

Abstract
Systemic lupus erythematosus (SLE) is a complex autoimmune disease characterized by the loss of tolerance to self-nuclear antigens. The symptoms of SLE, progression of pathology and the array of autoantibodies present in the serum differ significantly from patient to patient, which calls for a personalized approach to treatment. SLE is polygenic and strongly influenced by gender, ethnicity, and environmental factors. Data from genome-wide association studies suggests that polymorphisms in as many as 100 genes contribute to SLE susceptibility. Recent research has focused on genes associated with Toll-like receptors (TLRs), type I interferons, immune regulation pathways, and immune-complex clearance. TLR7 and TLR9 have been extensively studied using lupus-prone mouse models. In multiple systems overexpression of TLR7 drives disease progression but interestingly, a loss of TLR9 results in an almost identical phenotype. While TLR7 overexpression has been linked to human SLE, the possible role of TLR9 in human disease remains elusive. In the present review, we focus on TLR polymorphisms and TLR expression in SLE patients and discuss their potential as biomarkers for individualized treatment.
Keywords: systemic lupus erythematosus, TLR7, TLR9, type I interferons, pharmacogenomics
INTRODUCTION
Systemic lupus erythematosus (SLE) is a chronic autoimmune disease with a variety of clinical manifestations that differ from patient to patient. The heterogeneity of the symptoms represents a challenge for both diagnosis and treatment. While the diagnostic criteria for SLE have evolved over the past decades (reviewed in Yu et al., 2014), the treatment has remained largely symptomatic using non-specific conventional therapies which include non-steroidal anti-inflammatory drugs (NSAIDs), glucocorticoids, hydroxychloroquine (HCQ), and immunosuppressants (Murphy et al., 2013). In addition to their modest therapeutic effect, these drugs have severe side effects leading to substantial morbidity and mortality, resulting in a substantial economic burden to many societies (Lau and Mak, 2009). In this short review, we briefly discuss novel therapeutic agents and emerging immunological targets. We will focus on the burgeoning data on Toll-like receptor (TLR) 7 and TLR9 in SLE and on the factors that could influence their expression. We propose altered expression of TLR7/9 as a biomarker for identification of a subset of SLE patients that might benefit from a targeted therapeutic approach.
RECENT DEVELOPMENTS IN B CELL THERAPEUTICS
Recently, therapeutic avenues in SLE have pursued biologic drugs which either deplete B cells or reduce their activity. Belimumab (Benlysta®) is the only biological drug and the first therapeutic in 50 years to be approved by the Federal Drug Administration (FDA) for the treatment of lupus. Belimumab is a human monoclonal antibody (mAb) specific for B lymphocyte stimulator (BLyS) protein/B cell activating factor (BAFF; Hahn, 2011). Multicenter randomized controlled trials demonstrated a significant reduction in the Safety of Estrogen in Lupus Erythematosus National Assessment–SLE Disease Activity Index (SELENA–SLEDAI) score and risk of severe flares (Furie et al., 2011; Navarra et al., 2011). Recent results also suggest that belimumab may improve renal disease; however, further studies are needed to demonstrate a benefit in patients with severe active renal nephritis (Dooley et al., 2013). Based on the success of belimumab, other B cell targeting therapies are being assessed in clinical trials, including rituximab (anti-CD20), epratuzumab (anti-CD22), blisibimod and tabalumab (anti-BLyS), and atacicept [anti-BLyS/APRIL (A proliferation-inducing ligand); Kamal, 2014]. So far, the outcomes of B cell depletion with rituximab in clinical trials have been disappointing, partially due to poor trial design, problems with outcome measures and lack of long-term follow-up (Looney et al., 2010; Murphy et al., 2013; Kamal, 2014). In a randomized, double-blind, placebo-controlled Phase III trial rituximab successfully depleted B cells in lupus nephritis patients and decreased anti-dsDNA antibody levels, but failed to improve renal disease (Rovin et al., 2012). Since human prospective studies and mouse models have suggested a multi-step hypothesis to the development of SLE, it is perhaps unsurprising that reducing B cells has only moderate effects in end organ disease such as nephritis (Arbuckle et al., 2003; Fairhurst et al., 2006; Kanta and Mohan, 2009). Based on this hypothesis, the B cells have a critical role in autoantibody production due to initial loss of tolerance. However, an additional amplification step within the innate immune system is needed for the development of end organ disease.
Accordingly, new therapeutic approaches which focus on targeting innate immune cells are being developed (Figure 1).
FIGURE 1.
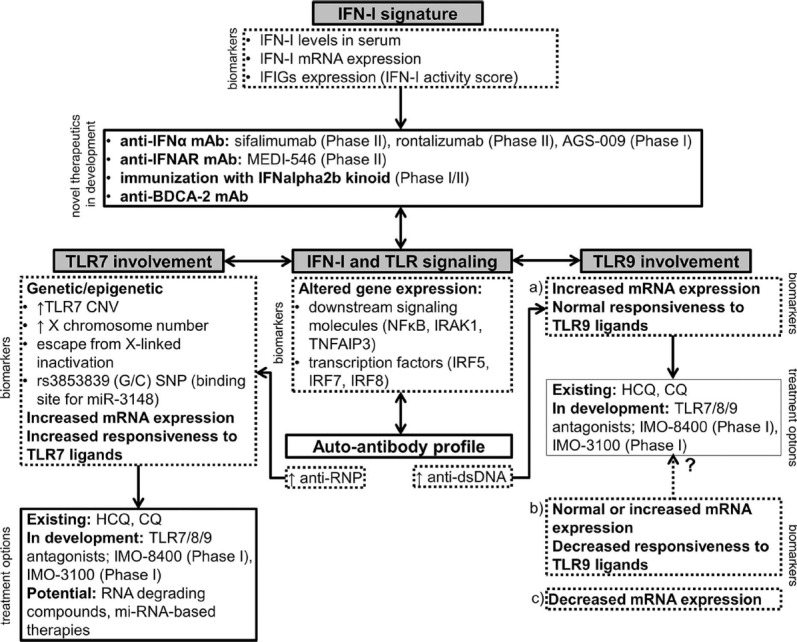
A schematic representation of novel therapeutic approaches in SLE involving the IFN-I signature, TLR7 and TLR9. The choice of the appropriate treatment option should ideally be based on the expression of relevant biomarkers in an individual patient. Drugs that are currently in clinical development are listed with the corresponding phase of the clinical trial (Clinicaltrials.gov, 2014). IFIGs, interferon-inducible genes; HCQ, hydroxychloroquine; CQ, chloroquine.
TLR7 AS A TARGET FOR SLE THERAPY
IMMUNOLOGICAL EVIDENCE FOR TLRs IN SLE
The type I-interferon (IFN-I) gene signature is one of the main immunological characteristics which was identified just over a decade ago by multiple groups (Baechler et al., 2003; Bennett et al., 2003; Crow, 2014). The increase in IFN-I-related genes was identified in the peripheral blood monocytes (PBMCs) from patients with SLE using gene expression profiling and has been identified in the majority of pediatric SLE patients and in the majority of adult SLE patients with active disease (Baechler et al., 2003; Bennett et al., 2003). Interestingly, the gene expression signature does not correlate with either elevated IFN-I levels in SLE serum, which is detectable only in a fraction of patients, or with elevated IFN-I mRNA (Blanco et al., 2001; Baechler et al., 2003). The reason for this discrepancy might be the low sensitivity of enzyme-linked immunosorbent assays (ELISAs) to detect serum IFN-I and the migration of IFN-I-producing cells to the tissues. However, there is also the possibility that the IFN-I signature is caused by another stimulus aside from IFN-I itself (Baechler et al., 2003; Bennett et al., 2003).
