

Abstract
The Chagos Archipelago was designated a no-take marine protected area (MPA) in 2010; it covers 550 000 km2, with more than 60 000 km2 shallow limestone platform and reefs. This has doubled the global cover of such MPAs.
It contains 25–50% of the Indian Ocean reef area remaining in excellent condition, as well as the world’s largest contiguous undamaged reef area. It has suffered from warming episodes, but after the most severe mortality event of 1998, coral cover was restored after 10 years.
Coral reef fishes are orders of magnitude more abundant than in other Indian Ocean locations, regardless of whether the latter are fished or protected.
Coral diseases are extremely low, and no invasive marine species are known.
Genetically, Chagos marine species are part of the Western Indian Ocean, and Chagos serves as a ‘stepping-stone’ in the ocean.
The no-take MPA extends to the 200 nm boundary, and. includes 86 unfished seamounts and 243 deep knolls as well as encompassing important pelagic species.
On the larger islands, native plants, coconut crabs, bird and turtle colonies were largely destroyed in plantation times, but several smaller islands are in relatively undamaged state.
There are now 10 ‘important bird areas’, coconut crab density is high and numbers of green and hawksbill turtles are recovering.
Diego Garcia atoll contains a military facility; this atoll contains one Ramsar site and several ‘strict nature reserves’. Pollutant monitoring shows it to be the least polluted inhabited atoll in the world. Today, strict environmental regulations are enforced.
Shoreline erosion is significant in many places. Its economic cost in the inhabited part of Diego Garcia is very high, but all islands are vulnerable.
Chagos is ideally situated for several monitoring programmes, and use is increasingly being made of the archipelago for this purpose.
Keywords: Chagos, British Indian Ocean Territory, marine protected area, coral recovery, reef fishes, seamounts, reef disease, marine invasives, fisheries, island conservation
INTRODUCTION
This paper reviews the scientific work and historic information that demonstrates the outstanding ecological values of the Chagos Archipelago, which led in 2010 to the creation of the world’s largest marine protected area (MPA), and which became fully a no-take MPA later the same year. The archipelago (Figure 1) forms the British Indian Ocean Territory (BIOT), created in 1965 for UK and USA defence purposes. It is a large group of atolls and submerged banks in the central Indian Ocean, lying at the southernmost end of the Lakshadweep–Maldives–Chagos ridge. Its central 200 × 300 km area contains five atolls with islands, one atoll which is awash at high tide, and a dozen more which are submerged to depths of 6–25 m. The Great Chagos Bank is the world’s largest atoll in area, although it contains only eight islands on its western and northern rim. The area within the total BIOT 200 nm zone is about 550 000 km2.
Figure 1.

The Chagos Archipelago. Inset shows location and MPA boundary (circular shape with flattened northern border). Main map: the five atolls with land are shown in bold, the islands on Great Chagos Bank and submerged reefs and atolls are not bold. All are located in the central area of the MPA.
During recent decades, most of the tropical ocean has been heavily affected by pollution, over-exploitation and various unwise forms of development (Millennium Ecosystem Assessment, 2005). Almost all indicators of ‘ocean health’ continue to show worsening trajectories, and many attempted remedial measures have failed to arrest the decline of habitats and ecosystems essential for both human welfare and maintenance of biodiversity and productivity. The world’s oceans are affected by overfishing, pollution, agriculture and industry, shoreline construction, and climate change (Jackson et al., 2001). Coral reefs in particular are overexploited because they support growing populations of some of the world’s poorest people (Wilkinson, 2008a; Burke et al., 2011). All reefs are highly vulnerable to increasing intensity of human exploitation, which reduces both biomass and productivity, and consequences of reef deterioration may be greater than previously anticipated (Mora et al., 2011). Most conventional forms of marine management are failing to arrest the decline, so many marine science bodies, conservation organizations and international conventions have called for more and larger marine protected areas (MPAs) that have effective levels of protection (United Nations, 2002; Wood et al., 2008; Nelson and Bradner, 2010). MPAs remain one of the only extensive tools being used directly for conservation purposes (Spalding et al., 2010).
A recent assessment (Toropova et al., 2010) showed that there are about 5900 MPAs, covering 4.2 million km2, which covers only 1.17% of the oceans. Recently their median size was only 5 km2 (Wood et al., 2008). The small proportion of ocean covered by MPAs must be compared with estimated need. The Convention on Biological Diversity called initially for 10% of the world’s EEZ to be protected by some form of MPA by 2010, though the target date was later put back to 2020. The World Parks Congress called for 20% of the oceans to be protected by 2012, and the World Summit on Sustainable Development called for a global network of comprehensive, representative and effectively managed MPAs by 2012 (United Nations, 2002). In developed countries of the Atlantic, the OSPAR Commission called for an ecologically coherent, well managed network of MPAs also by 2012. Most targets will manifestly fail, and participating parties would need to create over 19 million of the median size MPAs to achieve these targets. In 2010, 260 marine scientists made an urgent call for more and larger MPAs (http://www.globaloceanlegacy.org/), and a figure of 30% (not 10–20%) has increasingly emerged as the area needed to avert permanent damage.
Unfortunately, the meaning of ‘protected’ varies widely, with most allowing only partial protection and many also allowing fishing (one of the most ecosystem distorting activities), while many lack any protection at all. The latter are commonly called ‘paper parks’ and, regrettably, human pressures make this category the large majority. For coral reefs, only 6% are effectively managed, 21% are ineffectively managed, and 73% lie outside any MPA (Burke et al., 2011). Reasons for MPA failures range from being declared solely to meet ‘targets’ which are then inadequately resourced, to being simply overwhelmed by close proximity to large human populations.
Chagos and the marine protected area
Chagos was occupied from the 18th century, during which time most of its native vegetation was converted to coconut plantations. This industry lasted until the 1970s. Two of the five atolls were abandoned for economic reasons and social problems in the 1930s, and in the early 1970s the plantations on the remaining three atolls were closed due to the establishment of a military facility. The people were moved to various countries, notably Mauritius and the Seychelles, and some eventually to Europe, especially England (Edis, 2004). The remaining plantations were already in some decline given diminishing world demand for coconuts and the ascendency of palm oil from elsewhere, but nevertheless political issues surrounding the forced removal of the last inhabitants has been mired in controversy ever since. For the last 40 years the islands have been uninhabited except for the southernmost atoll of Diego Garcia, the western arm of which contains the military facility and at least 1000 Asian contractors and others. That facility does not depend on local food resources but is provisioned and supported entirely from outside the archipelago.
Over the period of BIOT’s existence, there have been a dozen scientific visits, involving more than 50 visiting scientists. It has become clear during this period when coral reefs in most of the Indian Ocean have become seriously degraded that those of Chagos persist in an exceptionally good state. This led increasingly to calls to extend its conservation, and data to support the concept came from over 200 papers arising from both those scientific expeditions and, to a lesser extent, from unpublished information, from regional data and from modelling.
Recently, the Pew Ocean Legacy Program included Chagos as one of five areas selected for protection, and promoted efforts to convince the UK Government to declare it a no-take MPA to the 200 nm boundary (Nelson and Bradner, 2010). Part of this process was the creation of the Chagos Environment Network (CEN), a loose association of several leading UK science bodies and NGOs, whose role was to support efforts to ensure that Chagos’ globally important natural environment would be conserved as a unique and valuable resource for present and future generations. In 2010, CEN responded to the UK Government’s Consultation, saying that only designation as a no-take MPA ‘…guarantees full protection for the ecosystems and species of the Chagos Archipelago and its surrounding reefs, lagoons and waters. Only [this] provides the complete protection needed to underpin the Chagos Protected Area’s value as an important global reference site for a wide range of scientific ecological, oceanographic and climate studies, as well as its continued benefits to humans into the future’ (CEN, 2010).
BIOT is a UK Overseas Territory and as such has its own government, which in this case is the office of the BIOT Administration located in the UK Foreign and Commonwealth Office in London. The senior UK military officer in the archipelago is the British Representative of the Commissioner. The UK Foreign Secretary announced the creation of BIOT as a no-take MPA, instructing the Commissioner to declare it as such in April 2010 (http://www.fco.gov.uk/en/news/latest-news/?view=News&id=22014096). The Commissioner, in Proclamation Number 1 of 2010, proclaimed it such ‘in the name of the Queen’. Existing environmental laws are currently being revised and consolidated to accommodate this status. Diego Garcia atoll to its 3 nm boundary is excluded from the MPA. The area thus excluded is less than 1% of the total area although it has several pre-existing, strict environmental laws of its own, and contains a Ramsar site (see later). Existing tuna fishing licences were discontinued, and the deficit for BIOT finances of approximately $1 million per year was subsequently replaced from private sources. It is currently the largest MPA in the world (Nelson and Bradner, 2010) and is now also part of the ‘Big Ocean Network’, an information exchange network of managers and partners of existing and proposed large-scale marine managed areas (www.bigoceanmanagers.org/). Monitoring and enforcement are undertaken in large part by a patrol vessel which serves as a mobile base for both the military and civilian research expeditions.
Island and reef areas
Figure 2(a) shows the areas of islands and reefs. Diego Garcia contains half the total land area, the rest being split among over 50 small islands, the number varying to some degree with tidal height and shifts of sand banks. All islands are very low lying, and are typical coral cays constructed of limestone. Beneath these lie freshwater lenses sustained by high rainfall.
Figure 2.

a) island areas in the Chagos Archipelago (scale in ha). The largest islands are named. b) area of submerged substrate in the archipelago (scale in km2) (from Dumbraveanu and Sheppard, 1999).
In contrast to the small island areas, sublittoral substrate in the photic zone is calculated to be approximately 60 000km2 (Figure 2(b)) (Dumbraveanu and Sheppard, 1999). How much of this huge area is actively growing reef is uncertain; more than 95% of the territory has never been studied, though some areas are apparently eroding and others support sand and/or large seagrass beds. There is enormous opportunity for new discoveries: as recently as 2010 an expedition discovered many hectares of seagrass and a previously unknown 3 km2 mangrove forest. Other parts of Chagos have been mapped using bathymetric or satellite data-based modelling (Yesson et al., 2011, see later Figures 9 and S12 in Supplementary Material).
Figure 9.

Seamounts of the Chagos MPA as identified in Yesson et al. (2011). Bathymetry data from shuttle radar topography mission 30 arc-second grid (http://www2.jpl.nasa.gov/srtm/).
REEF CONDITION
Coral cover and changes due to mortality episodes
There were no quantitative studies of reef condition on Chagos reefs before the 1970s, although descriptive studies, notably Stoddart and Taylor (1971), described land and reef flat in Diego Garcia. From the 1970s, episodic visits enabled a series of coral cover measurements to be taken on reef slopes.
Coral cover declined between the first survey in 1978 (Sheppard, 1980a) and the next in 1996 (Sheppard, 1999a) (Figure 3). This was mainly due to loss of shallow and mid-depth branching species, particularly Acropora palifera and table corals including Acropora cytherea, and causes were only speculated upon at the time (Sheppard, 1999a). Later, after much work globally and more surveys in Chagos, the cause was suggested to be several warming events. This modest decline occurred in many Indian Ocean islands groups over this period (Ateweberhan et al., 2011) and in Kenya (Muthiga et al., 2008) though it was not universal.
Figure 3.

Percentage coral on reefs by the main coral types in 4 years, for the major live categories identifiable by snorkelling in 1999. Arrows along the top are dates and approximate relative severity (arrow thickness) of previous warming events (from Sheppard, 1999a).
Severe warming in 1998 then caused severe mortality on all Chagos reefs (Sheppard, 1999b; Sheppard et al., 2002; Figures 3 and S1 in Supplementary Material) as it did throughout the Indian Ocean (Ateweberhan et al., 2011). Coral and soft coral mortality was almost total on several Chagos ocean-facing reefs to clearly defined depths, below which corals provided much higher cover. This killed zone extended deeper in southern atolls, and in Diego Garcia for example was greater than 40 m depth, while in more northern atolls it extended to only about 10–15 m depth (Sheppard et al., 2002). Such variability was mirrored in the Indian Ocean as a whole (Sheppard, 2006). Lagoon reefs of Chagos atolls were much less affected than ocean facing reefs (Figure S1 in Supplementary Material), with many retaining high coral cover, including stands of Acropora. Post-1998, coral species diversity was greatest in deep lagoon areas.
Coral recovery
Several ocean-facing transects around the atolls have been monitored repeatedly from 1999 onwards (Figure 4). On these, no increase of hard coral cover was seen for 3 years following the mortality, although by 2001 large numbers of juveniles were present, at densities of up to 28 m−2, the highest recorded globally at that time (Sheppard et al., 2002; Harris and Sheppard, 2008). Coral spat provided 6% cover on easily measured (but disintegrating) dead coral tables, with a further 5% cover provided by juvenile soft corals, indicating good recovery potential (Figure S2 in Supplementary Material). Increase in coral cover became evident by 2006, especially in shallow water (Harris and Sheppard, 2008; Sheppard et al., 2008) where restoration to pre-1998 levels occurred by 2010. Deeper recovery has been slower. Coral cover in 2011 reached values recorded in 1978 in a few transects in Diego Garcia, but most atolls were not surveyed during that year.
Figure 4.

Coral cover in depths to 25 m on ocean-facing slopes in different years (Sheppard 1980a, 1999a, 1999b; Sheppard et al., 2008; Sheppard unpublished data). Data are of all ocean-facing transects in this series measured on each date. Bars are error bars (error data lost for 1978).
In 2006, extensive video surveys were taken and archived, showing mean percentage cover of several benthic categories (Bayley, 2009) (Figures 5 and S3, S4 in Supplementary Material). These showed no significant differences that year in hard coral cover in pooled data between atolls, although significant differences existed at depth and site levels between ocean-facing reefs. Hard coral cover was significantly higher in lagoons (63.04 ± 3.19%) than ocean-facing slopes (39.69 ± 2.03%). Soft coral was higher on ocean-facing slopes (14.02 ± 1.58%) than in lagoons (2.65 ± 0.72%). Hard coral cover decreased between 6 and 25 m, but sponge and soft corals showed an increase with depth. Dead standing coral at most sites was low at 3–13%, and rubble did not change significantly at different depths. Structural complexity was reduced to 15 m in the outer atolls, and to deeper zones on the Great Chagos Bank and Diego Garcia atoll. By 2006 all shallow regions had developed sufficiently to form a canopy, with colonies competing with one another for space (O’Farrell, 2007). Furthermore, deep lagoonal areas exhibited the highest numbers of small, juvenile coral colonies. Modelling studies indicate that such deep reef areas could be responsible for relatively rapid recolonization of denuded shallow reefs (Riegl and Piller, 2003).
Figure 5.

Mean percentage cover values of life form and substrate categories pooled from all depths and all sites for each of four atolls (GCB:Great Chagos Bank, DG: Diego Garcia, SAL: Salomon atoll, PB: Peros Banhos atoll), surveyed by video during 2006. Bars are error bars.
The past decade has seen further coral bleaching events in Chagos, in 2003, 2004, 2005, and a mild one in 2010. None were sufficient to cause mass mortality, although species-specific coral mortality was recorded of many Acropora cytherea tables in 2010 (Pratchett et al., 2010). Given that warming episodes sufficient to kill corals are predicted to increase (Sheppard, 2003; Hoegh-Guldberg et al., 2007) it is likely that intermittent interruptions to coral growth will continue. However, models based on recruit availability scaled to the present coral cover, suggest that Chagos reefs will long be able to withstand recurring strong mortality events – even each decade – and still maintain high coral cover. Significant declines in cover are expected only if both larval supply decreases and coral mortality events increase in intensity and frequency.
Pre- and post-1998 studies have also revealed some changes in the soft coral component. The principal octocorals before and after the 1998 ENSO shared many common taxa (Reinicke and van Ofwegen, 1999; Schleyer and Benayahu, 2010), but a few discontinuities in their biodiversity indicate subtle changes in more persistent genera (Lobophytum, Sarcophyton). Some fast-growing ‘fugitive’ genera (e.g. Cespitularia, Efflatounaria, Heteroxenia) disappeared after the ENSO-related coral bleaching (Reinicke and van Ofwegen, 1999; Schleyer and Benayahu, 2010), suggesting that such transient fugitives might be eliminated from soft coral communities on isolated reef systems by bleaching disturbance of this nature. Carijoa riseii, a species often considered a fouling organism, and even an invasive in some places (Concepcion et al., 2010), was found in 2006. The observed post-ENSO recovery gives cause for hope for soft coral survival in the face of climate change.
Reef fish
Biomass of reef fish around the northern atolls of Chagos was quantified for the first time in 2010, using underwater visual census (Graham, 2010). Biomass estimates for Chagos exceed values from both fished and protected reefs elsewhere in the region, such as Kenya, Seychelles, Madagascar and even the Maldives, by several orders of magnitude (McClanahan et al., 2009; McClanahan, 2011; Graham, 2010). The Chagos biomass estimates are matched only by some remote, unfished locations in the Pacific Ocean (Sandin et al., 2008; Williams et al., 2011).
