

Abstract
Background
Necroptosis is an important mode of cell death, which is due to oxidant stress accumulation. Our previous study indicated that oxidant stresses could be reduced by Timosaponin B-II (TBII), a kind of Chinese herb RhizomaAnemarrhenae monomer extraction. We wonder the possible effect of Timosaponin B-II, whether it can protect cells from necroptosis via reducing the oxidant stress, in RGC-5 following hydrogen peroxide (H2O2) insult.
Methods
RGC-5 cells were grown in DMEM, the model group was exposed in H2O2 with the concentration of 300 μM, and the experimental group was pre-treated with Timosaponin B-II at different concentrations (1 μM, 10 μM, 100 μM and 1000 μM) for 24 hrs. MTT assay was carried out to measure the cytotoxicity of H2O2, MDA concentration assay was executed to evaluate the degree of oxidative stress, TNF-α ELISA Assay was used to measure the concentration of TNF-α, finally, the degree of necrosis were analyzed using flow cytometry.
Results
We first constructed the cell injury model of necroptosis in RGC-5 upon H2O2 exposure. Morphological observation and MTT assay were used to evaluate the degree of RGC-5 death. MDA assay were carried out to describe the degree of oxidant stress. Annexin V/PI staining was used to detect necroptotic cells pre-treated with or without Timosaponin B-II following H2O2 injury. TNF-α ELISA was carried out to detect the TNF-α accumulation in RGC-5. Upon using Timosaponin B-II with concentration of 100 μM, the percentage of cell viability was increased from 50% to 75%, and the necrosis of cells was reduced from 35% to 20% comparing with H2O2 injury group. Oxidant stress and TNF-α was reduced upon injury which decreased the ratio of RGC-5 necroptosis.
Conclusion
Our study found out that Timosaponin B-II might reduce necroptosis via inhibition of ROS and TNF-α accumulation in RGC-5 following H2O2 injury.
Keywords: Retinal ganglion cells-5, Timosaponin B-II, TNF-α, Necroptosis, Oxidative stress
Background
Necroptosis is a novel kind of cell death which has similar morphological features of necrosis [1, 2]. The previous studies pointed out that necroptosis was usually initiated by TNF-α or FasL, etc. and transmitted by certain specific cytokines, which results in the accumulation of oxidative products and eventually leading to necrosis. However, it can be blocked by necrostatin-1 (Nec-1, a specific inhibitor of necroptosis) [1]. Necroptosis existed in different cell types, such as epidermal keratinocytes [3], Jurkat T cells [4], L929 cells [5, 6], HepG2 cells [7], hippocampal neurons [8], cortical neurons [9] and photoreceptor cells [10], etc. upon injury. Furthermore, Rosenbaum, et al. have found a large number of necrotic cells exist in rat retinal ganglion cell layer by PI staining at the early stage of acute high intra-ocular pressure (aHIOP) [11]. The numbers of PI-positive neuron decreased, and the "b" wave was partially restored in flash ERG with Nec-1 pretreatment. Dvoriantchikova’s recent works showed that retinal ganglion cells (RGCs) necroptosis promoted in aHIOP-induced retinal damage [12], which has the similar finding with our previous work [13]. Additionally, our recent studies have also indicated that necroptosis occurs in retinal ganglion cells-5 (RGC-5) at an early stage following elevated hydrostatic pressure (EHP) in vitro[14] or hydrogen peroxide (H2O2) treatment (our unpublished data) detected by flow cytometry (a typical way to monitor necroptosis) [15].
Although some scientists recently have synthesized necroptosis inhibitors, such as Nec-3 [16], Nec-5 [17], Nec-7 [18], Nec-21 [19], etc., the mechanism still needs to be further clarified. Nec-1 is a synthetic small molecular compound which is the most commonly used in necroptosis inhibition [1]. So far, there are no reports about whether it has side effects when it is applied to animal or cellular models. Traditional Chinese herb rhizomaanemarrhenae is the dried rhizome of anemarrhenaasphodeloidesbge. It contains lots of steroidal saponins including sarsasapogenin, markosapogenin, negitogenin, diosgenin and its glycosylated derivatives, like Timosaponin A-I, A-II, A-III, A-IV, B-I, B-II,C, D and Timosaponin E [20]. Li [21] indicated that Timosaponin B-II (Figure 1, the chemical structure of Timosaponin B-II, CAS number: 136656-07-0) might improve the impairment of learning and memory caused by cerebral ischemia in a dose-dependent manner. Lu [11] and Kim’s [22] research suggested that, Timosaponin B-II could not only inhibit the production of IL-1β and IL-6, but also TNF-α (one of important molecules which initiates necroptosis), these results suggested the effect might be related to Timosaponin B-II which has anti-inflammatory activity. Besides, Zhang [23] and Kaname [24] found Timosaponin E-I, E-II, B-II, B-III and A-III played an important role in superoxide dismutase-generation. Deng [25] demonstrated that culturing primary rat neurons (Aβ25–35 insult model) with Timosaponin B-II at a certain range of concentrations showed remarkable anti-oxidative damage effects, which indicated Timosaponin B-II could remove oxygen radicals, and keep the intra-cellular redox reactions in a dynamic equilibrium [26]. Our previous study also showed that Timosaponin B-II participated in the protection of rat RGCs which was treated by FeCl3 solution via anti-oxidation [27]. Of note, whether Timosaponin B-II could inhibit oxidative stress-induced RGCs necroptosis or not, it is still under investigation. Therefore, our present experiments focused on Timosaponin B-II’s potential roles in RGC-5 necroptosis suppression upon hydrogen peroxide (H2O2) treatment. Meanwhile, the effect of Timosaponin B-II on malondialdehyde (MDA, biomarker of oxidative stress) and TNF-α in RGC-5 has been investigated. Our study may give a better understanding of the protective effect of Timosaponin B-II upon neuronal injury, and provide the experimental basis of novel mechanism on cell death.
