

Significance
Sea stars inhabiting the Northeast Pacific Coast have recently experienced an extensive outbreak of wasting disease, leading to their degradation and disappearance from many coastal areas. In this paper, we present evidence that the cause of the disease is transmissible from disease-affected animals to apparently healthy individuals, that the disease-causing agent is a virus-sized microorganism, and that the best candidate viral taxon, the sea star-associated densovirus (SSaDV), is in greater abundance in diseased than in healthy sea stars.
Keywords: virus, Asteroidea, disease, densovirus, wasting
Abstract
Populations of at least 20 asteroid species on the Northeast Pacific Coast have recently experienced an extensive outbreak of sea-star (asteroid) wasting disease (SSWD). The disease leads to behavioral changes, lesions, loss of turgor, limb autotomy, and death characterized by rapid degradation (“melting”). Here, we present evidence from experimental challenge studies and field observations that link the mass mortalities to a densovirus (Parvoviridae). Virus-sized material (i.e., <0.2 μm) from symptomatic tissues that was inoculated into asymptomatic asteroids consistently resulted in SSWD signs whereas animals receiving heat-killed (i.e., control) virus-sized inoculum remained asymptomatic. Viral metagenomic investigations revealed the sea star-associated densovirus (SSaDV) as the most likely candidate virus associated with tissues from symptomatic asteroids. Quantification of SSaDV during transmission trials indicated that progression of SSWD paralleled increased SSaDV load. In field surveys, SSaDV loads were more abundant in symptomatic than in asymptomatic asteroids. SSaDV could be detected in plankton, sediments and in nonasteroid echinoderms, providing a possible mechanism for viral spread. SSaDV was detected in museum specimens of asteroids from 1942, suggesting that it has been present on the North American Pacific Coast for at least 72 y. SSaDV is therefore the most promising candidate disease agent responsible for asteroid mass mortality.
Since June 2013, millions of sea stars (asteroids) of the west coast of North America have wasted away into slime and ossicle piles, due to a disease known as sea-star wasting disease (SSWD). SSWD has been used to collectively describe die-offs of sea stars in the Northeast Pacific since at least 1979; however, this SSWD event differs from other asteroid mass mortalities (1–5) due to its broad geographic extent (from Baja California, Mexico to Southern Alaska; pacificrockyintertidal.org) and many (n = 20) species affected, representing several major lineages of Asteroidea (Fig. 1, Table S1, and SI Text). The extensive geographic range and number of species infected might make SSWD the largest known marine wildlife epizootic to date. Outward signs of SSWD vary slightly among species but generally start with behavioral changes, including lethargy and limb curling, followed by lesions, ray autotomy, turgor loss (deflation), and end with animal death (Fig. 1). Histology of dead and dying asteroids from geographically widespread natural habitats and aquaria, showed epidermal necrosis and ulceration, and dermal inflammation and edema in the body wall. Clinically affected (i.e., symptomatic) individuals rarely recover in the laboratory and only occasionally in the field.
Fig. 1.

Photographs of SSWD-affected stars (A) asymptomatic P. helianthoides, (B) symptomatic P. helianthoides, and (C) symptomatic P. ochraceus. Disease symptoms are consistent with loss of turgor, loss of rays, formation of lesions, and animal decomposition. (D) Map showing occurrence of SSWD based on first reported observation. (E) Transmission electron micrograph of negatively stained (uranyl acetate) viruses extracted from an affected wild E. troschelii from Vancouver . The sample contained 20–25-nm diameter nonenveloped icosohedral viral particles on a background of cellular debris (primarily ribosomal subunits) and degraded viral particles of similar morphology. (Scale bar: 100 nm.)
The cause of SSWD remains a mystery. Scientific hypotheses given for other asteroid mortality events include storms (6–11), temperature anomalies (1, 3, 12), starvation (13), and infection by unidentified pathogens (5). For instance, pathogens in the bacterial genus Vibrio (12, 14, 15) and an unidentified eukaryotic parasite (4) were seen in die-offs of the tropical asteroid Acanthaster planci and the Mediterranean asteroid Astropecten jonstoni. However, it is difficult to distinguish the cause of an infectious disease from the associated microbial community that can flourish in a sick or injured animal.