With recent declines in coral cover globally (Ateweberhan et al., 2011) there is growing concern over impacts on reef fish assemblages (Pratchett et al., 2008). Much work has followed the 1998 bleaching event around the world in trying to document changes to reef fish populations. A compilation of short-term impacts (<3 years post-coral mortality) of coral loss on fishes indicated that species displaying population declines were those that specialized on coral for food, shelter or settlement (Wilson et al., 2006). However, subsequent studies have shown that longer term effects of coral loss and reef structural collapse on fish are much wider reaching, with a large portion of the fish community affected, including reductions in species richness, reduced abundance of many groups and changes in the size structure of the community (Jones et al., 2004; Graham et al., 2006, 2007, 2008; Wilson et al., 2008). Reef fish surveys in Chagos conducted in 1996 were repeated in 2006 using identical methodology. This was part of a study across seven Indian Ocean nations. One of the groups of fish most affected by coral loss is coral-feeding butterflyfishes, but a comparison between Chagos and Seychelles demonstrated that, although both specialized and generalist coral feeding butterflyfishes showed declines in abundance in the Seychelles through the 1998 bleaching event, there was no detectable difference in Chagos (Graham et al., 2009a).
Across the Indian Ocean as a whole there were declines in fish species richness, and in the abundance of corallivores, planktivores and small bodied fish (<20 cm maximum attainable size) (Graham et al., 2008; MacNeil and Graham, 2010) as a result of reef degradation. In contrast, the populations of Chagos remained remarkably stable (Figure 6). Changes in fish species richness for Chagos were extremely small and either side of zero, compared with the regional decline. This highlights the temporal stability of Chagos fish assemblages in response to a large-scale disturbance, despite substantial negative responses elsewhere in the ocean, and accords with findings of Mora et al. (2011) and Sandin et al. (2008) that reef fish assemblages are very vulnerable to anthropogenic stressors.
Figure 6.

Change in reef fish species richness across seven countries in the Indian Ocean following the 1998 coral mortality event. Chagos sites represented by filled circles. Adapted from Graham et al. (2008).
Also of note are some different fish behaviours that are very rarely seen elsewhere around the world where human exploitation, coastal development, and other impacts have changed abundances, ecological interactions, and behaviour. One such example is the daytime feeding behaviour of the moray eel, Gymnothorax pictus, on shore crabs, leaping clear of the water to capture their prey (Graham et al., 2009b). Behaviours like this, and the exceptional stability and abundance of the reef fish communities, make Chagos a very important reference area with which scientists can understand ecological and behavioural changes elsewhere in the world.
Coral diseases
In 2006 a survey assessed corals along 37 transects at eight sites across the archipelago (Figure 7). Overall prevalence of disease was 5.2%, which sits at the low end of the global spectrum where regional averages for ‘white syndrome’ alone are around 5% in parts of Australia, Palau, and East Africa, 8% in the Philippines (Weil et al., 2002; Willis et al., 2004; Raymundo et al., 2005) up to around 13% at some sites in the Eastern Indian Ocean (Hobbs and Frisch, 2010), and 20% in the Caribbean (Weil et al., 2006; Miller et al., 2009).
Figure 7.

Percentage of adult colonies in different atolls showing disease or other adverse conditions. Bars are standard error bars.
Temperature has been shown to be a key factor triggering diseases, with infection occurring rapidly at elevated temperatures (Ben-Haim and Rosenberg, 2002; Bruno et al., 2007; Harvell et al., 2007). Thus the increasing frequency of raised temperature episodes gives cause for concern. Coral diseases often arise from changes to the normal, commensal relationship between the coral and the bacterial community in their mucus, skeleton, and tissues (Rohwer and Kelley, 2004; Lesser et al., 2007). Physiological stresses that cause corals to become overwhelmed by bacteria are often anthropogenic in origin, coming from sediment deposition, nutrient rise (Bruno et al., 2003; Kaczmarsky and Richardson, 2011), or sea temperature rise (Harvell et al., 2007; Zvuloni et al., 2009). Other factors correlated with the likelihood of coral disease include geographical range and predator diversity (Diaz and Madin, 2011), while a higher density of individuals also increases susceptibility (Willis et al., 2004; Bruno et al., 2007). Although remoteness from people is no guarantee of absence of disease, especially if temperature rises (Williams et al., 2007), mitigation of other human induced stress factors may reduce disease prevalence (Bruno et al., 2003; Harvell et al., 2007). At present, Chagos reefs have very low disease levels.
Marine invasive species
Marine invasive alien species (IAS) are recognized as one of the most significant threats to global biodiversity (Wilcove et al., 1998; Bax et al., 2010) and documented IAS are commonly significantly underestimated. IAS pressure is driving global declines in species diversity, with the overall impact apparently increasing (McGeoch et al., 2010). Notably, over 100 introduced marine species covering 14 phyla are known from ports in the Pacific (Coles et al., 1999). Article 8(h) of the Convention on Biological Diversity calls for prevention of introductions and control or eradication of alien species that threaten ecosystems, habitats or species, and the recently agreed Aichi Biodiversity Targets call for identifying pathways and putting in place by 2020 measures to prevent species introduction and establishment.
Ballast water and hull fouling provide the primary vectors for marine species introduction (Cohen and Carlton, 1998; Ruiz et al., 2000; Hewitt et al., 2004). Navy and supply ships frequently arrive in Diego Garcia, mainly from the USA, the Middle East, and Singapore, but any ships, including recreational yachts, may carry hull fouling organisms (Bax et al., 2002). Therefore pathways for species introductions to Chagos exist, as do preconditions for successful establishment (Tamelander et al., 2009). While most ships arrive at Chagos loaded, some may be empty and ballasted. Ballast water exchange occurs outside the lagoon and during mid-crossing in keeping with IMO ballast water management guidelines (IMO, 2004).
A survey of non-native marine biota in Chagos was carried out in 2006 in all atolls (Tamelander et al., 2009) based on standard port survey methods (Hewitt and Martin, 2001) but with a lower sensitivity. Hard and soft substrate benthic biota were sampled at 42 sites (19 sites in Diego Garcia, nine each on the Great Chagos Bank and at Peros Banhos, and five in the Salomon atoll). Twenty-four phyla were represented in 2672 samples, with four phyla (Bryozoa, Mollusca, Annelida and Porifera) each making up over 10% of the total number of specimens.
No non-native species were detected in the samples, the first time such a survey has not found species introduced as a result of human activities (Tamelander et al., 2009). This finding is testament to the ecological integrity of Chagos’ marine ecosystems. Shallow marine habitats are believed to be particularly vulnerable to bioinvasions when degraded (Heywood, 1995), but ecosystem health and high biodiversity confer higher resistance. Only 16% of marine ecoregions have no reported marine invasions, although the true figure may be lower because of under-reporting (Molnar et al., 2008).
Because controlling or eradicating a marine species once it is established is nearly impossible (Bax et al., 2002), management must focus on precautionary measures (Thresher and Kuris, 2004; Carlton and Ruiz, 2005). Successful prevention and management of IAS threats in Chagos is a prerequisite for effective management of the newly established MPA (Pomeroy et al., 2004; Tu, 2009). Further, this needs to be devised in the broader context of climate change and the potentially greater risk of species spread and establishment that this may bring (Bax et al., 2010; Burgiel and Muir, 2010).
Chagos reef condition in the Indian Ocean context
Most Indian Ocean reef areas are heavily exploited and many have shown limited recovery following the 1998 bleaching disturbance (Wilkinson, 2008b; Harris, 2010). Many which declined catastrophically in 1998 and which also suffer from local impacts have not recovered significantly, or at all (Harris, 2010). The 1998 bleaching event is the main determinant of coral cover change in the Indian Ocean since the 1970s (Ateweberhan et al., 2011), and the central regions, which had some of the highest coral cover estimates before 1998, suffered the worst during the bleaching event. Subsequent recovery for most of these reefs now remains below average for the region, but of the central Indian Ocean reefs, recovery in Chagos is higher than elsewhere (Ateweberhan et al., 2011).
Globally, a third of reef-building corals are threatened with extinction (Carpenter et al., 2008) and today, in the Indian Ocean, only about a third of reefs may be attributed to a ‘low threat level’ category (Wilkinson, 2008a, 2008b). Chagos reefs fall within this minority group and contain a substantial proportion of reef area in very good condition. Reef area estimations are difficult, and have been subject to wide variation. Spalding et al. (2001) suggested the Indian Ocean has 32 000 km2 of reefs (the Red Sea region and the Gulf region adding 17 400 and 4200 km2 more, respectively), and, based on this, Chagos has 3770 km2 of reefs (Rajasuriya et al., 2004) meaning Chagos comprises up to half of this ocean’s reefs in a ‘low threat level’ category (Figure 8). More recent calculations by Spalding (pers. comm. and see Burke et al., 2011) resulted in a revised estimate that Chagos provides 25% of reefs in the ‘low threat’ category. Even this ‘…25% of the region’s low threat reefs is still an extraordinary proportion, and it is also worth stressing that in addition to this, these are by far the largest contiguous reef tracts considered to be under low threat’ (Spalding pers. comm., 2011).
Figure 8.
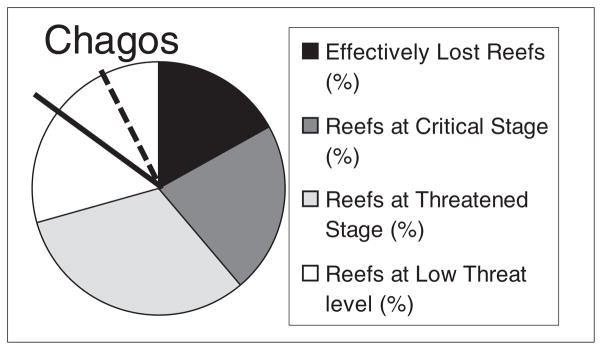
Percentage of reefs in different categories in the Indian Ocean. Categories are those from Wilkinson (2008a). The probable proportion occupied by Chagos (solid line to vertical) is about half of the reefs in the ‘best’ category. From the dashed line to vertical is an alternative estimate of the proportion of Chagos reefs according to Spalding using slightly different categories (pers. comm.).
While both area values are much less than the area of illuminated shallow limestone substrate which was calculated from detailed bathymetric plotting (Figure 2), the values used have the important benefit that they were calculated consistently throughout the world, thus permitting comparisons; direct measurements based on bathymetry do not yet have a counterpart in most other countries.
For other Indian Ocean areas, Tamelander and Rajasuriya (2008) report that ‘recovery of South Asian coral reefs since the 1998 mass bleaching has been patchy; Chagos has shown particularly good recovery, reefs in the western atoll chain of the Maldives and Bar Reef in Sri Lanka have also recovered relatively well, while many reefs near Sri Lanka and reefs in the eastern atoll chain of the Maldives have shown little or no recovery… Coral larval recruitment was very strong, such that the lowest Chagos recruit densities were at least 10 times higher than rates of recruitment at most other reefs in the central and western Indian Ocean.’ For island archipelagos further west, Wilkinson (2008a) reported that ‘some reefs of the Seychelles and Comoros that suffered major damage in 1998 have probably regained about half the lost coral cover; there has also been virtually no recovery on others’. Numerous other compilations (Wilkinson, 2008a; Burke et al., 2011) report patchy or poor coral recovery in most other parts of the region.
Reasons for the good condition of Chagos reefs are likely to include remoteness from compounding human activities, but some additional factors may contribute. Strong light adapted ‘Clade A’ forms of symbiotic zooxanthellae have been identified in shallow corals in Chagos, occurring in approximately half of the shallow water Acropora colonies that were heavily affected by warming but which are now recovering strongly (Yang et al., 2012; Figure S5 in Supplementary Material). Also, an array of temperature sensors at different depths has identified regular incursions of deep, cool water that rise to cover reefs, including during the annual periods of greatest warming (Sheppard, 2009).
But it is increasingly understood that direct human pressures are the main cause of reef degradation and this has often been underestimated in the past (Mora et al., 2011). Such activities impede recovery, and absence of herbivore extraction, pollution, and sedimentation increase reef resilience (Hughes et al., 2010). Most of Chagos has no human population at all. Diego Garcia imports all its requirements and for the last 15 years at least has had strong environmental management. Lack of human pressures is likely to be one major reason for the present good condition of these reefs.
In the Indian Ocean as a whole, direct human pressures can only increase further. Of this region, a decade ago Spalding et al. (2001) noted: ‘Human populations … are rapidly increasing. Most of the coastal populations are very poor, and heavily dependent on the adjacent reefs for food. Unfortunately there is little control over the utilization of these resources, either through traditional or formal management regimes, and large areas of reefs have been degraded through overfishing or destructive fishing techniques.’ With annual population growth rates of 2.5% being common in the region, compounded by migration to the coast in some countries that have experienced wars or drought, coastal populations and exploitation of reefs have increased substantially since Spalding’s (2001) account. As reefs degrade, the proportion of healthy reefs of the Indian Ocean contained in Chagos, already very high, continues to increase, so that a precautionary approach to their protection is merited.
DEEP-WATER ECOSYSTEMS
Yesson et al. (2011) determined that 86 seamounts (conical topographic rises >1000 m elevation) and 243 knolls (conical topographic rises of elevation 200–999 m) are predicted to occur within the Chagos MPA (Figure 9). Chagos thus contains more than 10% of all Indian Ocean seamounts and so the area is regionally important for these features as well. Given that globally only 506 seamounts and 606 knolls lie in protected areas (Yesson et al., 2011, based on the world database of protected areas 2009), this means that the Chagos MPA increased the world’s protection of seamounts by 17% and knolls by 40%. Previous emphasis of the Chagos MPA has been on shallow-water ecosystems, but protection of its seamounts is also important, especially considering their high biodiversity, often representing entirely unique ecosystems (Clark et al., 2006). Although the geology of some of the seamounts and ridges in the Indian Ocean has been explored, including the Chagos-Laccadives Ridge, seamount fauna is poorly known (Rogers et al., 2007). Some data on fish exist, mainly resulting from exploratory or commercial fishing, but no specific information relates to the Chagos-Laccadive Ridge. Recent modelling studies based on 30-arc second satellite bathymetry data indicate that the Indian Ocean hosts fewer seamounts than the Atlantic and Pacific Oceans (Yesson et al., 2011), and many are associated with ridges or originate at ridges.
The Indian Ocean suffers increasing pressure from deep-sea fishing that threatens both seamounts and other benthic habitats. The fact that there has never been deep-water fishing or trawling in Chagos makes it particularly important when considered in a regional context. Deep-sea fishing in the Indian Ocean was mostly undertaken by distant-water fleets, particularly from the USSR. These fisheries targeted redbait (Emmelichthys nitidus) and rubyfish (Plagiogeneion rubiginosus) with catches peaking about 1980 and then decreasing to the mid-1980s (Clark et al., 2007). Fishing then switched to alfonsino (Beryx splendens) in the 1990s as new seamounts were exploited. Some exploratory trawling was also carried out on the Madagascar Ridge and South-west Indian Ocean Ridge by French vessels in the 1970s and 1980s, particularly targeting Walter’s Shoals and Sapmer Bank (Collette and Parin, 1991). In the late 1990s, a new fishery developed on the South-west Indian Ocean Ridge with trawlers targeting deep-water species such as orange roughy (Hoplostethus atlanticus), black cardinal fish (Epigonus telescopus), southern boarfish (Pseudopentaceros richardsoni), oreo (Oreosomatidae) and alfonsino (Clark et al., 2007). This fishery rapidly expanded, with estimated catches of orange roughy being approximately 10 000 tonnes, until the fishery collapsed. Fishing then shifted to the Madagascar Plateau, Mozambique Ridge, and Mid-Indian Ocean Ridge, targeting alfonsino and rubyfish (Clark et al., 2007). Most of these areas therefore have probably been significantly affected by past deep-sea bottom fisheries.
Deep-sea fishing in most of the Indian Ocean is continuing and showing signs of increasing its geographic spread, mainly targeting orange roughy and alfonsino. Recent fishing has also taken place on the Broken Ridge (eastern Indian Ocean), 90 East Ridge, possibly the Central Indian Ridge, the Mozambique Ridge and Plateau and Walter’s Shoal (western Indian Ocean), where a deep-water fishery for lobster (Palinurus barbarae) has developed (Bensch et al., 2008). The banks around Mauritius and high seas portions of the Saya da Malha Bank have been targeted by fisheries for Lutjanus spp., and lethrinid fish (SWIOFC, 2009), and there are also reports of unregulated gillnet fishing in the Southern Indian Ocean such as at Walter’s Shoal, which target sharks (Shotton, 2006). Currently, there is little or no information available on impacts of deep-sea fishing in high seas areas of the Indian Ocean on populations of target or by-catch species, or on seabed ecosystems. Reporting of data is complicated by issues of commercial confidentiality in fisheries where individual stocks may be located across a wide area (e.g. the South-west Indian Ocean Ridge), and there is no adequate regional fisheries management body (see below). At present, new fisheries are developing in the region with no apparent assessment of resource size or appropriate exploitation levels to ensure sustainability of fisheries, or to estimate impacts of such fisheries on vulnerable marine ecosystems.