Figure 1.
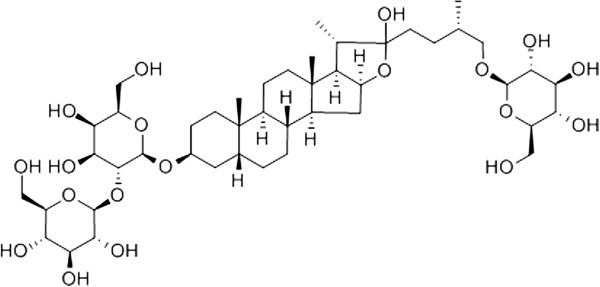
The chemical structure of Timosaponin B-II.
Methods
Cell culture
The mouse retinal ganglion cell line (RGC-5) was contributed by Department of Ophthalmology, Second Hospital of Ji Lin University, China [28]. RGC-5 cells were grown in Dulbecco’s Modified Eagle Medium (DMEM, HyClone Laboratories, Inc. UT, USA) and supplemented with 10% fetal bovine serum (FBS, HyClone Laboratories, Inc. UT, USA), 100 U/ml of penicillin and 100 μg/ml of streptomycin (HyClone Laboratories, Inc. UT, USA). The RGC-5 cells used in the experiment were with 2-3 passages post-thawed to minimize the variability in the assays based on our observations. The density of RGC-5 cells was around 80% in 6 ml culture media in 50 ml flask before insults (H2O2 treatment).
Reagent
The powder of Timosaponin B-II was provided by Professor Wan-Sheng Chen from Department of Pharmacology, School of Pharmacology, Second Military Medical University, Shanghai, China [29, 30] and its purity is above 98%. Timosaponin B-II was dissolved in sterile normal saline (NS) at 2 mM in room temperature (24°C), H2O2 (Sigma-Aldrich, MO, USA) with 0.01 M PBS at 3 mM in store to yield a low concentration working solutions.
Cell model construction and drug treatment
Cells were equally divided into three groups randomly. The normal control group (CTL), the model group (H2O2, 300 μM), and the experimental group (H2O2 + Timosaponin B-II). The experimental group was pre-treated with Timosaponin B-II at different concentrations (1 μM, 10 μM, 100 μM and 1000 μM) for 24 hrs. No drugs were exposed to the normal control or the model group. After that, we used H2O2 of 300 μM for 12 hrs to get cell insult and randomly selected three in each group for morphological studies while the remaining cells were used for biochemistry studies. For morphological study, the pictures of the attached cells were captured using inverted microscope in 10 × objective (Olympus, CKX41, Tokyo, Japan). We have captured ten pictures at least in each group, and selected the typical graphics to illustrate.
MTT assay
Cytotoxicity of H2O2 model group and Timosaponin B-II pretreated group (1 μM, 10 μM, 100 μM and 1000 μM) were assessed in RGC-5 cells by measuring the amount of insoluble formazan formed in live cells based on the reduction of 3-(4, 5 dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) salt (Nanjing Jian-Cheng Bio-engineering Institute, Jiangsu, China) according to the manufacturer's protocol. The cells were seeded in 96-well plates with a density of 104 cells/well incubated for 24 hrs at 37°C and 5% CO2. The cells were pretreated with different concentrations of Timosaponin B-II before H2O2 insult or H2O2 used alone and PBS as a negative control. Within 24 hrs pre-treatment of Timosaponin B-II, 50 μl of MTT labeling reagent (2 μg/ml) was added to each well. The plates were incubated at 37°C in a humidified atmosphere with 5% CO2 for 4 hrs. Thereafter, 100 μl of the solubilization solution was added to each well and followed by incubation overnight at 37°C to dissolve formazan crystals. Absorbance was ultimately read using an ELISA plate reader (Bio-tek, ELx800, IL, USA) at a wavelength of 570 nm. Where, H2O2 model group and Timosaponin B-II pretreated group are mean absorbance of treated cells and negative control, respectively.
MDA concentration assay
MDA levels in RGC-5 extractions were assayed using a commercial kit according to manufacturer’s instructions (Nanjing Jian-Cheng Biotechnical Co., Jiangsu, China) as in our previous study [27], the standard reference substance named tetraethoxypropane were used in 10 nmol/ml. Equal quantities (100 μg) of protein were loaded in each well and each analysis performed in duplicate.
TNF-α ELISA assay
A RGC-5 TNF-α concentration assay was performed using a commercial TNF-α ELISA kit (Invitrogen, CA, USA). The detailed processes were conducted according to the manual included in the kit. Positive control: the antibody tested in the kit was replaced by mouse TNF-α (provided in assay, concentration: 720 ng/L). Equal quantities of protein (80 μg) were analyzed in every tested well and the measurements were carried out by Bio-tek microplate-reader (ELx800, IL, USA). The percentage of TNF-α concentration in normal control group was set as 100%. All experiments were repeated at least twice.
Flow cytometry
The cells attached to the flasks were trypsinized followed by a gentle wash. The experimental group was pre-treated with Timosaponin B-II at 100 μM. The model group was treated with H2O2 at 300 μM. Resuspended the cells in 200 μl of 1× binding buffer, and then added 5 μl of 20 μg/ml Annexin V and 10 μl of 50 mg/ml PI, incubated at RT for 15 mins in the dark. After the cells were washed and analyzed by FACS Calibur (Becton, Dickinson Company, NJ, USA). The percentages of cells in each quadrant were analyzed using ModFit software (Verity Software House Topsham, NJ, USA). Statistical results of flow cytometry were conducted by calculating the PI+ cells numbers. All the results were repeated three times.