Some early patterns from the field supported the hypothesis that SSWD is contagious. Within a region, SSWD has sometimes moved from site to site similar to an infectious disease. For example, the disease spread north to south in Southern California. All of the major aquaria on the North American Pacific Coast were affected by SSWD in Fall 2013, with mass mortality of captive asteroids. However, SSWD did not spread into aquaria that sterilize inflowing seawater with UV light but did spread into nontreated aquaria. Furthermore, the incursion of SSWD in aquaria with sand-filtered intake suggests a microscopic infectious stage, rather than a disease that spread only via contact with an infected host or vector. Overall, these patterns suggest a microscopic, water-borne, infectious disease agent, rather than environmental pollutants.
Inoculation with Virus-Sized Homogenates Induces Wasting-Disease Signs
We turned to viruses as potential infectious agents in SSWD because histologic evaluation of hundreds of slides indicated no evidence of prokaryotic or eukaryotic microbial infection within tissues of asteroids affected by SSWD. Asymptomatic adult Pycnopodia helianthoides were collected from several sites in Puget Sound, Washington, in January–April 2014 before disease occurrence at the collection sites. Animals were placed in separate 10-gallon glass aquaria with sand-filtered and UV-treated flow-through seawater on separate draw lines (i.e., seawater was not recirculated between aquaria; US Geological Survey Marrowstone Laboratory) and held at ambient temperature (8–9 °C). Viral challenges were performed with homogenates prepared from symptomatic animals. Tissues from symptomatic asteroids were homogenized in 10 mL of seawater from the aquarium housing the symptomatic animal. The tissue homogenate was initially centrifuged at 1,000 × g for 5 min at 4 °C to remove large debris before being split into two 5-mL aliquots. One aliquot was boiled for 7 min to serve as a control before both heat-treated (control) and non–heat-treated (virus exposed) homogenates were serially syringe-filtered through 1-μm pore size polycarbonate and 0.22-μm pore size polyethersulfone filters, resulting in a virus-sized inoculum. Each asteroid was injected with 0.5 mL of either control (i.e., heat-treated) or viral inocula (0.5 mL) into the coelomic cavity. After injection, animals were observed every morning and evening for SSWD signs.
The animals inoculated with virus-sized material showed disease signs after 10–17 d whereas control (heat-treated inoculum) individuals did not develop SSWD signs (Fig. 2). From one of the virus-exposed animals showing SSWD signs in the first challenge, we again isolated virus-sized material to challenge a new set of asymptomatic animals. This second set of inoculated individuals had disease signs after 8–17 d (and again no control animals that received heat-treated inoculum developed SSWD signs). These experiments demonstrate that the disease is transmissible from symptomatic to asymptomatic individuals and that the pathogenic agent is virus-sized (i.e., <200 nm in diameter).
Fig. 2.

(A) Proportion of stars remaining asymptomatic after inoculation with control (heat-killed) or virus-sized fraction (VSF) of asteroid homogenates in first (Expt 1) and second (Expt 2) challenge. Survival analysis (20) indicates that the time to lesion development differs among treatment and control groups (log-rank test: χ2 = 18.6, df = 1, P = <0.0001), but there is no difference in time to development of lesions in the two experiments (log-rank test: χ2 = 0.2, df = 1, P = 0.698) (B) Change in SSaDV load between initiation of viral challenge and termination of experiment (i.e., animal expiry in live challenge or euthanasia of control animals). Note difference in scale. Viral load was determined by quantitative PCR (qPCR) targeting the NS1 gene of the SSaDV genome. Different letters above bars indicate that the mean viral abundance change is significantly different (P < 0.05, two-tailed Wilcoxon signed-rank test to account for heteroskedasticity; Fisher’s F-test P > 0.05). Error bars = SE.
