

Abstract
Legionella pneumophila has been recognized as the major cause of legionellosis since the discovery of the deadly disease. Legionella spp. other than L. pneumophila were later found to be responsible to many non-pneumophila infections. The non-L. pneumophila infections are likely under-detected because of a lack of effective diagnosis. In this report, we have sequenced the 16S-23S rRNA gene internal transcribed spacer (ITS) of 10 Legionella species and subspecies, including L. anisa, L. bozemanii, L. dumoffii, L. fairfieldensis, L. gormanii, L. jordanis, L. maceachernii, L. micdadei, L. pneumophila subspp. fraseri and L. pneumophila subspp. pasculleii, and developed a rapid oligonucleotide microarray detection technique accordingly to identify 12 most common Legionella spp., which consist of 11 pathogenic species of L. anisa, L. bozemanii, L. dumoffii, L. gormanii, L. jordanis, L. longbeachae, L. maceachernii, L. micdadei, and L. pneumophila (including subspp. pneumophila, subspp. fraseri, and subspp. pasculleii) and one non-pathogenic species, L. fairfieldensis. Twenty-nine probes that reproducibly detected multiple Legionella species with high specificity were included in the array. A total of 52 strains, including 30 target pathogens and 22 non-target bacteria, were used to verify the oligonucleotide microarray assay. The sensitivity of the detection was at 1.0 ng with genomic DNA or 13 CFU/100 mL with Legionella cultures. The microarray detected seven samples of air conditioner-condensed water with 100% accuracy, validating the technique as a promising method for applications in basic microbiology, clinical diagnosis, food safety, and epidemiological surveillance. The phylogenetic study based on the ITS has also revealed that the non-pathogenic L. fairfieldensis is the closest to L. pneumophila than the nine other pathogenic Legionella spp.
Introduction
Legionella acquired its name after an outbreak of a then-unknown “mystery disease” that affected 221 persons, and caused 34 deaths eventually, attending a convention of the American Legion in July 1976. This epidemic, which occurred within days of the 200th anniversary of the signing of the Declaration of Independence, was widely publicized and raised great concern in the United States [1]. A few months later, the causative agent was identified as a previously unknown bacterium, which was subsequently named Legionella. This gram-negative bacterium includes species responsible for Legionellosis or Legionnaire’s diseases, with Legionella pneumophila as the most notably species [2], [3]. Since then, more than 52 Legionella spp. have been identified (http://www.bacterio.cict.fr/l/legionella.html) [4], [5]. Although L. pneumophila remains as the major cause of legionellosis, non-pneumophila infections have been reported to be caused by Legionella micdadei (60%), Legionella bozemanii (15%), Legionella dumoffii (10%), Legionella longbeachae (5%), and other species (10%) [6].
Infections due to species other than L. pneumophila are likely to be underestimated because of a lack of appropriate diagnostic tests [6]. Since Legionella was first identified in 1977, various diagnostic tools for Legionella have been developed, including cell culture, antigen detection, serological typing, polymerase chain reaction (PCR), and microarray methods. The culture method is time-consuming due to the slow growth of Legionella spp., and it fails to distinguish Legionella spp. at the species level [7]. The detection of Legionella antigen in urine by enzyme immunoassays is a highly specific approach; and commercially available systems using this approach can detect L. pneumophila serogroup O1 but not other serogroups [8]. Serological typing methods with monoclonal and multiclonal antibodies can be used to detect L. pneumophila only with the aid of laborious pre-culture [9], [10].
Currently, most PCR methods target 5S rRNA, 16S rRNA, 23S-5S ribosomal RNA intergenic spacer, mip, rpoB, and gyrB genes [11]–[17]. However, the 5S, 16S, and 23S-5S rRNA genes are too conserved to differentially detect L. pneumophila from other Legionella spp. [18]. While the mip gene was initially used as an L. pneumophila-specific marker [19], other Legionella spp. were later found to harbor this gene as well [20], [21] A previous study has conducted a multilocus sequence analysis of 16S rRNA, mip, and ropB [22], and found 16S rRNA was useful for initial identification as it could recognize isolates robustly at the genus level, while mip, rpoB, and the mip-rpoB concatenation can be used to distinguish between different Legionella spp. However, multiplex PCR and sequencing are required for the identification, which render this method cumbersome and time-consuming. A gyrB gene-based single PCR method was developed for the differentiation of L. pneumophila subspp. pneumophila and L. pneumophila subspp. fraseri, but not for other Legionella spp. [16]. An oligonucleotide array based on mip gene sequences and digoxigenin-labeled PCR products was developed to identify 18 species of Legionella that have been reported to cause human infections, but the results are not reliable as some of the species only produced weak hybridization signals [13]. One other oligonucleotide microarray based on the wzm and wzt gene sequences and Cy3-labeled PCR products was developed to serotype all 15 distinct O-antigen forms within L. pneumophila [23].
In this study, we report the establishment of an oligonucleotide microarray method for the simultaneous detection of 11 pathogenic Legionella spp., L. anisa, L. bozemanii, L. dumoffii, L. gormanii, L. jordanis, L. longbeachae, L. maceachernii, L. micdadei, and L. pneumophila (including subspp. pneumophila, subspp. fraseri, and subspp. pasculleii), and one non-pathogenic spp., L. fairfieldensis, based on the 16-23S rRNA gene internal transcribed spacer (ITS) regions. The microarray method described here is specific, sensitive, and reliable and can be used as a better alternative to the traditional serotyping procedure, which is laborious and frequently cross-reactive.
Materials and Methods
Bacterial strains
The following standard Legionella spp. strains were used for ITS sequencing: L. anisa (DSMZ 17627), L. bozemanii (ATCC 33217), L. dumoffii (ATCC 33279), L. fairfieldensis (ATCC 49588), L. gormanii (ATCC 43769), L. jordanis (DSMZ 19212), L. maceachernii (DSMZ 16642), L. micdadei (NCTC 11371), L. pneumophila subspp. fraseri (ATCC 35251), and L. pneumophila subspp. pascullei (ATCC 4585). The 52 bacterial strains used for microarray are listed and described in Table 1, and included 30 strains of the Legionella target species and 22 other non-target bacterial species. Of these 52 strains, 41 were reference strains and 11 were clinical or environmental isolates. Legionella strains were cultured onto buffered charcoal yeast extract (BCYE) agar plates (Hope Bio-technology Co., Ltd, Qingdao, China) and incubated in a 5% CO2 incubator at 37°C for 2–4 days.
