

Abstract
Some eyespots are thought to deflect attack away from the vulnerable body, yet there is limited empirical evidence for this function and its adaptive advantage. Here, we demonstrate the conspicuous ventral hindwing eyespots found on Bicyclus anynana butterflies protect against invertebrate predators, specifically praying mantids. Wet season (WS) butterflies with larger, brighter eyespots were easier for mantids to detect, but more difficult to capture compared to dry season (DS) butterflies with small, dull eyespots. Mantids attacked the wing eyespots of WS butterflies more frequently resulting in greater butterfly survival and reproductive success. With a reciprocal eyespot transplant, we demonstrated the fitness benefits of eyespots were independent of butterfly behaviour. Regardless of whether the butterfly was WS or DS, large marginal eyespots pasted on the hindwings increased butterfly survival and successful oviposition during predation encounters. In previous studies, DS B. anynana experienced delayed detection by vertebrate predators, but both forms suffered low survival once detected. Our results suggest predator abundance, identity and phenology may all be important selective forces for B. anynana. Thus, reciprocal selection between invertebrate and vertebrate predators across seasons may contribute to the evolution of the B. anynana polyphenism.
Keywords: adaptive coloration, visual signalling, wing patterns
1. Introduction
Many organisms have evolved protective coloration to diminish and dissuade predator attack. Eyespot patterns, concentric rings of contrasting colours, are one form of protective coloration against predators. Broadly speaking, large singular eyespots function by startling or intimidating predators while smaller, numerous eyespots at the body margin divert predator attack to less vulnerable body parts (reviewed in [1,2]). Several studies have supported the intimidation hypothesis for eyespots (e.g. [3–6]), yet there is little empirical evidence for the deflection hypothesis (e.g. [7–11]).
Documenting the deflective function of eyespots, specifically ones on the margins of the animal, has proved difficult. Numerous studies suggest eyespots do not re-direct predator attack to the body margin or increase prey survival (e.g. [3–8]). There are a few experiments indirectly suggesting eyespots may be deflective [9,11]. For example, Olofsson et al. [9] demonstrated, under very specific lighting conditions of high ultraviolet (UV) and low visible light intensity, birds attack marginal wing eyespots on dead butterflies. The results of these few experiments beg the questions: can eyespots deflect predator attack of live prey? and does this deflection impact prey survival and reproduction?
Here, we investigated the deflection hypothesis and its impacts on prey survival and reproduction by assessing the behavioural response of an invertebrate predator to a butterfly species which exhibits a seasonal polyphenism in eyespot size (figure 1a). We presented live butterflies (Bicyclus anynana) to hand-reared Chinese mantids (Tenodera sinensis) in a series of laboratory experiments. In the first experiment, we observed the response of mantids to butterflies with small eyespots and large eyespots, noting the position of their attacks on the butterfly wing and the percentage of prey escapes. Second, we conducted a microcosm experiment with a single predator and multiple butterflies of one seasonal form evaluating both the length of butterfly survival and number of eggs laid. Finally, we controlled for the effect of butterfly behaviour across seasonal forms by transplanting eyespots from one form to the other and repeating the microcosm experiment.
Figure 1.

Mantid attack behaviours on dry season (DS) and wet season (WS) forms of Bicyclus anynana, and survival outcome for arena experiments. (a) The ventral surface of the two seasonal forms of B. anynana. Note the differences in the ventral hindwing eyespot size. (b) Latency for the invertebrate predator, Tenodera sinensis, to orient on each form of B. anynana. Means±95% CI presented. The DS form took longer for mantids to detect. (c) Percentage of butterfly escape once attacked by a praying mantid. The DS form was much less likely to escape once an attack was initiated. (d) Percentage of mantid first strike on various body parts of B. anynana. The WS form was attacked more frequently on the hindwings than the DS form. (e) Percentage of damage observed per hindwing eyespot in the WS form only. Eyespots Cu1, Cu2 and Pc were the most damaged. (Online version in colour.)
The seasonal eyespot polyphenism of B. anynana is determined by early developmental temperature conditions [12]. If the immature experiences ambient temperatures above 23°C, then the adult has large, bright and conspicuous marginal eyespots on its ventral hindwing. If the immature experiences ambient temperatures below 19°C, then the adult has small, dull and cryptic marginal eyespots on its ventral hindwing. This polyphenism is referred to as wet season (WS) and dry season (DS) forms, respectively (figure 1a). Previous research demonstrated the smaller, duller DS eyespots are more advantageous against vertebrate predators relative to WS eyespots, by delaying prey detection and increasing latency to attack [8]. However, once the prey is discovered, both DS and WS have a low probability of escape and survival [7,8,10], calling into question the functional benefits and adaptive importance of the WS form.
