

Significance
Although it is well known that reward enhances learning and memory, how extensively such enhancement occurs remains unclear. We examined how reward influences retrieval-induced forgetting (RIF) in which the retrieval of a nonpracticed item under the same category as a practiced item is worse than the retrieval of a nonpracticed item outside the category. RIF was abolished if a juice reward was given for correct answers in the practice phase. These results suggest that reward enhances processing of retrieval of unpracticed members by mechanisms such as spreading activation within the same category, irrespective of whether items were practiced or not.
Keywords: reward, retrieval-induced forgetting, learning, memory
Abstract
Although it is well known that reward enhances learning and memory, how extensively such enhancement occurs remains unclear. To address this question, we examined how reward influences retrieval-induced forgetting (RIF) in which the retrieval of a nonpracticed item under the same category as a practiced item is worse than the retrieval of a nonpracticed item outside the category. Subjects were asked to try to encode category-exemplar pairs (e.g., FISH–salmon). Then, they were presented with a category name and a two-letter word stem (e.g., FISH–sa) and were asked to complete an encoded word (retrieval practice). For a correct response, apple juice was given as a reward in the reward condition and a beeping sound was presented in the no-reward condition. Finally, subjects were asked to report whether each exemplar had been presented in the first phase. RIF was replicated in the no-reward condition. However, in the reward condition, RIF was eliminated. These results suggest that reward enhances processing of retrieval of unpracticed members by mechanisms such as spreading activation within the same category, irrespective of whether items were practiced or not.
Although it has been found that reward significantly enhances learning and memory (1–8), how extensively reward influences learning and memory processing has yet to be explored. It has been recently found that a subthreshold stimulus which was merely exposed in a visual field was learned if paired with reward, but was not if paired with no reward (5). These results suggest that in contrast to the role of attention which is to enhance only processing relevant to a given task, reward enhances learning of a presented item, irrespective of whether the item is task-relevant or not. An important question is how extensively reward influences learning and memory. To address this question, we examined how reward influences retrieval-induced forgetting (RIF) in which the retrieval of a nonpracticed item under the same category as a practiced item is worse than the retrieval of a nonpracticed item beyond the category (9). If reward completely unselectively enhances memory of any item, then it should enhance the retrieval of unpracticed items both within and beyond the category to which the practiced items belong. If reward selectively enhances only practiced items, it should not influence the retrieval of a nonpracticed item irrespective of whether it is within or beyond the category. In both cases, RIF should be observed because reward should not differentially influence the retrieval of nonpracticed items within and beyond the category. In contrast, if reward enhances items only within the category, irrespective of whether it was practiced or not, the degree of RIF should be significantly reduced or abolished.
In the present study, when no reward was given on each trial of the practice session, RIF was observed. However, when reward was given, there was no significant performance difference between recalls of nonpracticed items within the same category as practiced items and nonpracticed items outside the category. These results suggest that reward enhances retrieval of unpracticed members by mechanisms such as spreading activation within the same category, irrespective of whether items were practiced or not.
Experiment
Our experiment consisted of three phases: (i) study, (ii) retrieval-practice, and (iii) memory-test (Fig. 1). In the study phase, category-exemplar pairs (e.g., FISH–salmon) were presented sequentially on a 21-inch cathode ray tube (CRT) display for 5 s. Subjects (62 females and 29 males) were asked to try to memorize (encode) them. After 10 min of an irrelevant distracter task (Methods), the retrieval-practice phase started. In this phase, subjects were presented with a category name and a two-letter word stem (e.g., FISH–sa) and were asked to complete the encoded word by typing the rest of the letters on the computer keyboard. When a subject gave a correct response, drops of apple juice were delivered through a tube from a water dispenser into subject’s mouth as a reward (5, 8) in the reward condition (n = 47), whereas a beeping sound was presented instead of apple juice in the no-reward condition (n = 44). In the final memory-test phase, following a 15-min distracter task, a yes–no recognition test was conducted. Subjects were asked to judge whether each of category exemplars on the display had been presented in the study phase (not the retrieval-practice phase).
Fig. 1.