Investigation of Viral Agents in SSWD-Affected Tissues
We sought to identify candidate viruses in inoculum by conducting metagenomic surveys to compare viral consortia inhabiting asymptomatic and SSWD-affected tissues. Tissue samples (∼1-cm cross-section of a ray including body wall, epidermis, pyloric caeca, and gonad) were excised from 28 specimens (Tables S2–S5) with sterilized dissecting scissors and placed into sterile 50-mL plastic tubes. Viral metagenomes (i.e., metaviromes) were then prepared and analyzed following Gudenkauf et al. (16) (SI Text). All sequences have been deposited in GenBank under BioProject accession no. PRJNA253121.
Metaviromic analysis indicated a greater prevalence of parvovirus-like sequences in symptomatic relative to asymptomatic asteroid homogenates (in 2 of 13 asymptomatic libraries and 7 of 15 symptomatic libraries). Global assembly of all viral metagenomes (n = 28 libraries, representing 5 × 107 sequences and 1 × 1010 nt of information) resulted in assembly of a near-complete densovirus (Parvoviridae) genome, which we have named sea star-associated densovirus (SSaDV) (Fig. 3). The SSaDV genome fragment bore architectural features similar to insect densoviruses but lacked their characteristic palindromic repeats. However, SSaDV is related to densoviruses in the Hawaiian sea urchins (Echinoidea) Colobocentrotus atratus, Echinometra mathaei, and Tripneustes gratilla (16) (Fig. 4), placing it near the only other known viruses of echinoderms. Purified preparations from three Northeastern Pacific asteroids (Evasterias troschelii, P. helianthoides, and Pisaster ochraceus) that had wide SSaDV representation in metagenomic libraries all contained nonenveloped icosohedral viral particles ∼25 nm in size when negatively stained with uranyl acetate and viewed by transmission electron microscopy (TEM). The ultrastructure of these particles seems similar to other known viruses in the family Parvoviridae (Fig. 1). Comparison of viral metagenomes from symptomatic and asymptomatic asteroids did not reveal any other candidate viruses in tissue homogenates (see SI Text for details), confirming SSaDV as the sole virus associated with SSWD (Fig. S1).
Fig. 3.
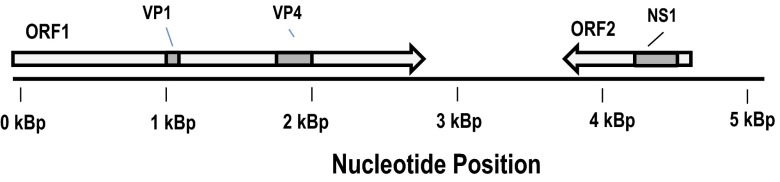
Genome architecture of the sea star-associated densovirus (SSaDV).
Fig. 4.

Phylogenetic representation of the sea star-associated densovirus (SSaDV) NS1 capsid protein. The phylogenetic tree is based on an amino acid alignment performed by MUSCLE. The tree was constructed based on maximum-likelihood distance.
To determine presence and load across a wider suite of individuals, quantitative PCR (TaqMan) primers were designed using Primer3 (4) specific to the nonstructural protein 1 (NS1) and viral gene product 4 (VP4) of SSaDV (SI Text). Quantitative PCR (qPCR) was performed following the approach of Hewson et al. (17), where template material comprised DNA extracted from small excisions of body wall or tube feet and where SSaDV copy number was divided by weight of tissue extracted.
In the experimental challenges with a virus-sized inoculum that elicited SSWD clinical signs, SSaDV load increased as disease signs appeared. In contrast, control asteroids receiving heat-killed virus-sized inoculum showed no SSWD signs, and SSaDV loads decreased over time. We attribute the initial low levels of SSaDV in the heat-treated qPCR to detection of heat-killed viral DNA that decayed after heat treatment (Fig. 2B). The inoculation experiments suggest that SSaDV is transmissible and can lead to wasting disease in exposed sea stars.
