

Abstract
Synthetic high polymer flocculants, frequently utilized for flocculating efficiency and low cost, recently have been discovered as producing increased risk to human health and the environment. Development of a more efficient and environmentally sound alternative flocculant agent is investigated in this paper. Bioflocculants are produced by microorganisms and may exhibit a high rate of flocculation activity. The bioflocculant ETH-2, with high flocculating activity (2849 mg Kaolin particle/mg ETH-2), produced by strain Enterobacter sp. isolated from activated sludge, was systematically investigated with regard to its production, characterization, and flocculation mechanism. Analyses of microscopic observation, zeta potential and ETH-2 structure demonstrates the bridging mechanism, as opposed to charge neutralization, was responsible for flocculation of the ETH-2. ETH-2 retains high molecular weight (603 to 1820 kDa) and multi-functional groups (hydroxyl, amide and carboxyl) that contributed to flocculation. Polysaccharides mainly composed of mannose, glucose, and galactose, with a molar ratio of 1∶2.9∶9.8 were identified as the active constituents in bioflocculant. The structure of the long backbone with active sites of polysaccharides was determined as a primary basis for the high flocculation activity. Bioflocculant ETH-2 is cation independent, pH tolerant, and thermally stable, suggesting a potential fit for industrial application.
Introduction
Flocculating agents, widely used in industrial processes such as wastewater treatment, are classified into three groups i.e., inorganic flocculants such as aluminium sulphate; organic synthetic high polymer flocculants such as polyacrylamide (PAM); natural flocculants or bioflocculant such as microbial flocculant [1], [2]. Synthetic high polymer flocculants have been most frequently utilized due to their flocculating efficiency and low cost, however, there is increasing evidence of risk to human health and the environment [1]–[3]. Acrylamide monomer, for example, is derived from PAM and acts as a strong carcinogen and neurotoxin to humans.
Bioflocculant research seeks the benefits of a treatment with high production rate and biodegradable, innocuous waste products while solving the challenges of its tendency for low flocculation activity and high cost. Varying types of microorganisms, including bacteria [4]–[10], fungi [11], [12] and algae [13], [14], have been isolated from soil and activated sludge, and shown to produce bioflocculant. Recent studies have more specifically focused on various bioflocculant producing bacteria including; (Enterobacter aerogenes [4], Halomonas sp. [5], Chryseobacterium daeguense [6], Klebsiella pneumoniae [7], Paenibacillus sp. [8], Corynebacterium glutamicum [9], Bacillus licheniformis [10], et al.).
Bioflocculant research has focused on the isolation of bioflocculant-producing microorganisms and their production, whereas flocculation mechanisms and active constituents of bioflocculation are generally undetermined; this is especially true for cation independent bioflocculant. Active constituents within bioflocculant structure must be understood to fully comprehend flocculation functions. Colorimetric methods for the quantification of bioflocculant chemicals composition, Gel Permeation Chromatography (GPC) for Molecular Weight (MW) characterization, Fourier Transform Infrared Spectroscopy (FTIR) and X-ray Photoelectron Spectroscopy (XPS) for flocculation functional group identification have been utilized in limited studies to elucidate the composition and structure of bioflocculant [10]; however, the relationship between their structure and flocculation activity remains unclear.
In this study we isolated a microorganism, ETH-2, from activated sludge and identified it as Enterobacter sp. (E. sp.), which secretes bioflocculant. High flocculation activity to kaolin suspension without the aid of cations occurs with this bioflocculant. The production culture medium composition of E. sp. ETH-2 was optimized and the main factors influencing flocculation efficiency (pH, temperature, dosage, and cation) were then investigated. Bioflocculant structure was characterized by combining the chemicals analysis, GC-MS, GPC, FTIR, and XPS with the flocculation activity. The flocculation mechanism was then proposed according to the results.
Materials and Methods
Ethic Statement
Activated sludge used in this study was obtained from the secondary settling tank of Tang Jia Qiao municipal wastewater treatment plant (WWTP) in Chongqing, China. All necessary permits for collection of the sludge from this system were granted by Tang Jia Qiao Municipal WWTP and Chongqing Municipal Construction Bureau. All field work has been conducted according to the relevant national guidelines.
Isolation of flocculant-producing strain ETH-2 of Enterobacter sp
Isolation of strain
Activated sludge samples were collected in 1L sterilized Nalgene HDPE big mouth sample bottles (Fisher Scientific, USA) and stored on ice during transport back to the laboratory with immediate treatment upon arrival at the lab. Following 10 fold series dilution, 100 ul dilutions were planted on Luria-Bertani (LB) agar plates and cultured at 30°C overnight. Strains with unique colony morphologies were selected and inoculated into 250 ml flasks containing 100 ml flocculation selecting mediums for 3 days at 30°C with 180 rpm shaking. One ml culture was utilized for flocculation activity test assay. Fifty-four strains were isolated. Within them, ETH-2 strain demonstrated the highest flocculating activity and was selected for further study. The SEM image of the strain was obtained by a scanning electron microscope (Helios, NanoLab 600i, FEI). Strain ETH-2 has been deposited into the China Center for Type Culture Collection (CCTCC) (Wuhan, China) (accession number CCTCC M 2013042).
