

Land-use change remains a major driver of biodiversity loss, and projecting extinction rates for different scenarios of habitat conversion is a key concern in conservation research (Pereira et al. 2010; Wright 2010; de Baan et al. 2013). Species–area relationships (SARs) have been one of the main models used to develop such projections, but they have been criticized recently for overestimating extinctions (He & Hubbell 2011). One problem is that classic projections are based on the assumption that all natural areas converted to human-dominated areas, such as agriculture and forestry, become completely hostile to biodiversity (Pereira et al. 2012). However, there is a growing recognition that many species are not constrained to fragments of their native habitat and that the matrix can play an important role in the conservation of biodiversity (Prugh et al. 2008; Karp et al. 2012). Recently a comparison of 2 models that incorporate the wider landscape context, the countryside SAR (Pereira & Daily 2006) and the matrix-calibrated SAR was conducted by Koh and Ghazoul (2010). Here we show that the results of that comparison are incorrect and that in contrast with their results, the countryside SAR outperforms both the matrix-calibrated SAR and classic SAR projections in projecting tropical bird extinctions.
The countryside SAR classifies species into functional groups with particular affinities for different habitats in the landscape. The richness of each functional group Si is given by
| (1) |
where hij is the affinity of functional group i to habitat j, Aj is the area of habitat j in the landscape, m is the number of habitat types, and ci and zi are the usual parameters of the classic SAR. Affinity can be interpreted as the proportion of area of habitat j that is usable by functional group i, so that 0 ≤ hij ≤ 1.
Consider a completely native landscape where habitat conversion takes place. Assuming there is a single functional group (i.e., dropping the subscript i in Eq. 1), the proportion of species remaining after habitat conversion is
| (2) |
where 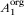 is the original area of the native habitat,
is the original area of the native habitat, 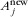 is the area of habitat j after conversion, Snew is the new number of species in the landscape, and Sorg is the original number of species. The original area of native habitat equals the sum of the new areas of all habitats,
is the area of habitat j after conversion, Snew is the new number of species in the landscape, and Sorg is the original number of species. The original area of native habitat equals the sum of the new areas of all habitats, 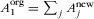 . Furthermore, we assume that species have maximum affinity for the native habitat, h1 = 1.
. Furthermore, we assume that species have maximum affinity for the native habitat, h1 = 1.
Koh and Ghazoul (2010) proposed instead the matrix-calibrated SAR, which gives the proportion of species remaining as
| (3) |
where pj is the proportional area of habitat j relative to the total converted area (area of the matrix), 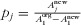 , and σj is the sensitivity of the taxon to the transformed habitat (σ1 = 0).
, and σj is the sensitivity of the taxon to the transformed habitat (σ1 = 0).
To compare the performance of different species–area models in projecting species extinctions, Koh and Ghazoul examined birds in 20 biodiversity hotspots in the world. For each hotspot they estimated the proportion of native habitat remaining and the proportion converted to disturbed forest, agricultural land, and urban area. For each hotspot, they estimated the number of species extinct or threatened with extinction as all endemic bird species in each hotspot classified as extinct, critically endangered, endangered, or vulnerable by the IUCN. Threatened species are included because they are expected to become extinct when species richness reaches an equilibrium with the amount of remaining habitat. Next they estimated sensitivities, σj, and affinities, hj, through the use of a database of studies of how many species disappear locally when natural habitat is converted to each type of human-dominated landscape.
For the countryside SAR, the affinity for habitat k can be derived from such a database with Eq. 2,
| (4) |
if one assumes full habitat conversion (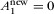 and
and 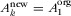 ). For the matrix-calibrated SAR, it is not possible to derive such an expression for full habitat conversion because Eq. 3 always tends to zero when
). For the matrix-calibrated SAR, it is not possible to derive such an expression for full habitat conversion because Eq. 3 always tends to zero when  . Instead Koh and Ghazoul assumed:
. Instead Koh and Ghazoul assumed:
| (5) |
This shows that affinities and sensitivities are related because  . Unfortunately, Koh and Ghazoul calculated the affinities simply as
. Unfortunately, Koh and Ghazoul calculated the affinities simply as 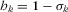 and ignored the exponent z. Using this incorrect calculation of affinities they found that the best projections of endemic bird extinctions are with the matrix model, followed by the classic SAR, and that the countryside SAR has the worst performance. We recalculated the projections of extinction rates with the data from Koh and Ghazoul, the z value they used (0.35), and the correct estimate of habitat affinities. We found that the countryside SAR outperformed both the matrix-calibrated SAR and the classic SAR in this data set (Table1, Fig.1).
and ignored the exponent z. Using this incorrect calculation of affinities they found that the best projections of endemic bird extinctions are with the matrix model, followed by the classic SAR, and that the countryside SAR has the worst performance. We recalculated the projections of extinction rates with the data from Koh and Ghazoul, the z value they used (0.35), and the correct estimate of habitat affinities. We found that the countryside SAR outperformed both the matrix-calibrated SAR and the classic SAR in this data set (Table1, Fig.1).
Table 1.
