

BIOLOGY has evidently borrowed the terms “heredity” and “inheritance” from everyday language, in which the meaning of these words is the “transmission” of money or things, rights or duties—or even ideas and knowledge —from one person to another or to some others: the “heirs” or “inheritors.”
The transmission of properties—these may be things owned or peculiar qualities—from parents to their children, or from more or less remote ancestors to their descendants, has been regarded as the essential point in the discussion of heredity, in biology as in jurisprudence. Here we have nothing to do with the latter; as to biology, the students of this science have again and again tried to conceive or “explain” the presumed transmission of general or peculiar characters and qualities “inherited” from parents or more remote ancestors. The view of natural inheritance as realized by an act of transmission, viz., the transmission of the parent's (or ancestor's) personal qualities to the progeny, is the most naive and oldest conception of heredity. We find it clearly developed by Hippocrates, who suggested that the different parts of the body may produce substances which join in the sexual organs, where reproductive matter is formed.
Darwin's hypothesis of “pangenesis” is in this point very consistent with the Hippocratic view, the personal qualities of the parent or the ancestor in question being the heritage.
Also the Lamarckian view as to the heredity of “acquired characters” is in accordance with those old conceptions. The current popular definition of heredity as a certain degree of resemblance between parents and offspring, or, generally speaking, between ancestors and descendants, bears the stamp of the same conceptions, and so do the modern “biometrical” definitions of heredity, e.g., as “the degree of correlation between the abmodality of parent and offspring.” In all these cases we meet with the conception that the personal qualities of any individual organism are the true heritable elements or traits!
This may be characterized as the “transmission-conception” of heredity or as the view of apparent heredity. Only superficial instruction can be gained by working on this basis. Certainly, medical and biological statisticians have in modern times been able to make elaborate statements of great interest for insurance purposes, for the “eugenics-movement” and so on. But no profound insight into the biological problem of heredity can be gained on this basis, for the transmission-conception of heredity represents exactly the reverse of the real facts, just as the famous Stahlian theory of “phlogiston” was an expression diametrically opposite to the chemical reality. The personal qualities of any individual organism do not at all cause the qualities of its offspring; but the qualities of both ancestor and descendant are in quite the same manner determined by the nature of the “sexual substances”—i.e., the gametes—from which they have developed. Personal qualities are then the reactions of the gametes joining to form a zygote; but the nature of the gametes is not determined by the personal qualities of the parents or ancestors in question. This is the modern view of heredity.
The main result of all true analytical experiments in questions concerning genetics is the upsetting of the transmission-conception of heredity, and the two different ways of genetic research: pure line breeding as well as hybridization after Mendel's model, have in that respect led to the same point of view, the “genotype-conception” as we may call the conception of heredity just now sketched.
Here we can not trace the historical evolution of the ideas concerning heredity, not even in the last ten years, but it must be stated as a fact that a very great number of the terms used by the modern biological writers have been created under the auspices of the transmission-conception, and that perhaps the greater number of botanists and zoologists are not yet emancipated from that old conception. Even convinced Mendelians may occasionally be caught using such words as “transmission” and other now obsolete terms.
The science of genetics is in a transition period, becoming an exact science just as the chemistry in the times of Lavoisier, who made the balance an indispensable implement in chemical research.
The “genotype-conception,” as I have called the modern view of heredity, differs not only from the old “transmission-conception” as above mentioned, but it differs also from the related hypothetical views of Galton, Weismann and others, who with more or less effectiveness tried to expel the transmission-idea, having thus the great merit of breaking the ground for the setting in of more unprejudiced inquiries. Galton, in his admirable little paper of 1875, and Weismann, in his long series of fascinating but dialectic publications, have suggested that the elements responsible for inheritance (the elements of Galton's “stirp” or of Weismann's “Keimplasma”) involve the different organs or tissue-groups of the individual developing from the zygote in question. And Weismann has furthermore built up an elaborate hypothesis of heredity, suggesting that discrete particles of the chromosomes are “bearers” of special organizing functions in the mechanism of ontogenesis, a chromatin-particle in the nucleus of a gamete being in some way the representative of an organ or a group of tissues.
These two ideas: that “elements” in the zygote correspond to special organs, and that discrete particles of the chromosomes are “bearers” of special parts of the whole inheritance in question are neither corollaries of, nor premises for, the stirp- or genotype-conception. Those special ideas may have some interest as expressions of the searching mind, but they have no support in experience; the first of them is evidently erroneous, the second a purely speculative morphological view of heredity without any suggestive value.
The genotype-conception of the present day, initiated by Galton and Weismann, but now revised as an expression of the insight won by pure line breeding and Mendel-ism, is in the least possible degree a speculative conception. Of all the Weismannian armory of notions and categories it may use nothing. It is a well-established fact that language is not only our servant, when we wish to express—or even to conceal—our thoughts, but that it may also be our master, overpowering us by means of the notions attached to the current words. This fact is the reason why it is desirable to create a new terminology in all cases where new or revised conceptions are being developed. Old terms are mostly compromised by their application in antiquated or erroneous theories and systems, from which they carry splinters of inadequate ideas, not always harmless to the developing insight.
Therefore I have proposed the terms “gene” and “genotype” and some further terms, as “phenotype” and “biotype,” to be used in the science of genetics. The “gene” is nothing but a very applicable little word, easily combined with others, and hence it may be useful as an expression for the “unit-factors,” “elements” or “allelomorphs” in the gametes, demonstrated by modern Mendelian researches. A “genotype” is the sum total of all the “genes” in a gamete or in a zygote. When a monohybrid is formed by cross fertilization, the “genotype” of this F1-organism is heterozygotic in one single point and the “genotypes” of the two “genodifferent” gametes in question differ in one single point from each other.2
As to the nature of the “genes” it is as yet of no value to propose any hypothesis; but that the notion “gene” covers a reality is evident from Mendelism. The Mendelian workers have the great merit of being prudent in their speculations. In full accordance with this restraint —a quite natural reaction against the morphologicophantastical speculations of the Weismann school—it may be emphatically recommended to use the adjectival term “genotypical” instead of the noun “genotype.” We do not know a “genotype,” but we are able to demonstrate “genotypical” differences or accordances. Used in these derivated ways the term “gene” and “genotype” will prejudice nothing. The very appropriate German term “Reaktionsnorm” used by Woltereck is, as may be seen, nearly synonymous with “genotype,” in so far as the “Reaktionsnorm” is the sum total of the potentialities of the zygotes in question. That these potentialities are partly separable (“segregating” after hybridization) is adequately expressed by the “genotype” as composed of “genes.” The “Reaktionsnorm” emphasizes the diversity and still the unity in the behavior of the individual organism; certainly, the particular organism is a whole, and its multiple varying reactions are determined by its “genotype” interfering with the totality of all incident factors, may it be external or internal. Thence the notion “Reaktionsnorm” is fully compatible with the genotype-conception.