The LB agar contains (per liter): tryptone, 10 g; yeast extract, 5 g; and NaCl, 10 g; agar, 15 g. The flocculation selecting medium contains (per liter): Glucose 10 g; KH2PO4 2 g; K2HPO4 5 g; MgSO4·7H2O 0.2 g; NaCl 0.1 g; urea 0.5 g; yeast extract 0.5 g. After optimization of carbon and nitrogen source (Materials S1 and Figure S1 in File S1), the culture media was slightly modified as containing (per liter): glucose, 10 g; NaNO3, 1 g; KH2PO4, 2 g; K2HPO4, 5 g; NaCl, 0.1 g; MgSO4·7H2O, 0.2 g. The initial pH of all media was adjusted to 7.3±0.1 with NaOH (1 M) and HCl (0.5 M).
Identification of strain
DNA was extracted, according to vendor protocol, utilizing the TIA Namp Bacteria DNA Kit (TianGen, China). The 16S rRNA gene was amplified utilizing bacterial universal primers 27F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492R (5′-GGTTACCTTGTTACGACTT-3′) under the following conditions: Initial denaturation at 95°C for 10 min, followed by 30 cycles (30 s at 95°C, 30 s at 55°C, and 1 min 30 s at 72°C) and a final 10 min extension at 72°C. The PCR production was then purified utilizing the QIAquick PCR Purification Kit (Qiagen, USA), cloned into pMD19-T vector (Takara, China), and then sequenced. The obtained 16S rRNA gene sequence was assembled with Seqman II 5.0 (DNASTAR) [15] and analyzed using the BLAST program. Phylogenetic tree construction was followed as previous described [16]. The 16S rRNA gene sequence has been deposited into the GenBank with accession number KF739069.
Purification and Characterization of ETH-2 Bioflocculant
Extraction and Purification of ETH-2 Bioflocculant
ETH-2 was inoculated into a 200 ml optimization medium and cultivated at 30°C for 24 h at 180 rpm and the cultured broth was harvested for bioflocculant production extraction. Comparisons of flocculation extraction efficiency through varying purification approaches were attained. Heating at 80°C demonstrated the highest extraction efficiency (Table S1 in File S1) and was used for bioflocculant extraction as previously described [17]. Briefly, the cultured broth was centrifuged at 12,000 rpm for 5 min and then washed three times with 0.9% NaCl. The suspended pellets were dissolved into 0.1 volume 0.9% NaCl and heated at 80°C for 15 min.
Bioflocculant purification was performed using a minor modification of the method described [10], [18]. After natural cooling, the heating extracted cell-free supernatant was obtained by centrifugation at 12,000 rpm for 10 min and filtered with 0.22 µm membrane. Cold ethanol was added to the supernatant and left overnight at 4°C. The precipitate was collected by centrifugation at 12,000 rpm for 10 min and dissolved in ultrapure water and then dialyzed with a molecular weight cutoff: 8000–14400 Da (36 MM, Biosharp, USA). Cetyltrimethyl ammonium bromide (CTAB) (2%, wt.%) was added to the solution with stirring at 100 rpm. After 3 h, the CTAB-bioflocculant complex precipitate was collected by centrifugation and then dissolved in 0.5 M NaCl solution. Two volumes of cold ethanol were added to solution to obtain the precipitate, and the precipitate was washed several times with ethanol. The precipitate was dissolved in ultrapure water and dialyzed against ultrapure water overnight. Finally, the purified bioflocculant, designated as ETH2, was obtained and lyophilized for further experiments.
Characterization of ETH-2 Bioflocculant
Determination of total sugar was completed utilizing the phenol-sulfuric acid method with glucose as the standard solution [4], [10]. Protein was determined by applying the Bradford method with the bovine serum albumin (TianGen, China) as the standard. Analysis of monosaccharide composition was determined by GC-MS with the following procedure: 10 mg lyophilized ETH-2 was hydrolyzed with 2 ml of 1 M H2SO4 in 100°Coil bath for 4 hours in a sealed glass tube. The hydrolysate was then centrifuged at 8000 rpm for 5 min after its pH was neutralized to 7.0 with BaCO3. The supernatant was filtered with 0.22 µm hydrophilic filter membrane (JinTeng, China) and the filtrate collected for lyophilized (Virtis, BT4KXL, USA). The lyophilized powder reacted with 10 mg hydroxylamine hydrochloride and 0.5 ml pyridine in 90°Cwater bath for 30 min and then was acetylated with 0.5 ml acetic anhydride in 90°C water bath for another 30 min. The acetylated aldononitrile of ETH-2 were analyzed on Agilent 7890A-5977 GC/MS operated in the EI mode and a HP-5MS fused silica capillary column (30 m×0.32 mm×0.25 mm). Helium was used as carrier gas with 1 ml/min flow rate. Temperatures of injector and detector were set at 300°C and 260°C, respectively, and the initial column temperature was 130°C for 5 min, increased to 240°C at a rate of 4°C/min and held for 5 min. Sugar identification was performed by comparison with reference sugars. Relative molar proportions were then calculated by the area normalization method.
The molecular weight of ETH-2 was analyzed by using Lc-10ADVP Gel Permeation Chromatography (GPC) equipped with RID-10A detector (Shimadzu, Japan) and TSK G4000PWxl columns (Shimadzu, Japan) at an optimal operation temperature 40°C. The column was calibrated by standard dextrans. DDI water was used as the mobile phase with 0.5 ml/min flow rate. Prior to injection, the sample was filtrated utilizing a 0.45 µm filter. Number and weight-average molecular weight served to characterize the bioflocculant MWs [19].
The element of ETH-2was analyzed by X-Ray Photoelectron Spectroscopy (XPS) (XSAM800, Kratos, UK) under FAT mode with a vacuum degree of 5×10−7 Pa, a power of 12 kv and electric current of 12 mA. The laser source was aluminum target and the 284.8 eV was used for correction.