Global modelling studies are currently also being undertaken of habitat suitability for deep-sea Scleractinia and Octocorallia, at 30-arc second resolution (Davies and Guinotte, 2011; Yesson et al., in press). These indicate that suitable habitat for these organisms exist on deep slopes and seamounts within the Chagos MPA (Davies and Guinotte, 2011; Yesson et al., in press). Given the lack of a history of deep-sea fishing in the region around the Chagos Archipelago, it is likely that associated communities of invertebrates and fish are still largely intact, unlike on most other ridges in the Indian Ocean that have been fished or which are subject to continuing or expanding fisheries.
Given the lack of research in general on equatorial seamounts, the MPA is thus particularly important for deep-water ecosystem conservation, both at a regional and global level. It also provides a unique opportunity to investigate the energetic links between production associated with shallow-water coral reefs and deep-water ecosystems, an area of marine ecology that has not been explored.
PELAGIC FISHING AND FISHERIES
While no deep-sea trawling has been recorded in the Chagos EEZ, there have been impacts from fisheries operating in both in-shore and pelagic environments. These fisheries provided most income for Chagos until the cessation of fishing licences after the MPA was created, with the last licences expiring on 31 October 2010. The main fisheries were a longline and purse seine fishery for tuna, and an inshore fishery. There is also a small recreational fishery in Diego Garcia.
The longline fishery in Chagos waters was active year-round, mainly under Taiwanese and Japanese flagged vessels targeting large pelagic species, including yellowfin tuna (Thunnus albacares) and bigeye tuna (Thunnus obesus), swordfish (Xiphias gladius), striped marlin (Tetrapturus audax) and Indo-Pacific sailfish (Istiophorus platypterus), with annual catches ranging from 371–1366 tonnes over the last 5 years (Koldewey et al., 2010). The purse-seine fishery targeted yellowfin and skipjack tuna (Katsuwonus pelamis) and was highly seasonal, operating between November and March with a peak usually in December and January (Mees et al., 2009). Log book records show that catches, mainly by Spanish and French flagged vessels, were highly variable, ranging from less than 100 to around 24 000 tonnes annually over the last 5 years (Koldewey et al., 2010).
The Mauritian inshore fishery targeted demersal species, principally snappers, emperors, and groupers, and logbook records indicated that catches were between 200 and 300 tonnes per year for the period 1991–1997, decreasing to between 100 and 150 tonnes from 2004 (Mees et al., 2008).
The recreational fishery on Diego Garcia is much smaller, taking (in 2008) 25.2 tonnes of tuna and tuna-like species (76% of the catch), the remainder being reef-associated species (Mees et al., 2009).
As with most fisheries, those in Chagos suffered from poor documentation of by-catch and illegal fishing. By-catch was inadequately recorded through a log book system supported by limited observer coverage – mean observer coverage was 1.24% per season for longline fishing and 5.56% for purse-seine fishing (Koldewey et al., 2010). Even with this uncertainty, the by-catch in the Chagos was clearly substantial, particularly for sharks, rays, and billfish (Pearce, 1996; Anderson et al., 1998; Roberts, 2007; Graham et al., 2010; Koldewey et al., 2010). There is also evidence of marked harvesting effects on holothurian (sea cucumber) populations as a result of poaching (Price et al., 2010).
Illegal fishing remains a management issue following the implementation of the MPA and enforcement will be key to its effectiveness. Reef sharks in Chagos have declined by over 90% in a 30 year period (1975 to 2006), attributed primarily to poaching by illegal vessels (Anderson et al., 1998; Graham et al., 2010). The size and location of Chagos as an MPA is particularly important as the western Indian Ocean has some of the most exploited, poorly understood, and badly protected and managed coastal and pelagic fisheries in the world (Kimani et al., 2009; van der Elst et al., 2005), while overall catches continue to dramatically increase (FAO, 2010). Chagos is within the remit of the Indian Ocean Tuna Commission (IOTC), although this Regional Fisheries Management Organization (RFMO) is recognized to have numerous legal and technical weaknesses (Anon., 2009). Tuna in the Indian Ocean are considered to be close to the maximum sustainable yield (bigeye) or overexploited (yellowfin) and even skipjack, which is generally considered a highly productive and resilient species, has been highlighted for close monitoring (IOTC, 2010). Illegal, unreported and unregulated fishing is not a trivial component of the catch and adds substantial uncertainty into assessments (Ahrens, 2010).
There is increasing evidence that large MPAs like Chagos can benefit pelagic species that exhibit highly mobile behaviours (reviewed in Game et al., 2009; Koldewey et al., 2010). In fisheries management, the phrase ‘highly migratory’ often has little biological meaning, with studies of tuna mobility demonstrating they would benefit from national-level closures (Sibert and Hampton, 2003). Pelagic fish demonstrate considerable stability and persistence, and predictability of some habitat features does occur within the pelagic realm (Hyrenbach et al., 2000; Baum et al., 2003; Worm et al., 2003; Etnoyer et al., 2004; Alpine, 2005). Migratory predators like tuna do not move randomly, but associate with certain environmental and/or physical features (Hughes et al., 2010; Schaefer and Fuller, 2010), meaning that positive, measurable reserve effects on pelagic populations exist (Hyrenbach et al., 2002; Roberts and Sargant, 2002; Baum et al., 2003; Worm et al., 2003, 2005; Jensen et al., 2010). Several studies have shown that migratory species can benefit from no-take marine reserves (Polunin and Roberts, 1993; Palumbi, 2004; Beare et al., 2010; Jensen et al., 2010).
Pelagic MPAs are an important tool in marine conservation management (Game et al., 2009) and are rapidly becoming a reality (Pala, 2009), although some of the challenges relating to their implementation may be both costly and difficult (Kaplan et al., 2010). Large MPAs are considered necessary to protect migratory species such as large pelagic fish and marine mammals (Wood et al., 2008) as well as offsetting the concentration of fishing effort outside them (Walters, 2000) and maintaining ecological value (Nelson and Bradner, 2010). Their importance for top predators has been highlighted by the most comprehensive, decade-long, open ocean tagging study in the Pacific that clearly demonstrated that top predators – including whales, seals, tuna, sharks, seabirds and turtles – exploit their environment in predictable ways, providing the foundation for spatial management of large marine ecosystems (Block et al., 2011). Extending to 200 nm, the Chagos MPA offers an extremely valuable opportunity to understand the effects of large-scale protection on pelagic, migratory species, both within the MPA and within a regional context. Ranges of skipjack and yellowfin tuna have not been measured in the Indian Ocean, but if their ranges in a Pacific archipelago (Sibert and Hampton, 2003) are superimposed onto the Chagos MPA, it is seen that the latter encompasses as much as the median lifetime displacement of these two key species (Figure 10).
Figure 10.

The median lifetime displacement of skipjack (red) and yellowfin tuna (yellow), superimposed on a map of the Chagos MPA
BIOLOGICAL CONNECTIONS OF CHAGOS IN THE INDIAN OCEAN
To the east of Chagos, there is no shallow water until the Cocos-Keeling islands 2750 km to the (ranges from Pacific: Sibert and Hampton, 2003). east, with Indonesia another 1000 km further on. To the west, distances to shallow reefs are much less, 1700 km to the Seychelles and only 1050 km to the commonly overlooked Saya de Malha submerged banks at the northern end of the ‘Shoals of Capricorn’ between the Seychelles and the Mascarenes (Figure 11). Ocean currents passing across Chagos flow towards south-east Asia from approximately January to April, and towards the western Indian Ocean for much of the rest of the year, with fluctuations (Couper, 1987). At a speed of 0.5 m s-1 (Bonjean and Lagerloef, 2002) planktonic larvae from reef species would need 65 days to reach shallow habitat in the east, but only 35 and 25 days to reach the Seychelles and Saya de Malha reef systems, respectively, well within the pelagic larval duration of many reef organisms. Due to its location, Chagos is thus likely to be an important ‘stepping-stone’ for marine organisms in the Indian Ocean.
Figure 11.

Reef substrate or limestone banks within the photic zone in the central and western Indian Ocean.
Fifteen years ago, mapping methods in which geographical distances were replaced by similarities of coral presences, showed that Chagos does appear to function as an east–west stepping stone for corals (Sheppard, 1999c). Another study has shown recent colonization of a fish species from the east, consistent with this stepping-stone function, especially with reefs in the southern part of the group (Craig, 2008).
Genetic programmes to examine connections between Chagos and other Indian Ocean reef sites have been initiated recently for numerous species, including about 24 reef fish species and several invertebrates. For hawksbill turtles (Eretmochelys imbricata), genetic linkages were demonstrated for nesting females and foraging juveniles between Chagos and Seychelles, but no linkages were demonstrated with hawksbill rookeries of Western Australia (Mortimer and Broderick, 1999; Mortimer et al., 2002). In the wider Indian Ocean, Vargas et al. (in press) subsequently identified nine genetic groupings, with those nesting in Chagos and Seychelles forming a single grouping distinct from those in the Arabian Gulf and from easterly sites including Western Australia (Vargas et al., in press). Analyses of DNA indicate that most foraging hawksbills in Chagos derive from rookeries in Chagos and Seychelles, which also contribute substantially to foraging aggregations in Cocos Keeling (FitzSimmons, 2010, unpublished report). Although most mtDNA haplotypes found in the Chagos and Seychelles were not found elsewhere, some uncommon haplotypes were identical to those observed from Iran, Oman, and Australia, supporting the stepping stone model.
The crown-of-thorns starfish, an important coral predator, was previously believed to be a single species, Acanthaster planci, but Vogler et al. (2008) have shown that the species includes four highly differentiated lineages with restricted distributions, which together form a species complex. Two of these lineages are found in the Indian Ocean, and data indicate (Figure 12(a)) that crown-of-thorns starfish from Chagos belong to the Southern Indian Ocean lineage. A more detailed phylogeographic study (Vogler et al., in prep.) reveals that there is high gene flow among populations of the Southern Indian Ocean lineage, indicating high connectivity among these geographically distant populations. In other parts of the Pacific, larvae have been found to extend their developmental period to seven weeks in marginal food regimes (Lucas, 1982). Although the occurrence of a facultative teleplanic larva remains to be confirmed (Birkeland and Lucas, 1990), the low productivity found over most of the southern Indian Ocean (<130 gC m−2 day−1; Reid et al., 2006) could result in extended larval durations there too, and hence the observed high connectivity. This would contribute to the low levels of genetic structure observed in the Southern Indian Ocean lineage, despite the geographic distances among populations.
Figure 12.

(a) Crown of thorns genetic groupings. (b) peacock hind (Cephalopholis argus). (c) brown surgeonfish (Acanthurus nigrofuscus). (d) coconut crab (Birgus latro). Colour coding for the crown of thorns (Vogler et al., 2008, in prep.) and peacock hind (Gaither et al., 2011) indicate distinct genetic lineages. Dashed lines for the brown surgeonfish (Eble et al., 2011) indicate genetically independent populations. Photo credit: www.aquaportail.com. Image 12(b) and 12(c) reprinted from Gaither et al. (2011) and Eble et al. (2011) with permission from the authors. For (d) solidity of arrow lines represents relative amounts of gene flow, so that for this terrestrial crab flow is mainly eastwards during the Equatorial Counter Current flow.
Genetic surveys of reef fish species (Eble et al., 2011; Gaither et al., 2011) show an affinity with the western Indian Ocean. The peacock hind (Cephalopholis argus, Figure 12(b)) and brown surgeonfish (Acanthurus nigrofuscus) (Figure 12(c)) demonstrate genetic similarity within sites in the western Indian Ocean and much less similarity with sites further east.
Preliminary examination of the coral Platygyra daedalea with five microsatellite loci, including samples from Chagos (Macdonald et al., personal communication), revealed the intuitive result that, while the Chagos population had the lowest allelic diversity among the sites studied, it proved to be a source of genetic diversity for this species. The role of Chagos as a stepping stone between the east and the west of the Indian Ocean, or a recipient of larvae, is further suggested by coral species diversity patterns (Obura, unpublished; Figure S6 in Supplementary Material). Based on consistent field samples from 2002–2010, the coral fauna of Chagos is more similar to that of the western Indian Ocean continental coastline, including northern Madagascar, than it is to the smaller and more dispersed islands in the central Indian Ocean (Seychelles, Mauritius, Reunion). In terms of species richness it groups with the continental sites, potentially due to both connectivity and habitat area (Figure S6 in Supplementary Material).
Despite being geographically part of the Indian Ocean, the eastern Indian Ocean locations at Cocos-Keeling and Christmas Islands, and Western Australia are more closely affiliated with the Pacific ichthyofauna, with only 5% of species at Cocos-Keeling being exclusively of Indian Ocean origin (Allen and Smith-Vaniz, 1994). The latter islands are considered to be a part of the Indo-Polynesian Province stretching from the eastern Indian Ocean to Easter Island (Briggs and Bowen, 2012) and have been shown to be sites of hybridization between Indian and Pacific Ocean populations of reef fishes (Hobbs et al., 2009). Exceptions to this pattern include a dispersive snapper (Lutjanus kasmira; Gaither et al., 2010), trumpetfish (Bowen et al., 2001) and two moray eels (Genus Gymnothorax, Reece et al., 2010) which freely intermix across all their Indo-Pacific range and Chagos may act as a bridge between western Indian Ocean and Pacific populations of these species.
For the coconut crab, Birgus latro, which is terrestrial but which breeds in the sea, mitochondrial genetic work has compared Chagos with sites in the Seychelles and East Africa, and showed that the Chagos population was significantly differentiated (P <0.05) from Seychelles and East African populations (Table S1 in Supplementary Material). Asymmetric gene flow, favouring migration from East Africa to Seychelles, and Seychelles to Chagos, comes from estimates on direction and mean number of migrants per generation between regions. The rate of immigration to Chagos from the west was measured at about five effective females per generation (breeding age commences after about 5 years), using a measured mean effective female population size in the study of about 3000, or about 0.1–0.2% per generation (NB this is not the counted population of individuals, which is orders of magnitude greater). Thus for this species, Chagos receives more larvae from the west than flow from Chagos to the west (Figure 12(d); Tables S1–S3 in Supplementary Material) partly because egg release coincides with the period of current flow towards the east, and there is a high level of genetic connectivity. Additionally, a strong genetic connectivity among three sites was also seen through population structure analysis. The data also show that there is a clear differentiation between the Indian Ocean clade and the west Pacific clade (Figure S7 in Supplementary Material).
Taken together, these results confirm that Chagos is part of the western Indian Ocean province as described by Briggs (1974), although Briggs and Bowen (2012) comment that it has faunal affinities with the Indo-Polynesian province, with respect to fishes, as well as to the western Indian Ocean. Interestingly, Chagos shows less connectivity in some groups than might be expected (considering the much shorter distances) with the much closer Maldives to the north, which may be a function of the predominantly east–west currents. The pattern is clearly complex: earlier fish surveys (Winterbottom and Anderson, 1997) in the Chagos Islands delineated the archipelago into two distinct assemblages, with the northern portion sharing affinities with the eastern Indian Ocean and the southern portion (including Diego Garcia) more closely aligned with faunal assemblages further west.
Thus the results from the genetic and distribution data indicate that Chagos is an important biogeographic crossroad between the eastern and western Indian Ocean. The so-far limited molecular data show that the distances between Chagos and the western banks and Seychelles is much less of a barrier than is the much larger expanse of water to the east. Development of so-called ‘teleplanic larvae’ by which many species show greatly expanded larval durations, especially in conditions of low nutrients (such as exist in the central Indian Ocean) and lack of suitable substrate, has been long recognized (Scheltema, 1988). Some coral larvae may be competent for up to 105 days (Wilson and Harrison, 1998), and while the pelagic larval duration of reef fishes averages about one month, it varies enormously (Brothers and Thresher, 1985; Sale, 2002). Although it is probable that Chagos is an important stepping stone in the western Indian Ocean, the rate at which this happens for most groups is still not known (though values show appreciable mixing of the island-requiring coconut crab as noted above). In fact, the number of migrants needed to maintain genetic coherence between populations is small (Slatkin, 1977, 1982). As noted by Hellberg (2007), for management purposes we need to know whether or not connections are made every several thousand generations, or if only a single founding event occurred, or whether connections occur in ecologically frequent intervals. Patterns of connectivity as they exist today are especially important for designing management strategies to restore and conserve reef populations. If Chagos is mainly a net recipient of larvae then its rich and relatively undamaged state affords it a very high conservation value. If Chagos is also a source of biological diversity for the over-exploited sites further west, then its value would be even greater.