The genotypes can be examined only by the qualities and reactions of the organisms in question. Supposing that some organisms of identical genotypical constitution are developing under different external conditions, then these differences will produce more or less differences as to the personal qualities of the individual organisms. By simple inspection of series of different individuals it will be quite impossible to decide if they have or have not the same genotypical constitution—even if we know them to be homozygotic.3 We may easily find out that the organisms in question resemble each other so much that they belong to the same “type” (in the current sense of this word), or we may in other cases state that they present a disparity so considerable that two or more different “types” may be discerned.
All “types” of organisms, distinguishable by direct inspection or only by finer methods of measuring or description, may be characterized as “phenotypes.” Certainly phenotypes are real things; the appearing (not only apparent) “types” or “sorts” of organisms are again and again the objects for scientific research. All typical phenomena in the organic world are eo ipso phenotypical, and the description of the myriads of phenotypes as to forms, structures, sizes, colors and other characters of the living organisms has been the chief aim of natural history, which was ever a science of essentially morphological-descriptive character.
Morphology, supported by the huge collections of the museums, has of course operated with phenotypes in its speculations concerning phylogenetic questions. The idea of evolution by continuous transitions from one “type” to another must have imposed itself upon zoologists and botanists, because the varying external conditions of life are often4 shifting the phenotypes in very fine gradations; but also—and that is an important point —because there may always be found considerable genotypical differences hidden in apparently homogeneous populations, exhibiting only one single “type” around which the individuals fluctuate. For the descriptive-morphological view the manifestations of the phenotypes in different generations are the main point, and here the transmission-conception immediately announces itself. Hence we may adequately define this conception as a “phenotype-conception” in opposition to the genotype-conception.
As already stated, the genotype-conception has been gained in two ways: pure line breeding and hybridization. The first way leads to an analysis of the existing stocks or populations, the second way may realize an analysis of the genotypical constitution of the individuals. The analysis of populations has its most obvious importance in all such cases, where the phenotypes are quantitatively characterized. Even where individuals with considerable genotypical differences co-exist, the population may—by simple inspection or by statistical appreciation—seem to exhibit only one phenotype, this being usually characterized by the average measure of the individuals (dimensions, weight, intensity of any quality, number of organs and so on). This is due to the fluctuating variability swamping all limits between the different special phenotypes in question (see the diagram).
Populations of self-fertilizing organisms (several cereals and beans, peas and others) have offered the starting point for pure line breeding as a scientific method of research. A pure line may be defined as the descendants from one single homozygotic organism, exclusively propagating by self-fertilization. “Pure line” is a merely genealogical term, indicating nothing as to the qualities of the individuals in question. A “line” ceases to be “pure” when hybridization (or even inter-crossing) disturbs the continuity of self-fertilization.
From a population of homozygotic self-fertilizers there can be started (isolated) as many pure lines as there are fertile individuals—of course very many of such pure lines will be quite identical in genotypical constitution and might in reality belong to one and the same pure line if the genealogy was but sure. The guarantee of the descendence is thus a main point in the principle of pure lines. Identity of genotypical nature is not at all a proof for identical genealogy: the widespread confusion of “resemblance” with “genealogical relation” is the root of much evil—of which the statistics of biometricians have given us some instances.
The isolation of pure lines from plant-populations has been the instrument for gaining the conviction that selection is not able to shift the nature of genotypes. The well-known displacement of the “type” of a population by selection—this displacement proceeding from generation to generation in the direction indicated by the selection—is due to the existence a priori of genotypical differences in such populations (see the diagram). By selection a relatively great number of those organisms, whose genotypical constitution is favorable for the realization of the desired degrees of any character, will be saved for reproduction; hence the result of the selection!
Within pure lines—if no mutation or other disturbances have been at work—or within a population in which there is no genotypical difference as to the character in question, selection will have no hereditary influence. This result has in recent years also been reached by several other experimenters in genetics. Here I also may recall the brilliant experiments of H. S. Jennings with Paramcœium, experiments which have been carried out quite independently of my own researches and which have been of great importance for the propagation and support of the genotype-conception. The bearing of these experiments has been attacked on the ground that the Paramcœiums multiply asexually; but this matter seems to me of no importance in the present case. The experience that pure-line breeding of plants and pure-strain cultures of micro-organisms, in full agreement, demonstrate the non-adequacy of selection as a genotype-shifting factor, is a circumstance of the greatest interest. Also Woltereck's experiments with Daphnias, the important researches of Wolff, and the highly interesting indications of C. O. Jensen as to bacteria may be mentioned here as further supports for this view. Quite recently Pearl has arrived at the same conclusion as to the egg-production by fowls.
The famous Galtonian law of regression and its corollaries elaborated by Pearson pretended to have established the laws of “ancestral influences” in mathematical terms. Now, by the pure-line explanation of the well known action of selection in poly-genotypic populations, these laws of correlation have been put in their right place; such interesting products of mathematical genius may be social statistics in optima forma, but they have nothing at all to do with genetics or general biology! Their premises are inadequate for insight into the nature of heredity.
Ancestral influence! As to heredity, it is a mystical expression for a fiction. The ancestral influences are the “ghosts” in genetics, but generally the belief in ghosts is still powerful. In pure lines no influence of the special ancestry can be traced; all series of progeny keep the genotype unchanged through long generations. A. D. Darbishire's laborious investigations as to the classical object of Mendel's researches, green and yellow peas, may even convince a biometrician that the ancestral influence is zero in “alternative inheritance.” Ancestral influence in heredity is, plainly speaking, a term of the “transmission-conception” and nothing else. The characters of ancestors as well as of descendants are both in quite the same manner reactions of the genotypical constitution of the gametes in question. Particular resemblances between an ancestor and one or more of his descendants depend—so far as heredity is responsible—on corresponding particular identities in the genotypical constitution, and, as we have urged here, perhaps to excess, the genotype is not a function of the personal character of any ancestor.
