonates. Amsterdam, The Netherlands: Elsevier Science; 1993. pp. 67–105. [Google Scholar]
- 37.Craig K, Prkachin K, Grunau RV. The facial expression of pain. In: Turk D, editor. Handbook of Pain Assessment. New York, NY: Guilford Press; 1992. [Google Scholar]
- 38.Cui JG, Meyerson BA, Lindeoth B. Opposite effects of spinal cord stimulation in different phases of carrageenan-induced hyperalgesia. Eur J Pain. 1999;3:365–74. doi: 10.1053/eujp.1999.0145. [DOI] [PubMed] [Google Scholar]
- 39.Davis DA, Luecken LJ, Zautra AJ. Are reports of childhood abuse related to the experience of chronic pain in adulthood?: A meta-analytic review of the literature. Clin J Pain. 2005;21:398–405. doi: 10.1097/01.ajp.0000149795.08746.31. [DOI] [PubMed] [Google Scholar]
- 40.de Graaf-Peters VB, Hadders-Algra M. Ontogeny of the human central nervous system: What is happening when? Early Hum Dev. 2006;82:257–66. doi: 10.1016/j.earlhumdev.2005.10.013. [DOI] [PubMed] [Google Scholar]
- 41.de Graaf J, van Lingen RA, Simons SH, Anand KJ, Duivenvoorden HJ, Weisglas-Kuperus N, et al. Long-term effects of routine morphine infusion in mechanically ventilated neonates on children's functioning: Five-year follow-up of a randomized controlled trial. Pain. 2011;152:1391–7. doi: 10.1016/j.pain.2011.02.017. [DOI] [PubMed] [Google Scholar]
- 42.Denk F, McMahon SB, Tracey I. Pain vulnerability: A neurobiological perspective. Nat Neurosci. 2014;17:192–200. doi: 10.1038/nn.3628. [DOI] [PubMed] [Google Scholar]
- 43.Fabrizi L, Slater R, Worley A, Meek J, Boyd S, Olhede S, et al. A shift in sensory processing that enables the developing human brain to discriminate touch from pain. Curr Biol. 2011;21:1552–8. doi: 10.1016/j.cub.2011.08.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Feldman DE, Knudsen EI. Experience-department plasticity and the maturation of glutamatergic synapses. Neuron. 1998;20:1067–71. doi: 10.1016/s0896-6273(00)80488-2. [DOI] [PubMed] [Google Scholar]
- 45.Fitzgerald M. The development of descending brainstem control of spinal cord sensory processing. In: Hanson M, editor. Foetal and Neonatal Brainstem: Development and Clinical Issues. Cambridge, England: Cambridge University Press; 1991. pp. 127–36. [Google Scholar]
- 46.Fitzgerald M. The development of nociceptive circuits. Nat Rev Neurosci. 2005;6:507–20. doi: 10.1038/nrn1701. [DOI] [PubMed] [Google Scholar]
- 47.Fitzgerald M. Development of pain mechanisms. Br Med Bull. 1991;47:667–75. doi: 10.1093/oxfordjournals.bmb.a072499. [DOI] [PubMed] [Google Scholar]
- 48.Fitzgerald M. Development of pain pathways and mechanisms. In: Anand KJ, McGrath PI, editors. Pain in Neonates. New York, NY: Elsevier; 1993. pp. 19–37. [Google Scholar]
- 49.Fitzgerald M. Development of the peripheral and spinal pain system. In: Anand KJ, Stevens BJ, McGrath PI, editors. Pain in Neonates. 2nd ed. Amsterdam, The Netherlands: Elsevier; 2000. pp. 9–22. [Google Scholar]
- 50.Fitzgerald M. Developmental neurobiology of pain. In: Wall PD, Melzack R, editors. Textbook of Pain. Edinburgh, Scotland: Churchill Livingstone; 1999. pp. 235–52. [Google Scholar]
- 51.Fitzgerald M. Pain and analgesia in neonates. Trends Neurosci. 1987;10:344–6. [Google Scholar]
- 52.Fitzgerald M. The sprouting of saphenous nerve terminals in the spinal cord following early postnatal sciatic nerve section in the rat. J Comp Neurol. 1985;240:407–13. doi: 10.1002/cne.902400408. [DOI] [PubMed] [Google Scholar]