ISLANDS OF THE MPA
Because of its relatively large land size (29.7 km2) Diego Garcia has been the site of most terrestrial research (Stoddart and Taylor, 1971). During its plantation period, Diego Garcia, along with most other islands of Chagos, was heavily planted for coconut at the expense of the native plant communities. Stoddart (1971) expressed surprise at the large proportion of land area used for coconut production: ‘…almost the whole area of the atoll (6250 out of 7488 acres) was being cropped for coconuts’ and
“Little attention has been paid at Diego Garcia to conservation: the atoll has simply been used as a supplier of coconut products, and to a lesser extent of dried fish and turtles, for Mauritius. Both the Green and Hawksbill turtle used to nest here in some numbers… The early settlers found the frigate birds, boobies, noddies, terns, herons and tropicbirds to ‘breed on these islands. They are considered good eating; the feathers too, make excellent bedding’ (Anon. 1845, 483).”
The island was severely damaged in ecological terms. Guano mining and habitat destruction accompanying the plantations destroyed most birds and nesting habitats, including some huge tern colonies, along with other species now listed in the IUCN Red List such as turtles and coconut crabs (IUCN, 2011). Stoddart reports that the first practical conservation measures were taken by a manager in the 1870s, but ‘in the absence of enforcing authority or of any clear need for conservation it is unlikely that much attention was paid to it.’ The same lack of conservation ethic applied to other Chagos atolls too, as with coral islands throughout the ocean.
The military facility was constructed mainly in the early 1970s. It was built on former coconut plantation – little or no intact natural vegetation or bird colonies remained. Partly because of conservation requirements developed over the last 20 years, Diego Garcia has several conservation sites and issues of its own (Figure 13). It is one of the most enclosed atolls in the world, being open in the north only, with three small islets in the mouth. The military facility is located on the western arm. The eastern arm is part of a Ramsar site (JNCC: Ramsar Site UK61002), which extends 3 nm to seaward, encompassing most of the lagoon also. There are four Strict Nature Reserves: on the eastern arm south to line ‘A’ in Figure 13, and around the three islets, into which access is prohibited. Access to the part of this arm south of line ‘A’ is permitted. ‘Turtle Cove’ has special conservation status and restrictions because of its large aggregations of foraging juvenile and subadult hawksbill turtles, which have been the subject of mark–recapture, genetic analysis, blood hormone analysis, and population research (Mortimer and Broderick, 1999; Mortimer and Crain, 1999; Mortimer and Day, 1999; Mortimer et al., 2002).
Figure 13.

Map of Diego Garcia atoll. The military facility is on the western arm. Gray line shows the Ramsar site, which encompasses most of the lagoon, extending seaward on the eastern side. The eastern arm south approximately to the line where the Ramsar boundary intersects the coast is a ‘Nature Reserve’ which covers land only. From the top of the eastern arm to the line marked ‘A’, plus the three circled islets in the mouth of the atoll, are ‘Strict Nature Reserves’, which each cover land plus the sea area extending out 200 m from shore.
Underwater, the northern and deepest third of the lagoon was subjected to significant blasting and removal of surface-reaching coral growths, to create the present large anchorage. This caused much damage in that area at that time, though a brief survey of eight locations in 1979 (Sheppard, 1980b) showed that by that time coral cover had recovered in areas where there was no anchoring, being similar to values in the northern atolls where no activities of any kind had occurred for several years. During the 1970s, there was no indication that coral growth was other than vigorous in most of the world, though the extent of the lagoon recovery was more rapid than expected. To the authors’ knowledge, no further dredging or blasting activities have taken place after that initial period.
Pollution monitoring
Extensive pollution monitoring takes place in Diego Garcia. ‘Final Governing Standards’ and routine procedures require regular analyses in US laboratories of over 100 metals and organic substances according to US operating procedures. Almost all analyses report levels below detectable or reporting limits. None have been found to be of concern, including oil and oil residues (data held by BIOT Administration). In addition to the US monitoring, several specific projects have set out to measure emerging compounds of particular interest or concern (see Supplementary Information text for more detail about all substances summarized below).
Hydrocarbons have been particularly focused upon, and most have a natural origin (Readman et al., 1999). There was negligible evidence of contamination from petroleum. Polycyclic aromatic hydrocarbons (PAH) were similarly very low, and Readman et al. (1999) also found no evidence of sewage contamination. PCBs and organic pesticides were mostly below instrument detection levels. Similarly, extremely low levels occurred for polyfluorinated compounds, brominated, chlorinated and organo-phosphorus flame retardants, fluorinated tensides, and surfactants (PFOS) (Wolschke et al., 2011). Antifouling booster biocides and triazine herbicides (Guitart et al., 2007) also were negligible, with levels generally below the limit of detection. The same pattern occurred with most metals, though of interest is that some elevated copper was detected in 1996 in some northern waters, attributed to copper in fungicides used in coconut agriculture several years earlier (Everaarts et al., 1999). Comparisons with Antarctic and remote deep sea samples for many showed this area to have the least chemical contamination so far recorded (Supplementary Information).
There has been no oil or tar seen on Diego Garcia beaches to date, although a little has been seen on some northern islands. Lagoon water near ships is likewise monitored and is also devoid of these substances. In summary, from a chemical contaminant perspective, the marine environment surrounding the Chagos Archipelago can be considered to be near pristine and in chemical pollution terms, Diego Garcia is likely to be the cleanest inhabited atoll in the world.
Shoreline debris
Despite their near pristine chemical status, Chagos beaches have a surprisingly high number of pieces of debris. Observations were made in 1996, 2006, and 2010 at 20 sites in the outer atolls, and one in Diego Garcia (Price, 1999; Price and Harris, 2009). Median scores of the number of litter pieces were high in all years; >1000 items per 500 m linear beach. Items were mainly plastics, polystyrene (Styrofoam) and rope, much being lost fishing gear or debris discarded from ships, most commonly of south-east Asian origin. Levels in Diego Garcia in all years were two orders of magnitude less than in other atolls, reflecting periodic clean-up events in that inhabited atoll. The method did not determine size categories or weight; most items were a few cm in size or less, but several northern islands appear to collect substantial volumes of larger flotsam. Similar numbers are found in remote Pacific atolls (Price and Harris, 2009) where ocean current gyres are the main transport vector. Driftwood and lost timber from ships was low on beaches in all years, but decreased over time from 1996 to 2006, attributed to use for fuel by illegal fishing camps on the islands during this period of increasing fishing pressure (Price and Harris, 2009; Price et al., 2010). Oil slicks were never seen, but tar balls were seen at eight beaches in 1996, at three in 2006 and at one in 2010, none being seen in Diego Garcia. The decrease may reflect improved international ship ballast cleaning measures over that period (Price, unpublished data).
Terrestrial flora
Being geologically young, remote, isolated, flat, and never connected to a continental land mass, the Chagos islands have a naturally impoverished native flora. There are probably 45 native species of higher plants, 41 flowering plants (12 trees) and four ferns, none of which are endemic (Topp and Sheppard, 1999), most having a widespread distribution across the region. There are several important native vegetation community types with examples remaining intact, especially on islands too small to have been used in plantation times. These include Pisonia forest and Lumnitzera mangroves on Moresby Island; Scaevola-Argusia beach communities on many islands and Calophyllum thickets on Ile Takamaka. Most plants, especially on Diego Garcia, were introduced for fruit or ornament, or accidentally as unwelcome passengers of incoming cargo (Topp and Sheppard, 1999). Some species are problematic. The native dodder (Cassytha filiformis), a parasitic vine, is spreading rapidly and over-topping some of the native vegetation. Pipturus argenteus, a shrub whose origin is in doubt, is spreading widely and is having some negative impacts on regenerating native trees such as Pisonia grandis (Clubbe, 2010).
Introduced plant species are far more numerous than the native species. An updated vascular plant checklist (Clubbe, unpublished data; Hamilton and Topp, 2009) shows that 232 non-native species have been recorded, with 128 listed as only occurring on Diego Garcia (Hamilton and Topp, 2009). Of these, few show signs of invasiveness and most are unlikely to pose significant problems. However, there are some species where control measures are needed. Most significant is coconut (Cocos nucifera), which was widely cultivated previously. Young palms form a dense understorey which exclude virtually all other plants. Control measures underway in a pilot scheme on Diego Garcia involve the removal of palms to enable regeneration of native species from either a recently established seed bank or from those few plants that are able to survive in the low-light environment created by the coconuts. Regeneration is supplemented by enrichment planting of key native tree species including Barringtonia asiatica, Cordia subcordata, Guettarda scabra and Calophyllum inophyllum, all of which provide important nesting sites for seabirds. In Diego Garcia, there is a ‘two for one’ policy whereby any coconut or other tree removal is replaced by two hardwood trees collected from seedlings beneath established trees or from the seed bank. If successful, this approach will be applied to northern islands also. Some species of introduced plants are spreading invasively, some associated specifically with former settlement, e.g. Tabebuia heterophylla, while others not specifically associated with settlement areas, e.g. Casuarina equisetifolia, exist on several islands. The latter, introduced for timber in plantation times, poses a threat to native trees on some of the islands since it exerts a tenacious hold, out-competing other trees owing to the ability of its roots to fix nitrogen and because of the litter it creates.
More work is needed to fully understand the distribution of non-native species and the potential impact of invasive plants in Chagos. Preliminary analyses indicate, however, that the spread of non-native species from Diego Garcia to other atolls is slow and there has not been a noticeable increase in the number of non-native species on the outer islands since the 1980s (Clubbe, 2010).
Regarding algae, mosses, liverworts, cyanobacteria, fungi, and lichens, diversity is relatively low in Chagos, although these provide high cover and are ecologically important (Seaward, 1999; Seaward and Aptroot, 2000; Watling and Seaward, 2004; Seaward et al., 2006). Cyanobacteria extensively clothe limestone, thin sandy soils and tree trunks, and fix nitrogen directly in the otherwise poor soils. The mosses and liverworts are important in stabilizing soils, and are often the primary stages in the succession of higher vegetation in exposed areas. In wooded areas, the mosses and liverworts clothe the bark of living trees; the nature of this epiphytic flora changes with age; as the trunks die, different mosses and liverworts support the decomposition process undertaken mainly by fungi. Lichens colonize all available tree bark surfaces, as well as being found on the living leaves of various tree species, the long-lived evergreen nature of the leaves allowing these slow growing organisms to establish themselves.
There are no endemic species of these groups. However, due to the geographical setting of the islands their establishment is remarkable, not only in terms of the distances travelled by the spores and propagules, but also in terms of their origin, and there are interesting affinities of these floras with south-east Asia, India or Africa, as yet to be fully determined. A new liverwort, Cololejeunea planissima var. chagosensis, known only from Chagos (Seaward et al., 2006), is likely to occur elsewhere, but there is always the possibility that it has disappeared (or will disappear) from its original site(s).
Terrestrial fauna
There are no native mammals, amphibians or endemic birds. Of invertebrates recorded to date, most are widespread species with distributions across the Indo-Australian tropics and include some human commensals (Barnett and Emms, 1999). There are thought to be three endemic sub-species and one endemic species of Lepidoptera (Barnett and Emms, 1999).
The black rat (Rattus rattus) is the main invasive animal, and was reported to be of plague proportions as early as the late 1700s. Only smaller islands which were not planted have no rats and have numerous birds, and even islands close to rat-infested islands remain uninvaded.
Other species introduced at various times in the islands’ history include sheep Ovis aries, cattle Bos sp., horses Equus ferus, donkeys Equus africanus asinus and (having the worst impact on the birds), dogs Canus lupus., pigs Sus sp., cats Felis catus, as well as the rats (Edis, 2004; Carr, 2011a). Most have gone, but remaining invasive animals include the cane toad (Bufo marinus), feral cats, donkeys and several birds (Carr, 2011b). Dates of introduction for most terrestrial invasives are unknown, one exception being the garden lizard (Calotes versicolor) in Diego Garcia. This was first observed on Diego Garcia in May, 2001, and is believed to have been introduced via cargo. It is arboreal, diurnal and adaptable (Daniel, 2002) and is one of the most widespread non-geckonid lizards in the world (Matyot, 2004). It has a population density of 172 lizards ha−1 (95% CI: 154–237) in the inhabited area, has since spread south and is likely to spread island wide. This illustrates the rapidity at which such species may spread. Since that time, strict quarantine measures have been introduced for imported materials, including rock for construction of hardened shorelines to combat shoreline erosion.
Seabirds
In contrast to the generally poor terrestrial fauna, the breeding seabirds of the Chagos are of international importance (Carr, 2006, 2011a). Islands never planted, inhabited or rat infested give a glimpse of what the islands used to look like because significantly higher bird abundances occur on these (Symens, 1999; Price and Harris, 2009).
Population trends and the seabirds’ breeding phenology are poorly understood. Comprehensive censuses were undertaken of breeding seabirds in February/March 1996 (Symens, 1999) and again in March 2006 (McGowan et al., 2008; Figure S8 in Supplementary Material). Eighteen seabird species breed in Chagos (Carr, 2011a), five in globally important numbers, resulting in ten islands being classified as Important Bird Areas (IBAs) (Carr, 2004, 2006, 2011b), with two further islands being proposed as IBAs (McGowan et al., 2008). Recently more regular monitoring of the Chagos IBAs showed that many species of seabird either breed continuously throughout the year with spikes in breeding in certain months, e.g. red-footed (Sula sula) and brown (Sula leucogaster) boobies, or breed at different times of the year on different atolls, e.g. lesser noddy (Anous tenuirostris) and sooty tern (Sterna fuscata) (Figure S8 in Supplementary Material). This realization of continuous breeding has increased the estimates of some of the breeding populations of some species, making the Chagos seabirds an even more important asset than was previously believed, to both the Indian Ocean and globally. Diego Garcia and its three associated islets hold one of the Indian Ocean’s largest breeding colonies of red-footed booby, which has grown from three or four pairs in 1984 to over 5000 pairs in 2011, spread over 30 km of shoreline (Carr, 2011b). This growth may be attributable to the area being out of bounds to all personnel since 1984.
The suggestion has been made that the ten IBAs should be clustered into four grouped IBAs, which would ‘…be consistent with IBA philosophy in designating inter-connected, proximal, protected habitats for species that … appear to utilise different islands for breeding in different seasons’ (Carr, 2011b). These four would consist of Diego Garcia, the Western Great Chagos Bank Islands, Nelson’s Island in the northern Great Chagos Bank, and the eastern islands of Peros Banhos atoll. To initiate successful conservation management plans for seabirds, a thorough understanding of their breeding phenology is essential, including breeding triggers and dietary requirements. Long-term, scheduled monitoring is required to achieve this, in order to preserve and protect one of the last seabird strongholds in the Indian Ocean.
Coconut crabs
The coconut or robber crab (Birgus latro) has a wide distribution throughout the Indian and Pacific Oceans but is usually over-harvested. The largest land dwelling invertebrate in the world, it can exceed 5 kg. They mate on land but the female releases eggs in the ocean after a few months, where they immediately hatch, the oceanic larval stage usually lasting 2–3 weeks (Fletcher and Amos, 1994). Once on land their growth is slow; they may be mature at 5 years but it takes approximately 8–10 years (depending on diet quality) for crabs to reach a size of ~500 g, with maximum size at 40 years (Fletcher and Amos, 1994).
Many of the northern islands in the 1970s had very few coconut crabs, and they were virtually absent from some. Those seen had a size positively correlated with the length of time that the island had been uninhabited, ranging from the 1930s when the first two of the five atolls were abandoned, to the 1970s (Sheppard, 1979). In 2010 in Diego Garcia, crab populations showed an overall average density in the Ramsar area of 298 crabs ha−1 (95% CI: 229–387, Figure S9(a) in Supplementary Material), the highest average number ever recorded, with two transects containing 467 and 489 crabs ha−1 (Vogt, unpublished). The southern tip of the island had an average 147 crabs ha−1 (95% CI: 95–226), while the side containing human habitation had only 39 crabs ha−1 (95% CI: 24–63) indicating capture despite regulations, and the inhabited side also lacked crabs in the largest size classes (Figure S9(b) in Supplementary Material). The Ramsar side appears to be unharvested, a rarity for this species.
Turtles
Hawksbill (Eretmochelys imbricata) and green turtles (Chelonia mydas) nest in Chagos, and surveys conducted in 1996 and 2006 recorded both species at each of the five atolls. Estimated numbers of females nesting annually were 400–800 green turtles and 300–700 hawksbills in 1996 (Mortimer and Day, 1999). Diego Garcia, by far the largest island in the group, has nesting populations of both species that rival those of any of the other atolls, and nesting density and habitat quality are particularly good along the east coast.
Historical records indicate that nesting numbers of both species declined significantly over the period of human settlement (Mortimer and Day, 1999; Mortimer, 2009). In 1786, more nesting green turtles were captured at Salomon atoll in 4 days than were estimated to nest there annually in 1996 (Mortimer and Day, 1999). Likewise for hawksbills: trade statistics show that at the turn of the 20th century nearly as many were killed annually to provide shell for export, as were estimated to nest each year in 1996 (Mortimer, 2009; Figure S10 in Supplementary Material). A comparison of nesting census data from 1996 and 2006 (Mortimer, 2007) indicates little change in hawksbill nesting activity in the northern atolls, but a possible increase at Diego Garcia. Levels of green turtle nesting recorded at all island groups were greater in 2006 than in 1996. However, this does not necessarily indicate population increase, given that green turtle nesting activity oscillates from year to year and trends can only be discerned when nesting activity is monitored consistently over extended periods of time (usually more than 10 years).