Table 1. Bacterial strains used in this study.
| Bacterium | No. of strains of each source | Total number |
| Target Legionella spp. used to test the specificity of the probes (n = 30) | ||
| Legionella pneumophila (subspp. pneumophila) | 1a, 10b, 2c, 1 k | 14 |
| Legionella pneumophila (subspp. fraseri) | 1c, 1d, 1k | 3 |
| Legionella pneumophila (subspp. pascullei) | 1c | 1 |
| Legionella anisa | 1d | 1 |
| Legionella bozemanii | 1c, 1j | 2 |
| Legionella dumoffii | 1c | 1 |
| Legionella fairfieldensis | 1c | 1 |
| Legionella gormanii | 1c | 1 |
| Legionella jordanis | 1d | 1 |
| Legionella longbeachae | 1d, 1j | 2 |
| Legionella maceachernii | 1d | 1 |
| Legionella micdadei | 1b, 1j | 2 |
| Other bacterial species used to test the specificity of the probes (n = 22) | ||
| Legionella feeleii | 1j | 1 |
| Legionella steigerwaltii | 1b | 1 |
| Legionella worsleiensis | 1c | 1 |
| Acinetobacter baumannii | 1g | 1 |
| Citrobacter freundii | 1f | 1 |
| Escherichia coli | 1f | 1 |
| Enterococcus faecalis | 1e | 1 |
| Klebsiella pneumoniae | 1f | 1 |
| Listeria monocytogenes | 1h | 1 |
| Proteus mirabilis | 1i | 1 |
| Proteus penneri | 1a | 1 |
| Proteus vulgaris | 1i | 1 |
| Pseudomonas aeruginosa | 1e | 1 |
| Salmonella paratyphi | 1f | 1 |
| Salmonella typhi | 1f | 1 |
| Shigella boydii | 1k | 1 |
| Shigella flexneri | 1c | 1 |
| Streptococcus pneumoniae | 1c | 1 |
| Streptococcus pyogenes | 1f | 1 |
| Staphylococcus aureus | 1e | 1 |
| Staphylococcus epidermidis | 1f | 1 |
| Vibrio parahaemolyticus | 1j | 1 |
| Bacterial species used to perform the blind test (n = 22) | ||
| Legionella pneumophila (subspp. pneumophila) | 1a | 1 |
| Legionella pneumophila (subspp. fraseri) | 1d | 1 |
| Legionella pneumophila (subspp. pascullei) | 1c | 1 |
| Legionella anisa | 1d | 1 |
| Legionella bozemanii | 1c | 1 |
| Legionella dumoffii | 1c | 1 |
| Legionella fairfieldensis | 1c | 1 |
| Legionella gormanii | 1c | 1 |
| Legionella jordanis | 1d | 1 |
| Legionella longbeachae | 1d | 1 |
| Legionella maceachernii | 1d | 1 |
| Legionella micdadei | 1b | 1 |
| Legionella steigerwaltii | 1b | 1 |
| Legionella worsleiensis | 1c | 1 |
| Escherichia coli | 1f | 1 |
| Enterococcus faecalis | 1e | 1 |
| Klebsiella pneumoniae | 1f | 1 |
| Pseudomonas aeruginosa | 1e | 1 |
| Salmonella typhi | 1f | 1 |
| Staphylococcus aureus | 1e | 1 |
| Streptococcus pneumoniae | 1c | 1 |
Czech Collection of Microorganisms (CCM), Masaryk University, Brno, Czech Republic.
National Collection of Type Cultures (NCTC), Central Public Health Laboratory, London, United Kingdom.
American Type Culture Collection (ATCC), USA.
German Collection of Microorganisms and Cell Cultures (DSMZ), Germany.
Institute of Microbiology, Chinese Academy of Sciences (IMCAS).
National Center for Medical Culture Collections (CMCC), Beijing, China.
Universityät zu Köln, Deutschland, Gernamy.
Agricultural Culture Collection of China (ACCC), Beijing, China.
University of Lodz, Poland.
Shanghai Municipal Center for Disease Control and Prevention.
Shenzhen NanShan Center for Disease Control and Prevention.
Genomic DNA preparation
Genomic DNA was extracted from pure cultures using bacterial genomic DNA purification kit (Tiangen Biotech Co., Ltd., Beijing, China).
Amplification of Legionella spp. ITS regions
The primer pair wl-5793 and wl-5794 was designed on the basis of the 16S rRNA gene and 23S rRNA gene sequences, respectively, using Primer Premier 5.0 software (Premier Boost International, CA) [24], [25]. These primers were used to amplify the ITS region of all Legionella spp. The primer sequences and concentrations used for multiplex PCR are listed in Table 2. The PCR mixture contained 1× PCR buffer (50 mM KCl, 10 mM Tris-HCl; pH 8.3), 2.5 mM MgCl2, 200 µM dNTP, 1.0 U Taq DNA polymerase, 10 nM of each of the primers, and 100 ng of DNA template in a final volume of 50 µl. PCR was performed by initial denaturation at 95°C for 5 min; followed by 30 cycles of 94°C for 30 s, 50°C for 30 s, and 72°C for 1 min; and a final extension at 72°C for 5 min. The amplified fragment was then checked by agarose gel electrophoresis of 2-µL aliquots of the PCR products.
Table 2. Primers and their concentrations in multiplex PCR.
| Primer name | Target gene | Sequence (5′–3′)a | PCR product size (bp) | GenBank accession no. | Primer Conc. in multiplex PCR (µM) | Primer Conc. for Labelling (µM) |
| wl-5793 | ITS | (F)TGTACACACCGCCCGTC | 500–1000 | CP000675.2 | 0.2 | |
| wl-5794 | (R)GGTACTTAGATGTTTCAGTTC | 0.2 | 0.2 |
F, forward primer; R, reverse primer.