Data analysis
Figure panels were assembled by using Photoshop CC (Adobe, CA, USA). The data were analyzed by using SPSS 19.0 (SPSS, IL, USA). One-way analysis of variance (one-way ANOVA) was performed to test differences in average value between groups. All results were presented as mean ± SD. A value of p < 0.05 was considered statistically significant.
Results
Timosaponin B-II could protect RGC-5 from H2O2 injury
The normal RGC-5 cell graphic were shown in Figure 2A. The cells grew in apposite density and adherent well with axon-elongated and interweave each other to be a mesh, each of them presented as identical size and consistent morphology. The number of the cells decreased in H2O2 injury group (Figure 2B), and plenty of them were detached and floated in culture media. As shown in Figure C-E, the number of adherent cells increased following the incubated concentration of Timosaponin B-II increase, it is worth noting that the number of adherent cells in 1000 μM Timosaponin B-II treatment group is less than those in 100 μM Timosaponin B-II treatment group (Figure E and F). These results indicated that Timosaponin B-II could protect RGC-5 from H2O2 insult yield to the concentration manner, but the effect may reverse with higher concentrations of Timosaponin B-II in 1000 μM.
Figure 2.
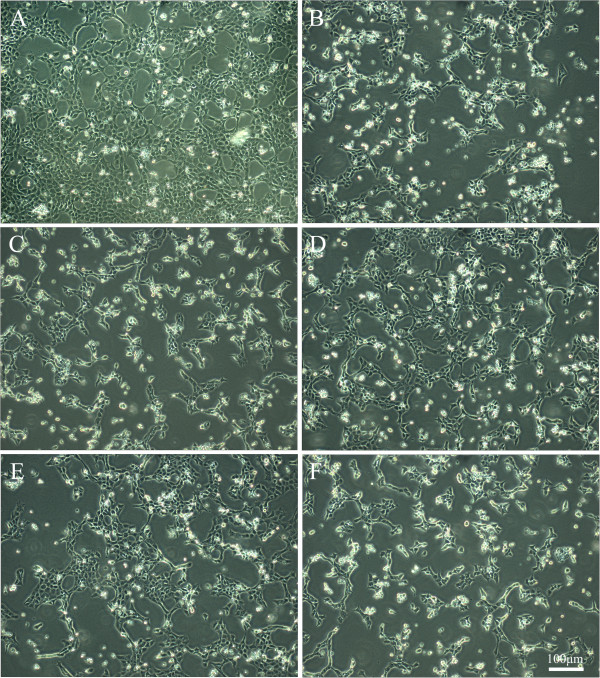
The morphological graphic of RGC-5 cell line insult with H 2 O 2 after 12 hrs pretreating in different concentration of TBII by inverted microscope; A: Normal Control; B: Model group (H 2 O 2 treatment); C: TBII pretreatment group (H 2 O 2 + 1 μM TBII); D: TBII pretreatment group (H 2 O 2 + 10 μM TBII); E: TBII pretreatment group (H 2 O 2 + 100 μM TBII); F: TBII pretreatment group (H 2 O 2 + 1000 μM TBII), Scale bar = 100 μm in A-E.
Timosaponin B-II could increase RGC-5 viability upon H2O2 injury
In order to evaluate the viability of RGC-5 in model group and Timosaponin B-II treatment group, we carried out MTT assay to detect cell viability. The statistical analysis of MTT result was shown in Figure 3. Comparing to normal control group (CTL), the cell viability was significantly decreased in model group and Timosaponin B-II treatment group. The cell viability kept gradually increasing upon elevating the concentration of Timosaponin B-II except 1000 μM Timosaponin B-II group. These results indicated that Timosaponin B-II could rescue RGC-5 viability, which was impaired by the H2O2.
Figure 3.
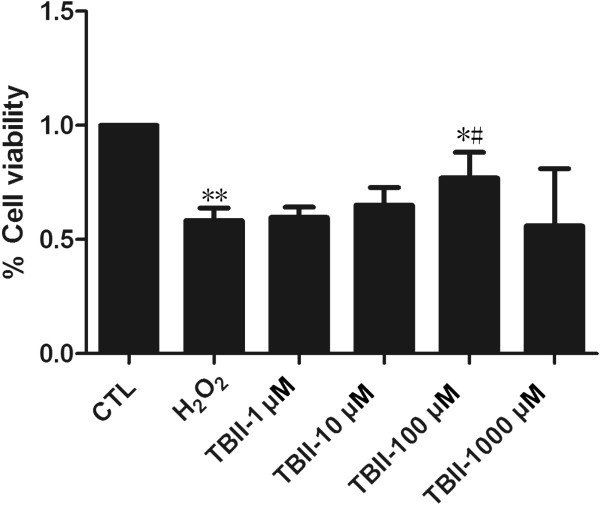
The percentage of MTT in RGC-5 cell line insult with H 2 O 2 after 12 hrs pretreating in different concentration of TBII (Normal Control were set as 100%). **vs CTL: p < 0.01; # vs Model group: p < 0.05; *vs CTL: p < 0.05.
