

Abstract
Generally, wounds are of two categories, such as chronic and acute. Chronic wounds takes time to heal when compared to the acute wounds. Chronic wounds include vasculitis, non healing ulcer, pyoderma gangrenosum, and diseases that cause ischemia. Chronic wounds are rapidly increasing among the elderly population with dysfunctional valves in their lower extremity deep veins, ulcer, neuropathic foot and pressure ulcers. The process of the healing of wounds has several steps with the involvement of immune cells and several other cell types. There are many evidences supporting the hypothesis that apoptosis of immune cells is involved in the wound healing process by ending inflammatory condition. It is also involved in the resolution of various phases of tissue repair. During final steps of wound healing most of the endothelial cells, macrophages and myofibroblasts undergo apoptosis or exit from the wound, leaving a mass that contains few cells and consists mostly of collagen and other extracellular matrix proteins to provide strength to the healing tissue. This review discusses the various phases of wound healing both in the chronic and acute wounds especially during diabetes mellitus and thus support the hypothesis that the oxidative stress, apoptosis, connexins and other molecules involved in the regulation of chronic wound healing in diabetes mellitus and gives proper understanding of the mechanisms controlling apoptosis and tissue repair during diabetes and may eventually develop therapeutic modalities to fasten the healing process in diabetic patients.
Keywords: Apoptosis, Diabetes mellitus, Diabetic foot, Chronic wound, Oxidative stress
Core tip: Uncontrolled diabetes mellitus lead to the chronic non healing wound which further can escort to the Ischemia and coronary artery disease. Reports suggested that the involvement of various mechanisms in the development of chronic non healing wound in patients with diabetes mellitus, among which the oxidative stress plays a pivotal role which then leading to the enhanced apoptosis of lymphocytes, may be playing a critical role in the delay of wound healing. Connexins are gap junction protein and their upregulation during diabetes might be leads to improper gap junction formation attributing to the passage of various, apoptotic and inflammatory signals thereby resulting in delayed healing of chronic diabetic ulcers.
INTRODUCTION
Diabetes mellitus (DM) is a complex, chronic metabolic disorder; affects almost all age group of patients which requires continuous medical care with multifactorial risk reduction strategies beyond glycemic control[1]. Prolonged and uncontrolled DM may leads various complications which is broadly divided into microvascular complications (due to damage to small blood vessels) and macrovascular complications (due to damage to the arteries) affecting several organs, including muscle, skin, heart, brain, and kidneys.
It is reported that patients with DM are increasing rapidly worldwide and it is now recognized that the developing countries like India and China presently face the greatest burden of diabetes. It is the fourth or fifth leading cause of death in most high income countries caused 5.1 million deaths in 2013 and every six seconds a person dies due to diabetes[2]. According to International Diabetes Federation 382 million peoples were diagnosed with diabetes in 2013 which can reach up to 592 million in 2035. Among the countries China and India are having 98.4 and 65.1 million DM patients respectively in 2013 and which could be reach up to 142.7 million in china and 109.0 million in India[2]. Patients with poorly controlled diabetes may be subject to acute complications of diabetes, such as dehydration, poor wound healing, and hyperglycemic hyperosmolar coma.
Patients with DM have 15% higher risk for amputation than the general population due to chronic ulcers. It leads to diabetic neuropathy, which inhibits nociception and the perception of pain[3]. Due to loss of sensation in the feet of DM patients they become unaware of small wounds in the legs and feet, and may consequently fail to prevent infection or repeated injury on time[4]. Further, DM causes immune suppression and damage to small blood vessels, preventing adequate oxygenation of tissue, which can cause chronic wounds[4]. Immune deficiency also takes place in patients with type 2 DM (T2DM) due to the increased apoptosis of lymphocytes[5] and also the increased generation of reactive oxygen species (ROS) in patients with T2DM, might be another factor, which then stimulates downstream apoptotic signalling pathways[6].
In this connection, Desmoulière et al[7] reported that the decrease cellularity in wound repair process is achieved by apoptosis of different cell types. It is reported that the reduced rate of apoptosis is correlated with reduced expression of early growth response protein 1 (EGR1) in the 13 d old wound of epidermis of transgenic animal and the EGR1 mediate the proapaptotic signal via p53[8] and it clearly vindicated that the induced Egr1 expression plays a critical role in the resolution phase of wound repair by inducing apoptosis in keratinocytes. Further, it is suggested that the Egr1 expression is induced by various proteins among which transforming growth factor beta (TGF-β) is well known[9].