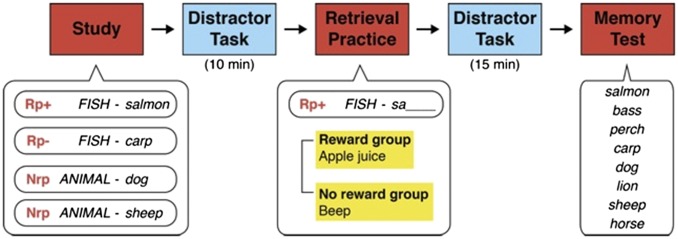
Schematic description of the experimental procedure. Apple juice (reward condition) or a beep (no-reward condition) was given to subjects in the retrieval-practice phase. Rp+, Rp−, and Nrp represent a practiced exemplar, an unpracticed exemplar under the same category as the practiced exemplar, and an unpracticed exemplar not under the same category as the practiced exemplar, respectively.
Results
Fig. 2 A and B shows the results of the no-reward and reward conditions, respectively. A two-way ANOVA [reward (presence vs. absence) as a between-subject factor] × exemplar [Rp+ (practiced exemplars) or Rp− (nonpracticed exemplars under the same category as Rp+), vs. Nrp (nonpracticed exemplars outside the category of Rp+) as a within-subject factor] was applied to “hit” percentages in both conditions. Note that there is no correct rejection for Rp+ or Rp− items (see Methods for a discussion on item balancing). The results showed a significant interaction between the two factors (F2,178 = 3.31, P = 0.036). Further t tests with Bonferroni correction showed that in the no-reward condition. Additionally, the hit percentage with Rp+ was significantly higher than with Rp− (P < 0.001) and Nrp (P < 0.001), and that the hit percentage with Nrp was significantly higher than with Rp−. These results indicate that in the no-reward condition RIF was replicated. On the other hand, in the reward condition, whereas the hit percentage with Rp+ was significantly higher than with Rp− (P < 0.001) and with Nrp (P < 0.001) as in the no-reward condition, there was no significant difference in the hit percentages between Rp− and Nrp. That is, RIF was eliminated by reward.
Fig. 2.

Results of the experiment. (A) Results of the no-reward condition. Percentage of hit for Rp− was significantly lower than for Nrp, showing a fine replication of RIF. (B) Results of the reward condition. RIF observed in the no-reward condition was eliminated. Vertical error bars represent the SEM. ***P < 0.001.
We also calculated an RIF index defined as [(Nrp − Rp−)/Nrp × 100] (%) for each of the reward and no-reward conditions, as shown in Fig. 3. The results of one-way ANOVA [reward (presence vs. absence) as a between-subject factor] indicate that the RIF index was significantly greater for the no-reward condition compared with the reward condition (F1,89 = 7.12, P = 0.009).
Fig. 3.

RIF index for no-reward and reward conditions. Vertical error bars represent the SEM. **P < 0.01.
We further analyzed false alarm rates to examine whether reward affected subjects’ response criteria. The results of two-way ANOVA [reward (presence vs. absence) as a between-subject factor] × category [Rp vs. Nrp as a within-subject factor] did not show any significant main effects nor significant interaction (false alarm rates in reward condition: Rp = 10.86% and SE = 1.64, Nrp = 12.77% and SE = 1.98; false alarm rates in no-reward condition: Rp, 12.12% and SE = 1.76, Nrp, 13.32%, SE = 2.12. See Methods for a discussion on item balancing.). These results are in accord with the hypothesis that subjects’ response criteria were kept constant irrespective of whether reward was given or not.
Discussion
Our result clearly demonstrated that that RIF is eliminated by reward. This suggests that reward enhances and spreads activation to an item that was not practiced but that the spreading does not go beyond the boundary of the category of practiced items.
What is the underlying mechanism of the elimination of RIF by reward? There are at least two models of RIF: inhibitory and noninhibitory. In the inhibitory model, RIF results from inhibition of a nonpracticed item within the category to which practiced items belong (9–14). In the noninhibitory model, RIF occurs as a result of blocking access to a nonpracticed item caused by stronger retrieval of a practiced item in later recall (15–18). Accordingly, the results of the present study suggest that reward either cancels the possible inhibition of unpracticed item or spreads activation to unpracticed items within the category of practiced items in a later recall. Our experiment was to examine the role of reward and was not designed to test which model of RIF is correct. Our finding indicates that reward spreads activation within the same category, irrespective of whether the items are practiced or not. This finding suggests that reward enhances signals in a different fashion than attention, which enhances signals that are relevant to a given task.