Timosaponin B-II reduced MDA production in RGC-5
The statistical analysis of MDA level is shown in Figure 4. The level of MDA in the experimental group (except 1000 μM Timosaponin B-II treatment group) kept decreasing gradually compared to the model group upon H2O2 insult after increasing the concentration of Timosaponin B-II, but it remained at higher level than those in normal control group (CTL). In 100 μM Timosaponin B-II treatment group, the level of MDA decreased significantly compared with H2O2 insult group (p < 0.05). These results indicated that Timosaponin B-II reduced the oxidative stress which is usually induced by H2O2 in RGC-5. Nevertheless, the capacity of anti-oxidant was limited, because the oxidative stress remained higher than normal control group in Timosaponin B-II treatment group.
Figure 4.
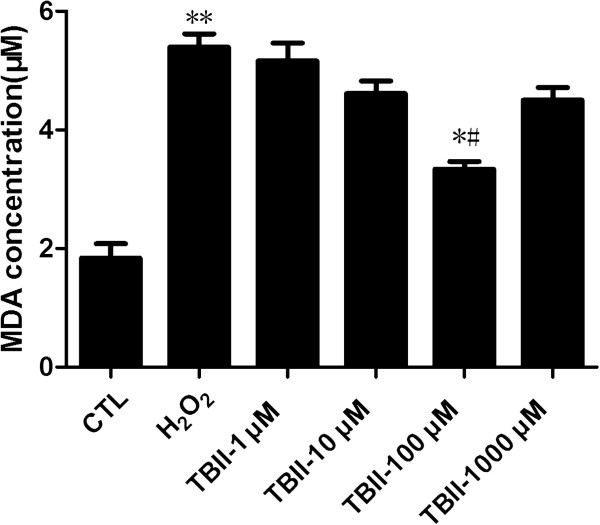
The concentration of MDA in RGC-5 cell line insult with H 2 O 2 after 12 hrs pretreating in different concentration of TBII. **vs CTL: p < 0.01; # vs Model group: p < 0.05; *vs CTL: p < 0.05.
Timosaponin B-II inhibited TNF-α production in RGC-5
The statistical analysis of ELISA assay for TNF-α level was described in Figure 5. The level of TNF-α in the experimental group (except 1000 μM Timosaponin B-II treatment) was gradually decreasing compared to the model group after increasing the concentration of Timosaponin B-II, but it remained at higher levels compared to those in normal control group. These results indicated that Timosaponin B-II remarkably inhibited TNF-α production. Nevertheless, the effect of Timosaponin B-II was impaired to a certain extent since the level of target peptides was still higher than those in normal condition.
Figure 5.
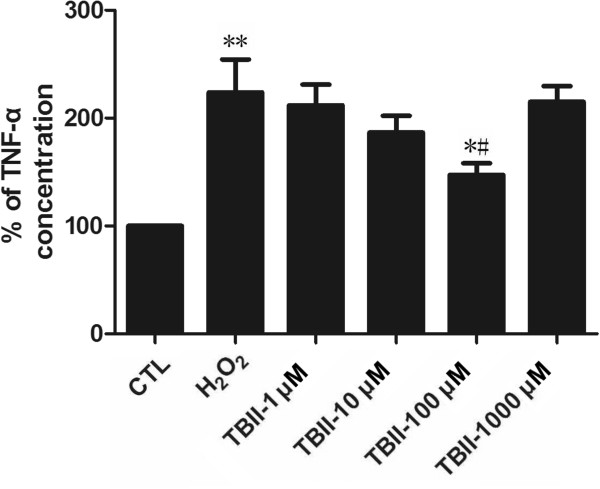
The percentage of TNF-α concentration in RGC-5 cell line insult with H 2 O 2 after 12 hrs pretreating in different concentration of TBII. **vs CTL: p < 0.01; # vs Model group: p < 0.05; *vs CTL: p < 0.05.
Timosaponin B-II may decrease the rate of RGC-5 necrosis
Upon 300 μM H2O2 insult, necrosis in RGC-5 occurred in our study. Therefore, the cells were treated under this condition with the addition of Timosaponin B-II at different concentrations. After that, we analyzed cellular necrosis by using flow cytometry with PI/Annexin V double staining, and detected whether it could decrease the rate of necrosis in RGC-5 with pretreatment of Timosaponin B-II upon H2O2 insult. These results showed that the ratio of necrosis cells is 30.3% (Figure 6B), the percentage decreased to 22.8% upon adding Timosaponin B-II (100 μM, Figure 6C). Meanwhile, statistical analysis indicated that there were significant changes in the rate of PI-positive RGC-5 in Timosaponin B-II pretreatment group compared with normal control group and H2O2 model group (Figure 6D). These results indicated that RGC-5 necroptosis in the early stage may decrease with Timosaponin B-II usage.
Figure 6.
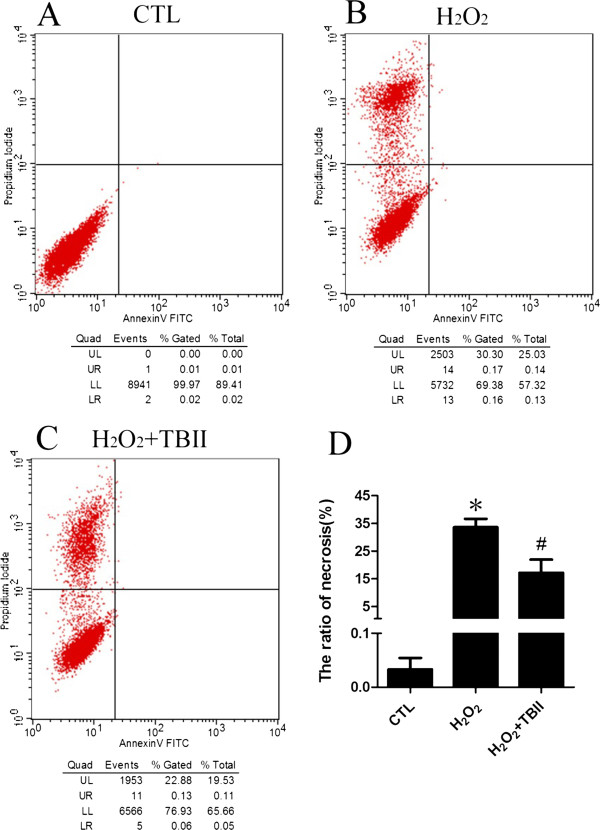
The flow Cytometry analysis of RGC-5 cell line insult with H 2 O 2 after 12 hrs pretreating in different concentration of TBII. A: Normal Control; B: Model group (H2O2 treatment); C: TBII pretreatment group (H2O2 + 100 μM TBII); D: Statistical analysis; *vs CTL: p < 0.01; # vs Model group: p < 0.01.