BASIC MECHANISM OF APOPTOSIS
The term “apoptosis” was coined by Kerr et al[10] for a morphologically distinct mode of cell death and the other type of cell death is known as necrosis. The key mechanism of apoptosis is endonuclease activation leading to internucleosomal double-stranded chromatin (DNA) fragmentation which occurs in most physiological cell death whereas cell membrane damage takes place in necrosis. Apoptosis is essential, as defects in apoptotic cell death regulation contribute to many diseases including disorders where deregulated cell proliferation occurs (cancer, restenosis) or where cell loss ensues (stroke, heart failure, neurodegeneration, Acquired Immune Deficiency Syndrome)[11]. In wound-healing process apoptosis is responsible for the removal of inflammatory cells and the evolution of granulation tissue into scar tissue[7]. In DM patients delayed wound healing is one of the major problems which are supposed to be takes place due to uncontrolled blood sugar level; it affects apoptosis during the wound healing process[12].
Apoptosis is also known as programmed cell death that may occur in multicellular organisms; leads to characteristic cell changes like blebbing, cell shrinkage, nuclear fragmentation, chromatin condensation, and chromosomal DNA fragmentation[13]. It is a complex process which initiates intracellular apoptotic signalling in response to a stress, which may bring about cell suicide. Cell suicide takes place in four separable but overlapping steps; induction, detection, effectors, and removal[14]. The dying cell remnants are removed by phagocytic cells of the macrophage/monocyte lineage. Interestingly, apoptotic bodies may also be engulfed by cells not specialized in phagocytosis (e.g., vascular smooth muscle cells) (Figure 1)[15].
Figure 1.
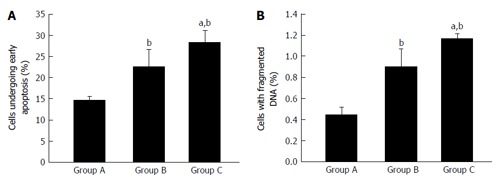
Percentage of apoptotic and dead cells in healthy (Group A), type 2 diabetes mellitus (Group B) and type 2 diabetes mellitus patients with chronic non healing wound (Group C) (A and B). bP < 0.01 vs healthy; aP < 0.05 vs uncontrolled diabetes without complication and uncontrolled diabetes with chronic non healing wound. First, second, and third bar in each panel represents healthy, uncontrolled diabetic and uncontrolled diabetic with chronic non healing wound, respectively.
T2DM is associated with elevated level of oxidative stress, which is one of the most important factors responsible for the development of chronic complications of this disease. Antioxidants like reduced glutathione (GSH), superoxide dismutase (SOD) and catalase protects cells against oxidative damages. In our own publication we have shown that oxidative stress is higher in T2DM patients. In T2DM patients with chronic non healing wound, lymphocyte apoptosis is initiated by the augmentation of reactive oxygen species which leads to the increased expression of proapoptotic proteins like Caspases, FAS, BAX and decreased expression of antiapoptotic proteins like B-cell lymphoma 2 genes (Bcl-2) (Figure 2)[6].
Figure 2.
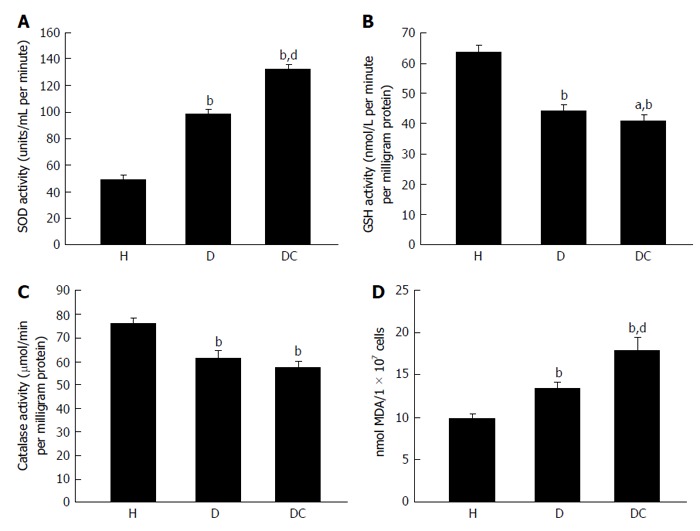
Concentration of superoxide dismutase (A), reduced glutathione (B), catalase (C) and malondialdehyde (D) in healthy (H), type 2 diabetes mellitus (D) and type 2 diabetes mellitus patients with chronic non healing (DC) groups. bP < 0.01 vs healthy; dP < 0.01 and aP < 0.05 vs uncontrolled diabetes without complication and uncontrolled diabetes with chronic non healing wound. First, second, and third bar in each panel represents healthy, uncontrolled diabetic and uncontrolled diabetic with chronic non healing wound, respectively. SOD: Superoxide dismutase; GSH: Reduced glutathione; MDA: Malondialdehyde.