To overcome the limitations of serum IFN-I measurements “IFN-I activity score” assays were developed by several groups (Hua et al., 2006; Burgi Mde et al., 2012). These assays use serum or plasma of patients and the activity score strongly correlates with the titer of antinuclear autoantibodies (ANAs; Kirou et al., 2005; Hua et al., 2006; Niewold et al., 2007). High IFN-I activity score and the tendency to develop anti-ribonucleoprotein (RNP) and anti-dsDNA antibodies appear to be independent risk factors for SLE, since healthy first-degree relatives of SLE patients frequently display elevated IFN-I activity but no detectable ANAs (Niewold et al., 2007). The crucial connection between both traits is nucleic acid-sensing by endosomal TLRs, which evolved as sensors of foreign RNA and DNA (Blasius and Beutler, 2010; Kawai and Akira, 2010). In SLE, where self-nucleic acids are associated with autoantibodies in immune complexes (ICs), endosomal TLRs might become aberrantly activated in the absence of foreign molecules (reviewed in Celhar et al., 2012). Multiple in vitro studies in both mouse and human cells proved that RNA/DNA-containing ICs activate TLR9 and TLR7 through B cell receptor (BCR)-mediated internalization in B cells (Leadbetter et al., 2002; Lau et al., 2005) and through Fc gamma receptor (FcγR)-mediated internalization in dendritic cells (DCs; Boule et al., 2004), plasmacytoid DCs (pDCs; Bave et al., 2003; Barrat et al., 2005; Means et al., 2005; Vollmer et al., 2005; Lovgren et al., 2006), macrophages (Henault et al., 2012), and neutrophils (Garcia-Romo et al., 2011). Upon such activation, pDCs produce IFNα (Bave et al., 2003; Barrat et al., 2005; Means et al., 2005; Vollmer et al., 2005; Lovgren et al., 2006), conventional DCs produce cytokines (Boule et al., 2004) and neutrophils release neutrophil extracellular traps (NETs; Garcia-Romo et al., 2011). These findings highlight the central role of TLR7 and TLR9 in the induction and modulation of immune responses in SLE, and their association with the IFN-I signature (recently reviewed in Kono et al., 2013; Shrivastav and Niewold, 2013; Crow, 2014). Genome-wide association studies (GWAS) have provided supporting evidence by identifying SLE susceptibility variants in both TLR and IFN-I pathways (Rullo and Tsao, 2013).
GENETIC EVIDENCE: POLYMORPHISMS IN TLR7-ASSOCIATED PATHWAYS
Overexpression of TLR7 causes severe lupus in multiple mouse models of SLE (reviewed in Celhar et al., 2012). Male BXSB lupus mice develop severe disease due to a translocation of a segment near the pseudoautosomal region of the X chromosome onto the Y chromosome, identified as the y-linked autoimmune accelerating (yaa) locus (Pisitkun et al., 2006; Subramanian et al., 2006). Among the duplicated genes, TLR7 was identified as the major gene responsible for the development of severe disease (Deane et al., 2007; Fairhurst et al., 2008; Santiago-Raber et al., 2008). Genetically modified mouse models have proven to be an indispensable tool for the study of single gene modifications. However, correlations with the highly genetically variable human population are rarely straightforward. This is particularly true for polygenic disorders such as SLE, where multiple genes are associated with disease susceptibility (Rullo and Tsao, 2013; Armstrong et al., 2014). Genomic data suggests that common copy number variations (CNVs) similar to yaa are very rare in human SLE (Conrad et al., 2010; Park et al., 2010; Shen et al., 2010). Nevertheless, they may be an important risk factor childhood-onset SLE, as recently shown in the Mexican population (Garcia-Ortiz et al., 2010). Male patients with more than one copy and female patients with greater than two copies of TLR7 had a higher disease susceptibility and the TLR7 copy number correlated with TLR7 mRNA expression levels (Garcia-Ortiz et al., 2010). In addition to altered gene copy number, the gene dosage of TLR7 can be altered in individuals affected by aneuploidy. Indeed, SLE is more common in men affected by conditions with additional X chromosomes, such as Klinefelter’s syndrome (47,XXY; Ortiz-Neu and LeRoy, 1969; Dillon et al., 2012).
Overall, however, it is unlikely that a single gene, in this case TLR7, would have such a large impact. Consistent with this theory, recent investigations have identified polymorphisms in genes that are shared among TLR and IFN-I signaling pathways, including the transcription factors interferon regulatory factor (IRF) 5, IRF7, and IRF8 and components of the downstream NFκB pathway, such as interleukin-1 receptor-associated kinase 1 (IRAK1) and tumor necrosis factor, alpha-induced protein 3 (TNFAIP3; Rullo and Tsao, 2013). Therefore, polymorphisms in several genes could ultimately together lead to an excessive response to RNA sensing by TLR7. Additionally, variants in regulatory single nucleotide polymorphisms (SNPs) which bind microRNA (miRNA), X-chromosome linked modifications and induction of TLR7 expression by various infectious agents may be involved in subgroups of SLE patients.
X-CHROMOSOME LINKED ALTERATIONS OF TLR7 EXPRESSION
Recent findings suggest that approximately 15% of genes on the X chromosome escape X-linked inactivation and are thus bi-allelicially expressed, ultimately increasing gene dosage (reviewed in Berletch et al., 2011). X-linked genes are particularly interesting in dissecting out causes of conditions that predominately affect women, including autoimmune diseases and some viral infections such as HIV and herpes simplex virus (HSV; Fish, 2008). Several studies have attributed these sexual dimorphisms to differential RNA-sensing by TLR7, since female immune cells produce higher levels of IFNα upon stimulation with TLR7 ligands or virus-derived RNA compared to male counterparts (Berghofer et al., 2006; Meier et al., 2009; Torcia et al., 2012). The underling mechanism for this sex bias is currently not clear. Berghofer et al. (2006) found equivalent TLR7 mRNA expression in pDCs and B cells from healthy males and females with no evidence of escaping X-linked inactivation or involvement of estrogen receptor signaling. However, these findings do not exclude escape from X-linked inactivation in SLE patients.
EPIGENETIC FACTORS AND microRNA BINDING
It has now been established that miRNAs can act to fine tune TLR signaling by targeting expression itself or by modulating adaptor molecules, downstream regulators and cytokines (reviewed in O’Neill et al., 2011). Variants in the 3′ untranslated region (UTR) regions may alter miRNAs binding and ultimately TLR7 expression and/or responsiveness. A large multi-centered and multi-ethnic study identified such SNP in the 3′ UTR region of TLR7 (rs3853839 (G/C)) as a risk factor for SLE (Shen et al., 2010; Deng et al., 2013). The G-allele carriers have increased TLR7 transcripts and are more likely to have anti-RNA associated autoantibodies than C-allele carriers (Shen et al., 2010). The non-risk C allele bears a binding site of microRNA-3148 (miR-3148), which confers faster degradation of the transcript and thus lower levels of TLR7 gene product (Deng et al., 2013).