SSaDV Is Linked to Wasting Disease in Field Surveys
Due to the association of SSaDV with diseased asteroid tissues, we examined the incidence of SSaDV among symptomatic (n = 286 individuals) and asymptomatic (n = 49 individuals) asteroids of 14 species. Viral load (number of SSaDV copies detected per mg of tissue) and prevalence (i.e., percentage of samples where SSaDV was detected) were higher in symptomatic than in the asymptomatic animals in all three species where both symptomatic and asymptomatic animals were obtained (Fig. 5). However, the virus was present in both asymptomatic and symptomatic individuals in species where animals in both health states were sampled, including P. ochraceus, P. helianthoides, and E. troschelii. Because SSaDV detection varied by tissue type and location on each animal (Fig. S2), the single tissue sample taken from each individual likely led to some false negatives (in repeated sampling of body-wall tissues from symptomatic P. ochraceus, SSaDV was detected in 11–38% of samples). Due to the potential for these false negatives, it was not surprising that we observed SSaDV in some asymptomatic asteroids. Conversely, SSaDV in asymptomatic animals almost certainly represents viral presence before disease signs develop because we know from our inoculation experiment that signs can take 2 wk to progress after inoculation (or could represent viruses present on animal surfaces that had not yet gained entry to animal tissues).
Fig. 5.

(A) Mean viral load and (B) prevalence (i.e., proportion of SSaDV-positive individuals) (B) as determined by qPCR targeting the VP4 gene of the SSaDV genome. qPCR was applied to P. helianthoides (n = 10 asymptomatic and 79 symptomatic), P. ochraceus (n = 26 asymptomatic and 72 symptomatic), and E. troschelii (n = 6 asymptomatic and 31 symptomatic) whole-tissue DNA extracts. (C) SSaDV load in sympatric asteroid species. All stars except Dermasterias imbricata, Orthasterias sp., and Astropecten polyacanthus were symptomatic. The number of individuals tested is indicated for each species. Different letters within bars represent significant difference in the percentage of viral reads between asymptomatic and symptomatic asteroids (P < 0.001; df = 113 for P. helianthoides, df = 42 for E. troschelii, and df = 117 for P. ochraceus; data log-transformed and corrected for heteroskedasticity by x/√x; t test). The probability of being infected with the virus was higher in symptomatic asteroids. Logistic regression (generalized linear model with binomial distribution and logit link) comparing models with species and disease status and their interaction indicated that a model including species (likelihood ratio test: χ = 19.7, df = 2; P < 0.0001) and disease status (likelihood ratio test: χ =7.4, df = 1; P = 0.0065) additively had the greatest support. This result indicates that viral prevalence differs among species and disease status, but the difference among disease status does not differ significantly among species (likelihood ratio test: χ = 2.50, df = 1; P = 0.29). The odds ratios suggest that symptomatic stars are 3.2 times more likely to be virus-positive than asymptomatic stars. Error bars = SE.
Despite these procedural challenges, asteroids were more likely to be diseased if they had a high viral load (Fig. 6). In our statistical models involving viral load, we started with all factors and their first-order interaction terms. To focus our interpretation of model effects and increase power, we sequentially removed interaction terms that were not statistically significant (in order of their associated P value) and then did the same for main effects (this removal generally followed Akeike information criterion model selection). For the relationship between SSaDV load and disease, we used a logistic model of symptomatic vs. asymptomatic to evaluate the potential independent effects of SSaDV load, asteroid species, geography (San Diegan province or south of Point Conception vs. Oregonian province or north of Point Conception), and asteroid size (measured as arm circumference) in a sample of 107 symptomatic and asymptomatic P. ochraceus, P. helianthoides, and E. troschelii for which we had size measurements. The main significant predictive variable for being symptomatic was the SSaDV load [logistic regression, square root-transformed load of SSaDV, estimate = 0.0013 (0.0008 SE) chance of being symptomatic increasing with viral load, P = 0.006]. In addition, for a given viral load, asteroids from southern sites were more likely to be symptomatic [logistic regression, estimate = 0.95 (0.53 SE), P = 0.03] (Fig. 6). This result was consistent with analyses limited to P. ochraceus, which was the only species sampled in the North and South. However, neither asteroid size nor species had significant independent or interactive effects with the other factors. Given that viral load was the main predictor of disease, we then asked what factors (species, geography, size) predicted viral load in 106 infected asteroids (from five species). A general linear model found a significant interaction between species and size (P = 0.01) due to a negative association between viral load and size in P. helianthoides (small individuals of P. helianthoides were more likely to have high viral counts), which was absent in the other four asteroid species. Therefore, our results provide strong evidence for a link between SSaDV and disease in wild asteroids.