In streptozotocin-induced diabetic rats, the elevated blood sugar level increases cellular apoptosis and the least expression of Bcl-2 protein causes deregulation of the wound healing processes (Tables 1 and 2)[16].
Table 1.
Mean blood glucose level, apoptotic index and DNA fragmentation in control rats (P value < 0.01)
| 5th day | 10th day | 20th day | 30th day | |
| Control (n =10) blood glucose (mg/dL) | 75.62 ± 6.41 | 80.79 ± 11.45 | 92.05 ± 9.56 | 90.77 ± 9.7 |
| Apoptotic index (mean ± SD) | 1.50 ± 0.60 | 1.60 ± 0.99 | 1.64 ± 0.86 | 1.69 ± 1.12 |
| DNA fragmentation (%) (mean ± SD) | 42.25 ± 3.95 | 44.15 ± 5.61 | 45.45 ± 5.88 | 46.58 ± 5.95 |
Table 2.
Mean blood glucose level, apoptotic index, and DNA fragmentation in rats with diabetes (P value < 0.01)
| 5th day | 10th day | 20th day | 30th day | |
| With diabetes (n = 10) blood glucose (mg/dL) | 467.25 ± 48.2 | 506.33 ± 35.89 | 474.99 ± 39.76 | 488.15 ± 34.36 |
| Apoptotic index (mean ± SD) | 3.50 ± 2.60 | 4.20 ± 2.99 | 3.60 ± 3.56 | 3.69 ± 2.75 |
| DNA fragmentation (mean ± SD) | 62.80 ± 9.56 | 74.95 ± 10.45 | 66.55 ± 8.67 | 70.48 ± 6.21 |
The mechanism of apoptosis has been linked with several proteins but two of them are extensively recognised for their regulation in the pathways (Figure 3)[17]: (1) targeting mitochondria functionality, or directly transducing the signal via adaptor proteins, known as intrinsic pathway; and (2) extrinsic pathway of initiation as identified in several toxin studies is an increase in calcium concentration within a cell caused by drug activity, which can also cause apoptosis via calcium binding protease calpain.
Figure 3.
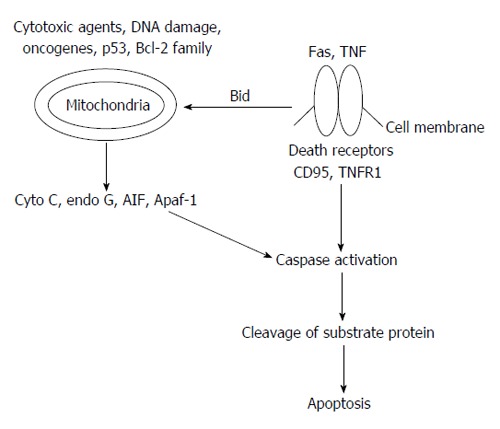
Basic outline of apoptosis mechanism. Bcl-2: B-cell lymphoma 2; TNF: Tumor necrosis factor; AIF: Apoptosis-inducing factor; Apaf-1: Apoptotic protease activating factor-1; TNFR1: Tumor necrosis factor receptor 1.
In the wound healing process various expression patterns of apoptosis key regulators have been studied which shows that the healing in mucosa takes place predominantly through the intrinsic pathway whereas skin healing is predominantly through the extrinsic pathway. The identification of differences in the apoptotic pathways involved in wound healing of various organs may allow the development of therapeutics to improve wound healing[18].
INTRINSIC PATHWAY
The intrinsic signalling pathways involve various arrays of non-receptor-mediated stimuli that produce intracellular signals to work immediately on objects within the cell and are mitochondrial-initiated events. Intrinsic pathway acts both as proapoptotic or antiapoptotic fashion and depends upon the intracellular signals. Negative signals involve the lack of certain growth factors, hormones and cytokines that can escort to collapse of death programs inhibition, thereby triggering apoptosis. Other stimuli that act in encouraging fashion of apoptosis include radiation, toxins, hypoxia, hyperthermia, viral infections, and free radicals, etc.
Stimulus of apoptotic proteins targeting inner membrane of mitochondria may cause mitochondrial swelling through the formation of mitochondrial permeability transition (MPT) pore, or they may increase the permeability of the mitochondrial membrane and cause apoptotic effectors to leak out[19]. Formation of MPT is achieved by the group of proteins consist of cytochrome c, Smac/DIABLO, and the serine protease HtrA2/Omi. The release of cytochrome c into the cytoplasm appears to be a crucial step for the activation of caspase. Once cytochrome c is released it binds with Apoptotic protease activating factor-1 and ATP, which then tie up to pro-caspase-9 to create a protein complex known as apoptosome. The apoptosome cleaves the pro-caspase to its active form of caspase-9, which in turn activates the effector caspase-3. Smac/DIABLO and HtrA2/Omi promote apoptosis by inhibiting inhibitors of apoptosis proteins activity[20].