INDUCTION OF TLR7 EXPRESSION
Several studies have reported the induction of TLR7 transcription in immune cells (B cells, eosinophils, monocytes, macrophages, pDCs) following stimulation with various inflammatory agents, including bacteria (Zarember and Godowski, 2002; Bourke et al., 2003; Miettinen et al., 2008), viruses (Miettinen et al., 2001) CpG (Hornung et al., 2002; Bourke et al., 2003), IFNα (Miettinen et al., 2001; Sirén et al., 2005), and IFNγ (Miettinen et al., 2001; Nagase et al., 2003). This increase in TLR7 during infection may be part of a positive feedback mechanism to increase IFNα, which is essential for a fast and robust anti-microbial response (Rönnblom et al., 2006). It is possible that this pathway is deregulated in SLE patients and explains why many features of SLE resemble a chronic viral infection in the absence of a detectable virus (Crow, 2014). Indeed, upregulated TLR7 and TLR9 mRNA expression have been reported in PBMCs from SLE patients and levels correlate with the expression of IFNα (Komatsuda et al., 2008; Lyn-Cook et al., 2014). Upregulation of TLR7, but not other TLRs, has also been observed when healthy neutrophils were cultured with sera from SLE patients with active disease (Garcia-Romo et al., 2011). This may be due to IFNα present in patient’s sera, since pre-treatment with purified IFNα resulted in a similar response of increased TLR7. Furthermore, IFNα increased susceptibility to anti-RNP antibody-induced NETosis (Garcia-Romo et al., 2011). Consistent with these findings, monocyte-derived macrophages produce IFNα upon TLR7/8 ligand stimulation only after pre-treatment with IFNα and subsequent induction of TLR7 (Sirén et al., 2005). TLR7 may also be induced by serum-derived ICs containing TLR7 ligands. A recent study by Chauhan et al. (2013) showed that TLR7 was preferentially increased in SLE patients with antibodies against RNA-associated antigens, while TLR9 induction correlated with anti-dsDNA antibody titers. Moreover, flu and synthetic TLR7 ligands have the capacity to induce early IFN-inducible genes in pDCs independently of IFN-α production (Di Domizio et al., 2009). Overall, accumulating data suggest that persistently higher levels of TLR7 may lead to an acquired responsiveness in cells which are normally unresponsive to TLR7 ligands. This is supported by the observation that pDCs are responsible for the majority IFNα produced by healthy PBMCs, but account only for 57% of IFNα produced by PBMCs from SLE patients (Blanco et al., 2001).
TLR9 AS A TARGET FOR THERAPY FOR SLE
While TLR7 hyper-responsiveness in human SLE is consistent with the TLR7-associated nephritis shown by mouse models, the role of TLR9 remains controversial. Multiple mouse studies have shown the importance of TLR9 expression in B cells for the generation of anti-dsDNA, anti-chromatin, and anti-nucleosome autoantibodies (Christensen et al., 2005, 2006; Lartigue et al., 2006; Yu et al., 2006; Nickerson et al., 2010). However, the deletion of TLR9 in these lupus-prone models did not lead to amelioration, but rather to exacerbation of disease, suggesting a protective/regulatory role of TLR9 in cells other than B cells. Nonetheless, the manifestation of the disease in TLR9-deficient mice was dependent on TLR7 expression (Nickerson et al., 2010).
Increases in TLR9 expression have been shown in PBMCs from SLE patients and levels usually correlate with IFNα expression and anti-dsDNA antibodies (Komatsuda et al., 2008; Mu et al., 2012; Chauhan et al., 2013; Lyn-Cook et al., 2014). Additionally, B cells and monocytes from patients with active disease express higher TLR9 levels compared to patients with inactive disease (Papadimitraki et al., 2006; Nakano et al., 2008). An increase in the frequency of TLR9-expressing B cells, but not monocytes, correlated with anti-dsDNA antibodies (Papadimitraki et al., 2006). These results suggest that, similarly to murine studies, increased TLR9 expression in B cells may be involved in autoantibody production, especially anti-dsDNA
However, paradoxically, the increased expression of TLR9 does not lead to increased responsiveness to TLR9 ligands. Despite an increase in TLR9 on B cells and BDCA3+ DCs from patients with severe disease, cells were less activated and hyporesponsive to TLR9 stimulation (Zorro et al., 2009). Similarly, despite an increased TLR9 expression in SLE PBMCs, TLR9-induced IFN-α production was markedly reduced (Kwok et al., 2008). This desensitization has been directly attributed to HCQ treatment (Kwok et al., 2008; Sacre et al., 2012). However, in patients not taking HCQ, alterations in downstream TLR9-signaling or induction of negative regulators may be involved (Kwok et al., 2008; Zorro et al., 2009).
Limited data is available on the effect of decreased TLR9 expression in human SLE. A recent study conducted in a cohort of Danish SLE patients found significantly decreased TLR9 mRNA expression in lupus PBMCs, which did not correlate with dsDNA antibody titers (Laska et al., 2014). It would be interesting to further identify the main cell type within the PBMCs responsible for the downregulation and assess the responsiveness to TLR9 ligands in this cohort.
Overall, further research is required to establish the role of TLR9 in mediating the progression of benign autoimmunity to severe disease.
TLR9 POLYMORPHISMS
Several TLR9 polymorphisms have been associated with SLE, particularly in patients of Asian or European ancestry (Tao et al., 2007; Xu et al., 2009; Lu et al., 2011; dos Santos et al., 2012; Huang et al., 2012; Piotrowski et al., 2013; Laska et al., 2014). However, these results were not confirmed by other studies or by a recent meta-analysis (Hur et al., 2005; Ng et al., 2005; De Jager et al., 2006; Yang et al., 2012). Similarly, as mentioned earlier for TLR7, polymorphisms in a single gene are not very likely to recapitulate a polygenic disease like SLE. Additional genetic variants in TLR9-downstream signaling pathways, regulators of TLR signaling and epigenetic modifications should also be considered and need further examination.
DEVELOPMENT OF NOVEL THERAPEUTICS FOR SLE
NOVEL THERAPIES DIRECTED TOWARD IFNα
IFNα signaling may be suppressed by several strategies: direct neutralization by an anti-IFNα mAb, suppression of the IFN-I signature using an anti-IFNα receptor (IFNAR) antibody, or immunization to achieve endogenous anti-IFNα production (reviewed in Crow, 2014). Three anti-IFN-α mAbs, sifalimumab, AGS-009, and rontalizumab, have achieved safety and dose-dependent reduction of IFN-I signature in Phase I SLE trials (Yao et al., 2009; Merrill et al., 2011; Petri et al., 2013; Stohl, 2013). Furthermore, a Phase II trial has shown that rontalizumab decreased the incidence of flares and reduced prednisone dosage in a subgroup of patients who had a low IFN-I signature at baseline (Stohl, 2013).