The genotypic constitution of a gamete or a zygote may be parallelized with a complicated chemico-physical structure. This reacts exclusively in consequence of its realized state, but not in consequence of the history of its creation. So it may be with the genotypical constitution of gametes and zygotes: its history is without influence upon its reactions, which are determined exclusively by its actual nature.
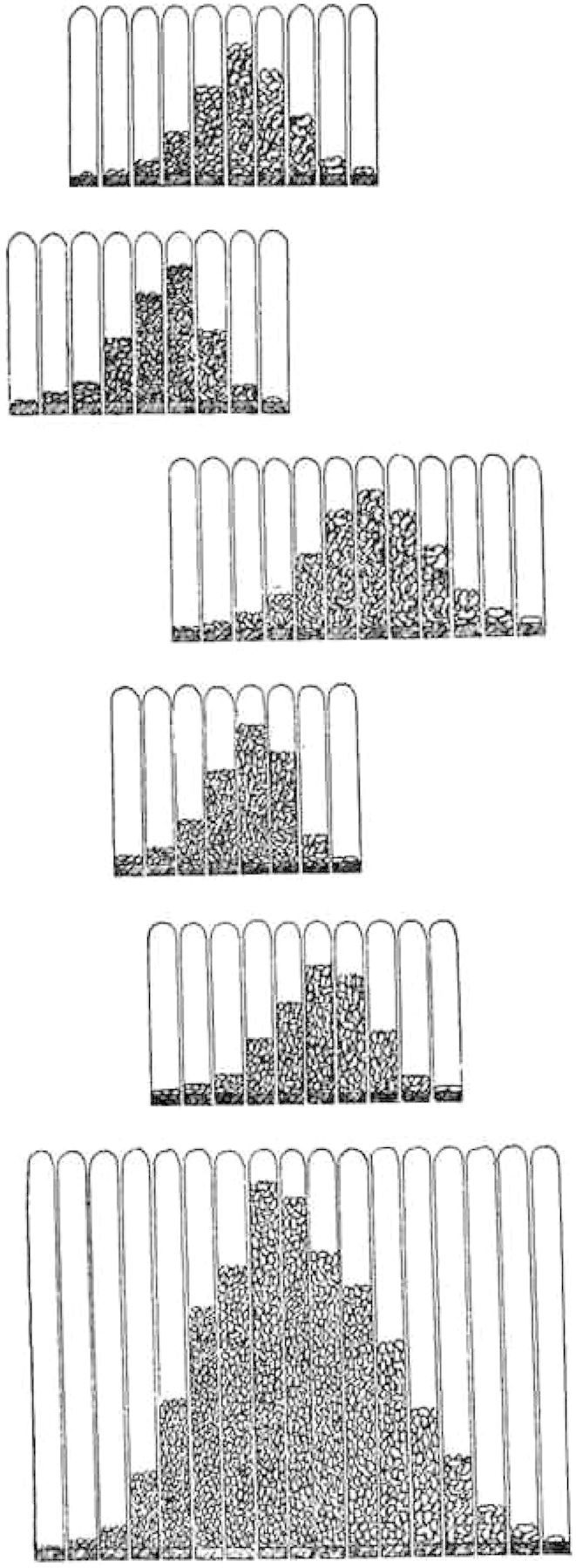
EXPLANATION OF DIAGRAMS DIAGRAMS SHOWING FIVE DIFFERENT PURE LINES OF BEANS AND A “POPULATION” FORMED BY THEIR UNION. In each case the beans enclosed in glass-tubes are marshalled in equidistant classes of length; identical classes are superposed. The pure lines show transgressive fluctuation: it is mostly impossible to state by simple inspection of any individual bean the line to which it belongs.—The fluctuations about the average length (the phenotype) within the pure lines as well as in the mixed population show no characteristic difference.5
The genotype-conception is thus an “ahistoric” view of the reactions of living beings—of course only as far as true heredity is concerned. This view is an analog to the chemical view, as already pointed out; chemical compounds have no compromising ante-act, H2O is always H2O, and reacts always in the same manner, whatsoever may be the “history” of its formation or the earlier states of its elements. I suggest that it is useful to emphasize this “radical” ahistoric genotype-conception of heredity in its strict antagonism to the transmission-or phenotype-view.
As to the evolution of human civilization we meet with true ancestral influences, viz., the tradition (comprising literature, monuments of art, etc., and all forms of teaching). Tradition is playing a very great rôle, but tradition is quite different from heredity. Nevertheless there may often be danger of confusion; and here the use of false analogs is not harmless. So an obscure metaphor is involved in archeologists' reference to Greek temples as “ancestors” of some types of Christian churches, or in their speaking of the descent of violins from more primitive “ancestors.” Certainly, evolution of types of tools, instruments and implements of all kinds is—at least partially—going on by means of selective factors combined with tradition, the latter not only conserving the valuable types but actively stimulating their improvement. But all this has nothing at all to do with the biological notion of heredity. It is of course interesting to see that the idea of “evolution by selection” has won credit in archeology, sociology, etc., but this involves nothing as to genetics, for which “tradition” is irrelevant.
The very “radical” form of the genotype-conception advocated here may be too “theoretical” to be carried through in all its consequences in cases of practical experiments in genetics. In nature and even in the chemical factories the chemical compounds are not always to be had in quite pure state. The history of a preparation may sometimes be traced by accompanying impurities. As to the analogy with the genotypes we touch here the question whether the genotypical constitution of a gamete may not be accompanied by some accessorial or accidental “impurities” from the individual organism in which the gamete was developed.