- 53.Fitzgerald M, Jennings E. The postnatal development of spinal sensory processing. Proc Natl Acad Sci U S A. 1999;96:7719–22. doi: 10.1073/pnas.96.14.7719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Fitzgerald M, Koltzenburg M. The functional development of descending inhibitory pathways in the dorsolateral funiculus of the newborn rat spinal cord. Brain Res Dev Brain Res. 1986;24:261–70. doi: 10.1016/0165-3806(86)90194-x. [DOI] [PubMed] [Google Scholar]
- 55.Fitzgerald M, Millard C, MacIntosh N. Hyperalgesia in premature infants. Lancet. 1988;6:292. doi: 10.1016/s0140-6736(88)90365-0. [DOI] [PubMed] [Google Scholar]
- 56.Fitzgerald M, Millard C, McIntosh N. Cutaneous hypersensitivity following peripheral tissue damage in newborn infants and its reversal with topical anaesthesia. Pain. 1989;39:31–6. doi: 10.1016/0304-3959(89)90172-3. [DOI] [PubMed] [Google Scholar]
- 57.Fitzgerald M, Shaw A, McIntosh N. Postnatal development of the cutaneous flexor reflex: Comparative study of preterm infants and newborn rat pups. Dev Med Child Neurol. 1988;30:520–6. doi: 10.1111/j.1469-8749.1988.tb04779.x. [DOI] [PubMed] [Google Scholar]
- 58.Fitzgerald M, Shortland P. The effect of neonatal peripheral nerve section on the somatodendritic growth of sensory projection cells in the rat spinal cord. Brain Res. 1988;470:129–36. doi: 10.1016/0165-3806(88)90208-8. [DOI] [PubMed] [Google Scholar]
- 59.Fitzgerald M, Walker SM. Infant pain management: A developmental neurobiological approach. Nat Clin Pract Neurol. 2009;5:35–50. doi: 10.1038/ncpneuro0984. [DOI] [PubMed] [Google Scholar]
- 60.Flechsig PE. Leipzig: Veit; 1897. Gerhirn und Seele. [Google Scholar]
- 61.Flechsig PE. Ueber die entwickelungsgeschichtliche (myelogenetische) Flachengliederung der grosshirnrinde des menschen. Arch Ital Biol. 1901;36:30–9. [Google Scholar]
- 62.Flint AC, Maisch US, Weishaupt JH, Kriegstein AR, Monyer H. NR2A subunit expression shortens NMDFA receptor synaptic currents in developing neocortex. J Neurosci. 1997;17:2469–76. doi: 10.1523/JNEUROSCI.17-07-02469.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Gibbins S, Stevens B. The influence of gestational age on the efficacy and short-term safety of sucrose for procedural pain relief. Adv Neonatal Care. 2003;3:241–9. [PubMed] [Google Scholar]
- 64.Gibbins S, Stevens B, McGrath PJ, Yamada J, Beyene J, Breau L, et al. Comparison of pain responses in infants of different gestational ages. Neonatology. 2008;93:10–8. doi: 10.1159/000105520. [DOI] [PubMed] [Google Scholar]
- 65.Gilles F, Shankle W, Dooling E. Myelinated tracts: Growth patterns. In: Gilles F, Leviton A, Dooling E, editors. The Developing Human Brain: Growth and Epidemiologic Neuropathology. Boston: John Wright; 1983. pp. 117–83. [Google Scholar]
- 66.Gliess J, Stuttgen G. Morphologic and functional development of the skin. In: Stave U, editor. Physiology of the Perinatal Period: Functional and Biochemical Development in Mammals. New York: Appleton-Century-Crofts; 1970. pp. 889–906. [Google Scholar]