Island prospects, erosion and sea levels
Though tiny in total area compared with marine habitats, the islands are central to conservation measures. Much has been discussed recently about loss of coral islands from climate change; indeed the nearest archipelago, the Maldives, has taken a lead among small nations in this matter. Controversy about island erosion and sea level rise has been strenuous because the subject has many apparent contradictions.
Sea-level rise in the southernmost atoll Diego Garcia has been recorded by tide gauges since 1988 (Figure S11 in Supplementary Material). The data are in two series, (1988–2000 and 2003 to present), the former being ‘research grade’, the latter being ‘fast delivery’ data only at present. Both series have gaps and, while the graph shows a substantial rise, both series are short. The two nearest tide gauges to Chagos are on Gan in the southern Maldives, 475 km north at about 1°S and on the capital Malé at about 4°N (Singh et al., 2001). These show rises of similar order to that in Diego Garcia. Further, there are marked seasonal differences in monthly averages, both in Diego Garcia and in the Maldives, which Singh et al. (2001) link to the monsoon cycle. Highest monthly average tides in Diego Garcia occur in November–February, and the lowest ones between April and July.
Newer satellite altimetry data shows a much smaller rise compared with tide gauge measurements for the central Indian Ocean (www.sealevel.colorado.edu), including both Chagos and the Maldives. Local land movements may account for an unknown proportion of the discrepancy between gauges and satellite altimetry, but whether gauges or satellite altimetry are more correct, it is high tides, not monthly averages, which cause flooding, and several other oceanographic and climate driven changes may be more important. Further, different atolls may respond slightly differently to Diego Garcia in terms of relative sea level rise because the area is tectonic, lying on a minor plate margin (Figure S12 in Supplementary Material). Local effects are also dependent on supply of sediments, which is likely to have changed markedly following the massive coral mortality of 1998.
The important immediate consideration is erosion of shores and inundation by the sea, rather than sea-level rise per se. Coral islands have long been known to have mobile shores because of natural annual or decadal shifts in sedimentation driven by climate, which affects wave height, swell, and swell direction (Young et al., 2011 in press) as well as sea level. Chagos has suffered considerable overall erosion of many shorelines, most notably in the last decade. This has not yet been studied in detail, although in Diego Garcia it is most noticeable if only because of the high cost of sea defence measures employed there. In parts of the northern atolls, horizontal attrition may exceed 1 m yr−1 in several areas although this high rate is episodic and spatially patchy. On several islands, remnants of mature coconut trees are now in the intertidal zone, and pits dug for coconut cultivation are now at or below high tide levels in some places, all indicating net erosion over the past 100–150 years in such places. Blenheim atoll in the north used to have three low sandy islands, with low bushes (report by Archibald Blair, 1787) but this atoll is now submerged, with only some sections of its western rim emergent at low tide. This atoll’s coral and algal ridge growth are as prolific as on nearby islanded atolls, so a deficiency in reef growth is an unlikely cause of the drowning. In contrast, Egmont atoll, was known in the 19th century as the ‘Six Islands’ (each named separately), yet today most have fused to form only three islands at low tide, possibly because of tectonic movement or because of changing sedimentary patterns, although erosion as described above also occurs in some sectors. Coral mortality from warming, discussed earlier, is likely to lead to massive if temporary changes in sand production, known to have had marked consequences on shorelines elsewhere in the Indian Ocean for example (Sheppard et al., 2005).
Island heights and cross-sections are likely to be important to future events. The highest points of probably all islands occur on the rims. Profiles of several Chagos islands (Figure 14) show a central depression, caused by acidic rain eroding the coral limestone rock (Sheppard, 2002), and by accumulation of sand around island rims. Future erosion patterns are unlikely to be smoothly progressive, and may be episodic. There is evidence of this in several locations, and a broach could destroy the freshwater lens in that area. Figures 14(d)–(e) also shows potential inundation levels in two islands in two northern atolls, determined by Royal Haskoning (2002) in a resettlement feasibility study. In Diego Garcia atoll also, similar central depressions are evident on both western and eastern sides, although much of the eastern arm is very narrow and low lying too, such that extensive flooding occurs at high spring tides.
Figure 14.

Cross-sections of (a) Sipaille, (b) Lubine, (c) Sudest islands in Egmont atoll, (d) Isle de Coin on Peros Banhos atoll, and (e) Ile Boddam on Salomon atoll. Y-axis is metres above mean high tide in (a)–(c), and from a relative datum in (d)–(e). (a) Lagoon is on right, total width is 500 m, (b) lagoon on right, total width 450 m, (c) lagoon is on left, total width 250 m. (d) and (e) have no horizontal dimensions but are larger islands. Solid horizontal lines are mean high water springs. (a)–(c) from Sheppard (2002), (d)–(e) from Royal Haskoning (2002). Dashed lines in (d)–(e) are estimated flooding levels following severe storm overtopping.
A further phenomenon, noted over 100 years ago but which has been largely overlooked, is that of ‘self-gravitation’, whereby the large volume of ice added to the poles creates significant gravitational attraction of its own (Vermeersen, pers. comm.; Vermeersen and Schotman, 2009), raising water level nearer the poles. Thus, when ice melts and water redistributes, polar gravitational attraction lessens, so sea-level rise increases the further the distance from the ice sheet (along with a marked fall near the poles). The magnitudes are likely to be significant to tropical coral islands because, in the absence of the Greenland ice sheet’s gravitational attraction, sea-level rise in equatorial regions would probably be 10–20% greater than would occur from ice volume melt alone (Vermeersen, pers. comm.).
CLIMATE AND LONG TERM ENVIRONMENTAL MONITORING PROGRAMMES
Work is increasingly needed on climate change projections, yet the Indian Ocean forms a very large gap in many global monitoring programmes. Chagos is situated in a key region of climate variability. It lies at the eastern margin of an oceanic feature known as the ‘Seychelles–Chagos thermocline ridge’ (Hermes and Reason, 2008), along which the thermocline rises close to the surface and upwelling of cold water from below the thermocline occurs (Vialard et al., 2009). In contrast to most upwelling regions, surface water of the Seychelles–Chagos ridge is extremely warm. In austral summer, the main rainy season, sea surface temperature (SST) varies between 28.5 °C and 30 °C only, a range in which the atmosphere is very sensitive to small oceanic changes (Timm et al., 2005). The high SST combined with the shallow thermocline makes the Seychelles–Chagos ridge a region with very strong air–sea interactions.
However, climate observations from this region are extremely sparse due to the paucity of measurements in most Indian Ocean countries and to the remoteness of Chagos. Further, instrumental SST data are associated with errors that are as large as the SST anomalies across a range of time scales (Annamalai et al., 1999). There is currently only one weather station at Diego Garcia, whose weather records contain several gaps (Sheppard, 1999d). ‘Conventional’ infrared satellite measurements of SST are not adequate to capture important SST perturbations that occur in the rainy season when convective activity and cloud cover is highest (Vialard et al., 2009), and only the increased use of orbiting microwave instruments in the late 1990s allowed better quality SST to reveal the role of Chagos with respect to air–sea interactions (Vialard et al., 2009). Therefore, a continuous monitoring programme of important climatic and oceanographic parameters at Chagos is needed for a better understanding of air–sea interactions that are crucial in global climate issues.
The Seychelles–Chagos region has a distinct oceanic and atmospheric variability at multiple time scales, each with significant climatic consequences. It has the strongest intraseasonal SST variance in the Indo-Pacific warm pool, because the shallow thermocline is very responsive to atmospheric fluxes (Vialard et al., 2008, 2009). SST cooling of 1–1.5 °C, lasting for 1 to 2 months, may occur during austral summer, followed by a short lag and sharp increase in atmospheric convective activity (Vialard et al., 2009) associated with the Madden–Julian–Oscillation (MJO, Madden and Julian, 1994). The MJO has a time scale of 30–80 days, and it explains much of the variance of tropical convection, and modulates cyclonic activity.
On interannual time scales, the Seychelles–Chagos ridge is affected by the El Nino-Southern Oscillation (ENSO) and the Indian Ocean Dipole (IOD) (Saji et al., 1999; Webster et al., 1999). ENSO events lead to a displacement of the West Pacific warm pool and affect the Indian Ocean via the so-called atmospheric bridge. Anomalous subsidence over Indonesia during El Nino years leads to a cooling over the maritime continent and a basin-scale warming of the Indian Ocean, particularly the western part. The IOD is a coupled ocean–atmosphere instability centred in the tropical Indian Ocean that affects the climate of the countries that surround the Indian Ocean basin (Marchant et al., 2007). A positive IOD period is characterized by cooler than normal water and below average rainfall in the eastern Indian Ocean, Indonesia and over parts of Australia, while warmer water and increased rainfall is observed in the western Indian Ocean and equatorial East Africa. A negative IOD period is characterized by warmer than normal water and above average rainfall in the eastern Indian Ocean sector and cooler than normal water and below average rainfall in the western Indian Ocean. Some studies suggest that positive (negative) IOD events may be triggered by El Niño (La Niña), while others find that IOD events occur independently although they may overlap with El Niño/La Niña events, e.g. the strong 1998 ENSO (Saji and Yamagata, 2003). The IOD has two ‘centres of action’: one off the coast of Sumatra, in the eastern Indian Ocean sector, and one at the Chagos Archipelago (Saji and Yamagata, 2003). Reliable instrumental records of the IOD are currently limited to the past 50 years.
On decadal and longer time scales, information on climate variability at Chagos has been gained from geochemical proxies archived in long-lived corals (Pfeiffer et al., 2004, 2006, 2009; Timm et al., 2005). These data suggest that changes in the mean climate may influence the impact of large-scale climate phenomena at Chagos, possibly affecting climate in other countries surrounding the Indian Ocean. For example, a shift towards higher mean SSTs occurred in 1975 (Timm et al., 2005; Pfeiffer et al., 2006, 2009), after which small SST changes induced by ENSO caused much larger precipitation anomalies (Timm et al., 2005; Pfeiffer et al., 2006). Similar changes in rainfall variability have been found in South Africa (Richard et al., 2000), Sri Lanka and southern India (Zubair and Ropelewski, 2006).
The long-term variability of the IOD has also been investigated with coral proxies from the Seychelles and Indonesia (Abram et al., 2008). The coral reconstruction suggests an increase in the strength and frequency of IOD events during the 20th century, which may also influence the distribution of rainfall in the tropical Indian Ocean. The work highlights the importance of the Chagos Archipelago for climate variability research in the Indian Ocean sector and beyond, and emphasizes the need for high-quality in situ data recording of climatic parameters at Chagos.
CONCLUSIONS
The Chagos Archipelago is unquestionably a very valuable biological asset in an ocean showing significant and continuing decline. The exceptionally good condition of the Chagos marine ecosystems is important in terms of biodiversity and productivity, and for its likely biogeographic role in the ocean. Its protection is of great and increasing importance given that global pressures from overfishing and other activities resulting from human population growth are increasing. In this it joins a small handful of other large, protected sites in the world (Nelson and Bradner, 2010), and is the only one in the Indian Ocean. The importance of the Chagos MPA was further underpinned by a recent report across marine science disciplines suggesting that the world’s ocean is at high risk of entering a phase of disturbance unprecedented in human history (Rogers and Laffoley, 2011). Indeed, many more effective and very large MPAs need to be created if international goals for protecting the oceans are to be met, although the number of locations where this will be possible is diminishing. Regarding terrestrial value, several smaller islands that were not ravaged by coconut plantations remain intact and, by using them as source areas, much restoration is possible and indeed has been commenced.
The MPA therefore is needed for multiple reasons. It has met with opposition from various sources, including the oceanic fishing industry, the government of Mauritius that claims the territory, and from Chagossians who were removed approximately 40 years ago, and some of their representatives. While the decision to remove the inhabitants at that time was based on politics and defence rather than for any reasons of conservation, the present good condition of such a large area has been a fortuitous if unplanned consequence of the subsequent lack of exploitation of the area. The MPA was created ‘without prejudice’ to any future resettlement, and if resettlement does occur then management must be adequate to avoid the problems which almost all past global experience has demonstrated could too easily happen.
The high value of relatively undisturbed areas encompassing a range of functionally linked ecosystems is becoming increasingly recognized at the same time that their number world-wide is diminishing (http://www.globaloceanlegacy.org/). Whereas there are many ‘managed’ reef sites elsewhere in the tropical oceans, almost all are themselves in a poor condition compared with places like Chagos and a few Pacific sites. The ‘shifting baseline syndrome’ (Pauly, 1995) applies to marine management and in many places has led to situations where decisions are taken that are highly disadvantageous to both the natural systems and to the people dependent on them. Most reefs remain unprotected or protected in name only. The huge scale of the interconnected network of atolls and banks in Chagos, and its effective governance, are likely to become increasingly important both directly and as a scientific reference site in the Indian Ocean. BIOT has, at present and the foreseeable future, governance which will enable this situation to persist. Priorities now are management in an effective manner, whatever the political future holds for the area, so that the benefits of a well protected MPA are likely to extend to peoples and ecosystems far beyond the boundaries of the Chagos MPA.
Supplementary Material
Acknowledgments
The authors thank the Administration of the British Indian Ocean Territory for permission to visit the area on various occasions, the military commanders and personnel for much assistance on site, and to the officers and crew of the BIOT Patrol Vessel Pacific Marlin for exceptional help on all visits to atolls away from Diego Garcia. The OTEP fund provided core funds for most visits, and all scientists involved received funding from numerous sources to carry out their own programmes of work in the archipelago. For assistance with genetic work we thank Jiddawi Norriman and Mohammed Suleiman Mohammed (Zanzibar), and Nancy Bunbury (Seychelles).