Here we meet with the cases of “spurious” heredity, e.g., the infections of the gametes or zygotes as may be seen in certain cases of tuberculosis, syphilis, etc. Such and other forms of spurious heredity may have the appearance of “hereditary transmission” or “ancestral influence”; but theoretically they do not interfere at all with the genotype-conception of heredity. In such interesting cases as that detected by Correns, viz., the “heredity” of a special form of albinism by “transmission” through the plasm of the ovum—the sperm not transmitting this character—we may at the first glance be puzzled. Nevertheless, as Correns himself points out, here we have certainly to do with a pathological state of the plasm or the chromatophores in question, and that may perhaps be the reason for the lack of heredity through the sperm which carries no (?) plasm or only a small quantity. The etiology of such abnormalities being as yet quite unknown, it may often be very difficult to distinguish them clearly from “genotypically” determined abnormalities which show the normal form of heredity through both ovum and sperm. The case quoted demands further experience and seems not to be in accord with results of Baur ‘s experiments. At any rate, there may be several difficulties to overcome in the full and consistent application of the genotype-conception, difficulties that may be characterized as perturbations by infection or contamination. And hereby it must be remembered that theoretically, as well as practically, there are no sharp limits between “normal” and “pathological” manifestations of life. “Nature is beautiful, but not correct,” is a Danish saying.
The principle of pure lines or, generally, pure culture, is of importance also for elucidating the celebrated question of the inheritance of “acquired characters.” Mendelism and pure-line researches are here in the most beautiful accordance, both emphasizing the stability of genotypical constitution; the former operating with the constituent unities, the latter with the behavior of the totality of the genotypes in question. The brilliant work of Tower with Leptinotarsa and the highly suggestive injection experiments of MacDougal indicate that changes of the genotypical constitution are produced by steps, discontinuously. And as yet no experiment with genotypically homogeneous cultures has given any evidence for the Lamarckian view, the most extreme “transmission”-conception ever issued. As to bacteria, the important experiments recently made by C. O. Jensen for the purpose of changing their types through adaptation have given not only absolutely negative results, but have demonstrated the fallacy of some positive indications by previous authors. Lamarckism and selectionism are certainly at bottom the same thing: the belief in personal qualities being “transmitted” to the offspring. Observations in impure populations are now their places of resort; nevertheless, it is granted that their history in biology as suggestive ideas has been most glorious.
Apropos, some cases of apparent action of selection may have direct touch with Lamarckian ideas, as, e.g., De Vries's selection of buttercups, recently quoted by Jennings as “the only case that he has found” indicating hereditary action of selection: “Here, after selection the extreme was moved far beyond that before selection.” And Jennings says: “Possibly repetition with thorough analytical experimentation will show that something besides selection has brought about the great change. But at present the case stands sharply against the generalizations from the pure line work.”
Certainly Jennings is in reason, when he, on the ground of his own masterly researches, looks out for “something besides selection.” There are three directions for the inquiry here. First, the strong evidence that the buttercup-population was not at all homogeneous. Secondly, the possibility of intercrossing. I only need to point out the beautiful researches of Shull as to the effect of intercrossing in maize. The heterozygotes were here larger and more productive than the pure strains. The surprises of heterozygotic “constructions” or of new combinations in F2, may perhaps be responsible for the case of De Vries's buttercups; I shall not try to discuss it. But, thirdly, we have an instance pointed out several times by De Vries himself, viz., the combination of selection with nourishment: “la sélection c'est l’alimentation” as it has been said. I suppose that we have here the essential point. The buttercups in culture have been better nourished than before the experiments. Hence, the “best” genotypes having been selected from the population and submitted to “better” nourishment, the result would easily be a moving of the extremes far beyond those before selection. The buttercup-case seems to me to present no difficulties for the genotype-conception.
The practical breeders are a somewhat difficult people to discuss with. Their methods of selection combined with special training and “nurture” in the widest sense of this word are mostly unable to throw any light upon questions of genetics, and yet they only too frequently make hypotheses as to the nature of heredity and variability. Darwin has somewhat exaggerated the scientific value of breeders' testimonies, as if a breeder eo ipso must be an expert in heredity. As to the principle of pure lines it has been occasionally vindicated by German authors, e.g., K. V Rümker, that pure line breeding is a thing old and well known. This is quite true; nearly sixty years ago L. Vilmorin not only emphasized in a lucid manner the importance of pure breeding, but he even tried a little to use his experiences theoretically. But it can not be denied that the principle of pure lines, as a true scientific analytical implement, as an indispensable method of research in heredity—not merely as a questionable and, at any rate, unilateral and insufficient method of practical breeding—is a novelty from recent years. Had this analytical principle been used in the times of Darwin, or had it even been appreciated in due time by the biometric school, certainly the real bearing of selection might long since have been rightly understood also by the practical breeders of pure strains.
The genotypes may then be characterized as something fixed and may be, to a certain degree, parallelized with the most complicated molecules of organic chemistry consisting of “nuclei” with a multitude of “side-chains.” Continuing for a moment such a metaphor, we may even suggest that the genes may be looked upon as analogs of the “radicals” or “side-chains.” All such ideas may as yet be premature; but they are highly favored by the recent researches of Miss Wheldale.
The fixity of a genotypical constitution in question is the conception arrived at by Mendelian and pure line. work. Hence there is a discontinuity between different genotypes. This discontinuity has been energetically contested by several biologists, among whom Woltereck may be pointed out as an important representative. In his very interesting report on experiments with Daphnias, Woltereck indicates, as said above, that selection was as yet ineffective; moreover he describes a case of discontinuous alteration of type (mutation), and his experiments designed to confirm the Lamarckian view have given as yet negative results, even though these may be called “promising,” as he says. So all the evidence of his breeding experiments is in reality quite in favor of our genotype-conception!
But how much depends upon our mental eyesight, what we see. Woltereck confesses openly his belief in continuous evolution and remarks that for a convinced selectionist of the Weismann school the new genotype-conception is a “hard blow.” The aim of his paper in question is to parry off such blows. Of course this parry can not use his own statements just mentioned; as to their obvious but inconvenient accordance with our conception Woltereck might apply the famous words from Harvey's times: “video sed non credo.” Hence the arguments must be taken from other observations, and some very instructive results of cultures under varying conditions have supplied the pièce de reśistance for the discussion. Woltereck is within his right when asserting that we consider different genotypes as having constant differences (like different formulas in chemistry). This is an essential point; but Woltereck, admitting no constancy in the differences, tries to demonstrate that our view must be fallacious.