Fig. 6.

Square root-transformed viral load in asymptomatic and symptomatic asteroids in San Diegan (i.e., south of Point Conception) and Oregonian (i.e., north of Point Conception) biogeographical provinces. For the relationship between SSaDV abundance and disease, we used a logistic model of symptomatic vs. asymptomatic to evaluate the potential independent effects of SSaDV abundance, sea star species, geography (San Diegan vs. Oregonian province), and sea-star size (measured as arm circumference) in the 107 P. ochraceus, P. helianthoides, and E. troschelii for which we had both size measurements and a mix of asymptomatic and symptomatic stars. The main significant predictive variable for being symptomatic was the abundance of SSaDV [logistic regression, square root-transformed count of SSaDV, estimate = 0.0013 (0.0008 SE) chance of being symptomatic increasing with viral count, P = 0.006]. Error bars = SE.
To confirm the presence of viable SSaDV in host tissues, we performed quantitative reverse transcriptase PCR (qRT-PCR) on RNA extracted from symptomatic and asymptomatic individuals. SSaDV RNA, a proxy for viral presence and replication, was detected only in symptomatic P. helianthoides and E. troschelii and not in asymptomatic animals. However, SSaDV gene expression was observed in both symptomatic and asymptomatic P. ochraceus, suggesting that the host parasite dynamics may differ among species (Fig. 7).
Fig. 7.

Transcription of the SSaDV VP4 as assessed by qRT-PCR comparing asymptomatic and symptomatic tissues. qRT-PCR was performed on whole-tissue RNA extracts from E. troschelii (n = 5 asymptomatic and 5 symptomatic), P. ochraceus (n = 5 asymptomatic and 6 symptomatic), and P. helianthoides (n = 10 asymptomatic and 10 symptomatic), and normalized to quantities of SSaDV assessed by qPCR in cDNA extracts. Transcript levels were significantly higher in asymptomatic P. ochraceus than symptomatic individuals (Mann–Whitney U test; P = 0.039, df = 11). Error bars = SE.
SSaDV Present in Plankton, Sediments, and Nonasteroid Echinoderms
Given evidence for viral transmission between asteroids in the laboratory, we then sought to understand how SSaDV might move between wild host individuals and populations. To investigate whether uninfected asteroids could contact viruses free in the water, associated with suspended particles, or in sediments, we tested different environmental samples for SSaDV. SSaDV had its highest abundance in the virioplankton size fraction of the water (0.02–0.2 µm) and was present in the suspended particulate material size fraction > 0.2 µm (Fig. 8). SSaDV presence in particulate material is congruent with other observations of parvoviruses (16) and might represent viruses adsorbed to abiotic material, viruses in detrital particles from decayed animals, or viruses within larval asteroids. SSaDV was also found in sediments collected from public aquaria that had experienced SSWD several months earlier, and SSaDV was concentrated in sand filters used to treat incoming water and between aquaria (Fig. 9). Therefore, SSaDV might transmit between asteroids and among populations by mechanisms other than direct contact between diseased and healthy individuals, consistent with the observation that SSaDV-infected asteroids shed virus into the water column (Fig. S3). Water-column SSaDV transport helps explain how SSWD spreads among disjunct asteroid populations.
Fig. 8.

Viral abundance in particle (i.e., >0.2 µm) and virioplankton (0.2–0.02 µm) size fractions of water collected at field sites, experimental incubations, and public aquaria. Viral abundance was determined by qPCR targeting the VP4 gene of the SSWDAV genome. Means were not significantly different.
Fig. 9.

Viral abundance in sediments from aquaria and field sites and in aquarium sand filters, as determined by qPCR targeting the VP4 gene of the SSaDV genome.