Cloning and sequencing of the ITS regions of Legionella spp. and subspp
PCR amplicons were cloned into the pGEM-T Easy vector (Promega, MA) and transformed into E. coli DH5α. Transformants (observed as white colonies grown on an ampicillin plates containing isopropyl-beta-d-thiogalactopyranoside and X-gal) were selected randomly. Plasmid DNA was isolated using the conventional alkaline lysis method, digested with Eco RI, and visualized on agarose gels to confirm the presence of the corresponding inserts. Sequences were verified using an ABI 3730 automated DNA sequencer (Applied Biosystems, USA). Sixteen transformants per Legionella spp. or subspp. were examined.
Sequence analysis
Multiple sequence alignment of ITS sequences was carried out with the ClustalW program (http://www.ebi.ac.uk/clustalw/). The identity level was calculated using BioEdit software (http://www.mbio.ncsu.edu/BioEdit/page2.html). Phylogenetic trees were constructed using the neighbor-joining method and plotted by the molecular evolutionary genetics analysis (MEGA) 3.1 software package (http://www.megasoftware.net). Bootstrap analysis was carried out based on 1,000 replicates.
Target DNA amplification and labeling
Primer concentrations were optimized according to the final intensity of the microarray hybridization signals. The PCR mixtures contained 1×PCR buffer (50 mM KCl, 10 mM Tris-HCl; pH 8.3), 2.5 mM MgCl2, 400 µM dNTP, 0.2 µM ITS of each primer, 2.5 U Taq DNA polymerase, and 50–100 ng of DNA template in a final volume of 25 µL. The following PCR parameters were employed: initial denaturation at 95°C for 5 min; followed by 35 cycles of 95°C for 30 s, 50°C for 30 s, and 72°C for 1 min; and a final extension at 72°C for 5 min. The amplified DNA was analyzed by agarose gel electrophoresis of a 2-µL aliquot of the PCR product (Fig. S1). To label the PCR products, 10 µL of the PCR products generated from the first run and the reverse primer and 0.3 µL of 25 nM Cy3-dUTP were added to the PCR mixture, and PCR was carried out using the same PCR conditions described above.
Oligonucleotide probe design
The conserved and variable regions of the ITS sequences were defined by aligning multiple ITS sequences using ClustalW. For each type of pathogen, two to four probes were designed on MEGA 3.1 based on the sequences from the GenBank database or from our lab data, and checked by Primer Premier 5.0. One probe based on the 16S rRNA gene was designed as the positive control (OA-1993). A probe containing 40 poly(T) oligonucleotides was used as the negative control (WL-4006). A probe containing 40 poly(T) oligonucleotides labeled by 3′-Cy3 was used as the positional reference and printing control (Cy3). Each probe comprised a modified 5′ amino acid sequence followed by a spacer of 10 to 15 poly(T)s and a stretch of specific sequence (synthesized by AuGCT Biotechnology Corporation, Beijing, China). All the oligonucleotide probes used in this study are listed in Table 3.
Table 3. Oligonucleotide probes used in this study.
| Target species | Probe name | Tm (°C)a | Sequence (5′–3′) | GenBank accession no. |
| Legionella anisa | OA-3819 | 68.5 | GCATGCATCAGTATGTGACCAAGCGAGCGAG | KM609989 |
| OA-3820 | 70.1 | CGAGCGAGTGGATGCAATGAAAACAAATTT | ||
| Legionella bozemanii | OA-3821 | 68.5 | AAAGCCGTGACCGAGAGGAAGCGGGAAGA | KM609990 |
| OA-3822 | 78.6 | AAGCGGGAAGATGCGCGGTCACGCTGAAAGC | ||
| OA-3823 | 73.8 | TCGTGACCGAGAGGAAGCGGGAAGATGCGC | ||
| Legionella dumoffii | OA-3824 | 57.2 | TGAATGATGAATAAATCCTAAGCTTCTGAA | KM609992 KM609993 |
| OA-3825 | 62.4 | CTGAAAGGAAGCAAATGCTTGATAAAGC | ||
| OA-3826 | 63.8 | TCCTAACCGTAATTTTTTATGCGGAAAGAAT | ||
| Legionella fairfieldensis | OA-3827 | 66.6 | GATTGCCGTATTTTTTGGGTGGATTGGAATG | KM609994 |
| OA-3828 | 60.1 | AGAGTCTGCATTGTGTAGCATTGATTATTG | ||
| OA-3829 | 69.1 | TGGGTGGATTGGAATGGTTTCATGAA | ||
| Legionella gormanii | OA-3830 | 69.2 | TGAGCCCGGTTCATAACGTTGTGAGTGCGGC | KM609996 KM609997 |
| OA-3831 | 50.4 | AGATAATTTTTCTTTAGTTCAAGTAAGTGTT | ||
| OA-3832 | 65.2 | GTAAAATTGCACTGTCTTGCGTTGGAG | ||
| Legionella jordansis | OA-3833 | 77.3 | ACTCCGATGCGAGGGAGCGAAGCGACCAA | KM609999 |
| OA-3834 | 71.8 | TGCTGAGCGAGGGAGCTTCGTAACCAAGGGGT | ||
| OA-3835 | 62.4 | ACCTTTATTGATTTTAGCGATGGCTTTGAA | ||
| Legionella longbeachae | OA-3836 | 63.9 | AAGCGGTAACAAAAGAGTGACTCGAAGC | NC_013861 |
| OA-3837 | 60.5 | CCAATTTTAGGGTTTTCAAGGATAGTCCA | ||
| OA-3838 | 57.5 | CAGAAAGATGAAAAATCTTAAGCTGCG | ||
| Legionella micdadei | OA-3839 | 65.9 | ATTCCTTAATCGAGATGTCAACGCGAAGG | |
| OA-3840 | 69.3 | GCTCGGTATGTGACCGAGGAAAAGCATGTC | ||
| OA-3841 | 75.7 | TACCGATGGCGCTTGGAACGGGCTAATGAGCC | ||
| Legionella maceachernii | OA-3842 | 57.9 | AAGACAAGGAAAAGAATAGCAGATTCTGCG | |
| OA-3843 | 69.4 | GCATCGGTGCGAAAGAGTAAGGGAGCCTACG | ||
| OA-3844 | 68.0 | ACGCAAGTAGGTGGCTGAACGAAAGGGTAT | ||
| Legionella pneumophila (subspp pneumophila,subspp fraseri, subspp pascullei) | OA-3815 | 71.3 | CAAGAATCGGAACGCGGTCCAAGATTGG | CP000675.2 |
| OA-3816 | 65.3 | AAGCGATTGGTATTTGCATCATGTGATTT | ||
| OA-3817 | 59.2 | CATAGAAAGGCACAGAAGGAACTAGAGTGC | ||
| Positive control | OA-1993 | 68.5 | GTACACACCGCCCGTCACACCATb | X80725 |
| Negative control | WL-4006 | TTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTc | ||
| Positional reference & printing control | Cy3 | TTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTTT _Cy3d |
Tm was predicted using Primer Premier 5.0 software.