It has been found that when subjects had to perform two concurrent updating tasks that demanded executive attention, RIF was not observed, suggesting that executive-control processes play a significant role in RIF (14). This suggests that if RIF is a result of inhibitory control, the abolishment of RIF in the reward condition of our experiment was due specifically to the cancellation of the inhibitory executive control signals by reward.
A number of RIF studies have used a category-plus-stem-cued recall as a final memory test in which Rp− and corresponding Nrp items are shown during the first half of the final test and Rp+ and corresponding Nrp items during the second half of the final test to avoid certain output interference effects. However, the memory test in our experiment followed another line of studies in which a recognition test rather than the cued recall test was conducted, and Rp+, Rp−, and Nrp items along with distractor items were presented in a random order for each subject (11, 12).
We conducted the recognition test for several reasons. First, we had to rule out the possibility that the effect of reward could be contaminated with the association of a cue with reward or internal changes induced by reward because such an association is not directly related to the effect we aimed to examine. For example, because a word used as a category cue in a category-plus-stem-cued test during practice was paired with reward in each “correct” trial of the practice in the reward condition, the word itself could be associated with the activation of processing induced by reward. Thus, if the category-plus-stem-cued test was conducted in the final test as well, the mere presentation of the category word as a cue could activate some internal processing. To avoid such possible artifacts particularly caused by reward, we used a recognition test in which a category cue word was not presented. Second, because the order of presentations of Rp+, Rp−, Nrp, and distractor items was random for each subject, the interference effects may have been negligible, if not completely eliminated.
In human experiments reward is typically provided as monetary incentive. What would occur if monetary reward were given instead of juice reward in the present experiment? Although to our knowledge there is no study that directly compares the strengths of monetary reward and juice reward, it is possible that juice reward is usually stronger because juice reward is a type of primary reward, whereas monetary reward is a secondary reward whose value is derived from primary rewards. Thus, if monetary reward were given in the present experiment, it is possible that RIF would not be completely eliminated.
In summary, we found that reward eliminates RIF. This suggests that reward unselectively enhances processing of tem retrieval irrespective of whether items were practiced or not if these items are within the same category.
Methods
Subjects.
Ninety-one undergraduates of Boston University (62 females and 29 males, mean age = 18.60 y) participated in the current experiment for their course credits (Introductory Psychology course). Forty-seven subjects were assigned to the reward condition, and the remaining 44 subjects were assigned to the no-reward condition. The subjects gave informed consent in the protocol that was approved by the Internal Review Board at Boston University.
Materials.
Six categories (drinks, fish, professions, metals, insects, and animals) were drawn from the Battig and Montague category norm (19). Each category consisted of 12 exemplars. The 12 exemplars from each category were divided into two groups of six exemplars. The mean frequencies and word lengths of the two groups were equated as much as possible. The assignment of six exemplars to one group and that of the remaining six exemplars to the other group were counterbalanced across subjects.
There were six categories. Three categories were assigned as practiced categories and the remaining three categories as not practiced categories. Assignments of these six categories into practiced and nonpracticed categories were counterbalanced across subjects (a more complete counterbalancing would have rotated all items through all conditions over subjects).
Three exemplars from one group of each of the three practiced categories were used as three Rp+ and the remaining three exemplars were used as three Rp− exemplars. The remaining six exemplars from the other group in the same practiced category were used only in the memory-test phase and termed “distractors” (Table 1). The Rp+ exemplars in the practiced category were used in all three phases, whereas the Rp− exemplars in the same practiced category were used only in the study and memory-test phases but not in the retrieval-practice phase (Table 1). Assignment of these two groups to Rp+ or Rp− was counterbalanced across subjects.
Table 1.