- 67.Green PG, Chen X, Alvarez P, Ferrari LF, Levine JD. Early-life stress produces muscle hyperalgesia and nociceptor sensitization in the adult rat. Pain. 2011;152:2549–56. doi: 10.1016/j.pain.2011.07.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Grunau RE. Neonatal pain in very preterm infants: Long-term effects on brain, neurodevelopment and pain reactivity. Rambam Maimonides Med J. 2013;4:e0025. doi: 10.5041/RMMJ.10132. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Grunau RE, Whitfield MF, Petrie-Thomas J, Synnes AR, Cepeda IL, Keidar A, et al. Neonatal pain, parenting stress and interaction, in relation to cognitive and motor development at 8 and 18 months in preterm infants. Pain. 2009;143:138–46. doi: 10.1016/j.pain.2009.02.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Grunau RV, Holsti L, Haley DW, Oberlander T, Weinberg J, Solimano A, et al. Neonatal procedural pain exposure predicts lower cortisol and behavioral reactivity in preterm infants in the NICU. Pain. 2005;113:293–300. doi: 10.1016/j.pain.2004.10.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Grunau RV, Holsti L, Whitfield MF, Ling E. Are twitches, startles, and body movements pain indicators in extremely low birth weight infants? Clin J Pain. 2000;16:37–45. doi: 10.1097/00002508-200003000-00007. [DOI] [PubMed] [Google Scholar]
- 72.Grunau RV, Johnston CC, Craig KD. Neonatal facial and cry responses to invasive and non-invasive procedures. Pain. 1990;42:295–305. doi: 10.1016/0304-3959(90)91142-6. [DOI] [PubMed] [Google Scholar]
- 73.Grunau RV, Whitfield MF, Petrie JH, editors. Proceedings of the Neonatal Society Meeting. Clare, Ireland: Shannon, Co; 1994. Extremely low birth weight (ELBW) toddlers are relatively unresponsive to pain at 18 months corrected age compared to larger birth weight children. [Google Scholar]
- 74.Grunau RV, Whitfield MF, Petrie JH. Pain sensitivity and temperament in extremely low-birth-weight premature toddlers and preterm and full-term controls. Pain. 1994;58:341–6. doi: 10.1016/0304-3959(94)90128-7. [DOI] [PubMed] [Google Scholar]
- 75.Grunau RV, Whitfield MF, Petrie JH. Pain sensitivity in toddlers of birthweight<1000 grams compared with heavier preterm and full birth weight toddlers. Pediatr Res. 1991;29:256A. [Google Scholar]
- 76.Grunau RV, Whitfield MF, Petrie JH, Fryer EL. Early pain experience, child and family factors, as precursors of somatization: A prospective study of extremely premature and fullterm children. Pain. 1994;56:353–9. doi: 10.1016/0304-3959(94)90174-0. [DOI] [PubMed] [Google Scholar]
- 77.Hammer P, Banck MS, Amberg R, Wang C, Petznick G, Luo S, et al. mRNA-seq with agnostic splice site discovery for nervous system transcriptomics tested in chronic pain. Genome Res. 2010;20:847–60. doi: 10.1101/gr.101204.109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Hatfield LA, Ely EA. Measurement of acute pain in infants: A review of behavioral and physiological variables. Biol Res Nurs. 2014 doi: 10.1177/1099800414531448. 1099800414531448, first published on May 1, 2014 as doi: 10.1177/1099800414531448. [DOI] [PubMed] [Google Scholar]
- 79.Hatfield LA, Meyers MA, Messing TM. A systematic review of the effects of repeated painful procedures in infants: Is there a potential to mitigate future pain responsivity? J Nurs Educ Pract. 2013;3:99–112. [Google Scholar]
- 80.Hermann C, Hohmeister J, Demirakça S, Zohsel K, Flor H. Long-term alteration of pain sensitivity in school-aged children with early pain experiences. Pain. 2006;125:278–85. doi: 10.1016/j.pain.2006.08.026. [DOI] [PubMed] [Google Scholar]