The 16S rDNA based probe was used as the positive control.
The probe containing 40 poly(T) oligonucleotides was used as the negative control.
The probe labeled by 3′-Cy3 was used as the positional reference and printing control.
Microarray preparation
The probes were dissolved in 50% dimethyl sulfoxide (DMSO) to a final concentration of 1 µg/µL and coated onto aldehyde group-modified glass slides (CapitalBio Corporation, Beijing, China) using SpotArray 72 (Perkin-Elmer Corporation, CA, USA). Each probe was spotted in triplicate. Coated slides were dried and stored at room temperature in the dark. Each glass slide contained eight individual arrays framed with an 8-sample cover slip containing individual reaction chambers. A schematic diagram of the probe positions on the microarray is shown in Fig. 1.
Figure 1. Probe positions on the microarray.
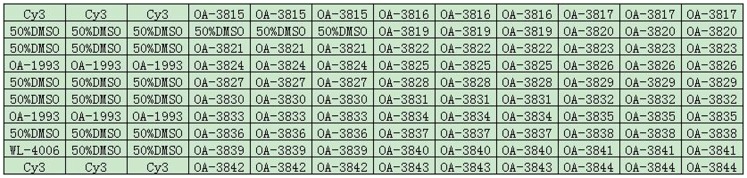
OA-1993, the positive control probe based on the 16S rRNA gene. WL-4006, the negative control probe. Cy3, the positional reference and printing control probe. Blank, 50%DMSO. The rest are the specific probes for the target strains.
Microarray hybridization and data analysis
All labeled PCR products were precipitated using 100% cold ethanol, centrifuged at 13,000 g for 10 min, washed with 75% ethanol, and dried at room temperature. The dried, labeled DNA was diluted in 16 µL of hybridization buffer (50% formamide, 6× SSC, 5× Denhardt, and 0.5% SDS) and then hybridized with the prepared microarray at 45°C for 12 h. After hybridization, the slide was washed with solution A (1× SSC and 0.1% SDS) for 3 min, solution B (0.05×SSC) for 3 min, and solution C (95% ethanol) for 1.5 min. The microarray was dried under a gentle air stream and scanned with a laser beam of 532 nm using the GenePix biochip scanner 4100A (Axon Instruments, CA, USA) set to the following parameters: photomultiplier tube gain, 600, and pixel size, 5 µm. The signal-to-noise ratio (SNR) was calculated for each spot using the built-in software, GenePix Pro 6.0, with the threshold set at 3.0. A signal was considered positive when 70% of the probes to a respective target gene generated hybridization signals above the SNR threshold.
Test of mock samples
BCYE medium was used for proliferation. Pure cultures of L. bozemanii, L. dumoffii, and L. gormanii were serially diluted from 101 to 106 CFU/mL, and 1 mL of the diluent was mixed with 100 mL of fresh tap water from the laboratory and vacuum filtered through a 0.22-µm membrane. The membrane was treated with 500 µL of diluted HCl (pH 3.0) for 1 min, placed face-down on BCYE agar plates, and incubated in a 5% CO2 incubator at 37°C for 3–5 days. The genomic DNA was then extracted from the cells for microarray hybridization.
Test of air conditioner-condensed water samples
A filter-enriched air conditioner-condensed water sample (800 µL) was plated onto a GVPC agar plate (Hope Bio-Technology Co., Ltd., Qingdao, China) and incubated in a 5% CO2 incubator at 37°C for 48 h. Then, the culture was collected and genomic DNA was extracted for use in the downstream PCR and hybridization process.
Nucleotide sequence and microarray accession numbers
The ITS sequences of L. anisa, L. bozemanii, L. dumoffii, L. fairfieldensis, L. gormanii, L. jordanis, L. maceachernii, L. micdadei, and L. pneumophila subspp. fraseri, and L. pneumophila subspp. pascullei were deposited into the GenBank database under the accession numbers KM609984-KM610004. The microarray dataset was deposited into the Gene Expression Omnibus database under the accession number GSE61962.
Results
Legionella spp. ITS regions reveal interspecies variations
We have sequenced the ITS regions of 10 Legionella spp. and subspp., including L. anisa, L. bozemanii, L. dumoffii, L. fairfieldensis, L. gormanii, L. jordanis, L. maceachernii, L. micdadei, L. pneumophila subspp. fraseri and L. pneumophila subspp. pasculleii. Next we analyzed the ITS regions of the 12 Legionella spp., the above 10 plus L. longbeachae and L. pneumophila subspp. pneumophila, whose sequences were previously published (NC_013861, CP0005672) using tRNA-ScanE software (http://lowelab.ucsc.edu/tRNAscan-SE/). The data indicated that except for L. jordanis, which has three ITS types of ITS-tRNAAla (with tRNAAla gene), ITS-tRNAIle (with tRNAIle gene), and ITS-tRNAnone (without tRNA gene), the 11 other Legionella spp. and subspp. all contain two distinct ITS types: ITS-tRNAAla (with tRNAAla gene) and ITS-tRNAIle (with tRNAIle gene). Alignments of the above ITS sequences revealed significant interspecies variations of 0.266–0.772 for ITS-tRNAAla and 0.280-0.774 for ITS-tRNAIle, but low intraspecies polymorphisms of 0.967–0.993 for L. pneumophila ITS-tRNAAla and 0.943–0.996 for L. pneumophila ITS-tRNAIle, suggesting that ITS is a good target for species-specific identification.