References
- Abram NJ, Gagan MK, Cole JE, Hantoro WS, Mudelsee M. Recent intensification of tropical climate variability in the Indian Ocean. Nature Geoscience. 2008;1:849–853. [Google Scholar]
- Ahrens RNM. PhD thesis. University of British Columbia; 2010. Global analysis of apparent trends in abundance and recruitment of large tunas and billfishes inferred from japanese longline catch and effort data. [Google Scholar]
- Allen GR, Smith-Vaniz WF. Fishes of the Cocos (Keeling) Islands. Atoll Research Bulletin. 1994;412:1–21. [Google Scholar]
- Alpine J. BSc thesis. CSIRO Marine Research and School of Zoology, University of Tasmania; 2005. The application of closed area management in pelagic ecosystems: a viable strategy for longline fisheries? [Google Scholar]
- Anderson C, Sheppard C, Spalding M, Crosby R. Shortage of sharks at Chagos. Shark News. 1998;10:1–3. [Google Scholar]
- Annamalai H, Slingo JM, Sperber KR, Hodges K. The mean evolution and variability of the Asian summer monsoon: comparison of the ECMWF and NCEP/NCAR reanalysis. Monthly Weather Review. 1999;127:1157–1186. [Google Scholar]
- Anonymous. Report of the IOTC Performance Review Panel: January 2009. Indian Ocean Tuna Commission; 2009. [Google Scholar]
- Ateweberhan M, McClanahan TR, Graham NAJ, Sheppard CRC. Episodic heterogeneous decline and recovery of coral cover in the Indian Ocean. Coral Reefs. 2011;30:739–752. [Google Scholar]
- Barnett LK, Emms C. The insects of the Chagos Archipelago. In: Sheppard CRC, Seaward MRD, editors. Ecology of the Chagos Archipelago. Linnean Society/Westbury Publishing; London: 1999. pp. 241–251. [Google Scholar]
- Baum J, Myers R, Kehler D, Worm B, Harley S, Doherty P. Collapse and conservation of shark populations in the Northwest Atlantic. Science. 2003;299:389–392. doi: 10.1126/science.1079777. [DOI] [PubMed] [Google Scholar]
- Bax N, Hayes K, Marshall A, Parry D, Thresher R. Man-made marinas as sheltered islands for alien marine organisms: establishment and eradication of an alien invasive marine species. In: Vetich, Clout, editors. Occasional Paper of the IUCN Species Survival Commission No. 27.Turning the Tide: the Eradication of Invasives Pecies, Proceedings of the International Conference on Eradication of Island Invasives; 2002. [Google Scholar]
- Bax N, Williamson A, Aguero M, Gonzalez E, Geeves W. Marine invasive alien species: a threat to global biodiversity. Marine Policy. 2010;27:313–323. [Google Scholar]
- Bayley D. MSc thesis. University of Wales; Bangor: 2009. Video analysis of the reef structure and community composition of the Chagos Archipelago, British Indian Ocean Territory. [Google Scholar]
- Beare D, Hölker F, Engelhard GH, McKenzie E, Reid DG. An unintended experiment in fisheries science: a marine area protected by war results in Mexican waves in fish numbers-at-age. Naturwissenschaften. 2010;97:797–808. doi: 10.1007/s00114-010-0696-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ben-Haim Y, Rosenberg E. A novel Vibrio sp. pathogen of the coral Pocillopora damicornis. Marine Biology. 2002;141:47–55. [Google Scholar]
- Bensch A, Gianni M, Grébroval D, Sanders JS, Hjort A. FAO Fisheries and Aquaculture Technical Paper 522. 2008. Worldwide review of bottom fisheries in the high seas. [Google Scholar]
- Birkeland CE, Lucas JS. Acanthaster planci: Major Management Problem of Coral Reefs. CRC Press; Boca Raton, FL: 1990. [Google Scholar]
- Block BA, Jonsen ID, Jorgensen SJ, Winship AJ, Shaffer SA, Bograd SJ, Hazen EL, Foley DG, Breed GA, Harrison AL, et al. Tracking apex marine predator movements in a dynamic ocean. Nature. 2011;475:86–90. doi: 10.1038/nature10082. [DOI] [PubMed] [Google Scholar]
- Bonjean F, Lagerloef GSE. Diagnostic model and analysis of the surface currents in the tropical Pacific Ocean. Journal of Physical Oceanography. 2002;32:2938–2954. [Google Scholar]
- Bowen BW, Bass AL, Rocha LA, Grant WS, Robertson DR. Phylogeography of the trumpetfishes (Aulostomus): ring species complex on a global scale. Evolution. 2001;55:1029–1039. doi: 10.1554/0014-3820(2001)055[1029:pottar]2.0.co;2. [DOI] [PubMed] [Google Scholar]
- Briggs JC. Marine Zoogeography. McGraw-Hill; New York: 1974. [Google Scholar]
- Briggs JC, Bowen BW. A realignment of marine biogeographic provinces with particular reference to fish distributions. Journal of Biogeography. 2012 doi: 10.1111/j.1365-2699.2011.02613.x. [DOI] [Google Scholar]
- Brothers EB, Thresher RE. Pelagic duration, dispersal and the distribution of Indo-Pacific coral reef fishes. In: Reaka ML, editor. The Ecology of Coral Reefs; Symposia Series for Undersea Research. NOAA’s Undersea Research Program; Washington DC: US Department of Commerce; 1985. pp. 53–69. [Google Scholar]
- Bruno JF, Peters LE, Harvell CD, Hettinger A. Nutrient enrichment can increase the severity of coral diseases. Ecology Letters. 2003;6:1056–1061. [Google Scholar]
- Bruno JF, Selig ER, Casey KS, Page CA, Willis BL, Harvell CD, Sweatman H, Melendy AM. Thermal stress and coral cover as drivers of coral disease outbreaks. PlosBiology. 2007;5:e124. doi: 10.1371/journal.pbio.0050124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burgiel SW, Muir AA. Invasive species, climate change and ecosystem- based adaptation: addressing multiple drivers of global change. Global Invasive Species Programme (GISP); Washington, DC, USA and Nairobi, Kenya: 2010. [Google Scholar]
- Burke L, Reytar K, Spalding M, Perry A. Reefs at Risk Revisited. World Resources Institute; Washington, DC: 2011. [Google Scholar]
- Carlton JT, Ruiz GM. Vector science and integrated vector management in bioinvasion ecology: conceptual frameworks. In: Mooney HA, Mack RN, McNeely JA, et al., editors. Invasive Alien Species: a New Synthesis. Island Press; Washington, DC: 2005. [Google Scholar]
- Carpenter K, Abrar M, Aeby G, Aronson RB, Banks S, Bruckner A, Chiriboga A, Cortes J, Delbeel C, De Vantier L, et al. One-third of reef-building Corals face elevated extinction risk from climate change and local impacts. Science. 2008;321:560–563. doi: 10.1126/science.1159196. [DOI] [PubMed] [Google Scholar]
- Carr P. Important Bird Areas in Asia. Key Sites for Conservation. BirdLife International; Cambridge: 2004. British Indian Ocean Territories. (BirdLife Conservation Series No. 13) [Google Scholar]
- Carr P. British Indian Ocean territory. In: Sanders SM, editor. Important Bird Areas in the United Kingdom Overseas Territories. RSPB; Sandy, UK: 2006. pp. 37–55. [Google Scholar]
- Carr P. Birds of the British Indian Ocean Territory. NatureBureau; Newbury: 2011a. [Google Scholar]
- Carr P. Important Bird Areas – the British Indian Ocean Territory. British Birds. 2011b;104:642–666. [Google Scholar]
- Chagos Environment Network. Consultation on whether to establish a Marine Protected Area in the British Indian Ocean Territory. 2010 protectchagos.org/wp-content/uploads/CEN_Submission-Final-FINAL.pdf.
- Clark MR, Tittensor D, Rogers AD, Brewin P, Schlacher T, Rowden A, Stocks K, Consalvey M. Seamounts, deep-sea corals and fisheries: vulnerability of deep-sea corals to fishing on seamounts beyond areas of national jurisdiction. UNEP/WCMC; Cambridge, UK: 2006. [Google Scholar]
- Clark MR, Vinnichenko VI, Gordon JDM, Beck-Bulat GZ, Kukharev NN, Kakora AF. Large-scale distant-water trawl fisheries on seamounts. In: Pitcher TJ, Morato T, Hart PJB, Clark MR, Haggan N, Santos RS, editors. Seamounts: Ecology, Fisheries & Conservation. Blackwell Publishing; Oxford: 2007. pp. 361–399. Fish and Aquatic Resources Series 12. [Google Scholar]
- Clubbe C. Botanical explorations of the Chagos Archipelago. Chagos News. 2010;36:8–10. [Google Scholar]
- Cohen AN, Carlton JT. Accelerating invasion rate in a highly invaded estuary. Science. 1998;279:55–58. doi: 10.1126/science.279.5350.555. [DOI] [PubMed] [Google Scholar]
- Coles SL, DeFelice RC, Eldredge LG, Carlton JT. Historical and recent introductions of non- indigenous marine species into Pearl Harbor, Oahu, Hawaiian Islands. Marine Biology. 1999;135:147–158. [Google Scholar]
- Collette BB, Parin NV. Shallow-water fishes of Walters Shoals, Madagascar Ridge. Bulletin of Marine Science. 1991;48:1–22. [Google Scholar]
- Concepcion G, Kahng SE, Crepeau M, Franklin EC, Coles S, Toonen RJ. Resolving natural ranges and marine invasions in a globally distributed octocoral (genus Carijoa) Marine Ecology Progress Series. 2010;401:113–127. [Google Scholar]
- Couper A. The Times Atlas of the Oceans. The Times; London: 1987. p. 272. [Google Scholar]
- Craig MT. The goldrim surgeonfish (Acanthurus nigricans; Acanthuridae) from Diego Garcia, Chagos Archipelago: first record for the central Indian Ocean. Zootaxa. 2008;1850:65–68. [Google Scholar]
- Cripps GC. Hydrocarbons in the Antarctic marine environment: monitoring and background. International Journal of Environmental Analytical Chemistry. 1994;55:3–13. [Google Scholar]
- Daniel JC. The Book of Indian Reptiles and Amphibians. Bombay Natural History Society, Oxford University Press; Mumbai: 2002. [Google Scholar]
- Davies AJ, Guinotte JM. Global habitat suitability for framework-forming cold-water corals. PLoS One. 2011;6 (4):e18483. doi: 10.1371/journal.pone.0018483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Diaz M, Madin J. Macroecological relationships between coral species’ traits and disease potential. Coral Reefs. 2011;30:73–84. [Google Scholar]
- Dumbraveanu D, Sheppard CRC. Areas substrate at different depths in the Chagos Archipelago. In: Sheppard CRC, Seaward MRD, editors. Ecology of the Chagos Archipelago. Linnean Society/Westbury Publishing; London: 1999. pp. 35–44. [Google Scholar]
- Eble JA, Craig MT, Rocha L, Bowen BW. Not all offspring stay close to home: trans-Pacific dispersal and gene flow in Indo-Pacific reef fishes. Journal of Marine Biology. 2011:12. doi: 10.1155/2011/518516. Article ID 518516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Edis R. Peak of Limuria. Anthony Rowe, Ltd; Chippenham: 2004. [Google Scholar]
- van der Elst R, Everett B, Jiddawi N, Mwatha G, Afonso PS, Boulle D. Fish, fishers and fisheries of the Western Indian Ocean: their diversity and status. A preliminary assessment. Philosophical Transactions of the Royal Society. B. 2005;363:263–284. doi: 10.1098/rsta.2004.1492. [DOI] [PubMed] [Google Scholar]
- Etnoyer P, Canny D, Mate B, Morgan L. Persistent pelagic habitats in the Baja California to Bering Sea (BSB) ecoregion. Oceanography. 2004;17:90–101. [Google Scholar]
- Everaarts JM, Booij K, Fisher CV, Maas YEM, Nieuwenhuize J. Assessment of the environmental health of the Chagos Archipelago. In: Sheppard CRC, Seaward MRD, editors. Ecology of the Chagos Archipelago. Linnean Society/Westbury Publishing; London: 1999. pp. 305–326. [Google Scholar]
- FAO Fisheries and Aquaculture Department. The State of World Fisheries and Aquaculture 2010 (SOFIA) Food and Agriculture Organisation of the United Nations; Rome: 2010. [Google Scholar]
- FitzSimmons N. Marine Turtle Conservation Fund Award 98210-7-G126. 2010. Final report: population genetic studies in support of conservation and management of hawksbill turtles in the Indian Ocean. Unpublished report to Multinational Species Conservation Fund. [Google Scholar]
- Fletcher WJ, Amos M. ACIAR Monograph No. 29. Australian Centre for International Agricultural Research; GPO Box 157 1, Canberra, Australia 2601: 1994. Stock assessment of coconut crabs. [Google Scholar]
- Fowler SW. PCBs in the environment: the Mediterranean marine ecosystem. In: Waid JS, editor. PCBs and the Environment. III. CRS Press; Boca Ratón, FL: 1990. pp. 209–239. [Google Scholar]
- Gaither MR, Toonen RJ, Robertson DR, Plane S, Bowen BW. Genetic evaluation of marine biogeographical barriers: perspectives from two widespread IndoPacific snappers (Lutjanus kasmira and Lutjanus fulvus) Journal of Biogeography. 2010;37:133–147. [Google Scholar]
- Gaither MR, Bowen BW, Bordenave TR, Rocha LA, Newman SJ, Gomez JA, van Herwerden L, Craig MT. Phylogeography of the reef fish Cephalopholis argus (Epinephelidae) indicates Pleistocene isolation across the Indo-Pacific Barrier with contemporary overlap in the Coral Triangle. BMC Evolutionary Biology. 2011;11:189. doi: 10.1186/1471-2148-11-189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Game ET, Grantham HS, Hobday AJ, Pressey RL, Lombard AT, Beckley LE, Gjerde K, Bustamante R, Possingham HP, Richardson AJ. Pelagic protected areas: the missing dimension in ocean conservation. Trends in Ecology and Evolution. 2009;24:360–369. doi: 10.1016/j.tree.2009.01.011. [DOI] [PubMed] [Google Scholar]
- Graham NAJ. Quantifying reef fish biomass and reef shark numbers in Chagos. Chagos News. 2010;36:20–21. [Google Scholar]
- Graham NAJ, Wilson SK, Jennings S, Polunin NVC, Bijoux JP, Robinson J. Dynamic fragility of oceanic coral reef ecosystems. Proceedings of the National Academy of Sciences of the USA. 2006;103:8425–8429. doi: 10.1073/pnas.0600693103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graham NAJ, Wilson SK, Jennings S, Polunin NVC, Robinson J, Bijoux JP, Daw TM. Lag effects in the impacts of mass coral bleaching on coral reef fish, fisheries, and ecosystems. Conservation Biology. 2007;21:1291–1300. doi: 10.1111/j.1523-1739.2007.00754.x. [DOI] [PubMed] [Google Scholar]
- Graham NAJ, McClanahan TR, MacNeil MA, Wilson SK, Polunin NVC, Jennings S, Chabanet P, Clark S, Spalding MD, Letourneur Y, et al. Climate warming, marine protected areas and the ocean-scale integrity of coral reef ecosystems. PLoS ONE. 2008;3(8):e3039. doi: 10.1371/journal.pone.0003039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Graham NAJ, Wilson SK, Pratchett MS, Polunin NVC, Spalding MD. Coral mortality versus structural collapse as drivers of corallivorous butterflyfish decline. Biodiversity and Conservation. 2009a;18:3325–3336. [Google Scholar]
- Graham NAJ, Purkis SJ, Harris A. Diurnal, land-based predation on shore crabs by moray eels in the Chagos Archipelago. Coral Reefs. 2009b;28:397. [Google Scholar]
- Graham NAJ, Spalding MD, Sheppard CRC. Reef shark declines in remote atolls highlight the need for multi-faceted conservation action. Aquatic Conservation: Marine and Freshwater Ecosystems. 2010;20:543–548. [Google Scholar]
- Guitart C, Sheppard A, Frickers T, Price ARG, Readman JW. Negligible risks to corals from antifouling booster biocides and triazine herbicides in coastal waters of the Chagos Archipelago. Marine Pollution Bulletin. 2007;54:226–246. doi: 10.1016/j.marpolbul.2006.10.012. [DOI] [PubMed] [Google Scholar]
- Hamilton M, Topp J. Chagos Plant Species Checklist. 2009 OnLine: www.chagos-trust.org/extrapage.asp?id=5.
- Harris AR. PhD thesis. University of Warwick; 2010. Analysis of recovery patterns of Indian Ocean coral reefs through examination of scleractinian communities and populations. [Google Scholar]
- Harris A, Sheppard CRC. Status and recovery of the coral reefs of the Chagos Archipelago, British Indian Ocean Territory. In: Obura D, Souter D, Linden O, editors. Indian Ocean Coral Reefs. CORDIO, Kalmar University; 2008. pp. 61–69. [Google Scholar]
- Harvell D, Jordán-Dahlgren E, Merkel S, Rosenberg E, Raymundo L, Smith G, Weil E, Willis B. Coral disease, environmental drivers, and the balance between coral and microbial associates. Oceanography. 2007;20:172–195. [Google Scholar]
- Hellberg ME. Footprints on the water: the genetic wake of dispersal among reefs. Coral Reefs. 2007;26:463–473. [Google Scholar]
- Hermes JC, Reason CJC. Annual cycle of the South Indian Ocean (Seychelles-Chagos) thermocline ridge in a regional ocean model. Journal of Geophysical Research. 2008;113 doi: 10.1029/2007JC004363. [DOI] [Google Scholar]
- Hewitt CL, Martin RB. CRIMP Technical Report Number 22. Centre for Research on Introduced Marine Pests - CSIRO Marine Research; Hobart: 2001. Revised protocols for baseline port surveys for introduced marine species: survey design, sampling protocols and specimen handling. [Google Scholar]
- Hewitt CL, Campbell ML, Thresher RE, Martin RB, Boyd S, Cohen BF, Currie DR, Gomon MF, Keough MJ, Lewis JA, et al. Introduced and cryptogenic species in Port Philip Bay, Victoria, Australia. Marine Biology. 2004;144:183–202. [Google Scholar]
- Heywood V, editor. Global Biodiversity Assessment. Cambridge University Press; Cambridge: 1995. [Google Scholar]
- Hobbs JPA, Frisch AJ. Coral disease in the Indian Ocean: taxonomic susceptibility, spatial distribution and the role of host density on the prevalence of white syndrome. Disease of Aquatic Organisms. 2010;89:1–8. doi: 10.3354/dao02181. [DOI] [PubMed] [Google Scholar]
- Hobbs JA, Frisch AJ, Allen GR, Herwerden LV. Marine hybrid hotspot at Indo-Pacific biogeographic border. Biology Letters. 2009;5:258–261. doi: 10.1098/rsbl.2008.0561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoegh-Guldberg O, Mumby PJ, Hooten AJ, Steneck RS, Greenfield P, Gomez E, Harvell CD, Sale PF, Edwards AJ, Caldeira K, et al. Coral reefs under rapid climate change and ocean acidification. Science. 2007;318:1737–1742. doi: 10.1126/science.1152509. [DOI] [PubMed] [Google Scholar]
- Hughes TP, Graham NAJ, Jackson JBC, Mumby PJ, Steneck RS. Rising to the challenge of sustaining coral reef resilience. Trends in Ecology & Evolution. 2010;25:633–642. doi: 10.1016/j.tree.2010.07.011. [DOI] [PubMed] [Google Scholar]
- Hyrenbach K, Forney K, Dayton P. Marine protected areas and ocean basin management. Aquatic Conservation: Marine and Freshwater Ecosystems. 2000;10:437–458. [Google Scholar]
- Hyrenbach K, Fernandez P, Anderson D. Oceanographic habitats of two sympatric North Pacific albatrosses during breeding season. Marine Ecology Progress Series. 2002;233:283–301. [Google Scholar]
- IMO. International Convention for the Control and Management of Ships’ Ballast Water & Sediments. Adopted by consensus in London on Friday 13 February 2004, not in force; May 2011.2004. [Google Scholar]
- IOTC. Report of the Fourteenth Session of the Indian Ocean Tuna Commission; Busan, Korea. 1–5 March 2010; 2010. IOTC-2010-S14-R[E] [Google Scholar]
- IUCN. IUCN Red List of Threatened Species. 2011 Version 2011.1. < http://www.iucnredlist.org>. Downloaded on 16 June 2011.