The Fourier Transorm Infrared Spectrometer (IR) (ThermoFisher, Nicolet 6700, USA) was used for the detection of functional groups of ETH-2 and IR of ETH-2 sample was measured from 400–4000 cm−1.
Flocculation of Kaolin by ETH-2 Bioflocculant
Flocculation dynamics
Flocculation dynamics of cell and produced biofloculant was measured according to the used kaolin suspension method. One ml agent was mixed into 50 ml of 4 g/L kaolin suspension in a 50 ml graduated cylinder covered with clean film. The test cylinder was gently shaken and settled at room temperature. At prescribed time, 3 ml of supernatant was carefully removed from the upper layer of solution and measured at a 550 nm absorbance. Control experiment was also conducted without flocculation agents.
Flocculation impact factors
Effects of impact factors (dosages, cations, pH and temperature) were investigated to study the flocculation activity of the ETH-2. Purified ETH-2 bioflocculant was dissolved in ultrapure water to get a 0.066 mg/ml concentration. A series volume (0.05, 0.1, 0.2, 0.5, 1, 2, 5, 10, 20 and 50 ml) of ETH-2 solution, equal to 0.003, 0.007, 0.013, 0.033, 0.066, 0.132, 0.330, 0.660, 1.320, and 3.300 mg ETH-2, were mixed into Kaolin suspension (200 mg Kaolin particles in 50 ml solution) to test the dosage impact. Cations effects were tested by adding varying cations, 1 milliliter/each (NaCl, KCl, CaCl2, MgCl2, AlCl3, FeCl3, concentration is 0.09 M) solution to the Kaolin suspension system. pH values of Kaolin clay solution were then adjusted to 3.0, 4.0, 5.0, 6.0, 7.0, 8.0, 9.0 and 10.0 respectively, with HCl (0.5 M) and NaOH (0.5 M), and then mixed with ETH-2 solution to test pH influence. Thermal stability of ETH-2 evaluation was determined by measuring the flocculation efficiency after ETH-2 was incubated at various temperatures (40, 50, 60, 70, 80, 90, 100°C) for 30 minutes [20].
Flocculation mechanism
The zeta potentials of ETH-2, Kaolin clay suspension, and mixture of ETH-2 and Kaolin clay were measured by a zeta potential analyzer (Malvern, UK) to test flocculation mechanism (bridging or charge neutralization).
Flocculation activity evaluation
Flocculation activity was measured according to the previously described kaolin suspension method [2], [21], [22]. One ml of agent was mixed with 50 ml of 4 g/L kaolin suspension in a 50 ml graduated cylinder covered by clean film. The test cylinder was gently shaken and settled for 5 min at room temperature. Three ml of supernatant were carefully removed from the upper layer of solution and measured at a 550 nm absorbance (TU-1901, Persee, China). A separate control experiment was conducted without flocculation agents. Flocculation activity was calculated according to the following equation:
Where A is the OD550 of sample, while B is the OD550 of control.
Results and Discussion
Isolation of flocculant-producing strain ETH-2 of Enterobacter sp
We compared the flocculation efficiency of ETH-2 strain culture broth (OD600 = 1.8) and PAM (0.1%, w/v %) in the test of flocculation of wine process wastewater and the former showed higher flocculation efficiency (Figure 1). Figure 2 shows the morphological analysis of the strain ETH-2. The strain formed rounded, cream-colored, smooth colonies on agar culture media, while the SEM image indicates the strain was short rod-shaped, flagellated, motile aerobe and had convex. The phylogenetic identity of the strain ETH-2 by 16S rDNA sequencing revealed that it was most closely to Enterobacter gergoviae (98% similarity) (Figure 3). Although in the past, several bioflocculant producing bacteria have been noted, few studies report bioflocculant production with Enterobacter sp. as the source. Currently, just four strains affiliated with Enterobacter, including Enterobacter sp. BY-29 [23] , Enterobacter cloacae WD7 [20], Enterobacter sp. EP3 [24] and Enterobacter aerogenes [4], have been isolated and their producing bioflocculant characterized.
Figure 1. Comparison of flocculation ability by ETH-2 and PAM for wine process wastewater.

Three hundred µl ETH-2 strain culture broth (OD600 = 1.8) and 0.1% PAM were added into 5 ml wine process wastewater (COD: 4299.0 mg/L; pH = 4.0; SS = 315.0 mg/L), respectively. The cylinder was gently shaken, settled at room temperature for 5 min, and measured the OD550.
Figure 2. SEM image of Enterobacter sp. ETH-2.

Figure 3. Phylogenetic tree based on 16S rRNA sequences (Bar: sequence dissimilarity of 0.02%).

Purification and Characterization of ETH-2 Bioflocculant
Characterization of the purified ETH-2
A total of 65.6 mg of purified ETH-2 was recovered from 1 L of fermentation broth. The total sugar and total protein content of ETH-2 was 91.7% and 1.8%, respectively, indicating a major polysaccharides component. Comparison of retention time and corresponding mass spectra with known standards (Figure 4a) was achieved through examination and identification of the alditol acetate derivatives from monosaccharide by GC-MS. Three major carbohydrate peaks at 23.79, 24.09, and 24.69 min were depicted on a gas chromatogram (Figure 4b), which were attributed to mannose, glucose, and galactose, respectively, with a molar ratio of 1∶2.9∶9.8.
Figure 4. Gas chromatogram of standard monosaccharide (a) and alditol acetate derivatives from ETH-2 (b).