Tenodera sinensis, the Chinese mantid, has a widespread distribution, is neurologically and behaviourally similar to other mantid species, and has been used as a model for investigating insect predator behaviour (e.g. [13,14]). Bicyclus anynana inhabits geographical areas with some of the highest global mantid diversity and abundance [15]. Mantids and other invertebrate predators have been observed as active predators in the field with B. anynana, especially during the wet season [16] making mantids good candidates as selective agents for butterflies with large eyespots. Here we investigated whether the large marginal eyespots of B. anynana WS form deflect mantid attack to the wing margin thereby increasing butterfly survival and reproduction when compared with the reduced margin eyespots of the WS form.
2. Material and methods
(a). Experimental animals
Eggs of B. anynana were collected from females from a laboratory colony. Larvae were raised on young maize plants (Zea mays) in a climate room at either 27°C (to produce the WS form) or 17°C (to produce the DS form) with a 12 L : 12 D cycle and 80% relative humidity. Males and females were separated on the day of pupal eclosion so they were virgins at the onset of the experiments. All adults were kept at 22°C and fed a maintenance diet of banana.
Egg cases of T. sinensis were purchased from Carolina Biological Supply Company and reared to adults in individual cages on a successive diet of fruit flies, houseflies and crickets. They were raised to the ultimate instar at 27°C (wet season temperatures) with a 12 L : 12 D cycle and 80% relative humidity. Mantids were not exposed to either butterflies or eyespots before any trial.
(b). Mantid response to individual wet season and dry season prey in an arena
The arena was designed to ritualize the encounter between prey and predator across trials; it consisted of three components: a rectangular ramp, a square floor and a cylindrical wall (electronic supplementary material, figure S1a). The ramp began outside the cylindrical wall, continued through a port in the wall, and continued upwards inside the wall at an 18° angle measuring a total of 35 cm long and 7 cm wide. Both ramp and floor were constructed of wood and covered in poster board paper, while the cylindrical wall was constructed of paper. Ramp, floor and wall were painted a uniform green (paint reflectance spectra in the electronic supplementary material, figure S1b). The arena was illuminated by two full-spectrum halogen lamps (Solux-Eiko 18003, 50W, 4700°K, CRI 91, 36° field of illumination; see irradiance spectra in the electronic supplementary material, figure S1c). Each lamp was positioned 23 cm above the highest point of the ramp and 20 cm from the other lamp to fully illuminate the arena.
A single butterfly was starved for 24 h then placed at the top of the ramp inside the arena wall. The posterior/anterior axis of the butterfly was perpendicular to the ramp. Butterflies were trained to remain in this position by a small food reward, a small piece of banana placed on a piece of removable parafilm. At the start of each trial, the butterfly acclimated for 5 min before the predator was introduced. After 5 min, a single mantid was placed at the ramp base outside the arena wall, out of view of the butterfly. Mantids are negatively geotatic, walking upwards from a lower position and were previously trained via a cricket food reward to walk directly up the ramp at a steady pace without stopping. Once a mantid demonstrated the desired walking behaviour consistently three times in a row, experimental trials with butterflies began. All mantids learned to walk up the ramp, without straying or stopping, over the course of three to five experiences until they completed the three trial series.
We used a repeated-measures design where each mantid (n = 20) encountered both WS and DS butterflies in random order. Each mantid experienced each butterfly form four times, a new butterfly was used for each trial. We measured latency to orient on prey, latency to attack prey and prey survival after attack. Almost all trials resulted in a mantid attack sequence, but in two trials the butterfly flew away before an attack was initiated (n = 2 out of 160 trials, 1% of trials resulted in no attack). These two butterflies were then removed from the experiment and the mantids were given another trial with a different butterfly to have even numbers of trials across mantids. Data were either square root (days, eggs) or arcsine transformed (per cent wing damage), evaluated for homoscedasticity and sphericity then analysed using one-way repeated measures ANOVA (R v. 2.13.0) with mantid as a random effect. We present p-values from two-tailed tests with α = 0.05. Means and 95% CI are reported.
We also noted the attack location on the butterfly. Because mantids remove the wings of butterflies before consuming them, we were able to record eyespot damage for both killed and escaped butterflies. WS wing damage data were evaluated with t-tests. We present p-values from two-tailed tests with α = 0.05.