Discussion
At present, mechanism of cell death is one of the hotspot in the field of life science research. As far as we are concerned, compared to apoptosis and autophagy, the researchers did not pay enough attention to necrosis due to the traditional viewpoint that it cannot be modulated and intervened. Necroptosis is the latest cell death mode which was conceptualized in last century in mid-1980s [31]. Since then, it started to catch more and more attention [32–34]. We speculate that damaged cells could be rescued if necroptosis can be intervened at an early stage, and it will also help us find a better strategy for rational interventional therapy in the future.
Chinese herb has a wealth of resources from natural materials with lower side effects, more economical and other advantages, etc. However, it also has the disadvantages such as its complicated composition which cannot be identified easily. Timosaponin B-II is a monomer which is extracted from Chinese herb rhizomaanemarrhenae, it has multiple pharmacological effects, including anti-inflammatory [21, 35], anti-diabetic [36], anti-oxidative stress [23, 26, 27], anti-senile dementia [37], learning ability or memory improvement [21] and neuro-protection [27] or anti-apoptosis effect in human umbilical vein endothelial cells [38], etc. Our MTT assay results showed that viability of RGC-5 in model group (H2O2 treatment) was significantly decreased, while RGC-5 was pre-incubated by Timosaponin B-II (1 μM, 10 μM and 100 μM groups) for 24 hrs, RGC-5 proliferation increased gradually in a dose-dependent manner. The RGC-5 proliferation of 100 μM treatment group reached a peak, but it significantly decreased in 1000 μM treatment group. The morphological results suggested that the number of RGC-5 decreased remarkably in model group, the survival of cells tends to be similar as MTT when pretreated with Timosaponin B-II. Taken all together, it indicated that Timosaponin B-II played a protective role on RGC-5 in H2O2-induced damage under a certain concentration. In addition, our results in flow cytometry showed that the number of RGC-5 cell necrosis significantly reduced in 100 μM Timosaponin B-II pretreated group, which suggested that Timosaponin B-II may be partly involved in inhibiting RGC-5 necroptosis under H2O2 exposed conditions. Moreover, our results on oxidation products measurement also showed that Timosaponin B-II could significantly decrease the accumulation of MDA in RGC-5 in certain range of concentrations. Furthermore, we had observed that TNF-α was also inhibited in Timosaponin B-II pretreatment group. Considering one of the most important pathways in necroptosis (TNF-α induced necroptosis pathway initiate-----necroptosis related molecular modulation-----ROS accumulation------cell necrosis [39, 40]), we speculated that Timosaponin B-II might inhibit the RGC-5 necroptosis by reducing oxidation products and TNF-α level.
Finally, it is worthy of noting that our experiments revealed the following two questions. Firstly, although Timosaponin B-II can reduce necroptosis by resisting oxidative stress and reduce pro-inflammatory molecule, but it is impossible to recover to normal level, which indicates its limited function as monomer herbal extraction, it also indirectly supports the compatibility usage of Chinese herbal drugs. Secondly, our results showed that higher concentrations of Timosaponin B-II (e.g. 1000 μM) could be toxic. Frank’s research showed Timosaponin B-II and Timosaponin A-III have the similar structure (such as steroid), but Timosaponin B-II presents one more glycosyl [41]. Timosaponin B-II has the trend to convert into Timosaponin A-III. However higher concentrations of Timosaponin A-III has been regarded as highly cytotoxic [41–43] and could cause cell death instead of protective effect. Therefore, we would consider discarding this concentration (1000 μM) in future studies. Until now, the molecule of regulation mechanism for cellular necroptosis upon injury was concerned, including receptor interacting proteins (RIPs) [8, 10, 12, 13], calpains [14, 44], CDGSH iron-sulfur domain-containing protein 1 (CISD1) [45] or ubiquitin C-terminal hydrolase (UCH-L1) [46], etc. Therefore, whether the possible pathway of Timosaponin B-II neuro-protection may mediate by these molecules mentioned above needs further investigation.
Conclusion
Timosaponin B-II with limited concentrations can partially protect RGC-5 from H2O2 induced-necroptosis, which may related to TNF-α and ROS accumulation inhibition.
Acknowledgements
This work is supported by National Natural Science Foundation of China (#81371011), Wu Jie-Ping Medical Foundation of the Minister of Health of China (320.6750.14118), Natural Science Foundation of Hunan Province (#2015JJ2187) and Young Teachers Training Program of Normal University of Hunan Province to KX. We are grateful for the provision of Timosaponin B-II from Professor Wan-Sheng Chen in Department of Pharmacology, School of Pharmacology, Second Military Medical University, Shanghai, China.