Targeting the IFNAR with MEDI-546, a fully human mAb against huIFNAR, achieved success in Phase I trials for systemic sclerosis (SSc; Wang et al., 2013). Pharmacogenomics and translational simulations were used to build a model to predict responses to MEDI-546 in SLE patients and facilitated the progression of this candidate molecule to a Phase II trial (Wang et al., 2013).
Immunization with human (hu) IFNalpha2b kinoid (IFN-K) generates endogenous polyclonal antibodies directed against all 13 subtypes of huIFNα (Mathian et al., 2011). The results from the first clinical study show that IFN-K is well tolerated and significantly reduces the IFN-I signature in IFN signature-positive patients (Lauwerys et al., 2013).
An additional approach is the blockage of BDCA-2 expressed on the surface of pDCs. Anti-BDCA-2 mAb successfully inhibited IFN-α production from healthy donors and SLE patients (Dzionek et al., 2001; Blomberg et al., 2003). A patent application has recently been submitted for an anti-BDCA-2 antibody in the context of preventing or treating pathological conditions involving pDC activation (De and Fournier, 2012).
TLR INHIBITION IN SLE
Several compounds that bind endosomal TLR7 and/or TLR9 activation have been developed with the goal to inhibit IFNα production and activation of autoimmune B cells (Barrat and Coffman, 2008; Kandimalla et al., 2013). This approach successfully ameliorated disease in lupus-prone mice but it needs further clinical evaluation (Barrat et al., 2007). IMO-3100, a TLR7/9 antagonist, and IMO-8400, a TLR7/8/9 antagonist, suppressed inflammation in a mouse model of psoriasis (Suarez-Farinas et al., 2013). Both compounds were well tolerated by healthy adults and are now undergoing Phase 2 trials in psoriasis patients (Clinicaltrials.gov, 2014). It is important to note that the therapeutic value of the standard SLE care drugs HCQ and chloroquine have been attributed to indirect inhibition of TLR7 and TLR9, through their inhibitory action on endosomal acidification which is necessary for TLR7/9 activation (Sacre et al., 2012).
Finally, miRNA-based therapies and therapeutics design to degrade RNA may also represent potential treatment options in specific groups of SLE patients (Deng et al., 2013; Sun et al., 2013).
CONCLUSION
Advances in the understanding of the innate immune system have revolutionized the development of novel SLE therapies. Owing to the heterogeneous nature of the clinical manifestations, much thought is being given to a stratification of lupus patients based on clinical symptoms and immunological dysfunctions, targeting disease development in a personalized medicine approach. This is particularly important for the design of clinical trials, where the correct recruitment and sub-grouping of patients may directly impact the outcome measures. Pharmacogenomic and pharmacogenetic studies will ultimately be essential to identify patients that will most likely benefit from a specific treatment.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
REFERENCES
- Arbuckle M. R., Mcclain M. T., Rubertone M. V., Scofield R. H., Dennis G. J., James J. A., et al. (2003). Development of autoantibodies before the clinical onset of systemic lupus erythematosus. N. Engl. J. Med. 349, 1526–1533. 10.1056/NEJMoa021933 [DOI] [PubMed] [Google Scholar]
- Armstrong D. L., Zidovetzki R., Alarcon-Riquelme M. E., Tsao B. P., Criswell L. A., Kimberly R. P., et al. (2014). GWAS identifies novel SLE susceptibility genes and explains the association of the HLA region. Genes Immun. 15, 347–354. 10.1038/gene.2014.23 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Baechler E. C., Batliwalla F. M., Karypis G., Gaffney P. M., Ortmann W. A., Espe K. J., et al. (2003). Interferon-inducible gene expression signature in peripheral blood cells of patients with severe lupus. Proc. Natl. Acad. Sci. U.S.A. 100, 2610–2615. 10.1073/pnas.0337679100 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barrat F. J., Coffman R. L. (2008). Development of TLR inhibitors for the treatment of autoimmune diseases. Immunol. Rev. 223, 271–283. 10.1111/j.1600-065X.2008.00630.x [DOI] [PubMed] [Google Scholar]
- Barrat F. J., Meeker T., Chan J. H., Guiducci C., Coffman R. L. (2007). Treatment of lupus-prone mice with a dual inhibitor of TLR7 and TLR9 leads to reduction of autoantibody production and amelioration of disease symptoms. Eur. J. Immunol. 37, 3582–3586. 10.1002/eji.200737815 [DOI] [PubMed] [Google Scholar]
- Barrat F. J., Meeker T., Gregorio J., Chan J. H., Uematsu S., Akira S., et al. (2005). Nucleic acids of mammalian origin can act as endogenous ligands for Toll-like receptors and may promote systemic lupus erythematosus. J. Exp. Med. 202, 1131–1139. 10.1084/jem.20050914 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bave U., Magnusson M., Eloranta M. L., Perers A., Alm G. V., Ronnblom L. (2003). Fc gamma RIIa is expressed on natural IFN-alpha-producing cells (plasmacytoid dendritic cells) and is required for the IFN-alpha production induced by apoptotic cells combined with lupus IgG. J. Immunol. 171, 3296–3302. 10.4049/jimmunol.171.6.3296 [DOI] [PubMed] [Google Scholar]
- Bennett L., Palucka A. K., Arce E., Cantrell V., Borvak J., Banchereau J., et al. (2003). Interferon and granulopoiesis signatures in systemic lupus erythematosus blood. J. Exp. Med. 197, 711–723. 10.1084/jem.20021553 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berghofer B., Frommer T., Haley G., Fink L., Bein G., Hackstein H. (2006). TLR7 ligands induce higher IFN-alpha production in females. J. Immunol. 177, 2088–2096. 10.4049/jimmunol.177.4.2088 [DOI] [PubMed] [Google Scholar]
- Berletch J. B., Yang F., Xu J., Carrel L., Disteche C. M. (2011). Genes that escape from X inactivation. Hum. Genet. 130, 237–245. 10.1007/s00439-011-1011-z [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blanco P., Palucka A. K., Gill M., Pascual V., Banchereau J. (2001). Induction of dendritic cell differentiation by IFN-alpha in systemic lupus erythematosus. Science 294, 1540–1543. 10.1126/science.1064890 [DOI] [PubMed] [Google Scholar]
- Blasius A. L., Beutler B. (2010). Intracellular Toll-like receptors. Immunity 32, 305–315. 10.1016/j.immuni.2010.03.012 [DOI] [PubMed] [Google Scholar]
- Blomberg S., Eloranta M. L., Magnusson M., Alm G. V., Ronnblom L. (2003). Expression of the markers BDCA-2 and BDCA-4 and production of interferon-alpha by plasmacytoid dendritic cells in systemic lupus erythematosus. Arthritis Rheum. 48, 2524–2532. 10.1002/art.11225 [DOI] [PubMed] [Google Scholar]
- Boule M. W., Broughton C., Mackay F., Akira S., Marshak-Rothstein A., Rifkin I. R. (2004). Toll-like receptor 9-dependent and -independent dendritic cell activation by chromatin-immunoglobulin G complexes. J. Exp. Med. 199, 1631–1640. 10.1084/jem.20031942 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bourke E., Bosisio D., Golay J., Polentarutti N., Mantovani A. (2003). The Toll-like receptor repertoire of human B lymphocytes: inducible and selective expression of TLR9 and TLR10 in normal and transformed cells. Blood 102, 956–963. [DOI] [PubMed] [Google Scholar]
- Burgi Mde L., Prieto C., Etcheverrigaray M., Kratje R., Oggero M., Bollati-Fogolin M. (2012). WISH cell line: from the antiviral system to a novel reporter gene assay to test the potency of human IFN-alpha and IFN-beta. J. Immunol. Methods 381, 70–74. 10.1016/j.jim.2012.04.010 [DOI] [PubMed] [Google Scholar]
- Celhar T., Magalhaes R., Fairhurst A. M. (2012). TLR7 and TLR9 in SLE: when sensing self goes wrong. Immunol. Res. 53, 58–77. 10.1007/s12026-012-8270-1 [DOI] [PubMed] [Google Scholar]
- Chauhan S. K., Singh V. V., Rai R., Rai M., Rai G. (2013). Distinct Autoantibody Profiles in Systemic Lupus Erythematosus Patients are Selectively Associated with TLR7 and TLR9 Upregulation. J. Clin. Immunol. 33, 954–964. 10.1007/s10875-013-9887-0 [DOI] [PubMed] [Google Scholar]
- Christensen S. R., Kashgarian M., Alexopoulou L., Flavell R. A., Akira S., Shlomchik M. J. (2005). Toll-like receptor 9 controls anti-DNA autoantibody production in murine lupus. J. Exp. Med. 202, 321–331. 10.1084/jem.20050338 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Christensen S. R., Shupe J., Nickerson K., Kashgarian M., Flavell R. A., Shlomchik M. J. (2006). Toll-like receptor 7 and TLR9 dictate autoantibody specificity and have opposing inflammatory and regulatory roles in a murine model of lupus. Immunity 25, 417–428. 10.1016/j.immuni.2006.07.013 [DOI] [PubMed] [Google Scholar]
- Clinicaltrials.gov (2014). Available at: https://clinicaltrials.gov [accessed November 21, 2014].