(c). Butterfly polyphenism and predation microcosm experiment
We used a 2 × 2 factorial design (mantid presence × butterfly form) to evaluate the effect of seasonal form on butterfly survival and reproduction in the presence of a mantid predator. There were four treatments: without mantid × DS butterflies, with mantid × DS butterflies, without mantid × WS butterflies and with mantid × WS butterflies. There were 12 trials per treatment (n = 48), each conducted under full-spectrum lights at 22°C. Each trial was performed in a microcosm cylindrical cage (50 cm diameter × 100 cm height) with a full-spectrum light source, adult food source and oviposition substrate (Zea mays). We used 10 virgin butterflies (five males and five females) and 0–1 mantid in each trial. All experimental animals were naive, i.e. did not experience each other before the trial. Trials began at 12.00 and lasted until all the butterflies were consumed. We counted the number of half days until all the B. anynana were consumed by the mantid. We also recorded the amount of wing damage on the remaining wings left by the mantids and the number of eggs laid by the five females in each cage. Data were transformed, evaluated for homoscedasticity and sphericity then analysed using linear models with an interaction (R v. 2.13.0). We present p-values from two-tailed tests with α = 0.05. Means and 95% CI are reported.
(d). Eyespot manipulation and predation microcosm experiment
The developmental polyphenism of B. anynana alters multiple traits other than eyespot size and brightness which may influence predator–prey dynamics [17,18]. WS butterflies may be able to anticipate and escape a predator attack better than DS butterflies, independently of their wing pattern because they are more active than DS at the same ambient temperature and light environment [19]. Additionally, WS and DS butterflies have different eye sizes and ommatidia dimensions, suggesting visual plasticity between forms [20]. We used a 2 × 2 factorial design (butterfly form × eyespot form) to evaluate the effect of ventral wing eyespots on butterfly survival and reproduction in the presence of a mantid predator. We transplanted a small strip of the ventral hindwing margin with all the eyespots to each butterfly securing the wing piece with Superglue. The transverse white band of the two seasonal forms was not transferred. There were four treatments, all with mantids: DS butterfly × DS eyespots, DS butterfly × WS eyespots, WS butterfly × DS eyespots and WS butterfly × WS eyespots. All other experimental and analyses protocols were identical to the polyphenism and predation microcosm experiment above.
3. Results and discussion
(a). Eyespots on wet season butterflies increase prey detectability, influence attack location and increase survival
In the arena experiment, we found WS butterflies were easier for predators to detect, but harder to capture and more likely to escape with eyespot damage (figure 1). Mantids oriented on WS butterflies much sooner (DS 16.9 ± 4.3 s, WS 8.5 ± 3.1 s, F1,19 = 9.18, p = 0.0016; figure 1b). Many more WS butterflies escaped once attacked by mantids (DS 26.1% escaped, WS 69.2% escaped F1,19 = 6.59, p = 0.0015; figure 1c; electronic supplementary material, video). We observed 68.8% of attacks on the margin of the ventral hindwing of WS butterflies compared with 5.0% on the wing margin of DS butterflies (F1,19 = 8.71, p < 0.001; figure 1d; electronic supplementary material, video). Within WS, the ventral hindwing Cu1 eyespot received the most damage (53.8%) followed by Cu2 (22.5%) and Pc (19.9%) (F1,9 = 7.52, p < 0.001; figure 1e).
(b). Wet season butterflies live longer and reproduce more with mantids
WS butterflies survived longer, laid more eggs and suffered more hindwing damage when mantids were present (figure 2). Both butterfly forms survived longer without mantids (without mantid 42.9 ± 2.6; with mantid 4.8 ± 1.9 days, F1,44 = 442.08, p < 0.0001), and both forms lived a similar number of days controlling for predator treatments (F1,44 = 0.09, p = 0.7595). Mantids negatively impacted DS butterflies more; DS had lower survival than WS (DS with mantid 2.3 ± 1.0 days, WS with mantid 7.4 ± 2.8 days, F1,44 = 63.76, p < 0.0001; figure 2a). Both butterfly forms laid more eggs without mantids (without mantid 510.3 ± 13.6 eggs, with mantid 18.5 ± 2.4 eggs, F1,44 = 1230.75, p < 0.0001). Across predator treatments, WS laid more eggs than DS (DS 469.2 ± 15.3 eggs, WS 550.9 ± 19.4 eggs, F1,44 = 14.35, p = 0.001) as noted in previous studies [17,18]. Within mantids, WS butterflies laid more eggs (DS 6.7 ± 2.3 eggs, WS 31.0 ± 3.4 eggs, F1,22 = 14.35, p = 0.001). There was no interaction effect between predator presence and butterfly form (F1,44 = 2.64, p = 0.1060; figure 2b). Both butterfly forms suffered less ventral hindwing damage without mantids (without mantid 8.9 + 0.4%, with mantid 44.6 ± 0.3%, F1,44 = 702.96, p < 0.0001). Across predator treatments, WS butterflies suffered more ventral hindwing damage than DS (DS 13.4 ± 3.9%, WS 59.5 ± 3.7%, F1,44 = 467.14, p < 0.001). Mantids damaged WS butterflies more (DS 10.6 ± 1.9%, WS 70.4 ± 2.1%, F1,44 = 350.92, p < 0.0001; figure 2c).