Footnotes
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
LS, J-FH and KX designed the experiment. S-HJ, LS and WD performed the experiment. S-HJ, LS and R-FM drafted the manuscript. SC analyzed the data. KX revised the manuscript and participated in paper modification. L-XX revised the manuscript for English writing. All authors participated in critical revision of the manuscript and approved the final manuscript.
Contributor Information
San-Hong Jiang, Email: 1462555868@qq.com.
Lei Shang, Email: shanglei1986@163.com.
Li-Xiang Xue, Email: lixiangxue@hsc.pku.edu.cn.
Wei Ding, Email: ldd2007200@163.com.
Shuang Chen, Email: 405237696@qq.com.
Ruo-Fei Ma, Email: 958888913@qq.com.
Ju-Fang Huang, Email: huangjufang@csu.edu.cn.
Kun Xiong, Email: xiongkun2001@163.com.
References
- 1.Wu W, Liu P, Li J. Necroptosis: an emerging form of programmed cell death. Crit Rev Oncol Hematol. 2012;82(3):249–258. doi: 10.1016/j.critrevonc.2011.08.004. [DOI] [PubMed] [Google Scholar]
- 2.Han J, Zhong CQ, Zhang DW. Programmed necrosis: backup to and competitor with apoptosis in the immune system. Nat Immunol. 2011;12(12):1143–1149. doi: 10.1038/ni.2159. [DOI] [PubMed] [Google Scholar]
- 3.Bonnet MC, Preukschat D, Welz PS, van Loo G, Ermolaeva MA, Bloch W, Haase I, Pasparakis M. The adaptor protein FADD protects epidermal keratinocytes from necroptosis in vivo and prevents skin inflammation. Immunity. 2011;35(4):572–582. doi: 10.1016/j.immuni.2011.08.014. [DOI] [PubMed] [Google Scholar]
- 4.Song KJ, Jang YS, Lee YA, Kim KA, Lee SK, Shin MH. Reactive oxygen species-dependent necroptosis in Jurkat T cells induced by pathogenic free-living Naegleria fowleri. Parasite Immunol. 2011;33(7):390–400. doi: 10.1111/j.1365-3024.2011.01297.x. [DOI] [PubMed] [Google Scholar]
- 5.Wu YT, Tan HL, Huang Q, Sun XJ, Zhu X, Shen HM. zVAD-induced necroptosis in L929 cells depends on autocrine production of TNFalpha mediated by the PKC-MAPKs-AP-1 pathway. Cell Death Differ. 2011;18(1):26–37. doi: 10.1038/cdd.2010.72. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Vanlangenakker N, Bertrand MJ, Bogaert P, Vandenabeele P, Vanden Berghe T. TNF-induced necroptosis in L929 cells is tightly regulated by multiple TNFR1 complex I and II members. Cell Death Dis. 2011;2:e230. doi: 10.1038/cddis.2011.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Jouan-Lanhouet S, Arshad MI, Piquet-Pellorce C, Martin-Chouly C, Le Moigne-Muller G, Van Herreweghe F, Takahashi N, Sergent O, Lagadic-Gossmann D, Vandenabeele P, Samson M, Dimanche-Boitrel MT. TRAIL induces necroptosis involving RIPK1/RIPK3-dependent PARP-1 activation. Cell Death Differ. 2012;19(12):2003–2014. doi: 10.1038/cdd.2012.90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Vieira M, Fernandes J, Carreto L, Anuncibay-Soto B, Santos M, Han J, Fernandez-Lopez A, Duarte CB, Carvalho AL, Santos AE. Ischemic insults induce necroptotic cell death in hippocampal neurons through the up-regulation of endogenous RIP3. Neurobiol Dis. 2014;68C:26–36. doi: 10.1016/j.nbd.2014.04.002. [DOI] [PubMed] [Google Scholar]
- 9.Chen WW, Yu H, Fan HB, Zhang CC, Zhang M, Zhang C, Cheng Y, Kong J, Liu CF, Geng D, Xu X. RIP1 mediates the protection of geldanamycin on neuronal injury induced by oxygen-glucose deprivation combined with zVAD in primary cortical neurons. J Neurochem. 2012;120(1):70–77. doi: 10.1111/j.1471-4159.2011.07526.x. [DOI] [PubMed] [Google Scholar]
- 10.Viringipurampeer IA, Shan X, Gregory-Evans K, Zhang JP, Mohammadi Z, Gregory-Evans CY. Rip3 knockdown rescues photoreceptor cell death in blind pde6c zebrafish. Cell Death Differ. 2014;21(5):665–675. doi: 10.1038/cdd.2013.191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Rosenbaum DM, Degterev A, David J, Rosenbaum PS, Roth S, Grotta JC, Cuny GD, Yuan J, Savitz SI. Necroptosis, a novel form of caspase-independent cell death, contributes to neuronal damage in a retinal ischemia-reperfusion injury model. J Neurosci Res. 2010;88(7):1569–1576. doi: 10.1002/jnr.22314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Dvoriantchikova G, Degterev A, Ivanov D. Retinal ganglion cell (RGC) programmed necrosis contributes to ischemia-reperfusion-induced retinal damage. Exp Eye Res. 2014;123C:1–7. doi: 10.1016/j.exer.2014.04.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Huang JF, Shang L, Zhang MQ, Wang H, Chen D, Tong JB, Huang H, Yan XX, Zeng LP, Xiong K. Differential neuronal expression of receptor interacting protein 3 in rat retina: involvement in ischemic stress response. BMC Neurosci. 