In a very suggestive manner be presents “phenotype-curves” for several pure strains. These curves are graphical schemes expressing (for the strain in question) the average degree or intensity of any particular character as it manifests itself under different conditions, e.g., the relative length of heads by poor, intermediate and rich feeding, etc. Such “phenotype-curves” may indeed be very useful as records of the behavior of the organisms in question, and they mark certainly a valuable progress in descriptive methods.
The phenotype-curves of the Daphnias in question sometimes show rather constant differences between the pure strains compared; but mostly this is not the case. Especially under extreme conditions, e.g., with poor or even with very rich feeding, some of the curves are confluent. So the differences between the phenotype-curves may vary considerably or may even vanish entirely. These experiences agree with numerous observations of Wesenberg Lund as to the Daphnias in the Danish lakes, and there is no doubt as to their correctness.
But when Woltereck thinks that these facts are inconsistent with the existence of constant differences between the genotypes, he shows himself to have totally misunderstood the question! Of course the phenotypes of the special characters, i.e., the reactions of the genotypical constituents, may under different conditions exhibit all possible forms of transition or transgression —this has nothing at all to do with constancy or inconstancy of genotypical differences.
Every student of genetics ought to know this; some few examples may suffice to enforce it: Temperature has great influence upon the intensity of color in flowers; all shades of intensity from saturated reddish-blue to pure white may be observed with different temperatures in lilac flowers of the “colored” varieties. Such pure white flowering individuals are—as to color—phenotypically not distinguishable from genotypically pure “white” varieties. Nobody will assume that there should be genotypical transitions here! Pure lines of beans may in one year be different in size, e.g., the average of the line A exceeding that of B. In another year B may exceed A, or their average sizes may be practically identical. Differences of soil may produce something similar, and it is well known to breeders that some strains of wheat yield relatively much better than others on rich soil, while the reverse is realized on poorer soils. In four subsequent years two pure lines of barley, both characterized by a considerable degree of disposition to produce vacant spikelets (aborted grains) in the heads, presented the phenotypes here indicated in percentages of such vacancies.
| Pure line L: | 30 | 33 | 27 | 29 |
| Pure line G: | 5 | 45 | 3 | 28 |
The genotype-differences are nevertheless constant; the “Reaktionsnorms” of the organisms in Woltereck's cases, as well as in the examples just cited, are of course eo ipso “constantly different” just as well as the “Reaktionsnorms” of different chemical compounds. And as to chemical analogies it may perhaps be useful to state that different chemical compounds (the structural or constitutional differences of which surely are granted to be discontinuous and constant) may sometimes show “reaction-curves” highly resembling Woltereck's “phenotype-curves.” It is, I suppose, quite sufficient to point out the temperature-curves of solubility for different salts of sodium and other metals. These curves interfere in different ways, cutting each other or partially confluent, in analogy with Woltereck's phenotype-curves.
The essential point in the whole matter is, of course, that a special genotypical constitution always reacts in the same manner under identical conditions—as all chemical or physical structures must do. Differences in genotypical constitution (as well as differences in chemical or physical nature) are not bound to manifest themselves at all—and still less to do so in the same sense—under all conditions. Sometimes even quite special conditions may be required for the realization of possibilities (“Potenzen,” as some German authors are saying), due to a special genotypical nature: This is a well-known fact in physiology as in the fine art of gardening. Baur has long since emphasized the importance of this point for the Mendelian researches.
So the criticisms of Woltereck as to the genotypical discontinuity and constancy are only based upon a regrettable misconception of the genotype-notion. Over and over we find in current literature this confusion of genotypes with phenotypes, and we even have met with the idea, that the Daphnias of a lake may in summer diverge in different races or varieties, but that in winter they converge into one single race! In this statement of Wesenberg Lund, the author regards of course only the phenotypes in a purely descriptive manner. It is evident that Woltereck's view has been influenced by Wesenberg Lund in this matter; but what might be fairly excused in the latter is not allowable for an experimenter pretending to work with cardinal questions of genetics.
Discontinuity and constant differences between the “genes” are the quotidian bread of Mendelism, and here the harmony of Mendelism and pure line work is perfect. We have dealt with some recent criticism of the pure line results; now it is time to look at Mendelism. The astonishing evolution of this mode of research has given an almost interminable stock of special results, and cases that at first might seem incompatible with the Mendelian views have been analyzed more thoroughly on a large scale and have shown themselves quite in accordance with Mendelism. The magnificent book of Bateson gives a full account of this prosperous state of Mendelian research. And it may be evident that Mendelism gives the most striking verification of the essential point in Galton's “stirp-hypothesis”: the inadequacy of the personal quality in heredity. At the same time it overthrows totally the idea of “organs” as being represented by the unities of the “stirp,” pointing out that the personal qualities of the organism in toto are the results of the reactions of the genotypical constitution. The segregation of one sort of “gene” may have influence upon the whole organization. Hence the talk of “genes for any particular character” ought to be omitted, even in cases where no danger of confusion seems to exist. So, as to the classical cases of peas, it is not correct to speak of the gene—or genes—for “yellow” in the cotyledons or for their “wrinkles,”—yellow color and wrinkled shape being only reactions of factors that may have many other effects in the pea-plants. It should be a principle of Mendelian workers to minimize the number of different genes as much as possible.
Here we meet with the questions of correlation and “coupling” of genes. I can not here enter into a discussion as to the notion of “correlation” with its several meanings; in my “Elemente der exakten Erblichkeits lehre” a rather full discussion is to be found. I may only point out here that many cases of presumed correlation may simply be cases of two or more characters (reactions) due to the presence—or even absence—of one single gene. The phenotypically distinct and even diversely localized “characters” convey easily the impression that they are reactions of different genes.
The highly interesting experiences of Correns, Doncaster, Morgan, Spillman and others as to the sex-determining factors, are in some way connected with researches of correlation and “coupling” of genes. The discussion of the ingenious Bateson-Punnett scheme for Abraxas and Morgan's suggestive schemes as to Drosophila may favor the idea of what may be called “ramified” genes. Castle has in his splendid researches as to color-factors in rabbits, etc., initiated a systematic description of the (partially) analyzed genotypes, somewhat resembling the formulas of organic “structural chemistry.” If we suggest an analogy between the radicals of chemistry and the genes, the (partial) genotype-formulas in Castle's manner may be able to demonstrate ramifications of the genes inserted upon the main group of the genotype-constituents. Pausing a moment on this metaphor, it may be suggested that the “branch,” or “branches” of a ramified gene may be more difficult to separate from its “trunk” than the whole gene from the totality of the genotype. I shall here only ask if such views may be of any use as working hypotheses. Their bearing as to the realization of mutations is obvious,—but the purely speculative nature of these suggestions can not as yet warrant a longer discussion here.