Phylogenetic analysis
We have constructed two phylogenetic trees of 12 Legionella spp. and subspp. based on the ITS-tRNAAla and ITS-tRNAIle gene sequences with Staphylococcus aureus and Enterococcus faecium as the outer group references for the two gene sequences (Fig. 2A and 2B). In the ITS-tRNAAla tree, there are two subgroups: the first subgroup consists of L. pneumophila subspp. pneumophila, L. pneumophila subspp. fraseri, L. pneumophila subspp. pascullei, L. fairfieldensis, L. jordanis, L. maceachernii, and L micdadei; while the second contained subgroup, L. gormanii, L. anisa, L. bozemanii, L. dumoffii, and L. longbeachae. In the ITS-tRNAIle tree, there are three subgroups: the first subgroup includes L. pneumophila subspp. pneumophila, L. pneumophila subspp. fraseri, L. pneumophila subspp. pascullei and L. fairfieldensis; the second subgroup, L. jordanis, L. maceachernii, and L micdadei; the third subgroup, L. dumoffii, L. longbeachae, L. gormanii, L. anisa, and L. bozemanii. In both phylogenetic trees, L. pneumophila is found to be most closely related to L. fairfieldensis; L. Maceachernii is closest to L micdadei; and L. jordanis is in the neighborhood of L. fairfieldensis and L. maceachernii; and L. anisa is closest to L. bozemanii.
Figure 2. Unrooted phylogenetic trees constructed by the neighbor-joining method based on the ITS-tRNAAla and ITS-tRNAIle genes.

Bootstrap values were based on 1,000 replications and only values greater than 50% are shown. A, Unrooted ITS-tRNAAla gene phylogenetic tree of Legionella spp. and subspp. constructed with the neighbor-joining method. B, Unrooted ITS-tRNAIle gene phylogenetic tree of Legionella spp. and subspp. constructed with the neighbor-joining method.
Probe specificity
A total of 52 strains were used to test the specificity of the designed probes. Probes that cross-hybridized or did not produce signals were eliminated from the test panel. After the screening, 32 probes (including 29 species-specific probes, one positive control probe, one negative control probe, and one positional and printing control probe) were selected (Table 3).
The microarray specifically identified the 30 target strains. For example, L. pneumophila (including subspp. pneumophila, subspp. fraseri, and subspp. pasculleii) produced positive signals with its specific probes of OA-3815, OA-3816, and OA-3817, as well as the positive control probe OA-1993 and the positional and printing control probe Cy3 but not with the other probes [Fig. 3(1)]. Likewise, L. anisa produced positive signals with its specific probes of OA-3819 and OA-3820 [Fig. 3(2)]; L. bozemanii, with OA-3821, OA-3822, and OA-3823 [Fig. 3(3)]; L. dumoffii, with OA-3824, OA-3825, and OA-3826 [Fig. 3(4)]; L. fairfieldensis, with OA-3827, OA-3828, and OA-3829 [Fig. 3(5)]; L. gormanii, with OA-3830, OA-3831, and OA-3832 [Fig. 3(6)]; L. jordanis, with OA-3833, OA-3834, and OA-3835 [Fig. 3(7)]; L. longbeachae, with OA-3836, OA-3837, and OA-3838 [Fig. 3(8)]; L. maceachernii, with OA-3839, OA-3840, and OA-3841 [Fig. 3(9)]; L. micdadei, with OA-3839, OA-3840, and OA-3841 [Fig. 3(10)]. At the same time, none of the 22 non-target bacteria produced positive signals with the 29 Legionella spp. specific probes, demonstrating that the designed probes were species-specific.
Figure 3. Microarray differentiation of the Legionella spp.

L. pneumophila; (2) L. anisa; (3) L. bozemanii; (4) L. dumoffii; (5) L. fairfieldensis; (6) L. gormanii; (7) L. jordanis; (8) L. longbeachae; (9) L. micdadei; (10) L. maceachernii; (11) L. anisa and L. dumoffii; (12) L. anisa and L. longbeachae; (13) L. fairfieldensis, L. gormanii and L. jordanis; and (14) L. longbeachae, L. maceachernii and L. micdadei.
Microarray sensitivity
The sensitivity of the microarray analysis was tested by hybridization with serially diluted genomic template DNA at concentrations of 0.1, 1.0, 10, and 100 ng. Based on the positive signals generated, the sensitivity of the assay using genomic DNA was 1.0 ng DNA for L. pneumophila, L. longbeachae, and L micdadei, and 0.1 ng DNA for L. dumoffii (Fig. S2).
Simultaneous detection of multiple pathogens
As the detection will be more desirable if multiple pathogens can be simultaneously detected. Genomic DNA of two groups of two pathogens: L. anisa and L. dumoffii or L. anisa and L. longbeachae were mixed and used as templates for the testing. The results revealed that the probes were able to hybridize specificly the target regions of these pathogens, demonstrating the designed probes are able to detect multiple samples simultaneously [Fig. 3(11)-3(12)]. Next, genomic DNA of two groups of three pathogens: L. gormanii, L. jordanis, and L. fairfieldensis or L. maceachernii, L micdadei, and L. longbeachae, were mixed and again the microarray probes were able to successfully identify multiple pathogens simultaneously [Fig. 3(13) and 3(14)].
Blind test
The specificity and sensitivity of the microarray detection system described was blind tested. Coded DNA samples from 22 species (Table 1) were randomly selected and hybridized to the microarrays. The bacterial species selected were L. anisa (n = 1), L. bozemanii (n = 1), L. dumoffii (n = 1), L. fairfieldensis (n = 1), L. gormanii (n = 1), L. jordanis (n = 1), L. longbeachae (n = 1), L. maceachernii (n = 1), L. micdadei (n = 1), L. pneumophila (n = 3), Legionella steigerwaltii (n = 1), Legionella worsleiensis (n = 1), Escherichia coli (n = 1), Enterococcus faecalis (n = 1), Klebsiella pneumoniae (n = 1), Pseudomonas aeruginosa (n = 1), Salmonella typhi (n = 1), Staphylococcus aureus (n = 1), and Streptococcus pneumoniae (n = 1). The results matched exactly with that of the conventional detection methods (data not shown).