Figure 2.
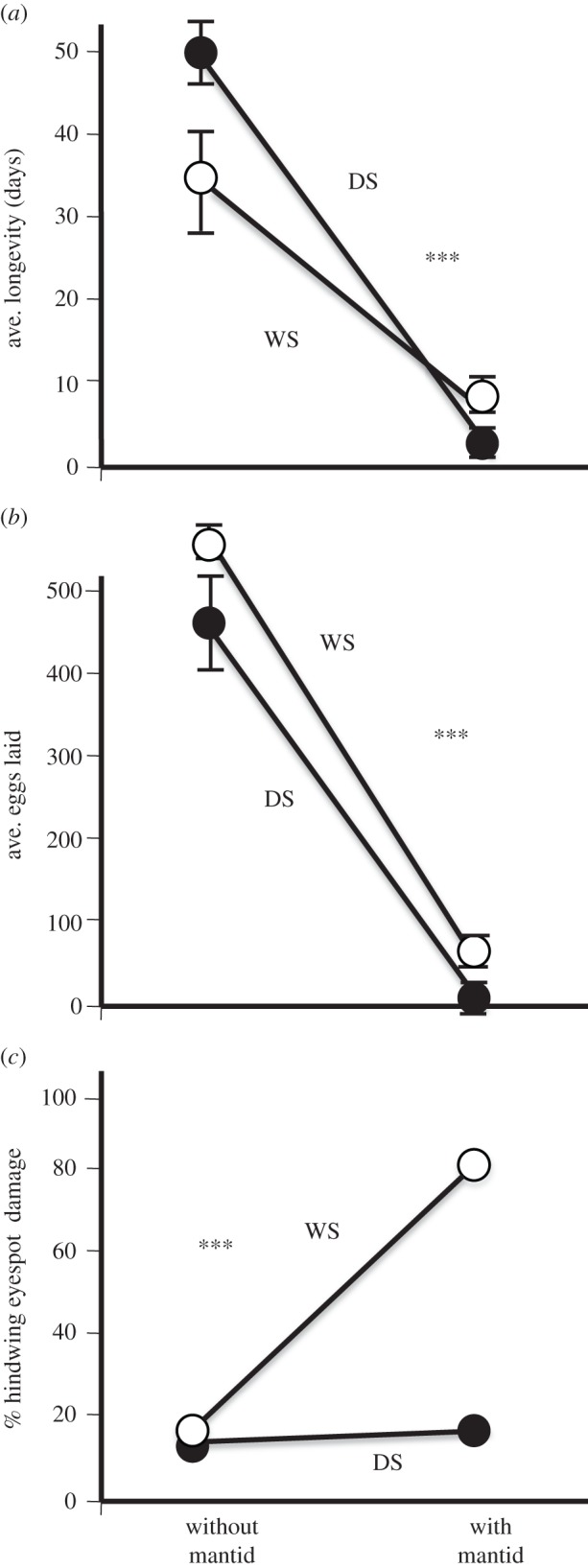
Longevity, fecundity and incurred wing damage for the DS and WS forms of B. anynana in microcosm experiments in the presence or the absence of mantids. (a) Average longevity measured in days. Means ± 95% CI are reported. DS forms survived longer in the absence of mantids while WS forms survived longer in the presence of mantids. (b) Average number of eggs laid. Means ± 95% CI are reported. WS form females laid more eggs, and mantids negatively impacted oviposition in both forms. (c) Percentage hindwing eyespot damage. WS forms experienced greater amounts of damage on their ventral hindwing eyespots in the presence of a mantid. Asterisks indicate statistical significance between the treatments.