It should always be borne in mind that the Mendelian analysis is purely relative. Baur and Shull and even several others have emphasized this fact when discussing the segregations in their experiments, and Shull has clearly pointed out that it may be quite impossible to indicate whether a particular reaction (character) is due to something positive or to the lack of a factor in the genotypical constitution. All that can as yet be determined in this regard by Mendelian analysis is the number of differing points between the two gametes forming a heterozygote. Such differences may be termed “genodifferences.” The well-known facts, that a “character” may be dominant in some hybrids but recessive in others, and that segregation in different cases may be very different, indicate that “characters” are complicated reactions. The famous case of Bateson's fowl-hybrids as to the form of comb may here be quoted as an example: In Walnut comb X Rose comb the latter is recessive, in Single comb X Rose comb it is dominant, and in both cases the segregation gives three dominants: one recessive. Now Bateson has shown that “Walnut” is a compound of Rose- and Pea-comb: Homozygotic Walnut differs from homozygotic Rose only in one point, as does Rose compared with Single. But Walnut-gametes differ from Single-gametes in two points; hence Walnut X Single, with Walnut as dominant, segregates in Walnut, Rose, Pea and Single in the proportions 9:3:3:1. Even with this analysis it is as yet not possible to decide whether Single or Walnut is the form of comb for the realization of which the greater number of positive factors are required. Suggesting—what seems to be the most probable assumption—that Walnut is the most geno-complicated case, Single may even be an expression for a multitude of genes in the fowl-constitution. The relativity of the analysis by segregation must in all such cases be remembered, and it is quite erroneous to think that dominance indicates the positivity of the “unit-factor” in question: So “Horns” are in Wood's cases dominant in male sheep but recessive in female sheep. And as to analogs with chemical reactions it must be kept in mind that a characteristic reaction may be the consequence of lack of any substance as well as dependent upon the presence of any special compound in the solution in question.
The elaborate work of Mendelians of recent years has shown very complicated segregations, and the most specialized segregation is almost the most specialized analysis still known of any “character”? in question. The “units” or “unit-factors” stated in Mendelian work are consequently quite provisory, depending essentially upon the number of genodifferences in the special crossing. Probably it may be discovered that several such “unit-factors” for one character may also be elements for the realization of quite other characters. If this be the truth, then the present state of Mendelism, characterized by the rapidly augmenting number of new “unit-factors” demonstrated in the organization of different biotypes able to hybridize, may be replaced by a period in which many such unit-factors will be identified. At any rate there is no reason to believe that the further Mendelian analysis will augment the number of genes into absurdity. The enormously increasing possibilities of combinations by augmentation of the number of segregable genes are a source of interest also in this connection.
As to cases of hybridization, in which segregation and combination do not suit the Mendelian “laws,” it must at first be stated that some apparent exceptions are probably caused by non-homogeneity of the initial material for experiments. The experiments of Correns, Castle, Miss Saunders, Tschermak and others have shown to excess that phenotypes may seem totally “pure” and nevertheless be heterogeneous (e.g., white flowering stocks or albino mice). Thus constancy as to the phenotype of the progeny is no sure proof for genotypical purity or unity. In discussing alternative inheritance we meet with difficulties of the same nature as in regarding fluctuating variability: the inadequacy of phenotype-description as the starting-point for genetic inquiries.
Secondly, the more or less high vitality of the different combinations of genes in F2 may perturb the Mendelian results, as Baur has illustrated; in other cases the different degree of facility with which the union of special gametes is realized may influence the relative numbers of representatives in the F2-generation, as Correns has demonstrated.
Here we can not discuss the difficulties in a complete carrying through of the Mendelian analysis; Bateson's recent book contains a richness of instances concerning this matter. Only one instance of special importance may be mentioned here, viz., the so-called “blended ‘inheritance” opposed to Mendelian segregation or “alternative inheritance.” In cases of blended inheritance the genes in question might be supposed to “fuse together” by the act of hybridization, or, in accordance with the presence- and absence-view, the gene unilaterally carried to the zygote might here in some manner be “diluted.” In this way, which certainly is very badly compatible with the conception of genes as unit-factors, failing segregation might be explained.
Cases of failing segregation seemed to be abundant in the beginning of the modern Mendelian era; Mendel himself pointed out some typical cases in the species-hybrids of Hieracium. And Correns's indication as to the constant intermediate stature of maize stems seemed to be a crucial case. Now the insight won by breeding experiments as well as by cytological researches concerning the phenomena of apogamy has put the question in a new light. The discoveries of Murbeck, Raunkiær, Ostenfeld, Rosenberg and others have led to quite other explanations as to the constancy of several intermediate hybrid forms. In such cases no segregation is realized, because no gametogenesis is going on—and in such cases there is no reason for supposing any “fusing” or “dilution” of genes. And as to Correns's experiments, this careful author has himself withdrawn the suggestion in question.
But still cases of “blending inheritance” remain. Among these Castle's experiences as to the dimensions of rabbits, especially the length of ears, are the most important and most discussed instances. Castle has in a convincing and suggestive manner demonstrated that the complicated color-characters in rabbits agree with the Mendelian laws. Therefore much stress might be laid upon his indication of cases contrary to these laws.
Crossing short-eared and long-eared races, he gained an F1-generation with almost intermediate ears, and here no. segregation was observed in F2.
But even this case may agree with Mendelian laws. The idea for such interpreting is won—as Lang has clearly pointed out–by means of Nilsson-Ehle's (and East's) experiments, the former concerning the colors of wheat-grains, the latter dealing with the number of “rows”-in the ears of maize. Nilsson-Ehle demonstrated that blending of red and white color in wheat is apparently a fiction: The red color is determined by several different genes, acting in the same sense and augmenting the effect of each other. Hence by segregation and new combinations of these approximately equipotent genes a whole series of gradations in red color will be realized. And these gradations must group themselves symmetrically around the phenotype of the F1 in question. If we have to consider say three genes, A, B and C, we shall for F1 use the formula AaBbCc, indicating the value 3 which is intermediate between aabbcc as zero and AABBCC as 6. Even in case of no fluctuation such a series must present itself as an almost continuous gradation, and it is not difficult to find out that the progeny of every “class” here will breed true, i.e., the average of the progeny's character will be like the “class” of the parent.