Test of mock samples
The mock samples containing L. bozemanii, L. dumoffii, and L. gormanii at various concentrations were tested, and the detection level was found to be at 18, 15, and 5 CFU/100 mL, respectively [Fig. 3(3), 3(4) and 3(5)]. On an average, Legionella spp. could be detected at concentrations of as low as 13 CFU/100 mL after filtering and culture enrichment.
Test of real water samples and its confirmation by sequencing
Seven samples of condensed water from air conditioners collected and provided by the Center for Disease Control and Prevention, Shanghai, China, were subjected to the microarray analysis. The hybridization profiles of the samples revealed that three of the seven samples were contaminated by L. pneumophila [Fig. 3(1)]. The remaining four samples generated signals with the positive control probe indicating the existence of bacteria other than the ten Legionella spp. (Fig. S3). The existence of L. pneumophila in the three water samples was confirmed by PCR amplification and DNA sequencing of the L. pneumophila wzt genes (23).
Discussion
Bacterial species have at least one copy of the 16S rRNA gene and the 16S-23S rDNA ITS region contains both highly conserved regions and hyper variable regions, which are useful molecular markers for bacterial identification at the species [26] and subspecies levels [27], [28], typing [29], [30], as well as in evolutionary studies [31], [32]. In this report, we describe a microarray method for the determination of pathogenic and non-pathogenic Legionella spp. on the basis of their ITS regions.
A number of techniques have been adopted to improve the reproducibility and sensitivity of the microarray First, we used a two-step PCR to amplify and label the samples. At the first step, the forward and reverse primers were used to amplify the target genes; followed by the labeling of single–strand DNA using the reverse primers at the subsequent step. The two-step PCR scheme enhances the amplification efficacy and generated intensively labeled probes as well.
The microarray is sensitive, and as little as 1.0 ng DNA or 13 CFU/100 mL can be reliably detected. This is achieved using a two-step procedure of vacuum filtering and culture process to enrich the target Legionella. After the collection of all the bacteria in the samples by the vacuum filtering, the acid-resistant Legionella were treated with HCl and selected in BCYE or GVPC [17]. The process described here allowed the detection of Legionella in a short time of 2–3 days, which is 9–10 day faster than the existing methods of identification (ISO11731∶1998).
The three probes OA-3815, OA-3816, and OA-3817 used to identify L. pneumophila are species-specific rather than subspecies-specific. Of the 18 strains of L. pneumophila obtained (Table 1), 14 were L. pneumophila subspp. pneumophila; three were L. pneumophila subspp. fraseri; and one was L. pneumophila subspp. pascullei. All the 18 strains were identified as L. pneumophila by the array. Only one or two reference strains were used for non-pneumophila Legionella species (Table 1) due to the limited availability of strains of these species in public culture collections.
In both ITS-tRNAAla and ITS-tRNAIle phylogenetic trees, the three L. pneumophila subspp., namely, pneumophila, fraseri, and pascullei, were grouped together as expected. L. pneumophila was found to be phylogenetically close to L. fairfieldensis but distant from L. longbeachae, while L micdadei was found to be in the neighborhood of these two species. As reported, most (approximately 90%) of the Legionnaire disease are caused by L. pneumophila, and of the 15 L. pneumophila serogroups identified, O1 alone is responsible for more than 84% of the cases of Legionnaire’s diseases worldwide [33], [34], [35]. Other Legionnaire’s diseases were caused by two common pathogenic L micdadei serogroups. The phenotypic characteristics of L. micdadei were reported to be quite similar to that of L. pneumophila and L. bozemanii [36]. L. longbeachae is another causative agent of Legionnaire’s diseases in Australia and New Zealand, and is associated with exposure to potting soil [37]. L. fairfieldensis was isolated from water in a cooling tower in Fairfield, Victoria, Australia, in February 1987, but has not yet been recognized as a human pathogen [6]. Although L. fairfieldensis is nonpathogenic, its ITS sequence was most closely related to L. pneumophila (the identity of 16S rRNA sequences between L. pneumophila and L. fairfieldensis is 0.909, while the identity of mip genes of these two is 0.587). A comparative study of L. pneumophila and L. fairfieldensis genomes could provide further insight into the pathogenesis of these bacteria.
Conclusion
We have sequenced and analyzed the ITS regions of 12 Legionella spp., L. anisa, L. bozemanii, L. dumoffii, L. fairfieldensis, L. gormanii, L. jordanis, L. longbeachae, L. maceachernii, L. micdadei, and L. pneumophila (including subspp. pneumophila, subspp. fraseri, and subspp. pascullei). We found that only L. jordanis contained three distinct ITS types: ITS-tRNANone (without a tRNA gene), ITS-tRNAAla (with tRNAAla gene) and ITS-tRNAIle (with tRNAIle gene), and the rest nine Legionella spp. contained two distinct ITS types: ITS-tRNAAla (with tRNAAla gene) and ITS-tRNAIle (with tRNAIle gene). Alignments of the above ITS sequences revealed significant interspecies variations of 0.266–0.772 for ITS-tRNAAla and 0.280-0.774 for ITS-tRNAIle, but low intraspecies polymorphisms of 0.967–0.993 for L. pneumophila ITS-tRNAAla and 0.943–0.996 for L. pneumophila ITS-tRNAIle, which provides a molecular basis for species-specific identification.
We have developed a rapid oligonucleotide microarray to identify the above 12 Legionella spp. and subspp. based on the polymorphism of 16–23S rRNA ITS sequences. A total of 52 strains were used to test the microarray assay, including 30 target pathogens and 22 closely-related bacteria. The 29 probes selected have reproducibly detected multiple pathogens with high specificity and sensitivity at 1.0 ng genomic DNA or 13 CFU/100 mL following filtering and culture enrichment. A 100% detection of the seven air conditioner-condensed water samples validated the microarray. Our findings revealed that the oligonucleotide microarray technique presented in this study is a promising method for basic microbiology, clinical diagnosis, food safety, and epidemiological surveillance.