The SSWD epizootic has hit an alarming number of asteroid species in all shallow water habitats. Eight of 11 asteroid species sampled from SSWD areas contained SSaDV (Fig. 4). The broad range of species in which SSaDV was detected is unexpected because most viruses infect a narrow range of host species. However, parvoviruses are known to infect across families, and variations in capsid protein secondary structure can result in variable host range (18). Parvoviruses gain entry to host cells via transferrin receptors, which are among the most highly expressed proteins in coelomic fluid (where, notably, it is found in coelomocytes, which are a major defense mechanism against cellular microbial infection) (19). It is also possible that receptors are shared between closely related sea-star species. This phenomenon is especially a possibility here because six of the eight species known to be affected by SSWD in which SSaDV was detected are within a single asteroid family (Asteriidae; the remaining two were within the Asterinidae) that may share common cell-surface features through which viruses may infect.
This broad host range inspired us to look for SSaDV in other Northeastern Pacific echinoderms. We observed SSaDV in sympatric, nonasteroid echinoderms, including echinoids (Strongylocentrotus purpuratus and Dendraster excentricus) and ophiuroids (Fig. S4). The impact of SSaDV on these taxa is unknown; however, the presence of the virus suggests that they could form a reservoir of SSaDV. If some echinoderms are tolerant reservoirs of infection, it might help keep SSaDV in the system long after it extirpates less tolerant hosts, helping explain how SSWD can extirpate some host species while still persisting in an ecosystem. However, we cannot eliminate the possibility that detection of SSaDV in these species may represent free viruses attached to their surfaces or drawn into their water vascular systems, as opposed to those infecting tissues.
SSaDV Present in Asteroids 72 Years Ago
Its broad host range suggested that SSaDV could be associated with asteroid mortalities in other times and places. To better understand the geographic distribution of SSaDV, we studied Northwest Atlantic Coast asteroids with SSWD-like signs in 2012 and 2013. We used qPCR targeting two loci (VP4 and NS1) on the SSaDV genome to detect SSaDV from diseased Asterias forbesii from the Mystic Aquarium (Connecticut). Although we did not detect SSaDV gene transcription, the SSaDV DNA detected in 9 of 14 samples suggests that this virus might be present in other oceanic basins. Additionally, to investigate whether SSaDV was present in Northeast Pacific Coast asteroids before 2013, we tested for SSaDV in ethanol-preserved museum specimens collected between 1923 and 2010 at sites in British Columbia, Washington, Oregon, and California (Table S6). qPCR amplification detected SSaDV DNA (NS1 and VP4 loci) in asteroids that had been field-collected and preserved in July 1942, October 1980, September 1987, and July 1991 (Fig. 10). We also detected one locus in at least nine other individual asteroids, suggesting that viruses with homologous NS1 or VP4 were present in these populations, too. Therefore, SSaDV and related viruses might have infected asteroids on the North American West Coast decades before the current SSWD event.
Fig. 10.

Detection and load of SSaDV in ethanol-preserved museum specimens from 1923 to the present. SSaDV load was assessed by qPCR targeting the VP4 gene on the SSaDV genome and normalized to extracted tissue weight. We targeted both NS1 (□) and VP4 (◇) for this analysis because homologous viruses and recombination may have led to spurious results in old asteroids. Both NS1 and VP4 were found in 6 (of 67 tested) specimens from 1942, 1980, 1987, and 1991. We also detected large loads of either NS1 or VP4 in 16 asteroids. These results suggest that SSaDV or perhaps related densoviruses have been present in populations of several Northeastern Pacific Coast asteroid species, at least since 1942.
If SSaDV is the cause of the current SSWD event, it is unclear why the virus did not elicit wide disease outbreaks in the past during periods in which it was detected; however, there are several possible reasons why the current SSWD event is broader and more intense than previous occurrences. SSaDV may have been present at lower prevalence for decades and only became an epidemic recently due to unmeasured environmental factors not present in previous years that affect animal susceptibility or enhance transmission. There are anecdotal reports from fishers and scuba divers that by 2012, the Salish Sea was severely overpopulated with adult P. helianthoides. Our finding of a strong relationship between size and SSaDV load, and anecdotal observations of SSWD commonly in adults but less so in juveniles in the field, suggest that the current event may be exacerbated by a large number of adult sea stars present in small bays and inlets. Because of its wide host range in the current event, we also speculate that variation of SSaDV (possibly by modification of capsid structure, as seen in other parvoviruses) (18) may have led to greater virulence. There remains much to be learned about the interactive effects of environmental transport, virulence, and environment on the dynamics of this disease.