(c). Eyespots alone deflect attack and increase butterfly fitness
WS eyespots, regardless of the form they were attached to, increased butterfly survival, reproductive output and ventral hindwing damage (figure 3). WS butterflies, controlling for eyespot treatment, survive longer than DS (DS form 5.1 ± 0.6 days, WS form 7.0 ± 0.3 days, F1,44 = 9.59, p = 0.0034). WS eyespots, regardless of the form they were attached to, increased butterfly longevity (DS eyespots 2.5 ± 0.4 days, WS eyespots 9.6 ± 0.3 days, F1,44 = 118.26, p < 0.0001). Mantids captured and consumed butterflies with DS eyespots faster than butterflies with WS eyespots regardless of form (DS form with DS eyespots 2.6 ± 0.5 days, WS form with DS eyespots 2.8 ± 0.5 days, DS form with WS eyespots 7.8 ± 0.4 days, WS form with WS eyespots 11.1 ± 0.6 days, F1,44 = 4.73, p = 0.0351; figure 3a). The WS form laid more eggs than the DS form controlling for eyespot treatment, but it was marginally significant (DS form 13.3 ± 1.4 eggs, WS form 18.2 ± 2.1 eggs, F1,44 = 3.88, p = 0.0666). Butterflies with WS eyespots laid more eggs regardless of form (DS eyespots 2.6 ± 0.4 eggs, WS eyespots 29.0 ± 0.8 eggs, F1,44 = 10.72, p < 0.0001). The interaction effect between form and eyespots had a greater impact on WS form than DS form (DS form with DS eyespots 3.5 ± 0.7 eggs, WS form with DS eyespots 1.6 ± 0.2 eggs, DS form with WS eyespots 23.2 ± 0.5 eggs, WS form with WS eyespots 34.9 ± 0.4 eggs, F1,44 = 6.84, p = 0.0122; figure 3b). The WS form experienced greater, albeit marginally significant, ventral hindwing damage (DS form 66.91 ± 2.71%, WS form 73.50 ± 4.34%, F1,44 = 1.835, p < 0.0733). WS eyespots regardless of form had much greater ventral hindwing damage (DS eyespots 17.92 ± 2.64%, WS eyespots 82.29 ± 2.64%, F1,44 = 24.647, p < 0.0001). The interaction between form and eyespot was not significant for per cent hindwing damage (F1,44 = 1.19, p = 0.2379; figure 3c).
Figure 3.
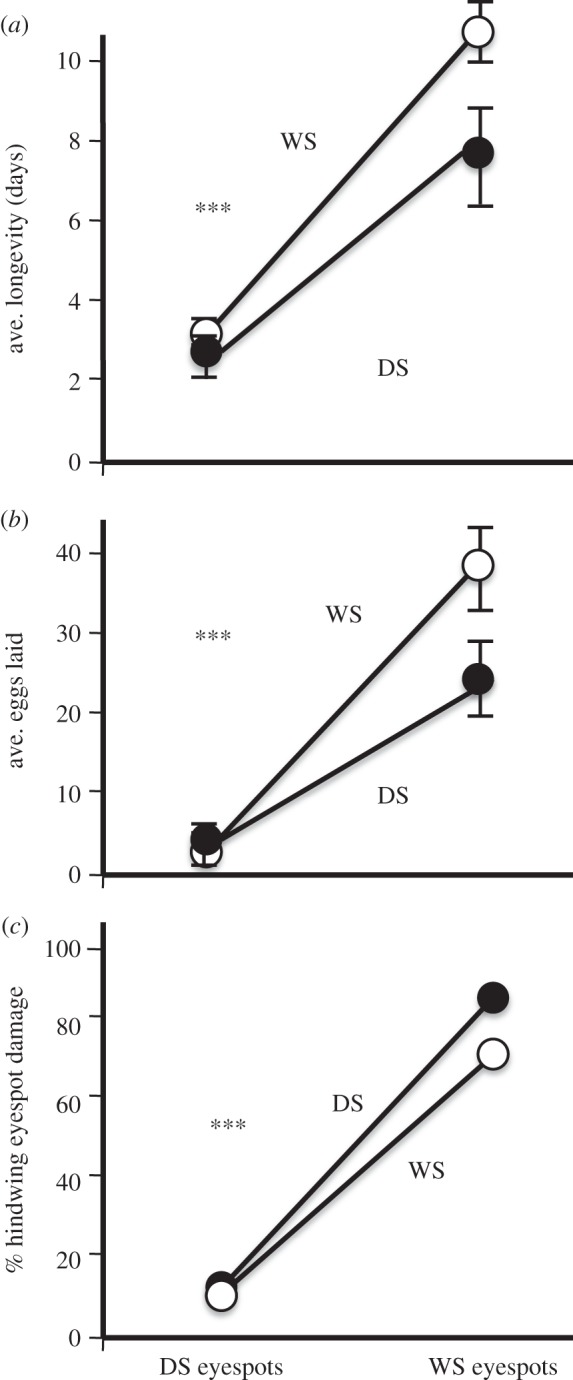
Longevity, fecundity and incurred wing damage for the DS and WS forms of B. anynana with transplanted ventral hindwing eyespots in microcosm experiments with mantids. (a) Average longevity measured in days. Means ± 95% CI are reported. Butterflies with WS eyespots survived much longer regardless of butterfly form. (b) Average number of eggs laid. Means ± 95% CI are reported. Butterflies with WS eyespots laid more eggs regardless of butterfly form. (c) Percentage hindwing eyespot damage. Butterflies with WS eyespots, regardless of butterfly form, exhibited greater amounts of damage on their ventral hindwing eyespots. Asterisks indicate statistical significance between the treatments.