Exemplars and their stimulus/response cases in the present study
| Category | Study phase | Retrieval-practice phase | Memory phase | |||
| Exemplar | Response | Stimulus/response | ||||
| Practiced | Presented | Rp+: FISH–salmon | Practiced | Salmon | Yes | Hit |
| No | Miss | |||||
| Rp−: FISH–carp | Not presented | Carp | Yes | Hit | ||
| No | Miss | |||||
| Not presented | Not presented | Perch | Yes | False alarm | ||
| No | Correct rejection | |||||
| Non practiced | Presented | Nrp: ANIMAL–dog | Not presented | Dog | Yes | Hit |
| No | Miss | |||||
| Not presented | Not presented | Lion | Yes | False alarm | ||
| No | Correct rejection | |||||
Six exemplars from one group of each of the three nonpracticed categories were used in the study and memory-test phases but not in the retrieval-practice phase (Table 1). The remaining six exemplars from the other group of the nonpracticed category were used only in the memory-test phase and termed distractors (Table 1).
In addition to the six categories there were three additional categories each containing four exemplars used as buffers. Buffers were used for the first and last several trials in each phase to eliminate/reduce the primacy and/or recency effect.
Procedure.
The experiment lasted ∼50 min for each subject. According to the procedure of typical RIF experiments, our experiment consisted of three phases: study, retrieval-practice, and memory-test phases.
Study phase.
Subjects studied each exemplar from one of the two groups in all six categories (a total 6 × 6 = 36 exemplars. Half of the exemplars corresponded to Rp+ and the other half to Rp− in the subsequent retrieval-practice phase). A category and its exemplar pair (e.g., FISH–salmon) were presented on the display (Hitachi; SuperScan CM801U 21-inch CRT). Subjects were asked to relate the exemplar to its category and to memorize them carefully and told that they would be asked to recall the pairs later in the experiment. Each pair was presented for 5 s.
The presentation order of categories and pairs was randomized for each subject. In addition to 36 exemplars, six buffer items were presented at the beginning of the study phase and the other six buffer items at the end.
After the study phase, subjects were asked to fill out an irrelevant questionnaire which took about 10 min. The task was conducted to avoid rehearsal of studied exemplars and possible interference between the study phase and the upcoming retrieval-practice phase.
Retrieval-practice phase.
Three of the six categories were used. Three exemplars of one of the two groups in each category were practiced (Rp+), and the remaining three exemplars from the same group were not (Rp−). The choices of exemplars for Rp+ and Rp− were counterbalanced across subjects. Retrieval cues were presented on the computer display in a category-exemplar format where the category label and the first two letters of its exemplar of Rp+ pairs were presented (e.g., FISH–sa_____). Subjects were asked to complete a blank space by typing the rest of the letters using a keyboard. Each category-exemplar pair was presented three times. The number of trials was 27 (3 categories × 3 exemplars × 3 repetitions). Subjects were asked to retrieve the item they had memorized in the study phase (e.g., salmon), type the rest of the letters of the retrieved word (e.g., lmon), and then press the “enter” key at the end of the word. They were allowed to delete the typed letters when they made a mistake.
Upon a correct response, drops of apple juice were delivered into their mouths in the reward condition (see below), whereas a beeping sound was presented in the no-reward condition. The presentation order was randomized for each subject. However, we avoided presenting the same Rp+ pairs in any successive trials. Similar to the study phase, the three buffer items were shown before and after the 27 trials.
Immediately after the retrieval-practice phase was completed, a paper-and-pencil task was given to the subjects for about 15 min as a distractor.
Memory-test phase.
A yes–no recognition test was administered in the final memory-test phase. All 12 exemplars per category were presented: six exemplars corresponded to the Rp+ and Rp− exemplars which came from the group used in the study phase (e.g., salmon and carp). The remaining six exemplars had not been shown in any of the previous phases (e.g., perch). Following a fixation point, an exemplar was presented in the center of the display. Subjects were asked to judge whether the exemplar was in the study phase or not by pressing a “yes” or “no” key as fast and accurately as possible. No feedback for response accuracy was given. When they pressed the key or 10 s elapsed without response, the display was cleared for the next trial. A total of 72 trials (6 categories × 12 exemplars) was conducted in randomized order across all categories for each subject. Six buffer items were presented before the 72 trials.
Exemplars.
As mentioned above, there were three phases: study, retrieval-practice phase, and memory-test. Below we explain step by step the subjects’ responses: hit, miss, false alarm, or correct rejection in the memory-test phase. In the memory-test phase, subjects were asked to report whether the exemplars were shown in the study phase. For the simplification purpose, we just use two categories (FISH and ANIMAL) here to explain.
Exemplars belonging to a category practiced in the retrieval-practice phase.