- 81.Holsti L, Grunau RE, Oberlander TF, Whitfield MF. Prior pain induces heightened motor responses during clustered care in preterm infants in the NICU. Early Hum Dev. 2005;81:293–302. doi: 10.1016/j.earlhumdev.2004.08.002. [DOI] [PubMed] [Google Scholar]
- 82.Holsti LP, Grunau RE, Whifield MF, Oberlander TF, Lindh VP. Behavioral responses to pain are heightened after clustered care in preterm infants born between 30 and 32 weeks gestational age. Clin J Pain. 2006;22:757–64. doi: 10.1097/01.ajp.0000210921.10912.47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Horii Y, Kandda K. Developmental alterations in NMDA receptor-mediated [Ca+]i elevation in substantia gelatinous neurons of neonatal rat spinal cord. Brain Res Dev Brain Res. 1994;80:141–8. doi: 10.1016/0165-3806(94)90098-1. [DOI] [PubMed] [Google Scholar]
- 84.Humphrey T. Some correlations between the appearance of human fetal reflexes and the development of the nervous system. Prog Brain Res. 1964;4:93–135. [Google Scholar]
- 85.Janig W. The sympathetic nervous system in pain. Eur J Anaesthesiol. 1995;12(Suppl 10):53–60. [PubMed] [Google Scholar]
- 86.Johnston CC, Fernandes AM, Campbell-Yeo M. Pain in neonates is different. Pain. 2011;152(3 Suppl):S65–73. doi: 10.1016/j.pain.2010.10.008. [DOI] [PubMed] [Google Scholar]
- 87.Johnston CC, Stevens BJ. Experience in a neonatal care unit affects pain response. Pediatrics. 1996;98:925–30. [PubMed] [Google Scholar]
- 88.Johnston CC, Stevens BJ, Horton L. Changes in physiological responses to heel stick in premature infants. Neonatal Netw. 1992;11:67. [Google Scholar]
- 89.Johnston CC, Stevens BJ, Yang F, Horton R. Developmental changes in response to heel stick in preterm infants: A prospective cohort study. Dev Med Child Neurol. 1996;38:435–45. doi: 10.1111/j.1469-8749.1996.tb15101.x. [DOI] [PubMed] [Google Scholar]
- 90.Kehlet H, Jensen TS, Woolf CJ. Persistent postsurgical pain: Risk factors and prevention. Lancet. 2006;367:1618–25. doi: 10.1016/S0140-6736(06)68700-X. [DOI] [PubMed] [Google Scholar]
- 91.Kim H, Dionne RA. Individualized pain medicine. Drug Discov Today Ther Strateg. 2009;6:83–7. doi: 10.1016/j.ddstr.2010.10.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Kim J, Foy M, Thompson R. Behavioral Stress modifies hippocampal plasticity through NMDA receptor activation. Proc Natl Acad Sci U S A. 1996;93:4750–3. doi: 10.1073/pnas.93.10.4750. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Koga K, Furue H, Rashid H, Takaki A, Katafuchi T, Yoshimura M. Selective activation of primary afferent fibers evaluated by sine-wave electrical stimulation. Mol Pain. 2005;1:13. doi: 10.1186/1744-8069-1-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Kwok CH, Devonshire IM, Bennett AJ, Hathway GJ. Postnatal maturation of endogenous opioid systems within the periaqueductal grey and spinal dorsal horn of the rat. Pain. 2014;155:168–78. doi: 10.1016/j.pain.2013.09.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Lee YS, Kim H, Wu TX, Wang XM, Dionne RA. Genetically mediated interindividual variation in analgesic responses to cyclooxygenase inhibitory drugs. Clin Pharmacol Ther. 2006;79:407–18. doi: 10.1016/j.clpt.2006.01.013. [DOI] [PubMed] [Google Scholar]