Conclusions
In summary, SSWD has caused widespread and, until now, unexplained mass mortality in asteroids. SSWD spread has been most consistent with an infectious agent, which we suggest is a virus. Based on our observations, the densovirus, SSaDV, is the most likely virus involved in this disease. We base this statement on finding virus and disease transmission to healthy asteroids after exposure in two trials with virus-sized particles from diseased asteroids, finding replicating densovirus in diseased tissue, and an association between viral load and disease. Furthermore, our observation of SSaDV from 72 y ago suggests that, like many marine pathogens, SSaDV was already present in the environment before the outbreak. The detection of SSaDV in diverse echinoderm species and some sediments suggests a high potential for persistence in nonasteroid reservoirs. SSaDV is present in environmental samples, suggesting that it can spread outside of a host. However, it remains to be seen how infection with SSaDV kills asteroids, what the role is for other microbial agents associated with dying asteroids, what triggers outbreaks, and how asteroid mass mortalities will alter near-shore communities throughout the North American Pacific Coast. More generally, viral pathogens are poorly known for all noncommercial invertebrates yet may play an unrecognized, yet important, role in marine ecosystems.
Supplementary Material
Acknowledgments
We thank Alice Nguyen, Armand Kuris, and Gretchen Hoffman [University of California (UC) Santa Barbara]; Elise Delaroque (UC Davis); Betsy Steele, Nathaniel Fletcher, Rani Gaddam, Melissa Redfield, Corianna Hume-Flannery, Sarah Sampson, Colin Gaylord, and Tristin McHugh (UC Santa Cruz); Steven Whittaker (National Parks Service); Stewart Johnson (Fisheries and Oceans Canada); Michael Garner (Northwest Zoopathology); Marina Krasnovid and Bryanda Wippel (University of Washington); Gene McKeen, Nate Schwarck, Jay Dimond, and Robert Boenish (Shannon Point Marine Center); Paul Hershberger [US Geological Survey (USGS) Marrowstone Island Laboratory]; Thierry Work (USGS); Sion Cahoon (Vancouver Aquarium); Kim Rotter (California Science Center); Allison Tuttle (Mystic Aquarium); Douglas Swanston and Neil McDaniel (Seacology Network); Shawn Larson and Bryan McNeil (Seattle Aquarium); Salvatore Frasca, Jr. (University of Connecticut); James Eaglesham, Sierra Helmann, Jacob Sangren (Cornell University); Rebecca Thurber (Oregon State University); and Laura James and Jan Kocian. Photographs in Fig. 1 B and C were provided by N. McDaniel. Research was supported by rapid response grants from the David R. Atkinson Center for a Sustainable Future (Cornell University), National Science Foundation Grant OCE-1401727 (to I.H. and B.M.), and Washington Sea Grant (to C.M.M. and B.M.).
Footnotes
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Data deposition: The sequence reported in this paper has been deposited in the GenBank database (accession no. PRJNA253121).
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1416625111/-/DCSupplemental.
References
- 1.Dungan ML, Miller TE, Thomson DA. Catastrophic decline of a top carnivore in the Gulf of California rocky intertidal zone. Science. 1982;216(4549):989–991. doi: 10.1126/science.216.4549.989. [DOI] [PubMed] [Google Scholar]
- 2.Eckert G, Engle JM, Kushner D. 1999. Sea star disease and population declines at the Channel Islands. Proceedings of the Fifth California Islands Symposium (Minerals Management Service, Washington, DC), pp 390–393.