Just so it is in the case of East's experiments with maize, as East himself has clearly illustrated. Thus, well-analyzed instances of heredity in plants, concerning both color-factors and meristic factors may be compared with Castle's case in question. Lang in his interesting criticisms points out that certain irregularities in Castle's F2-material give strong evidence for the view that we have no blended inheritance but true segregation here as well as in the cases of Nilsson-Ehle (and, as we may add, in the cases of East). Further analysis may then probably demonstrate in a more direct manner the true nature of the apparent blending in Castle's case; as yet we can only say that this case does not seem incompatible with Mendelian views. It must also be borne in mind that certainly there have been very many genodifferences between the differing races intercrossed in Castle's experiments. Hence these experiments are really operating with highly poly-heterozygotic F1-generations. And how great influence upon dimensions (of ears and other parts of the body) those color-determining genes may have exercised can not be easily determined.
As to beans, it is proved that genes, effective in color-reactions, may also have great influence upon the dimensions and forms. So in my crosses a special factor, which makes yellow color turn into brown and causes violet to be turned into black, has a very marked influence upon the size and form of the beans in question. Here exact data are not necessary; the instance exemplifies the two incident matters of fact, viz., that apparently simple “dimensional” or meristic characters may be determined by several different genes, and that one sort of gene may have influence upon several different reactions.
Then it seems that Mendelian analysis is proceeding in a very prosperous way; but there may be even very narrow limits for this analysis: the entire organization may never be “segregated” into genes! But still there is much to do in carrying through the genotype-conception as far as possible.
As to cytological researches the genotype-conception is as yet rather indifferent. Certainly the process of segregation must be a cell-action intimately connected with division. But all the innumerably detailed results of the refined cytological methods of to-day do not elucidate anything as to segregation. It seems to the unprejudiced observer that the much-discussed cytological phenomena of karyokinesis, synapsis, reduction and so on may be regarded rather as consequences or manifestations of the divisions, repartitions and segregations of genotypical constituents (and all other things in the cell) than as their causes. This view is applicable even in those cases where sex-determination can be diagnosticated cytologically.
In the discussion as to the existence of true graft-hybrids the cytological configurations have of course a high importance as precisely defined characters of cells in such cases where the cytological elements of the two species in question are different. And, as it may be well known, cytological evidence is not at all favorable for the idea of graft-hybrids. But the use of cytological configurations for diagnosis is quite different from the idea that special cytological elements might have importance for the phenomena of heredity.
The question of chromosomes as the presumed “bearers of hereditary qualities” seems to be an idle one. I am not able to see any reason for localizing “the factors of heredity” (i.e., the genotypical constitution) in the nuclei. The organism is in its totality penetrated and stamped by its genotype-constitution. All living parts of the individual are potentially equivalent as to genotype-constitution. In botany there has been no doubt as to this conception, and as to animals, O. Hertwig has for a long time advocated the same view against the views of Weismann and others, who have suggested that ontogenesis is partly determined or at any rate accompanied by a progressive simplification of the “anlagen” (as we say the “genotype-constitution”) in the cells of the growing embryo. The agencies of normally varying ambient conditions and the interactions of specialized parts in the developing individual may exercise their strong influence upon the whole phenotypical state of the resulting particular individual. But these factors will as a rule not change or shift the fundamental genotypical constitution of the biotype in question. Later on we shall touch the problem of such genotypical changes (the mutations) induced by external factors.
Here we have to point out the fact that “living matter”–or, with a more precise definition, those substances or structures the reactions of which we call “manifestations of life,”—is inter alia characterized by the property of autocatalysis. The autocatalysis of living beings must embrace the totality of their genotypical constituents. It seems to me that this autocatalysis as well as the compensative and complemental maintenance of genotypical equilibrium in the organisms, present some of the greatest enigmas of organic life.
The discussion of cytological problems leads us to the question of pure or impure segregation. In the beginning of modern Mendelian researches several instances of presumed impure segregation of genes in gametogenesis were discussed, e.g., as to color factors in animals. But more thorough analytical experiments have in many such cases demonstrated “purity” in the gametes, the characters in question having proved to be more complicated reactions than at first supposed. Recently Morgan has discussed the question in a quite new manner, suggesting—as a working hypothesis—that the segregation might be not of qualitative but of merely quantitative nature. Hence the gametes should as a rule not be pure. Nevertheless, as the author illustrates by means of interesting diagrams, the F2-generation of a monohybrid with normal dominance might be composed of two classes of individuals sharply defined. And the author suggests that this idea might be able to explain “the graded series of forms so often met with in experience and so often ignored or roughly classified by Mendelian workers.”
Here we again touch the question of “blended inheritance.” I suppose that the above-mentioned explanations by Lang and East are more consistent with the real nature of the graded series in question. Now the Mendelian work has not only been able to demonstrate that several cases of segregation apparently impure are pure segregations of complicated nature; but even the “spotted conditions” as to color in animals and plants, emphasized by Morgan as a puzzling case, does not seem to present any real difficulty for Mendelian explanation. Certainly such cases as Shull has pointed out, viz., heterozygotic nature being necessary for “mottling” in some special bean-hybrids, may at first glance favor the idea of “spotted conditions” being due to irregular segregation or to different repartition of color-determining factors in the tissues in question. But a closer examination seems to vindicate the real existence of special “spotting factors.” The very interesting researches of Lock as to the “Inheritance of certain invisible characters in peas” have clearly pointed out a “spotting” factor or a “pattern”-determiner in peas, independent of any color-manifestation. It must be borne in mind that a multitude of characteristic epidermal “patterns” are found in animals and plants, these patterns concerning all epidermal manifestations and often showing a widely fluctuating variability. As to the realization of all such spots it might be suggested that in neighboring parts of the developing epidermal tissue some little difference of ambient conditions may inhibit or even release reactions, the alternation of which produces the spots.