In addition to the release of cytochrome c; apoptosis-inducing factor (AIF), endonuclease G and Caspase Activated DNAse (CAD), discharge from the mitochondria during apoptosis. AIF translocates to the nucleus and causes DNA fragmentation into about 50-300 kb pieces and condensation of peripheral nuclear chromatin[21] whereas Endonuclease G translocates to the nucleus where it cleaves nuclear chromatin to produce oligonucleosomal DNA fragments[22]. CAD is subsequently discharged from the mitochondria and translocates to the nucleus where after cleavage by caspase-3, it leads to oligonucleosomal DNA fragmentation and chromatin condensation[23]. The control and regulation of these apoptotic mitochondrial events occur through members of the Bcl-2 family of proteins[24]. Bcl-2 proteins are able to promote or inhibit apoptosis by direct action on MAC/MOMPP. Bax and/or Bak form the pore, while Bcl-2, Bcl-xL or Mcl-1 inhibits its formation.
EXTRINSIC PATHWAY
The extrinsic signaling pathways involve death receptors that are members of the tumor necrosis factor (TNF) receptor gene superfamily[25]. Members of the TNF receptor family share similar cysteine-rich extracellular domains and have a cytoplasmic domain of about 80 amino acids called the “death domain”[26]. This death domain plays a critical role in transmitting the death signal from the cell surface to the intracellular signaling pathways.
TNF-α signaling is linked to the Fas signaling pathway through the interaction of TNF receptor-associated death domain protein with Fas-associated death domain protein and their activation is critically depends upon the activation of caspase[27]. Once caspase-8 is activated, the execution phase of apoptosis is triggered. The binding of three Fas molecules to a Fas ligand (FasL) homotrimer leads to the subsequent binding of Fas-associated death domain and procaspase-8 which finally triggers a cascade of caspase activation, including caspase-3, leading to cell death[28]. Diabetes-enhanced and prolonged expression of TNF-α and contributes in the direction of impaired healing[29]. TNF-α is found threefold higher in diabetic mouse wounds than wounds in normal mice[30] and threefold higher found in wound fluid from nonhealing venous leg ulcers than in healing ulcers[31].
EXECUTION PATHWAY OF APOPTOSIS
Execution pathways start from the end point of intrinsic and extrinsic pathways of apoptosis. In this phase execution caspase activates to start organized degradation of cellular organelles. Caspase-3 is considered to be the most important of the executioner caspases and is activated by any of the initiator caspases (caspase-8, caspase-9, or caspase-10)[23]. Phagocytic uptake of apoptotic cells is the last component of apoptosis. Mice lacking either of these caspases were deficient in skin wound healing and in liver regeneration[32].
Phospholipid asymmetry and externalization of phosphatidylserine on the surface of apoptotic cells and their fragments is the characteristic feature of cell death which can be measured by fluorescent activated cell sorter using annexin V tagged with fluorescent molecule[5].
DIABETIC WOUND HEALING AND APOPTOSIS
Usually wound healing process can be split into 4 temporarily and spatially overlapping phases: coagulation, inflammation, tissue formation (proliferative phase) and tissue remodelling or scar formation phase.
COAGULATION PHASE
Coagulation phase takes place immediately after injury to stop excessive blood flow from wound and provides provisional protection for the wounded area. Hemostatic reaction started with the adherence of platelets to damaged blood vessels giving rise to a blood-clotting cascade. To facilitate aggregation platelates express sticky glycoproteins on their cell membrane[33]. Platelets also released cytokines and growth factors which are a potent chemotactic agent; stimulates the deposition of extracellular membrane to the wound site[34]. In addition, platelets release proinflammatory factors like serotonin, bradykinin, prostaglandins, prostacyclins, thromboxane, and histamine to dilate blood vessel and increase cell proliferation and migration to the wound area[35].