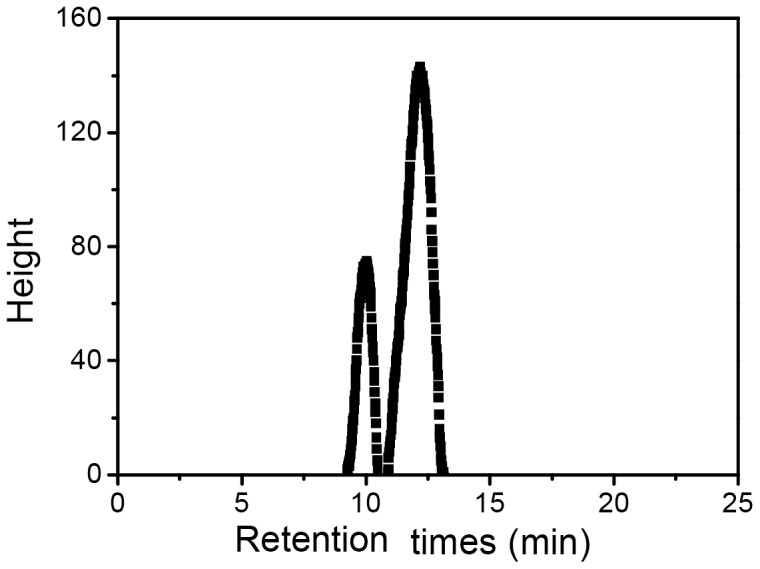
RI chromatograms with two retention times (10.03 and 12.18 minutes) of ETH-2 are represented in Figure 5. The apparent weight and number averaged MWs (M w and M n) of the two constituents are listed in Table S2 in File S1. The MWs of the main constituents in ETH-2 ranges from 603to 1820 kDa, which is within the high MWs range, demonstrating the high flocculation ability of ETH-2. High MW bioflocculant possess more adsorption points for bridging so greater flocs are obtained [1], [25]. The reported efficient bioflocculant usually has high MWs, ranging from 102 to 103 kDa. For example, Bacillus megaterium TF10 produced 1.0–2.5×103 kDa bioflocculant [26] and Bacillus licheniformis yield 1.8×103 kDa bioflocculant [10].
Figure 5. GPC chromatogram of ETH-2.
The elemental analysis of ETH-2 is indicated in Figure 6. Peaks of 1 s core level of C, O, and N are clearly visible, accounting for 64.7%, 28.6%, and 4.6%, mass fractions, respectively. A peak of 2 p core level of P is detected, presenting a small amount of mass fraction, 1.0%. XPS also detected lower (1.0% mass fraction) 1 s core level of Na, probably due to sodium contamination. High resolution scans of C 1s, O 1s, N1s, and P 2p were deconvoluted to obtain the corresponding functional groups. Total four C (1s) (Figure 6c), two O (1s) (Figure 6d), one N (1s) (Figure 6e), one P (2p) (Figure 6f), and one Na (1s) were observed with the assignment and quantification of these functional groups listed in Table S3 in File S1. The C 1s peak was resolved into four component peaks. The peaks at 284.8 eV are associated with C-(C,H) of lipids or amino acid side chains, accounting for the largest percentage in the spectral band (31.3%). The peak at 286.2 eV, which is attributed to C-(O) from alcohol, ether amine, or amide, also presents a large part (21.9%). The peaks at 287.4 eV are associated with O = CO, as in carboxylate, carbonyl, amide, acetal, or hemiacetal respectively, account for 8.7%. The peaks at 288.6 eV associated with C = O, as in carboxyl or ester, are much lower (2.8%). The O1s peak is decomposed into two peaks. The O1s peak at 531.9 eV (11.5%) is mainly attributed to the C = O, as in carboxylate, carbonyl, ester, or amide. The second O1s peak at 533.1 eV (17.1%) is attributed to C-O, as in alcohols, hemiacetal, or acetal groups. Single peak is detected in N 1s and P 2p, respectively. The peak at 399.9 eV is attributed to nonprotonated nitrogen (Nnonpr, 4.58%) from amines and amides. The peak at 133.6 eV is attributed to phosphate (0.09%).
Figure 6. FTIR spectra of ETH-2 (a), X-ray photoelectron spectra of ETH-2 (b) and high resolution 1 s XPS spectra of C, O,N, and P from ETH-2 are shown in (c), (d), (e) and (f), respectively.

As shown in Figure 6a, the intense absorption peak at 3,399 cm−1 (characteristic of a hydroxyl group) could be caused by the vibration of -OH or -NH in the sugar ring of polysaccharides. Two small stretching bands at 2,919 cm−1 and 2,850 cm−1 indicated = CH2 of carbohydrate. Predominantly assigned to C = O stretching associated with proteins, is the band at 1661 cm−1, while the band at 1593 cm−1 originates from N-H bending and C-N stretching vibrations in -CO-NH of proteins. The band at 1404cm−1 is attributed to C = O symmetric stretching of –COO groups. Assigned to C-H stretches associated with lipids, is the very small band at 1319 cm−1. Two weak peaks at 1075 cm−1 and 1024 cm−1 indicate C-O is also the characteristic absorption peak of polysaccharides.
The Infrared Spectrum Analysis revealed characteristic peaks for polysaccharides, protein and other materials such as lipids, and demonstrated the presence of the functional groups: hydroxyl (-OH), amide (-CO-NH) and carboxyl (-COO-). FTIR and XPS analysis demonstrates the presence of hydroxyl (-OH), amide (-CO-NH), and carboxyl (-COO−) in ETH-2. These function groups are preferred for the flocculation process for purposes similar to observations made in other bioflocculants [3], [25]–[27]. Two major roles in flocculation are important attributes of these function groups: hydrophilicity characteristics are utilized to extend the polymer chain [25]; and the groups span the gap between articles to adsorbs particles [3], [26].