2013;14:16. doi: 10.1186/1471-2202-14-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Shang L, Huang J-F, Ding W, Chen S, Xue L-X, Ma R-F, Xiong K. Calpain: a molecule to induce AIF-mediated necroptosis in RGC-5 following elevated hydrostatic pressure. BMC Neurosci. 2014;15(1):63. doi: 10.1186/1471-2202-15-63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Daoud A, Song J, Xiao F, Shang J. B-9-3, a novel beta-carboline derivative exhibits anti-cancer activity via induction of apoptosis and inhibition of cell migration in vitro. Eur J Pharmacol. 2014;724:219–230. doi: 10.1016/j.ejphar.2013.12.038. [DOI] [PubMed] [Google Scholar]
- 16.Choi S, Keys H, Staples RJ, Yuan J, Degterev A, Cuny GD. Optimization of tricyclic Nec-3 necroptosis inhibitors for in vitro liver microsomal stability. Bioorg Med Chem Lett. 2012;22(17):5685–5688. doi: 10.1016/j.bmcl.2012.06.098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Wang K, Li J, Degterev A, Hsu E, Yuan J, Yuan C. Structure-activity relationship analysis of a novel necroptosis inhibitor, Necrostatin-5. Bioorg Med Chem Lett. 2007;17(5):1455–1465. doi: 10.1016/j.bmcl.2006.11.056. [DOI] [PubMed] [Google Scholar]
- 18.Zheng W, Degterev A, Hsu E, Yuan J, Yuan C. Structure-activity relationship study of a novel necroptosis inhibitor, necrostatin-7. Bioorg Med Chem Lett. 2008;18(18):4932–4935. doi: 10.1016/j.bmcl.2008.08.058. [DOI] [PubMed] [Google Scholar]
- 19.Wu Z, Li Y, Cai Y, Yuan J, Yuan C. A novel necroptosis inhibitor-necrostatin-21 and its SAR study. Bioorg Med Chem Lett. 2013;23(17):4903–4906. doi: 10.1016/j.bmcl.2013.06.073. [DOI] [PubMed] [Google Scholar]
- 20.Xing J, Yi-fan F. Advances in studies on saponins in Anemarrhena asphodeloides. Chinese Tradit Herbal Drugs. 2010;41(4):supl12-15. [Google Scholar]
- 21.Li TJ, Qiu Y, Yang PY, Rui YC, Chen WS. Timosaponin B-II improves memory and learning dysfunction induced by cerebral ischemia in rats. Neurosci Lett. 2007;421(2):147–151. doi: 10.1016/j.neulet.2007.04.082. [DOI] [PubMed] [Google Scholar]
- 22.Kim JY, Shin JS, Ryu JH, Kim SY, Cho YW, Choi JH, Lee KT. Anti-inflammatory effect of anemarsaponin B isolated from the rhizomes of Anemarrhena asphodeloides in LPS-induced RAW 264.7 macrophages is mediated by negative regulation of the nuclear factor-kappaB and p38 pathways. Food Chem Toxicol. 2009;47(7):1610–1617. doi: 10.1016/j.fct.2009.04.009. [DOI] [PubMed] [Google Scholar]
- 23.Zhang J, Zhang M, Sugahara K, Sagara Y, Meng Z, Xu S, Kodama H. Effect of steroidal saponins of Anemarrhenae rhizoma on superoxide generation in human neutrophils. Biochem Biophys Res Commun. 1999;259(3):636–639. doi: 10.1006/bbrc.1999.0831. [DOI] [PubMed] [Google Scholar]
- 24.Kaname N, Zhang J, Meng Z, Xu S, Sugahara K, Doi Y, Kodama H. Effect of timosaponin E1 and E2 on superoxide generation induced by various stimuli in human neutrophils and on platelet aggregation in human blood. Clin Chim Acta. 2000;295(1–2):129–140. doi: 10.1016/S0009-8981(00)00196-0. [DOI] [PubMed] [Google Scholar]
- 25.Yun D, Bai-ping M, Yu-wen C, Yu-xian S, Jing-jing Z, Yu-jun S. Protective effects of timosaponin BII on primary neurons against beta amyloid peptide 25–35. Chinese Pharmacol Bull. 2009;02:244–247. [Google Scholar]
- 26.Yun D, Qiuping X, Zhenquan L, Baiping M, Chengqi X, Yang Z. Protective effects of saponin compound from anemarrhena asphodeloides on rat undergone cerebral ischemia/ reperfusion. J Beijing Univ Tradit Chinese Med. 2005;02:33–35. [Google Scholar]
- 27.Huang JF, Shang L, Liu P, Zhang MQ, Chen S, Chen D, Fan CL, Wang H, Xiong K. Timosaponin-BII inhibits the up-regulation of BACE1 induced by ferric chloride in rat retina. BMC Complement Altern Med. 2012;12:189. doi: 10.1186/1472-6882-12-189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Li GY, Li T, Fan B, Zheng YC, Ma TH. The D(1) dopamine receptor agonist, SKF83959, attenuates hydrogen peroxide-induced injury in RGC-5 cells involving the extracellular signal-regulated kinase/p38 pathways. Mol Vis. 2012;18:2882–2895. [PMC free article] [PubMed] [Google Scholar]
- 29.Zhang F, Zhan Q, Gao S, Dong X, Jiang B, Sun L, Tao X, Chen WS. Chemical profile- and pharmacokinetics-based investigation of the synergistic property of platycodonis radix in traditional Chinese medicine formula Shengxian decoction. J Ethnopharmacol. 2014;152(3):497–507. doi: 10.1016/j.jep.2014.01.033. [DOI] [PubMed] [Google Scholar]