- Conrad D. F., Pinto D., Redon R., Feuk L., Gokcumen O., Zhang Y., et al. (2010). Origins and functional impact of copy number variation in the human genome. Nature 464, 704–712. 10.1038/nature08516 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crow M. K. (2014). Type I interferon in the pathogenesis of lupus. J. Immunol. 192, 5459–5468. 10.4049/jimmunol.1002795 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deane J. A., Pisitkun P., Barrett R. S., Feigenbaum L., Town T., Ward J. M., et al. (2007). Control of Toll-like receptor 7 expression is essential to restrict autoimmunity and dendritic cell proliferation. Immunity 27, 801–810. 10.1016/j.immuni.2007.09.009 [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Jager P. L., Richardson A., Vyse T. J., Rioux J. D. (2006). Genetic variation in Toll-like receptor 9 and susceptibility to systemic lupus erythematosus. Arthritis Rheum. 54, 1279–1282. 10.1002/art.21755 [DOI] [PubMed] [Google Scholar]
- De R. C., Fournier N. (2012). Use of an Antibody Directed Against a Membrane Protein. Google Patents.
- Deng Y., Zhao J., Sakurai D., Kaufman K. M., Edberg J. C., Kimberly R. P., et al. (2013). MicroRNA-3148 modulates allelic expression of Toll-like receptor 7 variant associated with systemic lupus erythematosus. PLoS Genet. 9:e1003336. 10.1371/journal.pgen.1003336 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Di Domizio J., Blum A., Gallagher-Gambarelli M., Molens J. P., Chaperot L., Plumas J. (2009). TLR7 stimulation in human plasmacytoid dendritic cells leads to the induction of early IFN-inducible genes in the absence of type I IFN. Blood 114, 1794–1802. 10.1182/blood-2009-04-216770 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dillon S. P., Kurien B. T., Li S., Bruner G. R., Kaufman K. M., Harley J. B., et al. (2012). Sex chromosome aneuploidies among men with systemic lupus erythematosus. J. Autoimmun. 38, J129–J134. 10.1016/j.jaut.2011.10.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dooley M. A., Houssiau F., Aranow C., D’Cruz D. P., Askanase A., Roth D. A., et al. (2013). Effect of belimumab treatment on renal outcomes: results from the phase 3 belimumab clinical trials in patients with SLE. Lupus 22, 63–72. 10.1177/0961203312465781 [DOI] [PubMed] [Google Scholar]
- dos Santos B. P., Valverde J. V., Rohr P., Monticielo O. A., Brenol J. C., Xavier R. M., et al. (2012). TLR7/8/9 polymorphisms and their associations in systemic lupus erythematosus patients from southern Brazil. Lupus 21, 302–309. 10.1177/0961203311425522 [DOI] [PubMed] [Google Scholar]
- Dzionek A., Sohma Y., Nagafune J., Cella M., Colonna M., Facchetti F., et al. (2001). BDCA-2, a novel plasmacytoid dendritic cell-specific type II C-type lectin, mediates antigen capture and is a potent inhibitor of interferon alpha/beta induction. J. Exp. Med. 194, 1823–1834. 10.1084/jem.194.12.1823 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fairhurst A. M., Hwang S. H., Wang A., Tian X. H., Boudreaux C., Zhou X. J., et al. (2008). Yaa autoimmune phenotypes are conferred by overexpression of TLR7. Eur. J. Immunol. 38, 1971–1978. 10.1002/eji.200838138 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fairhurst A. M., Wandstrat A. E., Wakeland E. K. (2006). Systemic lupus erythematosus: multiple immunological phenotypes in a complex genetic disease. Adv. Immunol. 92, 1–69 10.1016/S0065-2776(06)92001-X [DOI] [PubMed] [Google Scholar]
- Fish E. N. (2008). The X-files in immunity: sex-based differences predispose immune responses. Nat. Rev. Immunol. 8, 737–744. 10.1038/nri2394 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Furie R., Petri M., Zamani O., Cervera R., Wallace D. J., Tegzova D., et al. (2011). A phase III, randomized, placebo-controlled study of belimumab, a monoclonal antibody that inhibits B lymphocyte stimulator, in patients with systemic lupus erythematosus. Arthritis Rheum. 63, 3918–3930. 10.1002/art.30613 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garcia-Ortiz H., Velazquez-Cruz R., Espinosa-Rosales F., Jimenez-Morales S., Baca V., Orozco L. (2010). Association of TLR7 copy number variation with susceptibility to childhood-onset systemic lupus erythematosus in Mexican population. Ann. Rheum. Dis. 69, 1861–1865. 10.1136/ard.2009.124313 [DOI] [PubMed] [Google Scholar]
- Garcia-Romo G. S., Caielli S., Vega B., Connolly J., Allantaz F., Xu Z., et al. (2011). Netting neutrophils are major inducers of type I IFN production in pediatric systemic lupus erythematosus. Sci. Transl. Med. 3:73ra20. 10.1126/scitranslmed.3001201 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hahn B. H. (2011). Targeted therapies in systemic lupus erythematosus: successes, failures and future. Ann. Rheum. Dis. 70(Suppl. 1), i64–i66. 10.1136/ard.2010.142208 [DOI] [PubMed] [Google Scholar]
- Henault J., Martinez J., Riggs J. M., Tian J., Mehta P., Clarke L., et al. (2012). Noncanonical autophagy is required for type I interferon secretion in response to DNA-immune complexes. Immunity 37, 986–997. 10.1016/j.immuni.2012.09.014 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hornung V., Rothenfusser S., Britsch S., Krug A., Jahrsdörfer B., Giese T., et al. (2002). Quantitative expression of Toll-like receptor 1–10 mRNA in cellular subsets of human peripheral blood mononuclear cells and sensitivity to CpG oligodeoxynucleotides. J. Immunol. 168, 4531–4537. [DOI] [PubMed] [Google Scholar]
- Hua J., Kirou K., Lee C., Crow M. K. (2006). Functional assay of type I interferon in systemic lupus erythematosus plasma and association with anti-RNA binding protein autoantibodies. Arthritis Rheum. 54, 1906–1916. 10.1002/art.21890 [DOI] [PubMed] [Google Scholar]
- Huang C. M., Huang P. H., Chen C. L., Lin Y. J., Tsai C. H., Huang W. L., et al. (2012). Association of Toll-like receptor 9 gene polymorphism in Chinese patients with systemic lupus erythematosus in Taiwan. Rheumatol. Int. 32, 2105–2109. 10.1007/s00296-011-1925-8 [DOI] [PubMed] [Google Scholar]