INFLAMMATORY PHASE
Inflammatory phase starts with the release of platelet-derived growth factor and TGF-A1 and TGF-2 from platelet which attract inflammatory cells, such as leukocytes, neutrophils, and macrophages[36]. Leukocytes release ROS that are antimicrobial and proteases that clear the wound of foreign bodies and bacteria. T lymphocytes playing central role in the wound healing[37] and its increased apoptosis leading to delayed wound healing in diabetic patients[17]. Neutrophils are important in wound healing as they serve to control infection by eliminating microorganisms. With the control of infections neutrophils also release harmful enzymes which damage healthy tissues surrounding the wound site. To prevent further inflammation neutrophils are engulfed by macrophages during the process of apoptosis[38]. Macrophages are the key scavengers for resolving inflammation and facilitating tissue regrowth[39]. These findings show that apoptosis of immune cells could be the major key to end inflammation and initiate healing[40].
Diabetes impaired wound healing by reducing macrophage number and activation which results in the reduced lymphatic vessel formation[41]. The anti proliferative protein p53 involved in apoptosis of inflammatory cells during the healing process and its expression during the healing of cutaneous wounds in swine has been reported by Antoniades et al[42].
PROLIFERATIVE PHASE
Proliferative phase of repair begins with the settling down of inflammatory phase and formation of granulation tissue. Granulation tissue formation takes place by growth factors which are released by basal keratinocytes, remaining inflammatory cells and migrating epidermal and dermal cells to support the epithelialization process of wound healing[36]. Diabetes mellitus affects re-epithelialization by affecting multiple proteins and genes including angiopoietin-4[43]. ANGPTL4 shows a potential effect on lipid homeostasis, glucose metabolism, re-epithelialization, inflammation, and potential effect on energy homoeostasis, which is required for wound healing. In corneal wound healing; apoptosis of stromal keratinocyte is well characterised. It triggers subsequent cellular processes that include bone marrow-derived cell infiltration, proliferation, and migration of residual keratinocyte cells and in some circumstances, generation of myofibroblast cells[44].
Diabetes mellitus affects signalling intermediates responsible for coordinating/regulating wound healing angiogenesis and vasculogenesis[45]. Due to the deficiencies in either endothelial progenitor cell or peripheral tissue homing and engraftment of bone marrow, diabetic patients are prone to the development of chronic wounds[46].
TISSUE REMODELING
Tissue remodeling is the process of reformation or restoration of existing tissues. Restoration of a normal blood supply offers an encouraging microenvironment for epidermal and dermal cell migration and proliferation. Fibroblasts proliferate within the wound and synthesize extra-cellular matrix (ECM) forming granulation tissue perfused with newly formed blood vessels.
Wound contraction and matrix remodeling occurs after the substitution of ECM from collagen III, fibrin, fibronectin, and hyaluronic acid[36]. Collagen homeostasis is aberrant in the wound of uncontrolled DM patients who suppose to be mediated by Hsp47; leading to the dysfunction of fibroblast cells. Such impairments could contribute to delayed wound healing[47]. With wound maturation, different cell populations need to be eliminated. Apoptosis of fibroblastic cells occurs, leading to the formation of a relatively acellular scar tissue whose tensile strength is equivalent with unwounded skin. Early studies suggest that endothelial cells undergo apoptosis followed by the removal of myofibroblasts[48].
The passage of various apoptotic and inflammatory signals via gap junctions play an important role in tissue remodelling during diabetic wound healing. Connexins (Cx), the gap junction proteins, form channels between two adjacent cells and their expression is highly regulated after wound formation at the transcriptional, translational and post translational levels[49]. In diabetic wounds significant increase in the levels of Cx26, Cx30.3, Cx31, Cx31.1, and Cx43 were observed as compared to non-diabetic wounds[50]. An up regulated connexin expression might lead to the improper gap junction formation attributing to the passage of various, apoptotic and inflammatory signals thereby resulting in delayed healing of chronic diabetic ulcers.
CONCLUSION
Diabetes mellitus delayed normal wound healing by various ways like narrowing of the blood vessels due to arteriosclerosis or leading decreased blood flow and oxygen to a wound, loss of sensation in feet and lowering down the efficiency of the immune system. DM is leading various complications like macroangiopathy and microangiopathy among which Chronic wounds such as venous ulcers are rapidly increasing. In chronic non healing DM patients various cytokines and chemokines are interacting together to lead various complications, e.g., strong positive association between interleukin-7 and monocyte chemoattractant protein 1 may be a possible cause of developing coronary artery disease in these patients[51]. Dysregulation of apoptosis in response to hyperglycemia is universal, leading to impaired wound healing along with the involvement of other target organs. Contrary to the accepted view that diabetic foot is caused by neuropathy and peripheral vascular disease, it now appears that dysregulated apoptosis is emerging as a major cause of the diabetic foot wound. Recent advances in management of DM and understanding of the molecular and cellular components of apoptosis involved during the wound healing phases may enable personalized diagnosis and therapy tailored to a particular patient’s needs and therefore lead to better therapeutic outcomes.