Goodness of fit of the classic species–area relationship (SAR), countryside SAR, and matrix-calibrated SAR projections of bird extinctions in 20 biodiversity hotspots (z = 0.35).a
| Model | ∑ε2 | AIC | w (%) | Evidence ratio |
|---|---|---|---|---|
| Countryside SAR | 3,417.2 | 46.7 | 77.4 | 1.00 |
| Matrix-calibrated SAR | 4,535.1 | 49.1 | 22.6 | 3.42 |
| Classic SAR | 34,320.6 | 66.7 | 0.0 | 22,446 |
Modified from Koh and Ghazoul (2010). The ∑ε2 is the sum of the squares of the differences between projected extinctions and observed number of extinct and threatened species; AIC is the Akaike's information criterion calculated as  , where n = 20 biodiversity hotspots and K (number of parameters) is 1; w is the Akaike weight
, where n = 20 biodiversity hotspots and K (number of parameters) is 1; w is the Akaike weight where ΔAIC is the difference between the AIC of that model and the best model; and evidence ratio is the ratio between the Akaike weight of the best model and that model.
where ΔAIC is the difference between the AIC of that model and the best model; and evidence ratio is the ratio between the Akaike weight of the best model and that model.
Figure 1.
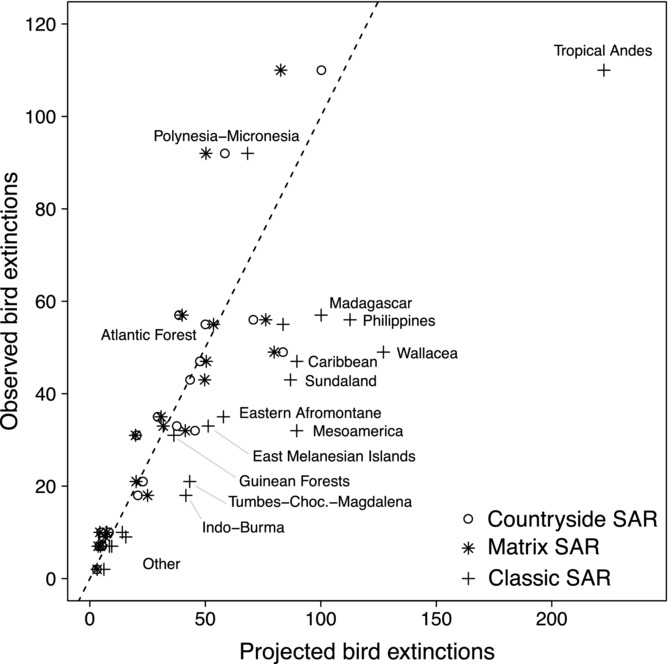
Comparison of observed and projected number of extinct and threatened endemic bird species in 20 biodiversity hotspots (dashed line, perfect fit between projections and observations). Modified from Koh and Ghazoul (2010).
There might be other data sets where the matrix-calibrated SAR outperforms the countryside SAR; more research is needed to compare the different SAR models. The countryside SAR is particularly suitable to describe diversity patterns in multi-habitat landscapes even when the original cover or species composition is not known. The results of 2 recent studies show that the performance of the countryside SAR is better than the classic SAR in describing bird (Guilherme & Pereira 2013) and plant (Proenca & Pereira 2013) diversity in such landscapes.
Literature Cited
- de Baan L, Mutel CL, Curran M, Hellweg S. Koellner T. Land use in life cycle assessment: global characterization factors based on regional and global potential species extinction. Environmental Science & Technology. 2013;47:9281–9290. doi: 10.1021/es400592q. [DOI] [PubMed] [Google Scholar]
- Guilherme JL. Pereira HM. Adaptation of bird communities to farmland abandonment in a mountain landscape. PLoS ONE. 2013;8:e73619. doi: 10.1371/journal.pone.0073619. [DOI] [PMC free article] [PubMed] [Google Scholar]
- He F. Hubbell SP. Species-area relationships always overestimate extinction rates from habitat loss. Nature. 2011;473:368–371. doi: 10.1038/nature09985. [DOI] [PubMed] [Google Scholar]
- Karp DS, Rominger AJ, Zook J, Ranganathan J, Ehrlich PR. Daily GC. Intensive agriculture erodes β-diversity at large scales. Ecology Letters. 2012;15:963–970. doi: 10.1111/j.1461-0248.2012.01815.x. [DOI] [PubMed] [Google Scholar]
- Koh LP. Ghazoul J. A matrix-calibrated species-area model for predicting biodiversity losses due to land-use change. Conservation Biology. 2010;24:994–1001. doi: 10.1111/j.1523-1739.2010.01464.x. [DOI] [PubMed] [Google Scholar]
- Pereira HM, et al. Scenarios for global biodiversity in the 21st century. Science. 2010;330:1496–1502. doi: 10.1126/science.1196624. [DOI] [PubMed] [Google Scholar]
- Pereira HM, Borda-de-Agua L. Martins IS. Geometry and scales in species-area relationships. Nature. 2012;482:E3–E4. doi: 10.1038/nature10857. [DOI] [PubMed] [Google Scholar]
- Pereira HM. Daily GC. Modeling biodiversity dynamics in countryside landscapes. Ecology. 2006;87:1877–1885. doi: 10.1890/0012-9658(2006)87[1877:mbdicl]2.0.co;2. [DOI] [PubMed] [Google Scholar]
- Proenca V. Pereira HM. Species-area models to assess biodiversity change in multi-habitat landscapes: the importance of species habitat affinity. Basic and Applied Ecology. 2013;14:102–114. [Google Scholar]
- Prugh L, Hodges K, Sinclair A. Brashares J. Effect of habitat area and isolation on fragmented animal populations. Proceedings of the National Academy of Sciences of the United States of. 2008;105:20770–20775. doi: 10.1073/pnas.0806080105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wright S. The future of tropical forests. Year in Ecology and Conservation Biology 2010. 2010;1195:1–27. doi: 10.1111/j.1749-6632.2010.05455.x. [DOI] [PubMed] [Google Scholar]