As shown in Table 1, there are three types of practiced exemplars that belong to a category practiced in the retrieval-practice phase (here FISH). Here we explain how the subjects’ hit, miss, false alarm, or correct rejection responses in the memory-test phase.
First, “salmon” is an example of Rp+. In the study phase, subjects were asked to relate it to FISH. In the retrieval-practice phase it was practiced (FISH–sa____). In the memory-test phase, if subjects responded yes to salmon, it was a hit. If they responded no, it was a miss. There was no false alarm or correct rejection for salmon (Rp+) because all of the Rp+ exemplars were shown in the study phase.
Second, “carp” is an example of Rp−. In the study phase, subjects were asked to relate it to FISH. In the retrieval-practice phase it was not presented (but when salmon as a Rp+ was presented, the category FISH, to which carp belongs, was practiced). In the memory-test phase, if subjects responded yes to carp, it was a hit. If they responded no, it was a miss. There was no false alarm or correct rejection for carp (Rp−) because all of the Rp− exemplars were shown in the study phase. Thus, for any exemplars for Rp+ and Rp−, a hit is equivalent to the percent correct. It is impossible to calculate false alarm or correction rejection for Rp+ or Rp− exemplars.
Third, “perch” belongs to FISH, which is a practiced category because salmon had been presented in the retrieval-practice phase. However, perch had been never presented in either the study or the retrieval-practice phase. Therefore, perch is neither Rp+ nor Rp−. In the memory-test phase, if subjects responded yes to perch, it was a false alarm. If they responded no, it was correct rejection. Thus, among the exemplars whose category was practiced in the retrieval-practice phase, false alarm and correct rejection were obtained only for exemplars that were neither Rp+ or Rp−. In other words, false alarm and correct rejection could be computed for the category that was practiced, but not for Rp+ or Rp− exemplars.
Exemplars belonging to a category that was not practiced in the retrieval-practice phase.
“Dog” is an example of Nrp, which belongs to ANIMAL that was not practiced in the retrieval-practice phase. In the study phase, subjects were asked to relate dog to ANIMAL. In the retrieval-practice phase, it was not presented. In addition, none of exemplars that belonged to ANIMAL were presented in the retrieval-practice phase. In the memory-test phase, if subjects responded yes to dog, it was marked as a hit. If they responded no, it was a miss.
As for “lion”, it was not presented in the study phase or the retrieval-practice phase. In the memory-test phase, if subjects responded yes to lion, it was a false alarm. If they responded no, it was a correct rejection.
Reward.
Subjects assigned to the reward condition were asked to put a medical-use tube in their mouths. Drops of apple juice (Mott’s Plus light apple juice; 50% less calories) were delivered as a reward via a water dispenser (Automate Scientific, Inc.; ValveLink8.2 system) connected to a Power Mac G4 (Apple) and controlled by the Psychophysics Toolbox (20, 21) for MATLAB (The MathWorks). Note that in our previous work, subjects were asked not to drink or eat for 5 h before each daily session to make drinking water rewarding (5). However, in the present experiment we used apple juice as the reward (8) and therefore subjects did not have to be deprived of water or food.
Acknowledgments
We thank Aaron V. Berard for comments on a draft. This research was supported by NIH Grants EY015980, EY019466, AG031941, and MH091801.