- Jackson J, Kirby M, Berger H, Bjorndal K, Botsford L, Bourque B, Bradbury R, Cooke R, Erlandson J, Estes J, et al. Historical overfishing and the recent collapse of coastal ecosystems. Science. 2001;293:629–638. doi: 10.1126/science.1059199. [DOI] [PubMed] [Google Scholar]
- Jensen OP, Ortega-Garcia S, Martell SJD, Ahrens RNM, Domeier ML, Walters CJ, Kitchell JF. Local management of a “highly migratory species”: the effects of long-line closures and recreational catch-and-release for Baja California striped marlin fisheries. Progress in Oceanography. 2010;86:176–186. [Google Scholar]
- Jones GP, McCormick MI, Srinivasan M, Eagle JV. Coral decline threatens fish biodiversity in marine reserves. Proceedings of the National Academy of Science. 2004;101:8251–8253. doi: 10.1073/pnas.0401277101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kaczmarsky L, Richardson LL. Do elevated nutrients and organic carbon on Philippine reefs increase the prevalence of coral disease? Coral Reefs. 2011;30:253–257. [Google Scholar]
- Kaplan DM, Chassot E, Gruss A, Fonteneau A. Pelagic MPAs: the devil is in the details. Trends in Ecology and Evolution. 2010;25:62–63. doi: 10.1016/j.tree.2009.09.003. [DOI] [PubMed] [Google Scholar]
- Kimani EN, Okemwa GM, Kazungu JM. Fisheries in the Southwest Indian Ocean: trends and governance challenges. In: Laipson E, Pandya A, editors. The Indian Ocean: Resource and Governance Challenges. The Henry L. Stimson Centre; Washington DC: 2009. [Google Scholar]
- Koldewey H, Curnick D, Harding S, Harrison L, Gollock M. Potential benefits to fisheries and biodiversity of the Chagos Archipelago/British Indian Ocean Territory as a no-take marine reserve. Marine Pollution Bulletin. 2010;60:1906–1916. doi: 10.1016/j.marpolbul.2010.10.002. [DOI] [PubMed] [Google Scholar]
- Lesser MP, Bythell JC, Gates RD, Johnstone RW, Hoegh-Guldberg O. Are infectious diseases really killing corals? Alternative interpretations of the experimental and ecological data. Journal of Experimental Marine Biology and Ecology. 2007;346:36–44. [Google Scholar]
- Lucas JS. Quantitative studies of feeding and nutrition during larval development of the coral reef asteroid Acanthaster planci (L.) Journal of Experimental Marine Biology and Ecology. 1982;65:173–193. [Google Scholar]
- Macdonald AHH, Lamb J, Schleyer MH. personal communication. Population structure of Platygyra daedalea on the south-east African coastline.
- MacNeil MA, Graham NAJ. Enabling coral-reef management in a changing climate through predictive Bayesian meta-analysis of large-scale disturbance. Global Ecology and Biogeography. 2010;19:412–421. [Google Scholar]
- Madden RA, Julian PR. Observations of the 40–50 day tropical oscillation – a review. Monthly Weather Review. 1994;122:814–837. [Google Scholar]
- Marchant R, Mumbi C, Behera S, Yamagata T. The Indian Ocean dipole – the unsung driver of climatic variability in East Africa. African Journal of Ecology. 2007;45:4–16. [Google Scholar]
- Matyot P. The establishment of the crested tree lizard, Calotes versicolor (Daudin, 1802), in Seychelles. Phelsuma. 2004;12:35–47. [Google Scholar]
- McClanahan TR. Coral reef fish communities in management systems with unregulated fishing and small fisheries closures compared with lightly fished reefs – Maldives vs. Kenya. Aquatic Conservation: Marine and Freshwater Ecosystems. 2011;21:186–198. [Google Scholar]
- McClanahan TR, Graham NAJ, Wilson SK, Letourneur Y, Fisher R. Fisheries closure size, age, and history of compliance effects on coral reef fish communities in the western Indian Ocean. Marine Ecology Progress Series. 2009;396:99–109. [Google Scholar]
- McGeoch MA, Butchart SHM, Spear D, Marais E, Kleynhans EJ, Symes A, Chanson J, Hoffmann M. Global indicators of biological invasion: species numbers, biodiversity impact and policy responses. Diversity and Distributions. 2010;16:95–108. [Google Scholar]
- McGowan A, Broderick AC, Godley BJ. Seabird populations of the Chagos Archipelago: an evaluation of Important Bird Area Sites. Oryx. 2008;42:424–429. [Google Scholar]
- Mees C, Pearce J, Clarke J, Wilson O, Carroll A. IOTC Eleventh Session of the Scientific Committee; Mahé, Seychelles. 1–5 December 2008; 2008. IOTC-2008-SC-INF12. [Google Scholar]
- Mees C, Pearce J, Clarke J, Wilson O. UK (Chagos/BIOT) national report. IOTC Twelfth Session of the Scientific Committee; Mahé, Seychelles. 30 November-4 December 2009; 2009. IOTC-2009-SC-INF08. [Google Scholar]
- Millennium Ecosystem Assessment. [Accessed August 2011];2005 http://www.maweb.org/en/index.aspx.
- Miller J, Muller E, Rogers C, Waara R, Atkinson A, Whelan KRT, Patterson M, Witcher B. Coral disease following massive bleaching in 2005 causes 60% decline in coral cover on reefs in the US Virgin Islands. Coral Reefs. 2009;28:925–937. [Google Scholar]
- Molnar JL, Gamboa RL, Revenga C, Spalding MD. Assessing the global threat of invasive species to marine biodiversity. Frontiers in Ecology and the Environment. 2008;6:485–492. [Google Scholar]
- Mora C, Aburto-Oropeza O, Ayala Bocos A, Ayotte PM, Banks S, Bauman AG, Beger M, Bessudo S, Booth DJ, Brokovich E, et al. Global human footprint on the linkage between biodiversity and ecosystem functioning in reef fishes. PLoS Biology. 2011;9(4):e1000606. doi: 10.1371/journal.pbio.1000606. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mortimer JA. Marine Turtle Conservation Fund Award 98210-6-G073. 2007. Final report: status and conservation of nesting turtles in the Chagos islands (BIOT) Unpublished report to Multinational Species Conservation Fund. [Google Scholar]
- Mortimer JA. History of turtle exploitation in Chagos. Chagos News. 2009;34:14–16. [Google Scholar]
- Mortimer JA, Broderick D. Population genetic structure and developmental migrations of sea turtles in the Chagos Archipelago and adjacent regions inferred from mtDNA sequence variation. In: Sheppard CRC, Seaward MRD, editors. Ecology of the Chagos Archipelago. Linnean Society/Westbury Publishing; London: 1999. pp. 184–194. [Google Scholar]
- Mortimer JA, Crain DA. Sex steroid concentrations in immature hawksbill turtles (Eretmochelys imbricata) in the Chagos Archipelago. In: Sheppard CRC, Seaward MRD, editors. Ecology of the Chagos Archipelago. Linnean Society/Westbury Publishing; London: 1999. pp. 173–184. [Google Scholar]
- Mortimer JA, Day M. Sea turtle populations and habitats in the Chagos Archipelago. In: Sheppard CRC, Seaward MRD, editors. Ecology of the Chagos Archipelago. Linnean Society/Westbury Publishing; London: 1999. pp. 159–172. [Google Scholar]
- Mortimer JA, Day M, Broderick D. Sea turtle populations of the Chagos Archipelago, British Indian Ocean Territory. In: NOAA Technical Memorandum NMFSSEFSC-477; Proceedings of the Twentieth Annual Symposium on Sea Turtle Biology and Conservation; 2002. pp. 47–49. [Google Scholar]
- Muthiga N, Costa A, Motta H, Muhando C, Mwaipopo R, Schleyer M. Status of coral reefs in East Africa: Kenya, Tanzania, Mozambique and South Africa. In: Wilkinson C, editor. Status of Coral Reefs of the World: 2008. Global Coral Reef Monitoring Network and Reef and Rainforest Research Center; Townsville, Australia: 2008. pp. 91–104. [Google Scholar]
- Nelson J, Bradner H. The case for establishing ecosystem-scale marine reserves. Marine Pollution Bulletin. 2010;60:635–637. doi: 10.1016/j.marpolbul.2010.04.009. [DOI] [PubMed] [Google Scholar]
- O’Farrell S. MSc thesis. University of Wales; Bangor: 2007. Competition and aggression between scleractinian corals and macroalgae on post-disturbance reefs in the Chagos Archipelago. [Google Scholar]
- Pala C. Protecting the last great tuna stocks. Science. 2009;324:1133. doi: 10.1126/science.324_1133. [DOI] [PubMed] [Google Scholar]
- Palumbi SR. Marine reserves and ocean neighbourhoods: the spatial scale of marine populations and their management. Annual Review of Environmental Resources. 2004;29:31–68. [Google Scholar]
- Pauly D. Anecdotes and the shifting baseline syndrome of fisheries. Trends in Ecology and Evolution. 1995;10:430. doi: 10.1016/s0169-5347(00)89171-5. [DOI] [PubMed] [Google Scholar]
- Pearce J. A Review of the British Indian Ocean Territory Fisheries Conservation and Management Zone Tuna Fishery 1991–1995. Marine Resources Assessment Group; London: 1996. [Google Scholar]
- Pfeiffer M, Dullo W-C, Eisenhauer A. Variability of the Intertropical Convergence Zone recorded in coral isotopic records from the central Indian Ocean (Chagos Archipelago) Quaternary Research. 2004;61:245–255. [Google Scholar]
- Pfeiffer M, Timm O, Dullo W-Chr, Garbe-Schönberg D. Paired Sr/Ca and delta18O records from the Chagos Archipelago: late 20th century warming affects rainfall variability in the tropical Indian Ocean. Geology. 2006;34:1069–1072. [Google Scholar]
- Pfeiffer M, Dullo W-Chr, Zinke J, Garbe-Schönberg D. Three monthly coral Sr/Ca records from the Chagos Archipelago covering the period of 1950 to 1995 A.D.: reproducibility and implications for quantitative reconstructions of sea surface temperature variations. International Journal of Earth Sciences. 2009;98:53–66. [Google Scholar]
- Polunin NVC, Roberts CM. Greater biomass and value of target coral-reef fishes in two small Caribbean marine reserves. Marine Ecology Progress Series. 1993;100:167–176. [Google Scholar]
- Pomeroy RS, Parks JE, Watson IM. How is your MPA doing? A Guidebook of Natural and Social indicators for Evaluating Marine Protected Area Management Effectiveness. IUCN; Gland, Switzerland and Cambridge UK: 2004. [Google Scholar]
- Pratchett MS, Munday PL, Wilson SK, Graham NAJ, Cinner JE, Bellwood DR, Jones GP, Polunin NVC, McClanahan TR. Effects of climate-induced coral bleaching on coral-reef fishes: ecological and economic consequences. Oceanography and Marine Biology: An Annual Review. 2008;46:251–296. [Google Scholar]
- Pratchett MS, Graham NAJ, Sheppard CRC, Mayes B. Are infestations of Cymo melanodactylus killing Acropora cytherea in the Chagos Archipelago? Coral Reefs. 2010;29:941. [Google Scholar]
- Price ARG. Broadscale coastal environmental assessment of the Chagos Archipelago. In: Sheppard CRC, Seaward MRD, editors. Ecology of the Chagos Archipelago. Linnean Society/Westbury Publishing; London: 1999. pp. 285–296. [Google Scholar]
- Price ARG, Harris A. Decadal changes (1996–2006) in coastal ecosystems of the Chagos Archipelago determined from rapid assessment. Aquatic Conservation: Marine and Freshwater Ecosystems. 2009;19:637–644. [Google Scholar]
- Price ARG, Harris A, McGowan A, Venkatachalam AJ, Sheppard CRC. Chagos feels the pinch: assessment of holothurian (sea cucumber) abundance, illegal harvesting and conservation prospects in British Indian Ocean Territory. Aquatic Conservation: Marine and Freshwater Ecosystems. 2010;20:117–126. [Google Scholar]
- Rajasuriya A, Zahir H, Venkataraman K, Islam Z, Tamelander J. Status of Coral Reefs in South Asia: Bangladesh, Chagos, India, Maldives and Sri Lanka. In: Wilkinson C, editor. Status of Coral Reefs of the World: 2004. Vol. 1. Australian Institute of Marine Science; Townsville, Queensland, Australia: 2004. pp. 213–234. [Google Scholar]
- Raymundo LJ, Rosell KB, Reboton C, Kaczmarsky L. Coral diseases on Philippine reefs: genus Porites is a dominant host. Diseases of Aquatic Organisms. 2005;64:181–191. doi: 10.3354/dao064181. [DOI] [PubMed] [Google Scholar]
- Readman JW, Tolosa I, Bartocci J, Cattini C, Price ARG, Jollife A. Contaminant levels and the use of molecular organic markers to characterize the coastal environment of the Chagos Archipelago. In: Sheppard CRC, Seaward MRD, editors. Ecology of the Chagos Archipelago. Linnean Society/Westbury Publishing; London: 1999. pp. 297–304. [Google Scholar]
- Reece JS, Bowen BW, Joshi K, Goz V, Larson L. Phylogeography of two moray eels indicates high dispersal throughout the Indo-Pacific. Journal of Heredity. 2010;101:391–402. doi: 10.1093/jhered/esq036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reid D, Lal K, Mackenzie-Dodds J, Kaligis F, Littlewood D, Williams S. Comparative phylogeography and species boundaries in Echinolittorina snails in the central Indo-West Pacific. Journal of Biogeography. 2006;33:990–1006. [Google Scholar]
- Reinicke GB, van Ofwegen LP. Soft corals (Alcyonacea: Octocorallia) from shallow water in the Chagos Archipelago: species assemblages and their distribution. In: Sheppard CRC, Seaward MRD, editors. Ecology of the Chagos Archipelago. Linnean Society/Westbury Publishing; London: 1999. pp. 67–90. [Google Scholar]
- Richard Y, Trzaska S, Roucou P, Rouault P. Modification of the southern African rainfall variability/ENSO relationship since the late 1960s. Climate Dynamics. 2000;16:883–895. [Google Scholar]
- Riegl B, Piller WE. Possible refugia for reefs in times of environmental stress. International Journal of Earth Sciences. 2003;92:520–531. [Google Scholar]
- Roberts J. Report for MRAG. London: 2007. Preliminary characterization of bycatch by tuna longliners operating in the Chagos/BIOT area. [Google Scholar]
- Roberts RC, Sargant H. Fishery benefits of fully protected marine reserves: why habitat and behaviour are important. Natural Resource Modelling. 2002;15:487–507. [Google Scholar]
- Rogers AD, d’A Laffoley D. Summary report. IPSO; Oxford: 2011. International Earth system expert workshop on ocean stresses and impacts. http://www.stateoftheocean.org/pdfs/1906_IPSO-LONG.pdf. [Google Scholar]
- Rogers AD, Baco A, Griffiths HJ, Hart T, Hall-Spencer JM. Corals on seamounts. In: Pitcher TJ, Morato T, Hart PJB, Clark MR, Haggan N, Santos RS, editors. Seamounts: Ecology, Fisheries and Conservation. Blackwell Publishing Ltd; Oxford: 2007. pp. 141–169. [Google Scholar]
- Rohwer F, Kelley S. Culture-independent analyses of coral-associated microbes. In: Rosenberg E, Loya Y, editors. Coral Health and Disease. Springer; Berlin: 2004. pp. 265–277. [Google Scholar]
- Royal Haskoning. Final Report. Royal Haskoning; 2002. Chagos feasibility study, Phase 2B. [Google Scholar]
- Ruiz GM, Rawlings TK, Dobbs FC, Drake LA, Mullady T, Huq A, Colwell RR. Global spread of microorganisms by ships. Ballast water discharged from vessels harbours a cocktail of potential pathogens. Nature. 2000;408:49–50. doi: 10.1038/35040695. [DOI] [PubMed] [Google Scholar]
- Saji NH, Yamagata T. Structure of SST and surface wind variability during Indian Ocean Dipole Mode events: COADS observations. Journal of Climate. 2003;16:2735–2751. [Google Scholar]
- Saji HH, Goswami BN, Vinayachandran PN, Yamagata T. A dipole mode in the tropical Indian Ocean. Nature. 1999;401:360–363. doi: 10.1038/43854. [DOI] [PubMed] [Google Scholar]
- Sale PF, editor. Dynamics and Diversity in a Complex Ecosystem. Academic Press; San Diego: 2002. Coral Reef Fishes. [Google Scholar]