(d). Eyespots are adaptive by deflecting invertebrate predator attack
We have demonstrated larger, brighter eyespots of the WS form of B. anynana are easier for an invertebrate predator to detect (figure 1b). Once detected, ventral hindwing eyespots direct predator attack to the wing margins, away from vital butterfly body parts, independently of butterfly form (figures 1d,e, 2c and 3c). Mantids attack and damage the largest eyespot on the ventral hindwing most frequently (Cu1) (figure 1e). Eyespots increase butterfly longevity and reproductive success when mantid predators are present in the local environment (figures 2a,b and 3a,b).
Past research indicated the large eyespots of WS B. anynana were not effective at deflecting vertebrate predator attack [7,8,10]. WS B. anynana were easier for vertebrate predators to detect; however, predators did not re-direct their attacks to the hindwing margin and WS B. anynana did not seem to accrue any fitness benefits for having large eyespots [7,8,10]. In these experiments, only approximately 4% of the numerous encounters between B. anynana and various vertebrate predators resulted in WS escape [7,8,10]. The discrepancy between vertebrate and invertebrate predator response to eyespots may be related to differences in the visual and nervous systems of the various experimental predators, or to the varying distances and angles from which distinct predators initiate their attack. Whatever the causes, mantids are misled by eyespot patterns more readily than vertebrate predators and alter their attack behaviour in ways that benefit survival and reproduction in WS B. anynana.
Across butterflies, eyespots have limited deflective effect on vertebrate predation under very specific light conditions. Low light conditions with accentuated UV promoted more avian attack to the wing margins of Lopinga achine [9]; however, these same butterflies suffered increased attacks under normal daylight conditions. Low light conditions are thought to increase the UV reflection of the white eyespot centre making the marginal eyespots more conspicuous to avian predators [9]. These findings are not transferable to mantids since mantids have monochromatic vision and are unable to see in the UV range [21], but the low light conditions may be relevant for other invertebrate predators, such as wasps, robber flies and spiders, which perceive signals in the UV range. More research is needed to understand the importance of predator identity, light conditions and their interaction on butterfly reproduction and survival.
Field surveys and mark recapture studies suggest butterfly and moth eyespot patterns deflect predator attack and are under selection in the wild. Individuals with more or larger eyespots have a higher recapture frequency and more damaged eyespots when recovered [22–24]. Also, the tensile strength of the wing regions where hindwing eyespots are found is often weaker than other areas of the wing and more easily torn [25,26]. These indirect observations have been attributed to selection by avian predators [22–26], but they are also consistent with pressure by invertebrate predators such as mantids. Mantids make similar damage patterns to beak marks and their attack has been noted for its speed and strength [13,27] (electronic supplementary material, video). Eyespot patterns on lepidopteran wings are probably under selection by a diverse community of both vertebrate and invertebrate predators.
Eyespots are found in a variety of other animals suggesting this colour pattern is of general adaptive importance. Investigations in marine and aquatic environments indicate eyespot patterns on fishes are used for predator intimidation or sexual attraction [21,28,29]. There remains at least some evidence eyespot may serve a deflective function. Fully reproductive butterfly fish are found in wild populations with up to 10% of their posterior body missing where eyespots are normally found [30]. Experiments using model prey found eyespots can attract attack by fish predators [31]. The deflection function may be working in conjunction with intimidation and sexual attraction in these systems. We hope our approach provides a useful experimental framework of combining ritualized predator–prey interactions with more general measurements of fitness for elucidating the various functions of eyespots across animals.
4. Conclusion
We leveraged a naturally occuring polyphenism in eyespot size to demonstrate larger, brighter eyespots deflected predator attack away from the more vulnerable body thereby increasing prey fitness. These marginal hindwing eyespots of WS B. anynana butterflies were more conspicuous to mantid predators; yet, these eyespots conferred greater survival and reproductive success compared to individuals with reduced, duller eyespots. Our results shed new light on the evolution of seasonal polyphenisms in B. anynana and other butterflies. Since ventral hindwing eyespots are beneficial to butterflies in the presence of invertebrate predators, these increases in reproduction and survival may offset the detection costs incurred against vertebrate predators [8]. Differences in phenology and longevity between invertebrate and vertebrate predators may explain the evolution of seasonal polyphenisms in eyespot size and colour.