- 30.Tsai CH, Yang CW, Wang JY, Tsai YF, Tseng LM, King KL, Chen WS, Chiu JH, Shyr YM. Timosaponin AIII Suppresses Hepatocyte Growth Factor-Induced Invasive Activity through Sustained ERK Activation in Breast Cancer MDA-MB-231 Cells. Evid Based Complement Alternat Med. 2013;2013:421051. doi: 10.1155/2013/421051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Laster SM, Wood JG, Gooding LR. Tumor necrosis factor can induce both apoptic and necrotic forms of cell lysis. J Immunol. 1988;141(8):2629–2634. [PubMed] [Google Scholar]
- 32.Liang X, Chen Y, Zhang L, Jiang F, Wang W, Ye Z, Liu S, Yu C, Shi W. Necroptosis, a novel form of caspase-independent cell death, contributes to renal epithelial cell damage in an ATP-depleted renal ischemia model. Mol Med Rep. 2014;10(2):719–724. doi: 10.3892/mmr.2014.2234. [DOI] [PubMed] [Google Scholar]
- 33.Lin W, Tongyi S. Role of Bax/Bcl-2 family members in green tea polyphenol induced necroptosis of p53-deficient Hep3B cells. Tumour Biol. 2014;35(8):8065–8075. doi: 10.1007/s13277-014-2064-0. [DOI] [PubMed] [Google Scholar]
- 34.Sawai H. Characterization of TNF-induced caspase-independent necroptosis. Leuk Res. 2014;38(6):706–713. doi: 10.1016/j.leukres.2014.02.002. [DOI] [PubMed] [Google Scholar]
- 35.Lu WQ, Qiu Y, Li TJ, Tao X, Sun LN, Chen WS. Timosaponin B-II inhibits pro-inflammatory cytokine induction by lipopolysaccharide in BV2 cells. Arch Pharm Res. 2009;32(9):1301–1308. doi: 10.1007/s12272-009-1916-4. [DOI] [PubMed] [Google Scholar]
- 36.Miura T, Ichiki H, Iwamoto N, Kato M, Kubo M, Sasaki H, Okada M, Ishida T, Seino Y, Tanigawa K. Antidiabetic activity of the rhizoma of Anemarrhena asphodeloides and active components, mangiferin and its glucoside. Biol Pharm Bull. 2001;24(9):1009–1011. doi: 10.1248/bpb.24.1009. [DOI] [PubMed] [Google Scholar]
- 37.Li Y. The protective effects of TSA after cerebral ischemia in rats. Master Degree. Beijing: Beijing University of Chinese Medicine; 2010. [Google Scholar]
- 38.Guo C, Li L, Yang X, Meng Z, Li F, Zhang C, Yang Z. Protective effects of timosaponin B-II on high glucose-induced apoptosis in human umbilical vein endothelial cells. Environ Toxicol Pharmacol. 2014;37(1):37–44. doi: 10.1016/j.etap.2013.11.009. [DOI] [PubMed] [Google Scholar]
- 39.Vandenabeele P, Declercq W, Van Herreweghe F, Vanden Berghe T. The role of the kinases RIP1 and RIP3 in TNF-induced necrosis. Sci Signal. 2010;3(115):re4. doi: 10.1126/scisignal.3115re4. [DOI] [PubMed] [Google Scholar]
- 40.Zhang DW, Zheng M, Zhao J, Li YY, Huang Z, Li Z, Han J. Multiple death pathways in TNF-treated fibroblasts: RIP3- and RIP1-dependent and independent routes. Cell Res. 2011;21(2):368–371. doi: 10.1038/cr.2011.3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.King FW, Fong S, Griffin C, Shoemaker M, Staub R, Zhang YL, Cohen I, Shtivelman E. Timosaponin AIII is preferentially cytotoxic to tumor cells through inhibition of mTOR and induction of ER stress. PLoS One. 2009;4(9):e7283. doi: 10.1371/journal.pone.0007283. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Wang N, Feng Y, Zhu M, Siu FM, Ng KM, Che CM. A novel mechanism of XIAP degradation induced by timosaponin AIII in hepatocellular carcinoma. Biochim Biophys Acta. 2013;1833(12):2890–2899. doi: 10.1016/j.bbamcr.2013.07.018. [DOI] [PubMed] [Google Scholar]
- 43.Sy LK, Yan SC, Lok CN, Man RY, Che CM. Timosaponin A-III induces autophagy preceding mitochondria-mediated apoptosis in HeLa cancer cells. Cancer Res. 2008;68(24):10229–10237. doi: 10.1158/0008-5472.CAN-08-1983. [DOI] [PubMed] [Google Scholar]
- 44.Cabon L, Galan-Malo P, Bouharrour A, Delavallee L, Brunelle-Navas MN, Lorenzo HK, Gross A, Susin SA. BID regulates AIF-mediated caspase-independent necroptosis by promoting BAX activation. Cell Death Differ. 2012;19(2):245–256. doi: 10.1038/cdd.2011.91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Shulga N, Pastorino JG. Mitoneet mediates TNFalpha-induced necroptosis promoted by exposure to fructose and ethanol. J Cell Sci. 2014;127(Pt 4):896–907. doi: 10.1242/jcs.140764. [DOI] [PMC free article] [PubMed] [Google Scholar] [Retracted]
- 46.Sosna J, Voigt S, Mathieu S, Kabelitz D, Trad A, Janssen O, Meyer-Schwesinger C, Schutze S, Adam D. The proteases HtrA2/Omi and UCH-L1 regulate TNF-induced necroptosis. Cell Commun Signal. 2013;11:76. doi: 10.1186/1478-811X-11-76. [DOI] [PMC free article] [PubMed] [Google Scholar]
Pre-publication history
- The pre-publication history for this paper can be accessed here:http://www.biomedcentral.com/1472-6882/14/459/prepub