Glycoproteins [28], [29], lipids [30] and especially polysaccharides [3], [10], [11], [31] and proteins [32], [33], have been found to be the key constituents of bioflocculants. For instance, Proteus mirabilis [3], Bacillus licheniformis [10], Aspergillus flavus [11], and Pullularia Pullulans [31] produced polysaccharides based bioflocculant, while Nocardia Amarae [33] and Pseudomonas sp. A-99 [32] produced protein based bioflocculant. Polysaccharides bioflocculants composed of mannose, glucose, and galactose are the key constituents in this study and are the active constituents of ETH-2 (as opposed to protein) as further demonstrated by the thermal stability qualities of ETH-2.
Flocculation of Kaolin by ETH-2 Bioflocculant
Flocculation behavior
Between 200–300 µm flocs were formed by the ETH-2 strain when tested for kaolin clay flocculation (Figure 7b), indicating its advantage on flocculation. The purified bioflocculant ETH-2 also formed 100–200 µm flocs during kaolin clay flocculation testing (Figure 7c), larger than flocs formed by Bacillus megaterium TF10 (20–30 µm) [26]. The flocculation dynamics indicated that flocculation efficiency of ETH-2 reached 81.8% at 0.5 min, peaked (92.5%) at 5.0 min, and kept high flocculation efficiency (above 90.4%) within 15 min (Figure 8), suggesting that ETH-2 has quick and high flocculation ability.
Figure 7. Microscopy imagine of Kaolin clay (a), microscopy imagine of Kaolin clay flocculated by Entercbacter sp. ETH-2 (b), and microscopy imagine of Kaolin clay flocculated by ETH-2 (c), respectively.

Figure 8. Flocculation dynamics of ETH-2.

Table 1 provides the comparison of Extracellular Polymeric Substances (EPS) based bioflocculants yields by different strains. The flocculation efficiency of ETH-2 is higher than the other reported bioflocculants. For instance, 1.3 mg/L ETH-2 could obtain 94.0% flocculation efficiency, adversely, 30.2 mg/L Bacillus megaterium TF10 gained 95.0% flocculation efficiency [26].
Table 1. The comparisons of different bioflocculants.
| Microorganism | Source | Optimized dose (mg/L) | Efficiency | Cation | References |
| Enterobacter aerogenes | Cell-free supernatant of broth | 90 | ND | Zn2+(30 mM) | [4] |
| Enterobacter cloacae WD7 | Cell-free supernatant of broth | 2 | ND | CaCl2(40 mM) | [20] |
| Enterobacter sp. BY-29 | The cell surface | 40 | ND | Fe3+(0.2 mM) | [23] |
| Enterobacter sp. EP3 | Cell-free supernatant of broth | 2 | 96% | CaCl2(8 mM) | [24] |
| Bacillus licheniformis | Cell-free supernatant of broth | 5.8 | >90% | CaCl2(5.14 mM) | [10] |
| Bacillus sp. F19 | Cell-free supernatant of broth | 2 | 97% | No cation | [41] |
| Bacillus mojavensis 32A | Cell-free supernatant of broth | 10 | 92.2% | CaCl2(1.35 mM) | [19] |
| Halomonas sp. V3a’ | Cell-free supernatant of broth | 4 | 96.9% | CaCl2(11.25 mM) | [5] |
| Bacillus circulans | Cell-free supernatant of broth | 2 | 99% | CaCl2(9 mM) | [27] |
| Bacillus firmus | Most (>85%) was in culture broth. Little localized on the cell surface. | 4 | >60* | CaCl2(6.8 mM) | [34] |
| Bacillus megaterium TF10 | The cell surface | 30.2 | 95.5% | CaCl2(5.6 mM) | [26] |
| Entercbacter sp. ETH-2 | The cell surface | 1.3 | 94.0% | No cation | This research |
ND: data not shown.
Flocculation Impact factors
Dosage effect of purified bioflocculant on flocculant efficiency was shown in Figure 9a. Flocculation efficiency of ETH-2 increased from 50.2% to 94.0% with the addition of bioflocculant dose at the range of 0.003 mg to 0.066 mg ETH-2, decreasing to 83.3% with further dose addition to 3.3 mg, indicating 0.066 mg ETH-2 provides optimum flocculation efficiency for 200 mg Kaolin particle. Correspondingly, the maximum adsorption capacity of ETH-2 is calculated as 2849 mg Kaolin particle/mg ETH-2. Similar to other flocculating agents, more or less dose would deteriorate flocculation [27], [34]. Less dose of ETH-2 caused inadequate bioflocculant molecules to adsorb the suspended Kaolin clay particles and to bridge between them. Adversely, more dose of ETH-2 inhibited flocs growth due to the stronger repulsion force between them [18], [26].
Figure 9. Effect of dosage (a), pH (b), cation (c), and temperature (d) on the flocculating efficiency of ETH-2.

Figure 9b illustrates the effect of pH on the reaction system for flocculant efficiency. Flocculation efficiency of ETH-2 kept stable (92.5%–95.2%) at a wide pH range of 3 to 9 and decreased at pH 10(78.9%). Deteriorating flocculation efficiency of ETH-2 with increasing pH possibly results from high alkaline degradation effects on polysaccharide, causing molecular rearrangement of its residue or fragmentation of the polysaccharide chain [35]. The stability of ETH-2 at the range of pH 3-9 was the same as that of the bioflocculant from Enterobacter cloacae WD7 [20], but has wider pH range than that of Gyrodinium impudicum KG03 that was stable at range of pH 3–6 [14].