- Sandin SA, Smith JE, DeMartini EE, Dinsdale EA, Donner SD, Friedlander AM, Konotchick T, Malay M, Maragos JE, Obura D, et al. Baselines and degradation of coral reefs in the northern Line Islands. PLoS ONE. 2008;3(2):e1548. doi: 10.1371/journal.pone.0001548. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schaefer KM, Fuller DW. Vertical movements, behavior, and habitat of bigeye tuna (Thunnus obesus) in the equatorial eastern Pacific Ocean, ascertained from archival tag data. Marine Biology. 2010;157:2625–2642. [Google Scholar]
- Scheltema RS. Initial evidence for the transport of teleplanic larvae of benthic invertebrates across the East Pacific barrier. Biological Bulletin. 1988;174:145–152. [Google Scholar]
- Schleyer MH, Benayahu Y. Pre- and post-1998 ENSO records of shallow-water octocorals (Alcyonacea) in the Chagos Archipelago. Marine Pollution Bulletin. 2010;60:2197–2200. doi: 10.1016/j.marpolbul.2010.08.019. [DOI] [PubMed] [Google Scholar]
- Seaward MRD. Cryptogamic flora of the Chagos Archipelago. In: Sheppard CRC, Seaward MRD, editors. Ecology of the Chagos Archipelago. Linnean Society/Westbury Publishing; London: 1999. pp. 207–224. [Google Scholar]
- Seaward MRD, Aptroot A. The lichen flora of the Chagos Archipelago, including a comparison with other island and coastal tropical floras. Tropical Bryology. 2000;18:185–198. [Google Scholar]
- Seaward MRD, Ellis LT, Pócs T, Wigginton MJ. Bryophyte flora of the Chagos Archipelago. Journal of Bryology. 2006;28:11–19. [Google Scholar]
- Sheppard CRC. Status of three rare animals on Chagos. Environmental Conservation. 1979;6:310. [Google Scholar]
- Sheppard CRC. Coral cover, zonation and diversity on reef slopes of Chagos atolls, and population structures of the major species. Marine Ecology Progress Series. 1980a;2:193–205. [Google Scholar]
- Sheppard CRC. The coral fauna of Diego Garcia lagoon, following harbour construction. Marine Pollution Bulletin. 1980b;11:227–230. [Google Scholar]
- Sheppard CRC. Changes in coral cover on reefs of Chagos over eighteen years. In: Sheppard CRC, Seaward MRD, editors. Ecology of the Chagos Archipelago. Linnean Society/Westbury Publishing; London: 1999a. pp. 91–100. [Google Scholar]
- Sheppard CRC. Coral decline and weather patterns over 20 years in the Chagos Archipelago, central Indian Ocean. Ambio. 1999b;28:472–478. [Google Scholar]
- Sheppard CRC. Corals of Chagos, and the role of Chagos reefs in the Indian Ocean. In: Sheppard CRC, Seaward MRD, editors. Ecology of the Chagos Archipelago. Linnean Society/Westbury Publishing; London: 1999c. pp. 53–66. [Google Scholar]
- Sheppard CRC. Changes in some weather patterns in Chagos over twenty five years. In: Sheppard CRC, Seaward MRD, editors. Ecology of the Chagos Archipelago. Linnean SocietyWestbury Publishing; London: 1999d. pp. 45–52. [Google Scholar]
- Sheppard CRC. Island elevations, reef condition and sea level rise in atolls of Chagos, British Indian Ocean Territory. In: Linden O, Souter D, Wilhelmsson D, Obura D, editors. Cordio Report 2002. Kalmar University; Kalmar, Sweden: 2002. pp. 202–211. [Google Scholar]
- Sheppard CRC. Predicted recurrences of mass coral mortality in the Indian Ocean. Nature. 2003;425:294–297. doi: 10.1038/nature01987. [DOI] [PubMed] [Google Scholar]
- Sheppard CRC. Longer term impacts of climate change. In: Cote I, Reynolds J, editors. Coral Reef Conservation. Cambridge University Press; Cambridge: 2006. pp. 264–290. [Google Scholar]
- Sheppard CRC. Large temperature plunges recorded by data loggers at different depths on an Indian Ocean atoll: comparison with satellite data and relevance to coral refuges. Coral Reefs. 2009;28:399–403. [Google Scholar]
- Sheppard CRC, Spalding MD, Bradshaw C, Wilson S. Erosion vs recovery of coral reefs after 1998 El Nino: Chagos reefs, Indian Ocean. Ambio. 2002;31:40–48. [PubMed] [Google Scholar]
- Sheppard CRC, Dixon DJ, Gourlay M, Sheppard ALS, Payet R. Coral mortality increases wave energy reaching shores protected by reef flats: examples from the Seychelles. Estuarine, Coastal and Shelf Science. 2005;64:223–234. [Google Scholar]
- Sheppard CRC, Harris A, Sheppard ALS. Archipelago-wide coral recovery patterns since 1998 in the Chagos Archipelago, central Indian Ocean. Marine Ecology Progress Series. 2008;362:109–117. [Google Scholar]
- Shotton R. FAO Fisheries Circular No. 1020. FAO; Rome, Italy: 2006. Managment of demersal fisheries resources of the Southern Indian Ocean. [Google Scholar]
- Sibert J, Hampton J. Mobility of tropical tunas and the implications for fisheries management. Marine Policy. 2003;27:87–95. [Google Scholar]
- Singh OP, Khan TMA, Aktar F, Sarker MA. Recent sea level and sea surface temperature changes along the Maldives coast. Marine Geodesy. 2001;24:209–218. [Google Scholar]
- Slatkin M. Gene flow and genetic drift in a species subject to frequent local extinctions. Theoretical Population Biology. 1977;12:253–262. doi: 10.1016/0040-5809(77)90045-4. [DOI] [PubMed] [Google Scholar]
- Slatkin M. Testing neutrality in subdivided populations. Genetics. 1982;100:533–545. doi: 10.1093/genetics/100.3.533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spalding MD, Ravilious C, Green EP. World Atlas of Coral Reefs. University of California Press; Berkely, CA: 2001. [Google Scholar]
- Spalding M, Wood L, Fitzgerald C, Gjerde K. The 10% target: where do we stand? In: Toropova C, Meliane I, Laffoley D, Matthews E, Spalding M, editors. Global Ocean Protection: Present Status and Future Possibilities. Brest, France: Agence des Aires Marines Protégées; Gland, Switzerland, Washington, DC and New York, USA: IUCN WCPA; Cambridge, UK: UNEP-WCMC; Arlington, USA: TNC; Tokyo, Japan: UNU; New York, USA: WCS; 2010. pp. 25–40. [Google Scholar]
- Stoddart DR. Settlement and development of Diego Garcia. Atoll Research Bulletin. 1971;149:208–237. [Google Scholar]
- Stoddart DR, Taylor DL, editors. Geology and ecology of Diego Garcia Atoll, Chagos Archipelago. Atoll Research Bulletin. 1971;149:237. [Google Scholar]
- SWIOFC. South West Indian Ocean Fisheries Commission. Report of the Third Session of the Scientific Committee; Mahe, Seychelles. 17–20 December 2007.2009. [Google Scholar]
- Symens P. Breeding seabirds of the Chagos Archipelago. In: Sheppard CRC, Seaward MRD, editors. Ecology of the Chagos Archipelago. Linnean Society/Westbury Publishing; London: 1999. pp. 35–44. [Google Scholar]
- Tamelander J, Rajasuriya A. Status of coral reefs in South Asia: Bangladesh, Chagos, India, Maldives and Sri Lanka. In: Wilkinson C, editor. Status of Coral Reefs of the World: 2008. Global Coral Reef Monitoring Network and Reef and Rainforest Research Center; Townsville, Australia: 2008. pp. 119–130. [Google Scholar]
- Tamelander J, Campbell M, Lundin CG. IUCN Global Marine Programme 2009. IUCN; Gland, Switzerland: 2009. Detecting bioinvasions on small islands in the Indian Ocean - project completion report. [Google Scholar]
- Thresher RE, Kuris AM. Options for managing invasive marine species. Biological Invasions. 2004;6:295–300. [Google Scholar]
- Timm O, Pfeiffer M, Dullo W-Chr. Nonstationary ENSO-precipitation teleconnection over the equatorial Indian ocean documented in a coral from the Chagos Archipelago. Geophysical Research Letters. 2005;32:L02701. doi: 10.1029/2004GL021738. [DOI] [Google Scholar]
- Tolosa I, Bayona JM, Albaiges J. Spatial and temporal distribution, fluxes, and budgets of organochlorinated compounds in northwest Mediterranean sediments. Environmental Science and Technology. 1995;29:2519–2527. doi: 10.1021/es00010a010. [DOI] [PubMed] [Google Scholar]
- Topp JMW, Sheppard CRC. Higher plants of the Chagos Archipelago. In: Sheppard CRC, Seaward MRD, editors. Ecology of the Chagos Archipelago. Linnean Society/Westbury Publishing; London: 1999. pp. 225–240. [Google Scholar]
- Toropova C, Meliane I, Laffoley D, Matthews E, Spalding M, editors. Global Ocean Protection: Present Status and Future Possibilities. Agence des aires marines protégées; Brest, France: IUCNWCPA; Gland, Switzerland, Washington, DC and New York: UNEP-WCMC Cambridge, TNC; Arlington, USA: UNU; Tokyo, Japan: WCS; New York: 2010. [Google Scholar]
- Tu M. Assessing and Managing Invasive Species within Protected Areas. In: Ervin J, editor. Protected Area Quick Guide Series. The Nature Conservancy; Arlington, VA: 2009. [Google Scholar]
- United Nations. Report of the World Summit on Sustainable Development. Johannesburg, South Africa; 26 August–4 September 2002; UN, NY, USA. 2002. A/CONF.199/20. [Google Scholar]
- Vargas SM, Jensen MP, Mobaraki A, Santos FR, Broderick D, Mortimer JA, Limpus C, Whiting S, FitzSimmons N. Phylogeography of the Hawksbill turtles (Eretmochelys imbricata) from the Indo Pacific Region. 29 April 2010; Symposium Proceedings. In press. [Google Scholar]
- Vermeersen LA, Schotman HHA. Constraints on glacial isostatic adjustment from GOCE and sea level data. Pure and Applied Geophysics. 2009;166:1261–1281. [Google Scholar]
- Vialard J, Foltz GR, McPhaden MJ, Duvel JP, de Boyer Montegut C. Strong Indian Ocean sea surface temperature signals associated with the Madden-Julian Oscillation in late 2007 and early 2008. Geophysical Research Letters. 2008;35:L19608. [Google Scholar]
- Vialard J, Duvel JP, McPhaden MJ, Bouruet-Aubertot P, Ward B, Key E, Bourras D, Weller R, Minnett P, Weill A, et al. Cirene: air-sea interactions in the Seychelles-Chagos thermocline ridge region. Bulletin of the American Meteorological Society. 2009;90 doi: 10.1175/2008BAMS2499.1. [DOI] [Google Scholar]
- Vogler C, Benzie J, Lessios H, Barber P, Worheide G. A threat to coral reefs multiplied? Four species of crown-of-thorns starfish. Biology Letters. 2008;4:696–699. doi: 10.1098/rsbl.2008.0454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vogler C, Benzie J, Barber P, Sheppard C, Tenggardjaja K, Gérard K, Wörheide G. Cryptic speciation: the crown-of-thorns starfish in the Indian Ocean. In preparation. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walters C. Impacts of dispersal, ecological interactions, and fishing effort dynamics on efficacy of marine protected areas: how large should protected areas be? Bulletin of Marine Science. 2000;66:745–757. [Google Scholar]
- Watling R, Seaward MRD. Some fungi of Indian Ocean islands. Botanical Journal of Scotland. 2004;56:65–84. [Google Scholar]
- Webster PJ, Moore A, Loschnigg J, Leban M. Coupled ocean-atmosphere dynamics in the Indian Ocean during 1997–1998. Nature. 1999;401:356–360. doi: 10.1038/43848. [DOI] [PubMed] [Google Scholar]
- Weil E, Urreiztieta I, Garzon-Ferreira J. Geographic variability in the incidence of coral and octocoral diseases in the wider Caribbean. Proceedings of the 9th International Coral Reef Symposium; Bali, Indonesia. 2002. pp. 1231–1237. [Google Scholar]
- Weil E, Smith GW, Gil-Agudelo DL. Status and progress in coral reef disease research. Diseases of Aquatic Organisms. 2006;60:1–7. doi: 10.3354/dao069001. [DOI] [PubMed] [Google Scholar]
- Wilcove DS, Rothstein D, Dubow J, Phillips A, Losos E. Quantifying threats to imperiled species in the United States. Bioscience. 1998;48:607–615. [Google Scholar]
- Wilkinson C. Executive summary. In: Wilkinson C, editor. Status of Coral Reefs of the World: 2008. Global Coral Reef Monitoring Network and Reef and Rainforest Research Center; Townsville, Australia: 2008a. pp. 5–19. [Google Scholar]
- Wilkinson C, editor. Status of Coral Reefs of the World: 2008. Global Coral Reef Monitoring Network and Reef and Rainforest Research Center; Townsville, Australia: 2008b. [Google Scholar]
- Williams GJ, Davy SK, Aeby GS. Coral disease at Palmyra Atoll, a remote reef system in the Central Pacific. Coral Reefs. 2007;27:207. [Google Scholar]
- Williams ID, Richards BL, Sandin SA, Baum JK, Schroeder RE, Nadon MO, Zgliczynski B, Craig P, McIlwain JL, Brainard RE. Differences in reef fish assemblages between populated and remote reefs spanning multiple archipelagos across the central and western Pacific. Journal of Marine Biology. 2011:Article ID 826234. doi: 10.1155/2011/826234. [DOI] [Google Scholar]
- Willis B, Page C, Dinsdale E. Coral diseases on the Great Barrier Reef. In: Rosenberg E, Loya Y, editors. Coral Health and Disease. Springer; Berlin: 2004. pp. 69–104. [Google Scholar]
- Wilson JR, Harrison PL. Settlement-competency periods of larvae of three species of scleractinian corals. Marine Biology. 1998;131:339–345. [Google Scholar]
- Wilson SK, Graham NAJ, Pratchett MS, Jones GP, Polunin NVC. Multiple disturbances and the global degradation of coral reefs: are reef fishes at risk or resilient? Global Change Biology. 2006;12:2220–2234. [Google Scholar]
- Wilson SK, Fisher R, Pratchett MS, Graham NAJ, Dulvy NK, Turner RA, Cakackak A, Polunin NVC, Rushton SP. Exploitation and habitat degradation as agents of change within coral reef fish communities. Global Change Biology. 2008;14:2796–2809. [Google Scholar]
- Winterbottom R, Anderson RC. A revised checklist of the epipelagic and shore fishes of the Chagos Archipelago, Central Indian Ocean. JLB Smith Institute of Ichthyology. Ichthyological Bulletin. 1997;66:1–28. [Google Scholar]
- Wolschke H, Sturm R, Ebinghaus R. Report to Save Our Seas Foundation. 2011. Analysis of flame retardants and polyfluorinated compounds in sediment samples from the Chagos Archipelago. [Google Scholar]
- Wood LJ, Fish L, Laughren J, Pauly D. Assessing progress towards global marine protection targets: shortfalls in information and action. Oryx. 2008;42:340–351. [Google Scholar]
- Worm B, Lotze K, Myers R. Predator diversity hotspots in the blue ocean. Proceedings of the National Academy of Science. 2003;100:9884–9888. doi: 10.1073/pnas.1333941100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Worm B, Sandow M, Oschlies A, Lotze HK, Myers RA. Global patterns of predator diversity in the open oceans. Science. 2005;309:1365–1369. doi: 10.1126/science.1113399. [DOI] [PubMed] [Google Scholar]
- Yang SY, Keshavmurthy S, Obura D, Sheppard CRC, Visram S, Chen CA. Diversity and distribution of Symbiodinium associated with seven common coral species in the Chagos Archipelago. PLoS One. 2012 doi: 10.1371/journal.pone.0035836. (Accepted) [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yesson C, Clark MR, Taylor M, Rogers AD. The global distribution of seamounts based on 30-second bathymetry data. Deep Sea Research Part I: Oceanographic Research Papers. 2011;58:442–453. [Google Scholar]
- Yesson C, Taylor M, Tittensor D, Davies A, Guinotte J, Baco A, Black J, Hall-Spencer M, Rogers AD. Global habitat suitability of cold water octocorals. Journal of Biogeography In press. [Google Scholar]
- Young IR, Zieger S, Babanin AV. Global trends in wind speed and wave height. Science. 2011;332:451–455. doi: 10.1126/science.1197219. [DOI] [PubMed] [Google Scholar]
- Zubair L, Ropelewski CF. The strengthening relationship between ENSO and northeast monsoon rainfall over Sri Lanka and southern India. Journal of Climate. 2006;19:1567–1575. [Google Scholar]
- Zvuloni A, Artzy-Randrup Y, Stone L, Kramarsky-Winter E, Barkan R, Loya Y. Spatio-temporal transmission patterns of black-band disease in a coral community. PlosONE. 2009;4:e4993. doi: 10.1371/journal.pone.0004993. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.