- Hur J. W., Shin H. D., Park B. L., Kim L. H., Kim S. Y., Bae S. C. (2005). Association study of Toll-like receptor 9 gene polymorphism in Korean patients with systemic lupus erythematosus. Tissue Antigens 65, 266–270. 10.1111/j.1399-0039.2005.00374.x [DOI] [PubMed] [Google Scholar]
- Kamal A. (2014). The efficacy of novel B cell biologics as the future of SLE treatment: a review. Autoimmun. Rev. 10.1016/j.autrev.2014.08.020 [Epub ahead of print]. [DOI] [PubMed] [Google Scholar]
- Kandimalla E. R., Bhagat L., Wang D., Yu D., Sullivan T., La Monica N., et al. (2013). Design, synthesis and biological evaluation of novel antagonist compounds of Toll-like receptors 7, 8 and 9. Nucleic Acids Res. 41, 3947–3961. 10.1093/nar/gkt078 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kanta H., Mohan C. (2009). Three checkpoints in lupus development: central tolerance in adaptive immunity, peripheral amplification by innate immunity and end-organ inflammation. Genes Immun. 10, 390–396. 10.1038/gene.2009.6 [DOI] [PubMed] [Google Scholar]
- Kawai T., Akira S. (2010). The role of pattern-recognition receptors in innate immunity: update on Toll-like receptors. Nat. Immunol. 11, 373–384. 10.1038/ni.1863 [DOI] [PubMed] [Google Scholar]
- Kirou K. A., Lee C., George S., Louca K., Peterson M. G., Crow M. K. (2005). Activation of the interferon-alpha pathway identifies a subgroup of systemic lupus erythematosus patients with distinct serologic features and active disease. Arthritis Rheum. 52, 1491–1503. 10.1002/art.21031 [DOI] [PubMed] [Google Scholar]
- Komatsuda A., Wakui H., Iwamoto K., Ozawa M., Togashi M., Masai R., et al. (2008). Up-regulated expression of Toll-like receptors mRNAs in peripheral blood mononuclear cells from patients with systemic lupus erythematosus. Clin. Exp. Immunol. 152, 482–487. 10.1111/j.1365-2249.2008.03646.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kono D. H., Baccala R., Theofilopoulos A. N. (2013). TLRs and interferons: a central paradigm in autoimmunity. Curr. Opin. Immunol. 25, 720–727. 10.1016/j.coi.2013.10.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kwok S. K., Lee J. Y., Park S. H., Cho M. L., Min S. Y., Kim H. Y., et al. (2008). Dysfunctional interferon-alpha production by peripheral plasmacytoid dendritic cells upon Toll-like receptor-9 stimulation in patients with systemic lupus erythematosus. Arthritis Res. Ther. 10, R29. 10.1186/ar2382 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lartigue A., Courville P., Auquit I., Francois A., Arnoult C., Tron F., et al. (2006). Role of TLR9 in anti-nucleosome and anti-DNA antibody production in lpr mutation-induced murine lupus. J. Immunol. 177, 1349–1354. 10.4049/jimmunol.177.2.1349 [DOI] [PubMed] [Google Scholar]
- Laska M. J., Troldborg A., Hansen B., Stengaard-Pedersen K., Junker P., Nexo B. A., et al. (2014). Polymorphisms within Toll-like receptors are associated with systemic lupus erythematosus in a cohort of Danish females. Rheumatology (Oxford) 53, 48–55. 10.1093/rheumatology/ket316 [DOI] [PubMed] [Google Scholar]
- Lau C. M., Broughton C., Tabor A. S., Akira S., Flavell R. A., Mamula M. J., et al. (2005). RNA-associated autoantigens activate B cells by combined B cell antigen receptor/Toll-like receptor 7 engagement. J. Exp. Med. 202, 1171–1177. 10.1084/jem.20050630 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lau C. S., Mak A. (2009). The socioeconomic burden of SLE. Nat. Rev. Rheumatol. 5, 400–404. 10.1038/nrrheum.2009.106 [DOI] [PubMed] [Google Scholar]
- Lauwerys B. R., Hachulla E., Spertini F., Lazaro E., Jorgensen C., Mariette X., et al. (2013). Down-regulation of interferon signature in systemic lupus erythematosus patients by active immunization with interferon alpha-kinoid. Arthritis Rheum. 65, 447–456. 10.1002/art.37785 [DOI] [PubMed] [Google Scholar]
- Leadbetter E. A., Rifkin I. R., Hohlbaum A. M., Beaudette B. C., Shlomchik M. J., Marshak-Rothstein A. (2002). Chromatin-IgG complexes activate B cells by dual engagement of IgM and Toll-like receptors. Nature 416, 603–607. 10.1038/416603a [DOI] [PubMed] [Google Scholar]
- Looney R. J., Anolik J., Sanz I. (2010). A perspective on B-cell-targeting therapy for SLE. Mod. Rheumatol. 20, 1–10. 10.1007/s10165-009-0213-x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lovgren T., Eloranta M. L., Kastner B., Wahren-Herlenius M., Alm G. V., Ronnblom L. (2006). Induction of interferon-alpha by immune complexes or liposomes containing systemic lupus erythematosus autoantigen- and Sjögren’s syndrome autoantigen-associated RNA. Arthritis Rheum. 54, 1917–1927. 10.1002/art.21893 [DOI] [PubMed] [Google Scholar]
- Lu K. C., Yang H. Y., Lin Y. F., Kao S. Y., Lai C. H., Chu C. M., et al. (2011). The T-1237C polymorphism of the Toll-like receptor-9 gene is associated with chronic kidney disease in a Han Chinese population. Tohoku J. Exp. Med. 225, 109–116. 10.1620/tjem.225.109 [DOI] [PubMed] [Google Scholar]
- Lyn-Cook B. D., Xie C., Oates J., Treadwell E., Word B., Hammons G., et al. (2014). Increased expression of Toll-like receptors (TLRs) 7 and 9 and other cytokines in systemic lupus erythematosus (SLE) patients: ethnic differences and potential new targets for therapeutic drugs. Mol. Immunol. 61, 38–43. 10.1016/j.molimm.2014.05.001 [DOI] [PubMed] [Google Scholar]
- Mathian A., Amoura Z., Adam E., Colaone F., Hoekman M. F., Dhellin O., et al. (2011). Active immunisation of human interferon alpha transgenic mice with a human interferon alpha Kinoid induces antibodies that neutralise interferon alpha in sera from patients with systemic lupus erythematosus. Ann. Rheum. Dis. 70, 1138–1143. 10.1136/ard.2010.141101 [DOI] [PubMed] [Google Scholar]