ACKNOWLEDGMENTS
Author AK Arya is thankful to Planning and Budgeting Committee, Council for Higher Education, Israel.
Footnotes
P- Reviewer: Liu C, Sanchez R S- Editor: Qi Y L- Editor: A E- Editor: Liu SQ
References
- 1.American Diabetes Association. Standards of Medical Care in Diabetes. Diabetes Care. 2014;37:S14–S80. doi: 10.2337/dc14-S014. [DOI] [PubMed] [Google Scholar]
- 2.International Diabetes Federation. IDF Diabetes Atlas, 6th ed. Brussels, Belgium: International Diabetes Federation; 2013. Available from: http: //www.idf.org/diabetesatlas. [Google Scholar]
- 3.Snyder RJ. Treatment of nonhealing ulcers with allografts. Clin Dermatol. 2005;23:388–395. doi: 10.1016/j.clindermatol.2004.07.020. [DOI] [PubMed] [Google Scholar]
- 4.Moreo K. Understanding and overcoming the challenges of effective case management for patients with chronic wounds. Case Manager. 2005;16:62–63, 67. doi: 10.1016/j.casemgr.2005.01.014. [DOI] [PubMed] [Google Scholar]
- 5.Arya AK, Garg S, Kumar S, Meena LP, Tripathi K. Estimation of lymphocyte apoptosis in patients with chronic non-healing diabetic foot ulcer. Int J Med Sci Pub Health. 2013;2:766–768. [Google Scholar]
- 6.Arya AK, Pokharia D, Tripathi K. Relationship between oxidative stress and apoptotic markers in lymphocytes of diabetic patients with chronic non healing wound. Diabetes Res Clin Pract. 2011;94:377–384. doi: 10.1016/j.diabres.2011.08.004. [DOI] [PubMed] [Google Scholar]
- 7.Desmoulière A, Redard M, Darby I, Gabbiani G. Apoptosis mediates the decrease in cellularity during the transition between granulation tissue and scar. Am J Pathol. 1995;146:56–66. [PMC free article] [PubMed] [Google Scholar]
- 8.Nair P, Muthukkumar S, Sells SF, Han SS, Sukhatme VP, Rangnekar VM. Early growth response-1-dependent apoptosis is mediated by p53. J Biol Chem. 1997;272:20131–20138. doi: 10.1074/jbc.272.32.20131. [DOI] [PubMed] [Google Scholar]
- 9.Amendt C, Mann A, Schirmacher P, Blessing M. Resistance of keratinocytes to TGFbeta-mediated growth restriction and apoptosis induction accelerates re-epithelialization in skin wounds. J Cell Sci. 2002;115:2189–2198. doi: 10.1242/jcs.115.10.2189. [DOI] [PubMed] [Google Scholar]
- 10.Kerr JF, Wyllie AH, Currie AR. Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics. Br J Cancer. 1972;26:239–257. doi: 10.1038/bjc.1972.33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Reed JC. Mechanisms of apoptosis. Am J Pathol. 2000;157:1415–1430. doi: 10.1016/S0002-9440(10)64779-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kumar H, Mishra M, Pokharia D, Kumar L, Arya AK, Shukla VK and Tripathi K. Glycemic control affects the apoptosis during delayed diabetic wound healing. Canada: Proceedings of the World Union of wound Healing societies; 2008. [Google Scholar]
- 13.Green Douglas R. Means to an End: Apoptosis and other Cell Death Mechanisms. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press; 2011. [Google Scholar]
- 14.Teraki Y, Shiohara T. Apoptosis and the skin. Eur J Dermatol. 1999;9:413–425; quiz 426. [PubMed] [Google Scholar]
- 15.Bennett MR, Gibson DF, Schwartz SM, Tait JF. Binding and phagocytosis of apoptotic vascular smooth muscle cells is mediated in part by exposure of phosphatidylserine. Circ Res. 1995;77:1136–1142. doi: 10.1161/01.res.77.6.1136. [DOI] [PubMed] [Google Scholar]
- 16.Bhan S, Mitra R, Arya AK, Pandey HP, Tripathi K. A study on evaluation of apoptosis and expression of bcl-2-related marker in wound healing of streptozotocin-induced diabetic rats. ISRN Dermatol. 2013;2013:739054. doi: 10.1155/2013/739054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Arya AK, Pokharia D, Kumar H, Mishra M, and Tripathi K. Impact of lymphocyte apoptosis in diabetes mellitus. Asian J Med Sci. 2011;2:1–6. [Google Scholar]