- 96.Leon AC, Davis LL, Kraemer HC. The role and interpretation of pilot studies in clinical research. J Psychiatr Res. 2011;45:626–9. doi: 10.1016/j.jpsychires.2010.10.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Low LA, Fitzgerald M. Acute pain and a motivational pathway in adult rats: Influence of early life pain experience. PLos One. 2012;7:e34316. doi: 10.1371/journal.pone.0034316. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Low LA, Schweinhardt P. Early life adversity as a risk factor for fibromyalgia in later life. Pain Res Treat. 2012;2012:140832. doi: 10.1155/2012/140832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Marin-Padilla M. Structural organization of the human cerebral cortex prior to the appearance of the cortical plate. Anat Embryol (Berl) 1983;168:21–40. doi: 10.1007/BF00305396. [DOI] [PubMed] [Google Scholar]
- 100.Marti E, Gibson SJ, Polak JM, Facer P, Springall DR, Van Aswegen G, et al. Ontogeny of peptid-and amine-containing neurones in motor, sensory, and autonomic regions of the rat and human spinal cord, dorsal root ganglia, and rat skin. J Comp Neurol. 1987;266:332–59. doi: 10.1002/cne.902660304. [DOI] [PubMed] [Google Scholar]
- 101.McGrath P. New York, NY: Guilford Press; 1993. Pain in Children. [Google Scholar]
- 102.Melzack R. Gate control theory: On the evolution of pain concepts. Pain Forum. 1996;5:128–38. [Google Scholar]
- 103.Melzack R. Recent concepts of pain. J Med. 1982;13:147–60. [PubMed] [Google Scholar]
- 104.Melzack R, Wall PD. Pain mechanisms: A new theory. Science. 1965;699:971–81. doi: 10.1126/science.150.3699.971. [DOI] [PubMed] [Google Scholar]
- 105.Melzack R, Wall PD. Pain mechanisms: A new theory: A gate control system modulates sensory input from the skin before it evokes pain perception and response Pain Forum. 1996;5:3–11. [Google Scholar]
- 106.Melzack R, Wall PD. Psychophysiology of pain. Int Anesthesiol Clin. 1970;8:3–34. doi: 10.1097/00004311-197000810-00003. [DOI] [PubMed] [Google Scholar]
- 107.Miranda A. Early life stress and pain: An important link to functional bowel disorders. Pediatr Ann. 2009;38:279–82. [PubMed] [Google Scholar]
- 108.Peters JW, Schouw R, Anand KJ, van Dijk M, Duivenvoorden HJ, Tibboel D. Does neonatal surgery lead to increased pain sensitivity in later childhood? Pain. 2005;114:444–54. doi: 10.1016/j.pain.2005.01.014. [DOI] [PubMed] [Google Scholar]
- 109.Porter FL, Grunau RE, Anand KJ. Long-term effects of pain in infants. J Dev Behav Pediatr. 1999;20:253–61. doi: 10.1097/00004703-199908000-00008. [DOI] [PubMed] [Google Scholar]
- 110.Prechtl HF. The behavioural states of the newborn infant (a review) Brain Res. 1974;76:185–212. doi: 10.1016/0006-8993(74)90454-5. [DOI] [PubMed] [Google Scholar]
- 111.Rabinowicz T, de Courten-Meyers GM, Petetot JM. Human cortex development: Estimates of neuronal numbers indicate major loss during late during gestation. J Neuropathol Exp Neurol. 1996;155:320–8. [PubMed] [Google Scholar]
- 112.Raja SN, Meyer RA, Ringkamp M. Peripheral neural mechanisms of nociception. In: Wall P D, Melzack R, editors. Textbook of Pain. Edinburgh, Scotland: Churchill Livingstone; 1999. pp. 11–58. [Google Scholar]
- 113.Rakic P, Goldman-Rakic PS. Development and modifiability of the cerebral cortex: Early developmental effects: Cell lineages, acquisition of neuronal positions, and areal and larninar development. Neurosci Res Program Bull. 1982;20:433–51. [PubMed] [Google Scholar]
- 114.Ranger M, Johnston CC, Anand KJ. Current controversies regarding pain assessment in neonates. Semin Perinatol. 2007;31:283–8. doi: 10.1053/j.semperi.2007.07.003. [DOI] [PubMed] [Google Scholar]