- Means T. K., Latz E., Hayashi F., Murali M. R., Golenbock D. T., Luster A. D. (2005). Human lupus autoantibody-DNA complexes activate DCs through cooperation of CD32 and TLR9. J. Clin. Invest. 115, 407–417. 10.1172/JCI23025 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meier A., Chang J. J., Chan E. S., Pollard R. B., Sidhu H. K., Kulkarni S., et al. (2009). Sex differences in the Toll-like receptor-mediated response of plasmacytoid dendritic cells to HIV-1. Nat. Med. 15, 955–959. 10.1038/nm.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Merrill J. T., Wallace D. J., Petri M., Kirou K. A., Yao Y., White W. I., et al. (2011). Safety profile and clinical activity of sifalimumab, a fully human anti-interferon alpha monoclonal antibody, in systemic lupus erythematosus: a phase I, multicentre, double-blind randomised study. Ann. Rheum. Dis. 70, 1905–1913. 10.1136/ard.2010.144485 [DOI] [PubMed] [Google Scholar]
- Miettinen M., Sareneva T., Julkunen I., Matikainen S. (2001). IFNs activate Toll-like receptor gene expression in viral infections. Genes Immun. 2, 349–355. [DOI] [PubMed] [Google Scholar]
- Miettinen M., Veckman V., Latvala S., Sareneva T., Matikainen S., Julkunen I. (2008). Live Lactobacillus rhamnosus and Streptococcus pyogenes differentially regulate Toll-like receptor (TLR) gene expression in human primary macrophages. J. Leukoc. Biol. 84, 1092–1100. 10.1189/jlb.1206737 [DOI] [PubMed] [Google Scholar]
- Mu R., Sun X. Y., Lim L. T., Xu C. H., Dai C. X., Su Y., et al. (2012). Toll-like receptor 9 is correlated to disease activity in Chinese systemic lupus erythematosus population. Chin. Med. J. (Engl.) 125, 2873–2877 10.3760/cma.j.issn.0366-6999.2012.16.014 [DOI] [PubMed] [Google Scholar]
- Murphy G., Lisnevskaia L., Isenberg D. (2013). Systemic lupus erythematosus and other autoimmune rheumatic diseases: challenges to treatment. Lancet 382, 809–818 10.1016/S0140-6736(13)60889-2 [DOI] [PubMed] [Google Scholar]
- Nagase H., Okugawa S., Ota Y., Yamaguchi M., Tomizawa H., Matsushima K., et al. (2003). Expression and function of Toll-like receptors in eosinophils: activation by Toll-like receptor 7 ligand. J. Immunol. 171, 3977–3982. [DOI] [PubMed] [Google Scholar]
- Nakano S., Morimoto S., Suzuki J., Nozawa K., Amano H., Tokano Y., et al. (2008). Role of pathogenic auto-antibody production by Toll-like receptor 9 of B cells in active systemic lupus erythematosus. Rheumatology (Oxford) 47, 145–149. 10.1093/rheumatology/kem327 [DOI] [PubMed] [Google Scholar]
- Navarra S. V., Guzman R. M., Gallacher A. E., Hall S., Levy R. A., Jimenez R. E., et al. (2011). Efficacy and safety of belimumab in patients with active systemic lupus erythematosus: a randomised, placebo-controlled, phase 3 trial. Lancet 377, 721–731 10.1016/S0140-6736(10)61354-2 [DOI] [PubMed] [Google Scholar]
- Ng M. W., Lau C. S., Chan T. M., Wong W. H., Lau Y. L. (2005). Polymorphisms of the Toll-like receptor 9 (TLR9) gene with systemic lupus erythematosus in Chinese. Rheumatology (Oxford) 44, 1456–1457. 10.1093/rheumatology/kei120 [DOI] [PubMed] [Google Scholar]
- Nickerson K. M., Christensen S. R., Shupe J., Kashgarian M., Kim D., Elkon K., et al. (2010). TLR9 regulates TLR7- and MyD88-dependent autoantibody production and disease in a murine model of lupus. J. Immunol. 184, 1840–1848. 10.4049/jimmunol.0902592 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niewold T. B., Hua J., Lehman T. J., Harley J. B., Crow M. K. (2007). High serum IFN-alpha activity is a heritable risk factor for systemic lupus erythematosus. Genes Immun. 8, 492–502. 10.1038/sj.gene.6364408 [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Neill L. A., Sheedy F. J., Mccoy C. E. (2011). MicroRNAs: the fine-tuners of Toll-like receptor signalling. Nat. Rev. Immunol. 11, 163–175. 10.1038/nri2957 [DOI] [PubMed] [Google Scholar]
- Ortiz-Neu C., LeRoy E. C. (1969). The coincidence of Klinefelter’s syndrome and systemic lupus erythematosus. Arthritis Rheum. 12, 241–246. 10.1002/art.1780120312 [DOI] [PubMed] [Google Scholar]
- Papadimitraki E. D., Choulaki C., Koutala E., Bertsias G., Tsatsanis C., Gergianaki I., et al. (2006). Expansion of Toll-like receptor 9-expressing B cells in active systemic lupus erythematosus: implications for the induction and maintenance of the autoimmune process. Arthritis Rheum. 54, 3601–3611. 10.1002/art.22197 [DOI] [PubMed] [Google Scholar]
- Park H., Kim J. I., Ju Y. S., Gokcumen O., Mills R. E., Kim S., et al. (2010). Discovery of common Asian copy number variants using integrated high-resolution array CGH and massively parallel DNA sequencing. Nat. Genet. 42, 400–405. 10.1038/ng.555 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Petri M., Wallace D. J., Spindler A., Chindalore V., Kalunian K., Mysler E., et al. (2013). Sifalimumab, a human anti-interferon-alpha monoclonal antibody, in systemic lupus erythematosus: a phase I randomized, controlled, dose-escalation study. Arthritis Rheum. 65, 1011–1021. 10.1002/art.37824 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piotrowski P., Lianeri M., Wudarski M., Olesinska M., Jagodzinski P. P. (2013). Contribution of Toll-like receptor 9 gene single-nucleotide polymorphism to systemic lupus erythematosus. Rheumatol. Int. 33, 1121–1125. 10.1007/s00296-012-2509-y [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pisitkun P., Deane J. A., Difilippantonio M. J., Tarasenko T., Satterthwaite A. B., Bolland S. (2006). Autoreactive B cell responses to RNA-related antigens due to TLR7 gene duplication. Science 312, 1669–1672. 10.1126/science.1124978 [DOI] [PubMed] [Google Scholar]
- Rönnblom L., Eloranta M. L., Alm G. V. (2006). The type I interferon system in systemic lupus erythematosus. Arthritis Rheum. 54, 408–420. [DOI] [PubMed] [Google Scholar]
- Rovin B. H., Furie R., Latinis K., Looney R. J., Fervenza F. C., Sanchez-Guerrero J., et al. (2012). Efficacy and safety of rituximab in patients with active proliferative lupus nephritis: the Lupus Nephritis Assessment with Rituximab study. Arthritis Rheum. 64, 1215–1226. 10.1002/art.34359 [DOI] [PubMed] [Google Scholar]