Figure 9c demonstrates the effect of different cations on flocculant efficiency. ETH-2 flocculation efficiency is not impacted by Na+, K+, Ca2+ and Mg2+, but dramatically influenced by Al3+ and Fe3+, especially by Fe3+, indicating that bioflocculant of ETH-2 is cation independent. Limited cation independent bioflocculants such as Chryseobacterium daeguense W6 [36] and Klebsiella pneumonia [37] have been reported. Cations stimulate flocculation by neutralization and stabilization of residual negative charges of carboxyl groups in polysaccharide by forming bridges between kaolin particles and consequently improving the flocculation ability for most previous reported bioflocculants [23]. For instance, bioflocculant from Enterobacter sp. required Al3+, Fe3+ and Ca2+ for high flocculating activity [23]. Calcium chloride produced a synergistic effect on kaolin flocculation for bioflocculant produced by Enterobacter cloacae WD7 [20] and Enterobacter sp. EP3 [24]. However, trivalent cations possibly alter the surface charge of kaolin particles and cover the adsorb sites [38]. The competition of the positively charged particles and less adsorb sites induce the low flocculating activity, explaining the flocculation activity reduction in the presence of Al3+ and Fe3+.
Figure 9d illustrates thermal stability of purified bioflocculant. With the rising of temperature in the range of 40–100°C, the flocculant efficiency reduced from 91.5% to 80.1%, revealing that ETH-2 is relatively stable at this range. Flocculant activity of bioflocculant with a key component of protein is strongly influenced by high temperature [33]. High temperature can destroy the dimensional structure and cause the denaturation of protein, leading to loss of flocculation activity. Adversely, the flocculation activity of polysaccharides bioflocculant cannot be reduced or less reduced under high temperature [27], [39] due to the heat resistance of polysaccharides. Some thermally stable bioflocculants have been reported, for example, Bacillus licheniformis [10], Bacillus mojavensis 32A [19], and Enterobacter cloacae WD7 [20] keep their flocculation ability below 80°C, 75°C, and 70°C, respectively. ETH-2 bioflocculant had 80.1%flocculation efficiency after being heated for 30 minutes under 100°C, indicating ETH-2 is more thermally stable.
Flocculation mechanism
Flocculation occurs through either bridging or charge neutralization and even by a combination of these two mechanisms [1], [40]. Bridging occurs when bioflocculants extend from the particles' surface into the solution for a distance greater than the distance over which the inter-particle repulsion acts and bioflocculants adsorb other particles to form flocs [1]. Charge neutralization occurs when bioflocculant is oppositely charged compared to the particles. Charge is neutralized and the repulsion between particles is eliminated [1]. The zeta potentials of bioflocculant and kaolin clay suspension were −28.7±8.23 and −35.6±1.66, respectively. If charge neutralization was the main mechanism for the flocculation, flocculation should occur when the zeta potential of the particles is sufficiently low to eliminate repulsion between them. However, the zeta potential of the mixture of ETH-2 and kaolin clay retains a large negative value, −46.9±6.72 mV, suggesting that bridging, instead of the charge neutralization, is probably the flocculation mechanism for ETH-2 bioflocculant.
The bridging mechanism depends on the structure of bioflocculants such as chemicals constitution, functional groups and the molecular weight. Polysaccharides produced by Bacillus megaterium TF10 have along “backbone” with a large number of active sites that adsorb particles through the bridging mechanism [26]. Polysaccharides produced by Proteus mirabilis TJ-1 [3] share the same mechanism. Charge neutralization also acts as the flocculation mechanism for these two bioflocculants. In this study, high MWs polysaccharides in ETH-2 had a long chain structure with small branches. The main backbone of the polysaccharides was composed of mannose, glucose, and galactose. Similar to the structure of polysaccharides produced by Bacillus megaterium TF10 [26], the long backbone of polysaccharides with a large number of active sites is believed to be the main reason for the high flocculation activity of the ETH-2. ETH-2 long backbones simultaneously adsorb many particles through large numbers of active sites, forming large flocs (Figure 10). Unlike bioflocculants produced by Bacillus megaterium TF10 and Proteus mirabilis TJ-1, cation could not improve the flocculation efficiency of ETH-2, indicating that charge neutralization was not the flocculation mechanism. Bridging mechanism then, was concluded as the main flocculation mechanism in this study.
Figure 10. Schematic illustration for the flocculation mechanisms of ETH-2 with Kaolin.

Conclusion
In summary, bioflocculant produced by the newly isolated microorganism Entercbacter sp. ETH-2 has high flocculating activity with unique characters such as cation independence, pH tolerance, and thermal stability. Polysaccharides in the bioflocculant were identified as the active constituents, and the long backbone structure with active sites and high MW were recognized to be responsible for the high flocculation activity through bridging mechanism.
Supporting Information
Combined file of supporting figures and tables. Figure S1, Optimization of carbon and nitrogen resource for strain ETH-2 growth. Table S1, Comparison of bioflocculants extraction methods. Table S2, Apparent molecular weight data from dextran standards and RI detector. Table S3, Binding energies (eV) and assignment/quantization of XPS spectral bands. Materials S1, Carbon and nitrogen source experiments.
(RAR)
Acknowledgments
The author would like to thank Professor YuanJian Xu for his critical suggestions and discussions. We especially thank the anonymous reviewers for offering helpful suggestions that greatly improved the manuscript.