Seasonal variation in eyespot size and other protective coloration has been primarily attributed to seasonal variation in avian predator intensity (reviewed in [1,2]). In B. anynana, the WS form is thought to have evolved as a result of relaxed avian predation during the rainy season coupled with increased avian predation in the dry season to produce the DS form [23]. Our results indicate increased invertebrate predation pressure may select for large, bright eyespots during the wet season while vertebrate predation pressure may select for small, dull eyespots in the dry season. This dynamic role of predator identity, their differences in vision, neurobiology and behaviour, in addition to predator abundance, may play an important role in the evolution of eyespot phenotypic plasticity.
Supplementary Material
Supplementary Material
Supplementary Material
Supplementary Material
Acknowledgements
We thank two anonymous reviewers and the associate editor for helpful comments.
Data accessibility
Data available from the Dryad Digital Repository: http://doi.org/10.5061/dryad.kr85.
Funding statement
We thank the Yale Institute for Biospheric Studies, the Donnelley Family, and Singapore Ministry of Education grant R-154-000-602-112 for funding this research.
References
- 1.Stevens M. 2005. The role of eyespots as anti-predator mechanisms, principally demonstrated in the Lepidoptera. Biol. Rev. 80, 573–588. ( 10.1017/S1464793105006810) [DOI] [PubMed] [Google Scholar]
- 2.Kodandaramaiah U. 2011. The evolutionary significance of butterfly eyespots. Behav. Ecol. 22, 1264–1271. ( 10.1093/beheco/arr123) [DOI] [Google Scholar]
- 3.Vallin A, Jakobsson S, Lind J, Wiklund C. 2005. Prey survival by predator intimidation: an experimental study of peacock butterfly defence against blue tits. Proc. R. Soc. B 272, 1203–1207. ( 10.1098/rspb.2004.3034) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Stevens M, Hopkins E, Hinde W, Adcock A, Connolly Y, Troscianko T, Cuthill IC. 2007. Field experiments on the effectiveness of ‘eyespots’ as predator deterrents. Anim. Behav. 74, 1215–1227. ( 10.1016/j.anbehav.2007.01.031) [DOI] [Google Scholar]
- 5.Kodandaramaiah U, Vallin A, Wiklund C. 2009. Fixed eyespot display in butterfly thwarts attacking birds. Anim. Behav. 77, 1415–1419. ( 10.1016/j.anbehav.2009.02.018) [DOI] [Google Scholar]
- 6.Olofsson M, Lovlie H, Tibblin J, Jakobsson S, Wiklund K. 2013. Eyespot display in the peacock butterfly triggers antipredator behaviors in naïve adult fowl. Behav. Ecol. 24, 305–310. ( 10.1093/beheco/ars167) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Lyytinen A, Brakefield PM, Mappes J. 2003. Significance of butterfly eyespots as an anti-predator device in ground-based and aerial attacks. Oikos 100, 373–379. ( 10.1034/j.1600-0706.2003.11935.x) [DOI] [Google Scholar]
- 8.Lyytinen A, Brakefield PM, Lindström L, Mappes J. 2004. Does predation maintain eyespot plasticity in Bicyclus anynana? Proc. R. Soc. B 271, 279–283. ( 10.1098/rspb.2003.2571) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Olofsson M, Vallin A, Jakobsson S, Wiklund C. 2010. Marginal eyespots on butterfly wings deflect bird attacks under low light intensities with UV wavelengths. PLoS ONE 5, e10798 ( 10.1371/journal.pone.0010798) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Vlieger L, Brakefield PM. 2007. The deflection hypothesis: eyespots on the margins of butterfly wings do not influence predation by lizards. Biol. J. Linn. Soc. 92, 661–667. ( 10.1111/j.1095-8312.2007.00863.x) [DOI] [Google Scholar]
- 11.Kjernsmo K, Meriliata S. 2013. Eyespots divert attacks by fish. Proc. R. Soc. B 280, 20131458 ( 10.1098/rspb.2013.1458) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Brakefield PM, Kesbeke F, Koch PB. 1998. The regulation of phenotypic plasticity of eyespots in the butterfly Bicyclus anynana. Am. Nat. 152, 853–860. ( 10.1086/286213) [DOI] [PubMed] [Google Scholar]
- 13.Prete FR, Wells H, Wells PH, Hurd LE. 1999. The preying mantids. Baltimore, MD: Johns Hopkins University Press. [Google Scholar]