- Rullo O. J., Tsao B. P. (2013). Recent insights into the genetic basis of systemic lupus erythematosus. Ann. Rheum. Dis. 72(Suppl. 2), ii56–ii61. 10.1136/annrheumdis-2012-202351 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sacre K., Criswell L. A., Mccune J. M. (2012). Hydroxychloroquine is associated with impaired interferon-alpha and tumor necrosis factor-alpha production by plasmacytoid dendritic cells in systemic lupus erythematosus. Arthritis Res. Ther. 14, R155. 10.1186/ar3895 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santiago-Raber M. L., Kikuchi S., Borel P., Uematsu S., Akira S., Kotzin B. L., et al. (2008). Evidence for genes in addition to Tlr7 in the Yaa translocation linked with acceleration of systemic lupus erythematosus. J. Immunol. 181, 1556–1562. 10.4049/jimmunol.181.2.1556 [DOI] [PubMed] [Google Scholar]
- Sirén J., Pirhonen J., Julkunen I., Matikainen S. (2005). IFN-alpha regulates TLR-dependent gene expression of IFN-alpha, IFN-beta, IL-28, and IL-29. J. Immunol. 174, 1932–1937. [DOI] [PubMed] [Google Scholar]
- Shen N., Fu Q., Deng Y., Qian X., Zhao J., Kaufman K. M., et al. (2010). Sex-specific association of X-linked Toll-like receptor 7 (TLR7) with male systemic lupus erythematosus. Proc. Natl. Acad. Sci. U.S.A. 107, 15838–15843. 10.1073/pnas.1001337107 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shrivastav M., Niewold T. B. (2013). Nucleic Acid sensors and type I interferon production in systemic lupus erythematosus. Front. Immunol. 4:319. 10.3389/fimmu.2013.00319 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stohl W. (2013). Future prospects in biologic therapy for systemic lupus erythematosus. Nat. Rev. Rheumatol. 9, 705–720. 10.1038/nrrheum.2013.136 [DOI] [PubMed] [Google Scholar]
- Suarez-Farinas M., Arbeit R., Jiang W., Ortenzio F. S., Sullivan T., Krueger J. G. (2013). Suppression of molecular inflammatory pathways by Toll-like receptor 7, 8, and 9 antagonists in a model of IL-23-induced skin inflammation. PLoS ONE 8:e84634. 10.1371/journal.pone.0084634 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Subramanian S., Tus K., Li Q. Z., Wang A., Tian X. H., Zhou J., et al. (2006). A Tlr7 translocation accelerates systemic autoimmunity in murine lupus. Proc. Natl. Acad. Sci. U.S.A. 103, 9970–9975. 10.1073/pnas.0603912103 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun X., Wiedeman A., Agrawal N., Teal T. H., Tanaka L., Hudkins K. L., et al. (2013). Increased ribonuclease expression reduces inflammation and prolongs survival in TLR7 transgenic mice. J. Immunol. 190, 2536–2543. 10.4049/jimmunol.1202689 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tao K., Fujii M., Tsukumo S., Maekawa Y., Kishihara K., Kimoto Y., et al. (2007). Genetic variations of Toll-like receptor 9 predispose to systemic lupus erythematosus in Japanese population. Ann. Rheum. Dis. 66, 905–909. 10.1136/ard.2006.065961 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Torcia M. G., Nencioni L., Clemente A. M., Civitelli L., Celestino I., Limongi D., et al. (2012). Sex differences in the response to viral infections: TLR8 and TLR9 ligand stimulation induce higher IL10 production in males. PLoS ONE 7:e39853. 10.1371/journal.pone.0039853 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vollmer J., Tluk S., Schmitz C., Hamm S., Jurk M., Forsbach A., et al. (2005). Immune stimulation mediated by autoantigen binding sites within small nuclear RNAs involves Toll-like receptors 7 and 8. J. Exp. Med. 202, 1575–1585. 10.1084/jem.20051696 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang B., Higgs B. W., Chang L., Vainshtein I., Liu Z., Streicher K., et al. (2013). Pharmacogenomics and translational simulations to bridge indications for an anti-interferon-alpha receptor antibody. Clin. Pharmacol. Ther. 93, 483–492. 10.1038/clpt.2013.35 [DOI] [PubMed] [Google Scholar]
- Xu C. J., Zhang W. H., Pan H. F., Li X. P., Xu J. H., Ye D. Q. (2009). Association study of a single nucleotide polymorphism in the exon 2 region of Toll-like receptor 9 (TLR9) gene with susceptibility to systemic lupus erythematosus among Chinese. Mol. Biol. Rep. 36, 2245–2248. 10.1007/s11033-008-9440-z [DOI] [PubMed] [Google Scholar]
- Yang Z., Liang Y., Qin B., Li C., Zhong R. (2012). TLR9 polymorphisms and systemic lupus erythematosus risk in Asians: a meta-analysis study. Cytokine 57, 282–289. 10.1016/j.cyto.2011.11.017 [DOI] [PubMed] [Google Scholar]
- Yao Y., Richman L., Higgs B. W., Morehouse C. A., De Los Reyes M., Brohawn P., et al. (2009). Neutralization of interferon-alpha/beta-inducible genes and downstream effect in a phase I trial of an anti-interferon-alpha monoclonal antibody in systemic lupus erythematosus. Arthritis Rheum. 60, 1785–1796. 10.1002/art.24557 [DOI] [PubMed] [Google Scholar]
- Yu C., Gershwin M. E., Chang C. (2014). Diagnostic criteria for systemic lupus erythematosus: a critical review. J. Autoimmun. 48–49, 10–13. 10.1016/j.jaut.2014.01.004 [DOI] [PubMed] [Google Scholar]
- Yu P., Wellmann U., Kunder S., Quintanilla-Martinez L., Jennen L., Dear N., et al. (2006). Toll-like receptor 9-independent aggravation of glomerulonephritis in a novel model of SLE. Int. Immunol. 18, 1211–1219. 10.1093/intimm/dxl067 [DOI] [PubMed] [Google Scholar]
- Zarember K. A., Godowski P. J. (2002). Tissue expression of human Toll-like receptors and differential regulation of Toll-like receptor mRNAs in leukocytes in response to microbes, their products, and cytokines. J. Immunol. 168, 554–561. [DOI] [PubMed] [Google Scholar]
- Zorro S., Arias M., Riano F., Paris S., Ramirez L. A., Uribe O., et al. (2009). Response to ODN-CpG by B Cells from patients with systemic lupus erythematosus correlates with disease activity. Lupus 18, 718–726. 10.1177/0961203309103098 [DOI] [PubMed] [Google Scholar]