Data Availability
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper and its Supporting Information files.
Funding Statement
We thank the Science and technology Cooperation Project (Science and Technology office of Guizhou Province, The people's Government of Bijie City, and Kunming Institute of Chinese Academy of Sciences) (Grant No. 2013-3) and the National Natural Science Foundation of China (Grant No. 21107091, Grant No. 21377109) for their financial support. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Salehizadeh H, Shojaosadati SA (2001) Extracellular biopolymeric flocculants - Recent trends and biotechnological importance. Biotechnol Adv 19:371–385. [DOI] [PubMed] [Google Scholar]
- 2. Salehizadeh H, Vossoughi M, Alemzadeh I (2000) Some investigations on bioflocculant producing bacteria. Biochem Eng J 5:39–44. [Google Scholar]
- 3. Xia SQ, Zhang ZQ, Wang XJ, Yang A, Chen L, et al. (2008) Production and characterization of a bioflocculant by Proteus mirabilis TJ-1. Bioresource Technol 99:6520–6527. [DOI] [PubMed] [Google Scholar]
- 4. Lu WY, Zhang T, Zhang DY, Li CH, Wen JP, et al. (2005) A novel bioflocculant produced by Enterobacter aerogenes and its use in defecating the trona suspension. Biochem Eng J 27:1–7. [Google Scholar]
- 5. He J, Zou J, Shao ZZ, Zhang JB, Liu ZD, et al. (2010) Characteristics and flocculating mechanism of a novel bioflocculant HBF-3 produced by deep-sea bacterium mutant Halomonas sp V3a'. World J Microb Biot 26:1135–1141. [Google Scholar]
- 6. Liu WJ, Wang K, Li BZ, Yuan HL, Yang JS (2010) Production and characterization of an intracellular bioflocculant by Chryseobacterium daeguense W6 cultured in low nutrition medium. Bioresource Technol 101:1044–1048. [DOI] [PubMed] [Google Scholar]
- 7. Nakata K, Kurane R (1999) Production of an extracellular polysaccharide bioflocculant by Klebsiella pneumoniae . Biosci Biotech Bioch 63:2064–2068. [DOI] [PubMed] [Google Scholar]
- 8. Oh HM, Lee SJ, Park MH, Kim HS, Kim HC, et al. (2001) Harvesting of Chlorella vulgaris using a bioflocculant from Paenibacillus sp AM49. Biotechnol Lett 23:1229–1234. [Google Scholar]
- 9. Wu HJ, Li QB, Lu R, Wang YP, Zhuang XL, et al. (2010) Fed-batch production of a bioflocculant from Corynebacterium glutamicum . J Ind Microbiol Biot 37:1203–1209. [DOI] [PubMed] [Google Scholar]
- 10. Xiong YY, Wang YP, Yu Y, Li QB, Wang HT, et al. (2010) Production and Characterization of a Novel Bioflocculant from Bacillus licheniformis . Appl Environ Microb 76:2778–2782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Aljuboori AHR, Idris A, Abdullah N, Mohamad R (2013) Production and characterization of a bioflocculant produced by Aspergillus flavus . Bioresource Technol 127:489–493. [DOI] [PubMed] [Google Scholar]
- 12. Kwon GS, Moon SH, Hong SD, Lee HM, Kim HS, et al. (1996) A novel flocculant biopolymer produced by Pestalotiopsis sp KCTC 8637P. Biotechnol Lett 18:1459–1464. [Google Scholar]
- 13. Bender J, Rodriguezeaton S, Ekanemesang UM, Phillips P (1994) Characterization of Metal-Binding Bioflocculants Produced by the Cyanobacterial Component of Mixed Microbial Mats. Appl Environ Microb 60:2311–2315. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14. Yim JH, Kim SJ, Ahn SH, Lee HK (2007) Characterization of a novel bioflocculant, p-KG03, from a marine dinoflagellate, Gyrodinium impudicum KG03. Bioresource Technol 98:361–367. [DOI] [PubMed] [Google Scholar]
- 15.Swindell SR, Plasterer TN (1997) SEQMAN: contig assembly. Methods in Molecular Biology 70. [PubMed]
- 16. Song L-Y, Wang Y-Q (2014) Investigation of microbial community structure of a shallow lake after one season copper sulfate algaecide treatment. Microbiological Research. doi:10.1016/j.micres.2014.08.008 [DOI] [PubMed] [Google Scholar]
- 17. Sun M, Li WW, Yu HQ, Harada H (2012) A novel integrated approach to quantitatively evaluate the efficiency of extracellular polymeric substances (EPS) extraction process. Appl Microbiol Biot 96:1577–1585. [DOI] [PubMed] [Google Scholar]
- 18. Zhang ZQ, Xia SQ, Zhang JA (2010) Enhanced dewatering of waste sludge with microbial flocculant TJ-F1 as a novel conditioner. Water Res 44:3087–3092. [DOI] [PubMed] [Google Scholar]
- 19. Elkady MF, Farag S, Zaki S, Abu-Elreesh G, Abd-El-Haleemn D (2011) Bacillus mojavensis strain 32A, a bioflocculant-producing bacterium isolated from an Egyptian salt production pond. Bioresource Technol 102:8143–8151. [DOI] [PubMed] [Google Scholar]
- 20. Prasertsan P, Dermlim W, Doelle H, Kennedy JF (2006) Screening, characterization and flocculating property of carbohydrate polymer from newly isolated Enterobacter cloacae WD7. Carbohyd Polym 66:289–297. [Google Scholar]