In conclusion, this study presents a new PCR-based microarray assay for the comprehensive and simultaneous detection and identification of ten Legionella spp. This new method provides an accurate and reliable approach to differentiate among Legionella isolates at the species level; contributes significantly to large-scale epidemiology studies; and can be used to monitor local, regional, and national trends in human legionellosis.
Supporting Information
Agarose gel electrophoresis of PCR products of Legionella spp. ITS regions. Lanes: M, molecular weight standards (DL2000 Marker); (1) L. anisa; (2) L. bozemanii; (3) L. dumoffii; (4) L. fairfieldensis; (5) L. gormanii; (6) L. jordanis; (7) L. longbeachae; (8) L. maceachernii; (9) L. micdadei; and (10) L. pneumophila.
(TIF)
The sensitivity of the microarray analysis with genomic DNA of L. dumoffii . (1) 100 ng; (2) 10 ng; (3) 1.0 ng; and (4) 0.1 ng.
(TIF)
Microarray pattern of bacteria other than the ten Legionella spp.
(TIF)
Data Availability
The authors confirm that all data underlying the findings are fully available without restriction. The ITS sequences were deposited into the GenBank database under the accession numbers KM609984-KM610004, and the microarray dataset was deposited into the Gene Expression Omnibus database under the accession number GSE61962.
Funding Statement
This work was supported by grants from the National Key Program for Infectious Diseases of China (2013ZX10004216-001-001), the National 863 Program of China (2012AA020103, 2011AA100901-2), the National 973 Program of China (2011CB504900), the National Natural Science Foundation of China (NSFC) Program (81171524, 31270144, 31170094, 31030002 and 31270003), and the Tianjin Research Program of Application Foundation and Advanced Technology (13JCQNJC09400, 12JCYBJC15100 and 10JCYBJC10000). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Newton HJ, Ang DK, van Driel IR, Hartland EL (2010) Molecular pathogenesis of infections caused by Legionella pneumophila . Clin Microbiol Rev 23:274–298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Neil K, Berkelman R (2008) Increasing incidence of legionellosis in the United States, 1990–2005: changing epidemiologic trends. Clin Infect Dis 47:591–599. [DOI] [PubMed] [Google Scholar]
- 3. Palusińska-Szysz M, Cendrowska-Pinkosz M (2009) Pathogenicity of the family Legionellaceae. Arch Immunol Ther Exp (Warsz) 57:279–290. [DOI] [PubMed] [Google Scholar]
- 4. Muder RR, Yu VL (2002) Infection due to Legionella species other than L. pneumophila . Clin Infect Dis 35:990–998. [DOI] [PubMed] [Google Scholar]
- 5. Berger P, Papazian L, Drancourt M, La Scola B, Auffray JP, et al. (2006) Ameba-associated microorganisms and diagnosis of nosocomial pneumonia. Emerg. Infect Dis 12:248–255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Fields BS, Benson RF, Besser RE (2002) Legionella and Legionnaires’ disease: 25 years of investigation. Clin Microbiol Rev 15:506–526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Inoue H, Noda A, Takama T, Ishima T, Agata K (2006) Enhanced antifungal effect of the selective medium for the detection of Legionella species by a combination of cycloheximide, amphotericin B and thiabendazole. Biocontrol Sci 11:69–74. [DOI] [PubMed] [Google Scholar]
- 8. Murdoch DR (2003) Diagnosis of Legionella infection. Clin Infect Dis 36:64–69. [DOI] [PubMed] [Google Scholar]
- 9. Doleans A, Aurell H, Reyrolle M, Lina G, Freney J, et al. (2004) Clinical and environmental distributions of Legionella strains in France are different. J Clin Microbiol 42:458–460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Qin T, Yan G, Ren H, Zhou H, Wang H, et al. (2013) High prevalence, genetic diversity and intracellular growth ability of Legionella in hot spring environments. PLOS One 8:e59018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Stolhaug A, Bergh K (2006) Identification and differentiation of Legionella pneumophila and Legionella spp. with real-time PCR targeting the 16S rRNA gene and species identification by mip sequencing. Appl Environ Microbiol 72:6394–6398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Yang G, Benson R, Pelish T, Brown E, Winchell JM, et al. (2010) Dual detection of Legionella pneumophila and Legionella species by real-time PCR targeting the 23S-5S rRNA gene spacer region. Clin Microbiol Infect 16:255–261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Su HP, Tung SK, Tseng LR, Tsai WC, Chung TC, et al. (2009) Identification of Legionella species by use of an oligonucleotide array. J Clin Microbiol 47:1386–1392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Wilson DA, Yen-Lieberman B, Reischl U, Gordon SM, Procop GW (2003) Detection of Legionella pneumophila by real-time PCR for the mip gene. J Clin Microbiol 41:3327–3330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15. Ko KS, Hong SK, Lee KH, Lee HK, Park MY, et al. (2003) Detection and identification of Legionella pneumophila by PCR-restriction fragment length polymorphism analysis of the RNA polymerase gene (rpoB). J Microbiol Methods 54:325–337. [DOI] [PubMed] [Google Scholar]
- 16. Zhou G, Cao B, Dou Y, Liu Y, Feng L, et al. (2011) PCR methods for the rapid detection and identification of four pathogenic Legionella spp. and two Legionella pneumophila subspecies based on the gene amplification of gyrB . Appl Microbiol Biotechnol 91:777–787. [DOI] [PubMed] [Google Scholar]
- 17. Zhou G, Wen S, Liu Y, Li R, Zhong X, et al. (2011) Development of a DNA microarray for detection and identification of Legionella pneumophila and ten other pathogens in drinking water. Int J Food Microbiol 145:293–300. [DOI] [PubMed] [Google Scholar]