The whole case seems to be somewhat analogous to the merely phenotypical phenomena of alternative variability first pointed out by De Vries, e.g., the alternation of decussated and contorted stems of Dipsacus. Here we touch the highly suggestive idea of “sensible periods” in ontogenesis or histogenesis emphasized with so good experimental arguments by De Vries. Of course there must be a genotypical fundament for the existence of the alternating character in question, e.g., for the particular nature of the surface of the spots (or for the contortion in Dipsacus, etc.); strains without such genotypical fundament will not be spotted (nor produce contorted individuals at all).—These remarks are made only to point out that Morgan may have exaggerated a little his criticisms as to “spotting factors,” but I confess that this question is in need of closer analysis.
Then the problem of pure or impure segregation may still be open; but the tendency in modern genetics goes certainly in the direction of establishing pure segregation as the normal case. If we accept the suggestion of autocatalysis as an essential factor for the propagation of living matter in general, and hence eo ipso, for the growth or multiplication of genotypical constituents, we might in case of impure segregation expect frequently to find “dominants” in the progeny of “recessives”; and the numerical proportions of the dominants and recessives in consecutive generations must be rather irregular. But this is not the case. The recent experiments of Darbishire quoted above demonstrate in a beautiful manner the purity of segregation during subsequent generations in Menders classical object, the pea.
Francis Bacon says; “Human understanding easily supposes a greater degree of order and equality in things than it really finds.” So we may in modern genetics be aware of the relativity and narrowness of our provisorial explanations, remembering Bacon's warning that “many things in nature may be sui genesis and irregular!” Among the irregularities in heredity we may reckon the mutations, observed in nature as well as in more precisely defined conditions of artificial experiments. From the famous observations of De Vries and the indications of several earlier authors, to the modern experimental researches of MacDougal, Standfuss, Tower, Blaringhem and others; all evidences as to mutations point out the discontinuity of the changes in question. Here we need not enter the question; it is sufficient to state that the essential point is the alteration, loss or gain of constituents of the genotype. The splendid experiments of Tower as to Leptinotarsa have in the most evident manner shown that the factors which produce the mutations in this case, viz., the temperature and state of moisture, are able to act in a direct manner upon the genotypical constitution of the gametes; and Tower has noted the occurrence of Mendelian segregation in hybridizing his mutants with the original unaltered biotypes. There may in some cases be certain puzzling irregularities to be explained by future researches, but it is evident that in all such mutations, discontinuity is the characteristic feature in the change of type.
As to populations, the biotypes of which may practically exhibit continuous transitions—like the case of my own populations of beans—the idea might be born that biotypes are evolved from each other by extremely small steps in genotypical change. Hence such mutations must be practically identical with “continuous” evolution. But there is no evidence for this view. Certainly in such populations the “static” transitions between the genotypical differences manifesting themselves in several characters may be called continuous—but such a “continuity of museums,” as it might be called, is not at all identical with genetic continuity. Galton himself has emphasized the capital difference between the notions of continuity in collections and continuity in origin, and as yet the mutations really observed in nature have all shown themselves as considerable, discontinuous saltations. So in my own still unpublished experiments with pure lines. Nutura facit saltus. The chemical analog to such mutations may be the formation of homologous alcohols, acids and so on. The greater mutations may be symbolized by more complicated molecular alterations. But such analogs are of very little value for the understanding of genetic evolution.
The genotype-conception supported by the great stock of experiments as to pure line work, Mendelism and mutations does not consider personal adaptation as a factor of any genetic importance. Phrases as “characters, won by adaptation and having successively been hereditarily fixed,” are without meaning from our point of view. Hence much talk of adaptive characters successively gained seems to us an idle matter. A closer study of desert-organisms and the like may elucidate such things; here the suggestive researches of Lloyd as to stomates desert plants may be pointed out. And as to the old question of “mimicry,” this problem in the famous cases of butterflies has in a most convincing manner been put into Mendelian terms by the observations and experiments of Punnett, de Meijere and others. This stronghold of the united Lamarckism and selectionism has now been conquered for Mendelism, i.e., for the genotype-conception.
The genotype-conception here advocated does not pretend to give a true or full “explanation” of heredity, but may be regarded only as an implement for further critical research, an implement that in its turn may be proved to be insufficient, unilateral and even erroneous—as all working-hypotheses may some time show themselves to be. But as yet it seems to be the most prosperous leading idea in genetics.
Heredity may then be defined as the presence of identical genes in ancestors and descendants, or, as Morgan says in full accordance with this definition; “The word heredity stands for those properties of the germ-cells that find their expression in the developing and developed organism.”
And now it is time to end this communication, too long for its real contents, but too short for the importance and diversity of the great problem of heredity.
In concluding this address I must highly emphasize the eminent merits of Hugo de Vries. His famous book “Die Mutationstheorie,” rich as well in positive indications as in ingenious views, has been the mediator for the new and the old era in genetics. This monumental work is a landmark in the progress of our science. Like the head of Janus it looks at once forward and backward, trying to reconcile—at least partly—the antagonistic ideas of continuity and discontinuity in evolution and heredity; hence a great deal of the charm of De Vries's work. But just these qualities have made the work of De Vries too eclectic for the stringent analytical tendencies of modern genetics—a tendency which has in recent years found a true home in American science.
Footnotes
1 Johannsen W. The Genotype Conception of Heredity. The American Naturalist 1911;45:129-159. Reprinted with permission.
2 They may therefore be characterized as “mono-genodifferent”; this term and the further terms “di-genodifferent” and so on, may or may not be of any use.
3 Here we are not concerned with the question of variable dominance, etc.
4 Not always, as Bateson has the merit of having emphasized.
5 It can not be detected by inspection that the five upper diagrams represent phenotypes which are genotypically homogeneous, while the nethermost diagram —the sum of the others—indicates a phenotype masking five others. That these five phenotypes all are genotypically different is known a priori, in this special case, but it could not be discerned by simple inspection.—In the population genotypical differences are combined with merely individual fluctuations; within the single pure line only such fluctuations are seen. Hence, while selection within a pure line will have no hereditary influence, it is evident that any selection in the population must shift or move the “type” of the progeny in the direction of selection.