- 21. Kurane R, Matsuyama H (1994) Production of a Bioflocculant by Mixed Culture. Biosci Biotech Bioch 58:1589–1594. [DOI] [PubMed] [Google Scholar]
- 22. Toeda K, Kurane R (1991) Microbial Flocculant from Alcaligenes Cupidus Kt201. Agr Biol Chem Tokyo 55:2793–2799. [Google Scholar]
- 23. Yokoi H, Yoshida T, Mori S, Hirose J, Hayashi S, et al. (1997) Biopolymer flocculant produced by an Enterobacter sp. Biotechnol Lett 19:569–573. [Google Scholar]
- 24. Liu CS, Guo YD, Zhao DF (2012) Characteristics and Flocculating Mechanism of a Bioflocculant M-1 Produced by Enterobacter sp EP3. Adv Mater Res-Switz 550–553:1482–1485. [Google Scholar]
- 25. Michaels A, Morelos O (1955) Polyelectrolyte adsorption by kaolinite. Industrial & Engineering Chemistry 47:1801–1809. [Google Scholar]
- 26. Yuan SJ, Sun M, Sheng GP, Li Y, Li WW, et al. (2011) Identification of Key Constituents and Structure of the Extracellular Polymeric Substances Excreted by Bacillus megaterium TF10 for Their Flocculation Capacity. Environ Sci Technol 45:1152–1157. [DOI] [PubMed] [Google Scholar]
- 27. Li Z, Chen RW, Lei HY, Shan Z, Bai T, et al. (2009) Characterization and flocculating properties of a novel bioflocculant produced by Bacillus circulans . World J Microb Biot 25:745–752. [Google Scholar]
- 28. Li Z, Zhong S, Lei HY, Chen RW, Yu Q, et al. (2009) Production of a novel bioflocculant by Bacillus licheniformis X14 and its application to low temperature drinking water treatment. Bioresource Technol 100:3650–3656. [DOI] [PubMed] [Google Scholar]
- 29. Mabinya LV, Cosa S, Nwodo U, Okoh AI (2012) Studies on Bioflocculant Production by Arthrobacter sp Raats, a Freshwater Bacteria Isolated from Tyume River, South Africa. Int J Mol Sci 13:1054–1065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Kurane R, Hatamochi K, Kakuno T, Kiyohara M, Tajima T, et al. (1995) Chemical-Structure of Lipid Bioflocculant Produced by Rhodococcus Erythropolis . Biosci Biotech Bioch 59:1652–1656. [Google Scholar]
- 31. Zajic JE, Leduy A (1973) Flocculant and Chemical Properties of a Polysaccharide from Pullularia Pullulans . Appl Microbiol 25:628–635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32. Yokoi H, Yoshida T, Hirose J, Hayashi S, Takasaki Y (1998) Biopolymer flocculant produced by an Pseudomonas sp. Biotechnol Tech 12:511–514. [Google Scholar]
- 33. Takeda M, Koizumi J, Matsuoka H, Hikuma M (1992) Factors Affecting the Activity of a Protein Bioflocculant Produced by Nocardia Amarae . J Ferment Bioeng 74:408–409. [Google Scholar]
- 34. Salehizadeh H, Shojaosadati SA (2002) Isolation and characterisation of a bioflocculant produced by Bacillus firmus . Biotechnol Lett 24:35–40. [Google Scholar]
- 35. Aspinall GO (1982) Chemical characterization and structure determination of polysaccharides. The polysaccharides 1:35–131. [Google Scholar]
- 36. Liu W, Wang K, Li B, Yuan H, Yang J (2010) Production and characterization of an intracellular bioflocculant by Chryseobacterium daeguense W6 cultured in low nutrition medium. Bioresource Technol 101:1044–1048. [DOI] [PubMed] [Google Scholar]
- 37. Zhao H, Liu H, Zhou J (2013) Characterization of a bioflocculant MBF-5 by Klebsiella pneumoniae and its application in Acanthamoeba cysts removal. Bioresource Technol 137:226–232. [DOI] [PubMed] [Google Scholar]
- 38. Gong W-X, Wang S-G, Sun X-F, Liu X-W, Yue Q-Y, et al. (2008) Bioflocculant production by culture of Serratia ficaria and its application in wastewater treatment. Bioresource Technol 99:4668–4674. [DOI] [PubMed] [Google Scholar]
- 39. Wang LL, Ma F, Qu YY, Sun DZ, Li A, et al. (2011) Characterization of a compound bioflocculant produced by mixed culture of Rhizobium radiobacter F2 and Bacillus sphaeicus F6. World J Microb Biot 27:2559–2565. [Google Scholar]
- 40. Ries HE, Meyers BL (1968) Flocculation mechanism: charge neutralization and bridging. Science 160:1449–1450. [DOI] [PubMed] [Google Scholar]
- 41. Zheng Y, Ye ZL, Fang XL, Li YH, Cai WM (2008) Production and characteristics of a bioflocculant produced by Bacillus sp F19. Bioresource Technol 99:7686–7691. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Combined file of supporting figures and tables. Figure S1, Optimization of carbon and nitrogen resource for strain ETH-2 growth. Table S1, Comparison of bioflocculants extraction methods. Table S2, Apparent molecular weight data from dextran standards and RI detector. Table S3, Binding energies (eV) and assignment/quantization of XPS spectral bands. Materials S1, Carbon and nitrogen source experiments.
(RAR)
Data Availability Statement
The authors confirm that all data underlying the findings are fully available without restriction. All relevant data are within the paper and its Supporting Information files.