- 18.Johnson A, Francis M, DiPietro L. Differential apoptosis in mucosal and dermal wound healing. Adv Wound Care. 2013 doi: 10.1089/wound.2012.0418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Saelens X, Festjens N, Vande Walle L, van Gurp M, van Loo G, Vandenabeele P. Toxic proteins released from mitochondria in cell death. Oncogene. 2004;23:2861–2874. doi: 10.1038/sj.onc.1207523. [DOI] [PubMed] [Google Scholar]
- 20.Schimmer AD. Inhibitor of apoptosis proteins: translating basic knowledge into clinical practice. Cancer Res. 2004;64:7183–7190. doi: 10.1158/0008-5472.CAN-04-1918. [DOI] [PubMed] [Google Scholar]
- 21.Joza N, Susin SA, Daugas E, Stanford WL, Cho SK, Li CY, Sasaki T, Elia AJ, Cheng HY, Ravagnan L, et al. Essential role of the mitochondrial apoptosis-inducing factor in programmed cell death. Nature. 2001;410:549–554. doi: 10.1038/35069004. [DOI] [PubMed] [Google Scholar]
- 22.Li LY, Luo X, Wang X. Endonuclease G is an apoptotic DNase when released from mitochondria. Nature. 2001;412:95–99. doi: 10.1038/35083620. [DOI] [PubMed] [Google Scholar]
- 23.Enari M, Sakahira H, Yokoyama H, Okawa K, Iwamatsu A, Nagata S. A caspase-activated DNase that degrades DNA during apoptosis, and its inhibitor ICAD. Nature. 1998;391:43–50. doi: 10.1038/34112. [DOI] [PubMed] [Google Scholar]
- 24.Cory S, Adams JM. The Bcl2 family: regulators of the cellular life-or-death switch. Nat Rev Cancer. 2002;2:647–656. doi: 10.1038/nrc883. [DOI] [PubMed] [Google Scholar]
- 25.Locksley RM, Killeen N, Lenardo MJ. The TNF and TNF receptor superfamilies: integrating mammalian biology. Cell. 2001;104:487–501. doi: 10.1016/s0092-8674(01)00237-9. [DOI] [PubMed] [Google Scholar]
- 26.Ashkenazi A, Dixit VM. Death receptors: signaling and modulation. Science. 1998;281:1305–1308. doi: 10.1126/science.281.5381.1305. [DOI] [PubMed] [Google Scholar]
- 27.Longthorne VL, Williams GT. Caspase activity is required for commitment to Fas-mediated apoptosis. EMBO J. 1997;16:3805–3812. doi: 10.1093/emboj/16.13.3805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Fadeel B, Orrenius S, Zhivotovsky B. Apoptosis in human disease: a new skin for the old ceremony? Biochem Biophys Res Commun. 1999;266:699–717. doi: 10.1006/bbrc.1999.1888. [DOI] [PubMed] [Google Scholar]
- 29.Giannone G, Mazzone O, Russo A, Trovato GM. [Electrocardiographic findings during colonoscopy] Boll Soc Ital Cardiol. 1978;23:441–446. [PubMed] [Google Scholar]
- 30.Siqueira MF, Li J, Chehab L, Desta T, Chino T, Krothpali N, Behl Y, Alikhani M, Yang J, Braasch C, et al. Impaired wound healing in mouse models of diabetes is mediated by TNF-alpha dysregulation and associated with enhanced activation of forkhead box O1 (FOXO1) Diabetologia. 2010;53:378–388. doi: 10.1007/s00125-009-1529-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Wallace HJ, Stacey MC. Levels of tumor necrosis factor-alpha (TNF-alpha) and soluble TNF receptors in chronic venous leg ulcers--correlations to healing status. J Invest Dermatol. 1998;110:292–296. doi: 10.1046/j.1523-1747.1998.00113.x. [DOI] [PubMed] [Google Scholar]
- 32.Li F, Huang Q, Chen J, Peng Y, Roop DR, Bedford JS, Li CY. Apoptotic cells activate the “phoenix rising” pathway to promote wound healing and tissue regeneration. Sci Signal. 2010;3:ra13. doi: 10.1126/scisignal.2000634. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Midwood KS, Williams LV, Schwarzbauer JE. Tissue repair and the dynamics of the extracellular matrix. Int J Biochem Cell Biol. 2004;36:1031–1037. doi: 10.1016/j.biocel.2003.12.003. [DOI] [PubMed] [Google Scholar]
- 34.Rosenberg L, de la Torre J. Wound Healing, Growth Factors. 2006. Available from: http: //emedicine.medscape.com/article/1298196-overview. [Google Scholar]
- 35.Stadelmann WK, Digenis AG, Tobin GR. Physiology and healing dynamics of chronic cutaneous wounds. Am J Surg. 1998;176:26S–38S. doi: 10.1016/s0002-9610(98)00183-4. [DOI] [PubMed] [Google Scholar]