- 18. Herpers BL, de Jongh BM, van der Zwaluw K, van Hannen EJ (2003) Real-time PCR assay targets the 23S-5S spacer for direct detection and differentiation of Legionella spp. and Legionella pneumophila . J Clin Microbiol 41:4815–4816. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Cianciotto NP, Eisenstein BI, Mody CH, Toews GB, Engleberg NC (1989) A Legionella pneumophila gene encoding a species-specific surface protein potentiates initiation of intracellular infection. Infect Immun 57:1255–1262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20. Cianciotto NP, O’Connell W, Dasch GA, Mallavia LP (1995) Detection of mip-like sequences and Mip-related proteins within the family Rickettsiaceae. Curr Microbiol 30:149–153. [DOI] [PubMed] [Google Scholar]
- 21. Ratcliff RM, Donnellan SC, Lanser JA, Manning PA, Heuzenroeder MW (1997) Interspecies sequence differences in the Mip protein from the genus Legionella: implications for function and evolutionary relatedness. Mol Microbiol 25:1149–1158. [DOI] [PubMed] [Google Scholar]
- 22. Guan W, Xu Y, Chen DL, Xu JN, Tian Y, et al. (2012) Application of multilocus sequence analysis (MLSA) for accurate identification of Legionella spp. Isolated from municipal fountains in Chengdu, China, based on 16S rRNA, mip, and rpoB genes. J Microbiol 50:127–136. [DOI] [PubMed] [Google Scholar]
- 23. Cao B, Yao F, Liu X, Feng L, Wang L (2013) Development of a DNA microarray method for detection and identification of All 15 distinct O-antigen forms of Legionella pneumophila . Appl Environ Microbiol 79:6647–6654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Wang M, Cao B, Yu Q, Liu L, Gao Q, et al. (2008) Analysis of the 16S-23S rRNA gene internal transcribed spacer region in Klebsiella species. J Clin Microbiol 46:3555–3563. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25. Cao B, Wang M, Liu L, Zhou Z, Wen S, et al. (2009) 16S-23S rDNA internal transcribed spacer regions in four Proteus species. J Microbiol Methods 77:109–118. [DOI] [PubMed] [Google Scholar]
- 26. Magray MS, Kumar A, Rawat AK, Srivastava S (2011) Identification of Escherichia coli through analysis of 16S rRNA and 16S-23S rRNA internal transcribed spacer region sequences. Bioinformation. 6:370–371. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Chun J, Rivera IN, Colwell RR (2002) Analysis of 16S–23S rRNA intergenic spacer of Vibrio cholerae and Vibrio mimicus for detection of these species. Methods Mol Biol 179:171–178. [DOI] [PubMed] [Google Scholar]
- 28. Xiong L, Kong F, Yang Y, Cheng J, Gilbert GL (2006) Use of PCR and reverse line blot hybridization macroarray based on 16S–23S rRNA gene internal transcribed spacer sequences for rapid identification of 34 mycobacterium species. J Clin Microbiol 44:3544–3550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Maggi RG, Chomel B, Hegarty BC, Henn J, Breitschwerdt EB (2006) A Bartonella vinsonii berkhoffii typing scheme based upon 16S–23S ITS and Pap31 sequences from dog, coyote, gray fox, and human isolates. Mol Cell Probes 20:128–134. [DOI] [PubMed] [Google Scholar]
- 30. Wojciech L, Staroniewicz Z, Jakubczak A, Ugorski M (2004) Typing of Yersinia Enterocolitica Isolates by ITS profiling, REP- and ERIC-PCR. J Vet Med B Infect Dis Vet Public Health 51:238–244. [DOI] [PubMed] [Google Scholar]
- 31. Gurtler V (1999) The role of recombination and mutation in 16S–23S rDNA spacer rearrangements. Gene 238:241–252. [DOI] [PubMed] [Google Scholar]
- 32. Perez Luz S, Rodriguez-Valera F, Lan R, Reeves PR (1998) Variation of the ribosomal operon 16S–23S gene spacer region in representatives of Salmonella enterica subspecies. J Bacteriol 180:2144–2151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33. Carvalho FR, Nastasi FR, Gamba RC, Foronda AS, Pellizari VH (2008) Occurrence and diversity of Legionellaceae in polar lakes of the Antarctic peninsula. Curr Microbiol 57:294–300. [DOI] [PubMed] [Google Scholar]
- 34. Daniels C, Vindurampulle C, Morona R (1998) Overexpression and topology of the Shigella flexneri O-antigen polymerase (Rfc/Wzy). Mol Microbiol 28:1211–1222. [DOI] [PubMed] [Google Scholar]
- 35. Doleans A, Aurell H, Reyrolle M, Lina G, Freney J, et al. (2004) Clinical and environmental distributions of Legionella strains in France are different. J Clin Microbiol 42:458–460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36. Thacker WL, Benson RF, Hawes L, Gidding H, Dwyer B, et al. (1991) Legionella fairfieldensis sp. nov. isolated from cooling tower waters in Australia. J Clin Microbiol. 29:475–478. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37. Wright AJ, Humar A, Gourishankar S, Bernard K, Kumar D (2012) Severe Legionnaire’s disease caused by in a long-term renal transplant patient: the importance of safe living strategies after transplantation. Transpl Infect Dis 14:E30–E33. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Agarose gel electrophoresis of PCR products of Legionella spp. ITS regions. Lanes: M, molecular weight standards (DL2000 Marker); (1) L. anisa; (2) L. bozemanii; (3) L. dumoffii; (4) L. fairfieldensis; (5) L. gormanii; (6) L. jordanis; (7) L. longbeachae; (8) L. maceachernii; (9) L. micdadei; and (10) L. pneumophila.
(TIF)
The sensitivity of the microarray analysis with genomic DNA of L. dumoffii . (1) 100 ng; (2) 10 ng; (3) 1.0 ng; and (4) 0.1 ng.
(TIF)
Microarray pattern of bacteria other than the ten Legionella spp.
(TIF)
Data Availability Statement
The authors confirm that all data underlying the findings are fully available without restriction. The ITS sequences were deposited into the GenBank database under the accession numbers KM609984-KM610004, and the microarray dataset was deposited into the Gene Expression Omnibus database under the accession number GSE61962.