- 14.Prudic KL, Skemp AK, Papaj DR. 2007. Aposematic coloration, luminance contrast, and the benefits of conspicuousness. Behav. Ecol. 18, 41–46. ( 10.1093/beheco/arl046) [DOI] [Google Scholar]
- 15.Picker M, Griffiths C, Weavings A. 2004. Field guide to insects of southern Africa. South Africa: Struik Publishers. [Google Scholar]
- 16.Spearman LA. 2009. Studies in the taxonomy and biodiversity of South African orthopteroids. Rutgers, NJ: Rutgers University Press. [Google Scholar]
- 17.Brakefield PM, Pijpe J, Zwaan BJ. 2007. Developmental plasticity and acclimation both contribute to adaptive responses to alternating seasons of plenty and of stress in Bicyclus butterflies. J. Biosciences 32, 465–475. ( 10.1007/s12038-007-0046-8) [DOI] [PubMed] [Google Scholar]
- 18.Geister TL, Lorenz MW, Meyering-Vos M, Hoffmann KH, Klaus F. 2008. Effects of temperature on reproductive output, egg provisioning, juvenile hormone and vitellogenin titres in the butterfly Bicyclus anynana. J. Insect Physiol. 54, 1253–1260. ( 10.1016/j.jinsphys.2008.06.002) [DOI] [PubMed] [Google Scholar]
- 19.Prudic KL, Jeon C, Cao H, Monteiro A. 2011. Developmental plasticity in sexual roles of butterfly species drives mutual sexual ornamentation. Science 331, 73–75. ( 10.1126/science.1197114) [DOI] [PubMed] [Google Scholar]
- 20.Everett A, Tong X, Briscoe AD, Monteiro A. 2012. Phenotypic plasticity in opsin expression in a butterfly compound eye complements sex role reversal. BMC Evol. Biol. 12, 232–242. ( 10.1186/1471-2148-12-232) [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Towner P, Gärtner W. 1994. The primary structure of mantid opsin. Gene 143, 227–231. ( 10.1016/0378-1119(94)90101-5) [DOI] [PubMed] [Google Scholar]
- 22.Swynnerton CFM. 1926. An investigation into the defences of butterflies of the genus Charaxes In 3rd International Congress of Entomology, Zurich, Switzerland, 1925, pp. 478–506. [Google Scholar]
- 23.Brakefield PM, Frankino WA. 2008. Polyphenisms in Lepidoptera: multidisciplinary approaches to studies of evolution. In Phenotypic plasticity of insects: mechanisms and consequences (eds Whitman DW, Ananthakrishnan TN.), pp. 121–152. Plymouth, UK: Science Publishers Inc. [Google Scholar]
- 24.Pinheiro CEG, Antezana MA, Machado LP. 2014. Evidence for the deflective function of eyespots in wild Junonia evarete Cramer (Lepidoptera, Nymphalidae). Neotrop. Entomol. 43, 39–47. ( 10.1007/s13744-013-0176-7) [DOI] [PubMed] [Google Scholar]
- 25.DeVries PJ. 2002. Differential wing toughness in distasteful and palatable butterflies: direct evidence supports unpalatability theory. Biotropica 34, 176–181. ( 10.1111/j.1744-7429.2002.tb00254.x) [DOI] [Google Scholar]
- 26.Hill RI, Vaca JF. 2004. Differential wing strength in Pierella butterflies (Nymphalidae, Satyrinae) supports the deflection hypothesis. Biotropica 36, 362–370. [Google Scholar]
- 27.Corrette BJ. 1990. Prey capture in the praying mantis Tenodera aridifolia sinensis: coordination of the capture sequence and strike movements. J. Exp. Biol. 148, 147–180. [DOI] [PubMed] [Google Scholar]
- 28.Gagliano M. 2008. On the spot: the absence of predators reveals eyespot plasticity in a marine fish. Behav. Ecol. 19, 733–739. ( 10.1093/beheco/arn013) [DOI] [Google Scholar]
- 29.Merilaita S, Vallin A, Kodandaramaiah U, Dimitrova M, Ruuskanen S, Laaksonen T. 2011. Number of eyespots and their intimidating effect on naïve predators in the peacock butterfly. Behav. Ecol. 22, 1326–1331. ( 10.1093/beheco/arr135) [DOI] [Google Scholar]
- 30.Neudecker S. 1989. Eye camouflage and false eyespots: chaetodontid responses to predators . Env. Biol. Fish. 25, 143–157. ( 10.1007/BF00002208) [DOI] [Google Scholar]
- 31.Kjernsmo K, Merilaita S. 2014. Eyespots divert attacks by fish. Proc. R. Soc. B 280, 20131458 ( 10.1098/rspb.2013.1458) [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Data Availability Statement
Data available from the Dryad Digital Repository: http://doi.org/10.5061/dryad.kr85.