- 36.Singer AJ, Clark RA. Cutaneous wound healing. N Engl J Med. 1999;341:738–746. doi: 10.1056/NEJM199909023411006. [DOI] [PubMed] [Google Scholar]
- 37.Barbul A, Regan MC. The regulatory role of T lymphocytes in wound healing. J Trauma. 1990;30:S97–100. doi: 10.1097/00005373-199012001-00021. [DOI] [PubMed] [Google Scholar]
- 38.Brubaker AL, Schneider DF, Kovacs EJ. Neutrophils and natural killer T cells as negative regulators of wound healing. Expert Rev Dermatol. 2011;6:5–8. doi: 10.1586/edm.10.66. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Li L, Yan B, Shi YQ, Zhang WQ, Wen ZL. Live imaging reveals differing roles of macrophages and neutrophils during zebrafish tail fin regeneration. J Biol Chem. 2012;287:25353–25360. doi: 10.1074/jbc.M112.349126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Wu YS, Chen SN. Apoptotic cell: linkage of inflammation and wound healing. Front Pharmacol. 2014;5:1. doi: 10.3389/fphar.2014.00001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Maruyama K, Asai J, Ii M, Thorne T, Losordo DW, D’Amore PA. Decreased macrophage number and activation lead to reduced lymphatic vessel formation and contribute to impaired diabetic wound healing. Am J Pathol. 2007;170:1178–1191. doi: 10.2353/ajpath.2007.060018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Antoniades HN, Galanopoulos T, Neville-Golden J, Kiritsy CP, Lynch SE. p53 expression during normal tissue regeneration in response to acute cutaneous injury in swine. J Clin Invest. 1994;93:2206–2214. doi: 10.1172/JCI117217. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Arya AK, Tripathi K, Das P. Promising role of ANGPTL4 gene in diabetic wound healing. Int J Low Extrem Wounds. 2014;13:58–63. doi: 10.1177/1534734614520704. [DOI] [PubMed] [Google Scholar]
- 44.Wilson SE, Chaurasia SS, Medeiros FW. Apoptosis in the initiation, modulation and termination of the corneal wound healing response. Exp Eye Res. 2007;85:305–311. doi: 10.1016/j.exer.2007.06.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Gallagher KA, Liu ZJ, Xiao M, Chen H, Goldstein LJ, Buerk DG, Nedeau A, Thom SR, Velazquez OC. Diabetic impairments in NO-mediated endothelial progenitor cell mobilization and homing are reversed by hyperoxia and SDF-1 alpha. J Clin Invest. 2007;117:1249–1259. doi: 10.1172/JCI29710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Liu ZJ, Velazquez OC. Hyperoxia, endothelial progenitor cell mobilization, and diabetic wound healing. Antioxid Redox Signal. 2008;10:1869–1882. doi: 10.1089/ars.2008.2121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Pokharia D, Kumar H, Mishra M, Arya Ak, Tripathi R, Singh RK, Kumar M Tripathi K. Study of heat Shock protein Hsp47 expression in chronic wound of controlled and uncontrolled type 2 diabetes mellitus. Diabetes. 2012;61 Suppl 10:A165. [Google Scholar]
- 48.Compton CC, Gill JM, Bradford DA, Regauer S, Gallico GG, O’Connor NE. Skin regenerated from cultured epithelial autografts on full-thickness burn wounds from 6 days to 5 years after grafting. A light, electron microscopic and immunohistochemical study. Lab Invest. 1989;60:600–612. [PubMed] [Google Scholar]
- 49.Wang CM, Lincoln J, Cook JE, Becker DL. Abnormal connexin expression underlies delayed wound healing in diabetic skin. Diabetes. 2007;56:2809–2817. doi: 10.2337/db07-0613. [DOI] [PubMed] [Google Scholar]
- 50.Bajpai S, Mishra M, Kumar H, Tripathi K. Singh S K, Srikrishna S, Prasad H, Singh R K. Comparative study of connexins expression in diabetic and nondiabetic wounds. Digest J Nanomat Biostruct. 2010;5:757–762. [Google Scholar]
- 51.Arya AK, Pokharia D, Bhan S, Tripathi R, Tripathi K. Correlation between IL-7 and MCP-1 in diabetic chronic non healing ulcer patients at higher risk of coronary artery disease. Cytokine. 2012;60:767–771. doi: 10.1016/j.cyto.2012.07.034. [DOI] [PubMed] [Google Scholar]