Footnotes
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
References
- 1.Nomoto K, Schultz W, Watanabe T, Sakagami M. Temporally extended dopamine responses to perceptually demanding reward-predictive stimuli. J Neurosci. 2010;30(32):10692–10702. doi: 10.1523/JNEUROSCI.4828-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bromberg-Martin ES, Hikosaka O. Lateral habenula neurons signal errors in the prediction of reward information. Nat Neurosci. 2011;14(9):1209–1216. doi: 10.1038/nn.2902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Liu J, Lu ZL, Dosher BA. Augmented Hebbian reweighting: Interactions between feedback and training accuracy in perceptual learning. J Vis. 2010;10(10):29. doi: 10.1167/10.10.29. [DOI] [PubMed] [Google Scholar]
- 4.Roelfsema PR, van Ooyen A, Watanabe T. Perceptual learning rules based on reinforcers and attention. Trends Cogn Sci. 2010;14(2):64–71. doi: 10.1016/j.tics.2009.11.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Seitz AR, Kim D, Watanabe T. Rewards evoke learning of unconsciously processed visual stimuli in adult humans. Neuron. 2009;61(5):700–707. doi: 10.1016/j.neuron.2009.01.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Dinse HR, Ragert P, Pleger B, Schwenkreis P, Tegenthoff M. Pharmacological modulation of perceptual learning and associated cortical reorganization. Science. 2003;301(5629):91–94. doi: 10.1126/science.1085423. [DOI] [PubMed] [Google Scholar]
- 7.Seymour B, et al. Opponent appetitive-aversive neural processes underlie predictive learning of pain relief. Nat Neurosci. 2005;8(9):1234–1240. doi: 10.1038/nn1527. [DOI] [PubMed] [Google Scholar]
- 8.O’Doherty J, et al. Dissociable roles of ventral and dorsal striatum in instrumental conditioning. Science. 2004;304(5669):452–454. doi: 10.1126/science.1094285. [DOI] [PubMed] [Google Scholar]
- 9.Anderson MC, Bjork RA, Bjork EL. Remembering can cause forgetting: Retrieval dynamics in long-term memory. J Exp Psychol Learn Mem Cogn. 1994;20(5):1063–1087. doi: 10.1037//0278-7393.20.5.1063. [DOI] [PubMed] [Google Scholar]
- 10.Anderson MC, Bjork EL, Bjork RA. Retrieval-induced forgetting: Evidence for a recall-specific mechanism. Psychon Bull Rev. 2000;7(3):522–530. doi: 10.3758/bf03214366. [DOI] [PubMed] [Google Scholar]
- 11.Hicks JL, Starns JJ. Retrieval-induced forgetting occurs in tests of item recognition. Psychon Bull Rev. 2004;11(1):125–130. doi: 10.3758/bf03206471. [DOI] [PubMed] [Google Scholar]
- 12.Spitzer B, Bäuml KH. Retrieval-induced forgetting in item recognition: Evidence for a reduction in general memory strength. J Exp Psychol Learn Mem Cogn. 2007;33(5):863–875. doi: 10.1037/0278-7393.33.5.863. [DOI] [PubMed] [Google Scholar]
- 13.Román P, Soriano MF, Gómez-Ariza CJ, Bajo MT. Retrieval-induced forgetting and executive control. Psychol Sci. 2009;20(9):1053–1058. doi: 10.1111/j.1467-9280.2009.02415.x. [DOI] [PubMed] [Google Scholar]
- 14.Kuhl BA, Dudukovic NM, Kahn I, Wagner AD. Decreased demands on cognitive control reveal the neural processing benefits of forgetting. Nat Neurosci. 2007;10(7):908–914. doi: 10.1038/nn1918. [DOI] [PubMed] [Google Scholar]
- 15.Raaijmakers JG, Shiffrin RM. Search of associative memory. Psychol Rev. 1981;88:93–134. [Google Scholar]
- 16.Raaijmakers JG, Jakab E. Retrieval-induced forgetting without competition: Testing the retrieval specificity assumption of the inhibition theory. Mem Cognit. 2012;40(1):19–27. doi: 10.3758/s13421-011-0131-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Verde MF. Retrieval-induced forgetting in recall: Competitor interference revisited. J Exp Psychol Learn Mem Cogn. 2013;39(5):1433–1448. doi: 10.1037/a0032975. [DOI] [PubMed] [Google Scholar]
- 18.Jakab E, Raaijmakers JG. The role of item strength in retrieval-induced forgetting. J Exp Psychol Learn Mem Cogn. 2009;35(3):607–617. doi: 10.1037/a0015264. [DOI] [PubMed] [Google Scholar]
- 19.Battig W, Montague W. Category norms for verbal items in 56 categories: A replication and extension of the Connecticut category forms. Journal of Experimental Psychology Monograph. 1969;80(3, Pt 2):1–46. [Google Scholar]
- 20.Brainard DH. The Psychophysics Toolbox. Spat Vis. 1997;10(4):433–436. [PubMed] [Google Scholar]
- 21.Pelli DG. Pixel independence: Measuring spatial interactions on a CRT display. Spat Vis. 1997;10(4):443–446. doi: 10.1163/156856897x00375. [DOI] [PubMed] [Google Scholar]