- 115.Rawlings DJ, Miller PA, Engle RR. The effect of circumcision on transcutaneous P02 in term infants. Am J Dis Child. 1980;134:676–8. doi: 10.1001/archpedi.1980.02130190044011. [DOI] [PubMed] [Google Scholar]
- 116.Reddi D, Curran N. Chronic pain after surgery: Pathophysiology, risk factors and prevention. Postgrad Med J. 2014;90:222–7. doi: 10.1136/postgradmedj-2013-132215. [DOI] [PubMed] [Google Scholar]
- 117.Reichling DB, Kyrozis A, Wang J, Mcdermott A. Mechanisms of GABA and glycine depolarization induced calcium transients in rat dorsal horn neurons. J Physiol. 1994;476:411–21. doi: 10.1113/jphysiol.1994.sp020142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Ren K, Dubner R. Interactions between the immune and nervous systems in pain. Nat Med. 2010;16:1267–76. doi: 10.1038/nm.2234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Reynolds ML, Fitzgerald M. Long-term sensory hyperinnervation following neonatal skin wounds. J Comp Neurol. 1995;358:487–98. doi: 10.1002/cne.903580403. [DOI] [PubMed] [Google Scholar]
- 120.Rodrigues AC, Guinsburg R. Pain evaluation after a non-nociceptive stimulus in preterm infants during the first 28 days of life. Early Hum Dev. 2013;89:75–9. doi: 10.1016/j.earlhumdev.2012.08.002. [DOI] [PubMed] [Google Scholar]
- 121.Sandkühler J. Models and Mechanisms of Hyperalgesia and Allodynia. Physiol Rev. 2009;89:707–58. doi: 10.1152/physrev.00025.2008. [DOI] [PubMed] [Google Scholar]
- 122.Schechter NL. The undertreatment of pain in children: An overview. Pediatr Clin North Am. 1989;36:781–94. doi: 10.1016/s0031-3955(16)36721-9. [DOI] [PubMed] [Google Scholar]
- 123.Scholz J, Woolf CJ. Can we conquer pain? Nat Neurosci. 2002;5(Suppl):1062–7. doi: 10.1038/nn942. [DOI] [PubMed] [Google Scholar]
- 124.Schug SA, Chong C. Pain management after ambulatory surgery. Curr Opin Anaesthesiol. 2009;22:738–43. doi: 10.1097/ACO.0b013e32833020f4. [DOI] [PubMed] [Google Scholar]
- 125.Schulte F. Gestation, wachsturn und hirnentwicklung. In: Linneweh F, editor. Fortscritte der Paedologie. Berlin: Springer-Verlag; 1968. pp. 46–64. [Google Scholar]
- 126.Schwaller F, Fitzgerald M. The consequences of pain in early life: Injury-induced plasticity in developing pain pathways. Eur J Neurosci. 2014;39:344–52. doi: 10.1111/ejn.12414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Seo S, Grzenda A, Lomberk G, Ou XM, Cruciani RA, Urrutia R. Epigenetics: A promising paradigm for better understanding and managing pain. J Pain. 2013;14:549–57. doi: 10.1016/j.jpain.2013.01.772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Shipton EA. The transition from acute to chronic post surgical pain. Anaesth Intensive Care. 2011;39:824–36. doi: 10.1177/0310057X1103900506. [DOI] [PubMed] [Google Scholar]
- 129.Simons SH, van Dijk M, Anand KJ, Roofthooft D, van Lingen RA, Tibboel D. Do we still hurt newborn babies? A prospective study of procedural pain and analgesia in neonates. Arch Pediatr Adolesc Med. 2003;157:1058–64. doi: 10.1001/archpedi.157.11.1058. [DOI] [PubMed] [Google Scholar]
- 130.Sivilotti LG, Gerber G, Rawat B, Woolf CJ. Morphine selectively depresses the slowest, NMDA-independent component of C-fiber-evoked synaptic activity in the rat spinal cord in vitro. Eur J Neurosci. 1995;7:12–8. doi: 10.1111/j.1460-9568.1995.tb01015.x. [DOI] [PubMed] [Google Scholar]