- 3.Bates AE, Hilton BJ, Harley CDG. Effects of temperature, season and locality on wasting disease in the keystone predatory sea star Pisaster ochraceus. Dis Aquat Organ. 2009;86(3):245–251. doi: 10.3354/dao02125. [DOI] [PubMed] [Google Scholar]
- 4.Zann L, Brodie J, Vuki V. History and dynamics of the crown-of-thorns starfish Acanthaster planci (L) in the Suva Area, Fiji. Coral Reefs. 1990;9:135–144. [Google Scholar]
- 5.Pratchett MS. An infectious disease in crown-of-thorns starfish on the Great Barrier Reef. Coral Reefs. 1999;18:272. [Google Scholar]
- 6.Tiffany WJ. Mass mortality of Luidia senegalensis (Lamarck, 1816) on Captiva Island, Florida, with a note on its occurrence in Florida Gulf coastal waters. Fla Sci. 1978;41:63–64. [Google Scholar]
- 7.Sieling FW. Mass mortality of the starfish, Asterias forbesi, on the Atlantic Coast of Maryland. Chesap Sci. 1960;1:73–74. [Google Scholar]
- 8.Lawrence JM. Mass mortality of echinoderms from abiotic factors. In: Jangoux M, Lawrence JM, editors. Echinoderm Studies. Vol 5. A.A. Balkema; Rotterdam: 1996. pp. 103–137. [Google Scholar]
- 9.Thorpe JP, Spencer EL. A mass stranding of the asteroid Asterias rubens on the Isle of Man. J Mar Biol Assoc U K. 2000;80:749–750. [Google Scholar]
- 10.Berger VY, Naumov AD. Effects of salinity on the substrate attachability of the starfish Asterias rubrens. Biologiya Morya. 1996;22:99–101. [Google Scholar]
- 11.Scheibling RE, Hennigar AW. Recurrent outbreaks of disease in sea urchins Strongylocentrotus droebachiensis in Nova Scotia: Evidence for a link with large-scale meteorologic and oceanographic events. Mar Ecol Prog Ser. 1997;152:155–165. [Google Scholar]
- 12.Staehli A, Schaerer R, Hoelzle K, Ribi G. Temperature induced disease in the starfish Astropecten jonstoni. Mar Biodivers Rec. 2008;2:e78. [Google Scholar]
- 13.Suzuki G, Kai S, Yamashita H. Mass stranding of crown-of-thorns starfish. Coral Reefs. 2012;31:821. [Google Scholar]
- 14.Sutton DC, Trott L, Reichelt JL, Lucas JS. 1988. Assessment of bacterial pathogenesis in crown-of-thorns starfish, Acanthaster planci. Proceedings of the Sixth International Coral Reef Symposium, eds Choat JH, Barnes D, Borowitzka MA, Coll JC, Davies PJ, Flood P, Hatcher BG, Hopley D, Hutchings PA, Kinsey D, Orme GR, Pichon M, Sale PF, Sammarco P, Wallace CC, Wilkinson C, Wolanski E, Bellwood O (Australian Museum, Townsville, Australia), Vol. 2, pp. 171–176, contributed papers.
- 15.Rivera-Posada JA, Pratchett M, Cano-Gomez A, Arango-Gomez JD, Owens L. Refined identification of Vibrio bacterial flora from Acanthasther planci based on biochemical profiling and analysis of housekeeping genes. Dis Aquat Organ. 2011;96(2):113–123. doi: 10.3354/dao02389. [DOI] [PubMed] [Google Scholar]
- 16.Gudenkauf BM, Eaglesham JB, Aragundi WM, Hewson I. Discovery of urchin-associated densoviruses (family Parvoviridae) in coastal waters of the Big Island, Hawaii. J Gen Virol. 2014;95(Pt 3):652–658. doi: 10.1099/vir.0.060780-0. [DOI] [PubMed] [Google Scholar]
- 17.Hewson I, et al. Metagenomic identification, seasonal dynamics and potential transmission mechanisms of a Daphnia- associated single-stranded DNA virus in two temperate lakes. Limnol Oceanogr. 2013;58:1605–1620. [Google Scholar]
- 18.Parker JS, Parrish CR. Canine parvovirus host range is determined by the specific conformation of an additional region of the capsid. J Virol. 1997;71(12):9214–9222. doi: 10.1128/jvi.71.12.9214-9222.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Harrington FE, Easton DP. A putative precursor to the major yolk protein of the sea urchin. Dev Biol. 1982;94(2):505–508. doi: 10.1016/0012-1606(82)90367-0. [DOI] [PubMed] [Google Scholar]
- 20.Harrington DP, Fleming TR. A class of rank test procedures for censored survival data. Biometrika. 1982;69:553–566. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.