- 131.Spehlmann R. New York: Elsevier/North-Holland; 1981. EEG primer. [Google Scholar]
- 132.Spehlmann R. New York, NY: Elsevier/North Holland; 1981. EEG Primer. [Google Scholar]
- 133.Sredl D. Myths and facts about pain in neonates. Neonatal Netw. 2003;22:69–71. doi: 10.1891/0730-0832.22.6.69. [DOI] [PubMed] [Google Scholar]
- 134.Stein C, Machelska H. Modulation of peripheral sensory neurons by the immune system: Implications for pain therapy. Pharmacol Rev. 2011;63:860–81. doi: 10.1124/pr.110.003145. [DOI] [PubMed] [Google Scholar]
- 135.Stevens BJ, Johnston CC. Premature infant's response to pain: A pilot study. Nurs Que. 1991;11:90–5. [PubMed] [Google Scholar]
- 136.Stone LS, Szyf M. The emerging field of pain epigenetics. Pain. 2013;154:1–2. doi: 10.1016/j.pain.2012.10.016. [DOI] [PubMed] [Google Scholar]
- 137.Teng CJ, Abbott FV. The formalin test: A dose-response analysis at three developmental stages. Pain. 1998;76:337–47. doi: 10.1016/S0304-3959(98)00065-7. [DOI] [PubMed] [Google Scholar]
- 138.Terman GW, Bonica JJ. Spinal mechanisms and their modulation. In: Loeser J, Butler S, Chapman C, Turk D, editors. Bonica's Management of pain. Philadelphia, PA: Lippincott Williams and Wilkins; 2001. pp. 73–152. [Google Scholar]
- 139.Tilney F, Rosett J. The value of brain lipoids as an index of brain development. Bull Neurol Inst. NY. 1931;1:28–71. [Google Scholar]
- 140.Van Cleve L, Johnson L, Andrews S, Hawkins S, Newbold J. Pain responses of hospitalized neonates to venipuncture. Neonatal Network. 1995;14:31–5. [PubMed] [Google Scholar]
- 141.Van Pragg H, Frenk H. The development of stimulation-induced analgesia (SPA) in the rat. Brain Res Dev Brain Res. 1991;64:71–6. doi: 10.1016/0165-3806(91)90210-a. [DOI] [PubMed] [Google Scholar]
- 142.Vinall J, Grunau RE. Impact of repeated procedural pain-related stress in infants born very preterm. Pediatr Res. 2014;75:584–7. doi: 10.1038/pr.2014.16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Volpe JJ. Intracranial hemorrhage: Subdural, primary subarchnoid, intracerebellar, intraventricular (term infant) and miscellaneous. In: Fletcher J, editor. Neurology of the Newborn. 3 ed. Philadelphia, PA: Saunders; 2000. pp. 397–428. [Google Scholar]
- 144.Voscopoulos C, Lema M. When does acute pain become chronic? Br J Anaesth. 2010;105(Suppl 1):i69–85. doi: 10.1093/bja/aeq323. [DOI] [PubMed] [Google Scholar]
- 145.Walden M, Penticuff JH, Stevens B, Lotas M, Kozinetz CA, Clark A, et al. Maturational changes in physiologic and behavioral responses of preterm neonates in pain. Adv Neonatal Care. 2001;1:94–106. [Google Scholar]
- 146.Walker SM, Tochiki KK, Fitzgerald M. Hindpaw incision in early life increases the hyperalgesic response to repeat surgical injury: Critical period and dependence on initial afferent activity. Pain. 2009;147:99–106. doi: 10.1016/j.pain.2009.08.017. [DOI] [PubMed] [Google Scholar]
- 147.Wang J, Reichling DB, Kyrozis A, Mcdermott AB. Developmental loss of GABA and glycine induced depolarization and Ca2+transients in embryonic rat dorsal horn neurons in culture. Eur J Neurosci. 1994;6:1275–80. doi: 10.1111/j.1460-9568.1994.tb00317.x. [DOI] [PubMed] [Google Scholar]
- 148.Williamson PS, Williamson ML. Physiological stress reduction by local anaesthetic during newborn circumcision. Pediatrics. 1983;71:36–40. [PubMed] [Google Scholar]