

Abstract
Long non-coding RNAs (lncRNAs) govern fundamental biochemical and cellular processes. lncRNA HOX transcript antisense RNA (HOTAIR) represses gene expression through recruitment of chromatin modifiers. The expression of HOTAIR is elevated in lung cancer and correlates with metastasis and poor prognosis. Moreover, HOTAIR promotes proliferation, survival, invasion, metastasis, and drug resistance in lung cancer cells. Here we review the molecular mechanisms underlying HOTAIR-mediated aggressive phenotypes of lung cancer. We also discuss HOTAIR’s potential in diagnosis and treatment of lung cancer, as well as the challenges of exploiting HOTAIR for intervention of lung cancer.
Electronic supplementary material
The online version of this article (doi:10.1186/s13045-014-0090-4) contains supplementary material, which is available to authorized users.
Keywords: lncRNA, HOTAIR, Lung cancer, PRC2, Metastasis
lncRNAs as novel master regulators of lung cancer
A surprising discovery of the ENCODE project is that 87.3% of the human genome is actively transcribed although only < 3% of the human genome encodes proteins [1]. One family of the non protein-coding RNAs is operationally defined as long non-coding RNAs (lncRNAs) based on their length > 200 nucleotides [2]. As published in GENCODE v7 (2012), the lncRNA catalogue comprises 9277 manually annotated lncRNA genes that produce 14,880 transcripts [3]. lncRNAs regulate fundamental biochemical and cellular processes, such as gene expression, RNA splicing, and ligand-receptor engagement, which mediates pathogenesis of benign and malignant respiratory disorders [4],[5].
lncRNAs have emerged as novel master regulators of initiation, progression, and response to therapy in a wide variety of solid tumors and hematological malignancies [6],[7]. Hundreds of IncRNAs have been associated with lung cancer through gene expression microarrays and massively parallel RNA sequencing of tumor tissues and paired adjacent non-tumor tissues in the lung [8]-[11]. As of September 2014, a PubMed search using lncRNA and lung cancer as key words yielded more than a dozen of lncRNAs that have been individually investigated in lung cancer (Table 1) [8],[12]-[38]. Despite their largely descriptive and correlative nature, these reports highlight a critical role of lncRNAs in lung cancer. The investigated lncRNAs regulate critical cellular processes in lung cancer, such as proliferation, invasion, and survival (Table 1). Moreover, dysregulated expression of these lncRNAs is correlated with metastasis, advanced pathological stages, and poor prognosis in patients with lung cancer (Table 1).
Table 1.
Lung cancer-associated lncRNAs
| lncRNA | Intersecting molecules and pathways | Cell processes | Associated clinical features |
|---|---|---|---|
| AK126698 | Reduces NKD2, activates β-catenin [12] | Anti-apoptosis, resistance to cisplatin [12] | Unknown |
| CARLo-5 | Unknown | Cell cycle, proliferation, invasion, EMT [13] | ↑ in NSCLC, lymph node metastasis, poor survival [13] |
| CCAT2 | Unknown | Proliferation, migration, invasion [14] | ↑ in LAC, lymph node metastasis [14] |
| H19 | Induced by cigarette smoke [15],[16] | Unknown | ↑ in NSCLC [17], poor survival [18] |
| HOTAIR | Induced by Col-1 [19]. Affects expression of gelatinases [20]. Represses cell-adhesion genes [21], p21waf1[22], and HOXA5 [23] | Proliferation, migration invasion [20],[21],[23]; resistance to cisplatin in vitro & in vivo [22] | ↑ in NSCLC, lymph node and brain metastasis, poor survival [19],[20],[23],[24]. ↑ in cisplatin-refractory LAC [22]. ↑ in SCLC, lymphatic invasion, relapse [21] |
| LCAL1 | Unknown | Proliferation [8] | ↑ in NSCLC [8] |
| MALAT1 | Affects expression of Bcl-2 [25] and metastasis related genes [26] | EMT [27], tumor growth in vivo [26], survival [25] | ↑ in NSCLC, brain metastasis, poor survival [25],[27]. ↑in periphery blood of NSCLC [28] |
| MVIH | Affects expression of MMP-2/-9 [29] | Proliferation & invasion [29] | ↑ in LAC and LSCC, advanced TNM stage, lymph node metastasis, poor prognosis [29] |
| SCAL1 | Induced by cigarette smoke and NRF2 [30] | Protection against oxidative stress [30] | ↑ NSCLC [8],[30] |
| SOX2ot | Affects expression of EZH2 [31] | Cell cycle, proliferation [31] | ↑ in LSCC, poor survival [31] |
| ZXF1 | Antisense to ACTA2 [32] | Migration & invasion [32] | ↑ in LAC, lymph node metastasis, advanced TNM stage, poor survival [32] |
| BANCR | Inhibits the expression of EMT markers [33] | Induces apoptosis, inhibits EMT, migration, invasion, metastasis in vivo [33] | ↓ in LAC and LSCC, lymph node metastasis, advanced TNM stage, poor survival [33] |
| GAS6-AS1 | Antisense to and represses expression of GAS6 [34] | Unknown | ↓ in NSCLC, advanced TNM stage, poor survival [34] |
| MEG3 | Induces p53 [35] | Inhibits proliferation & growth in vivo, pro-apoptosis [35] | ↓ in NSCLC, advanced TNM stage, poor survival [35] |
| SPRY4-IT1 | Intronic to SPRY4, silenced by EZH2 [36] | Inhibits invasion, growth & metastasis in vivo, induces apoptosis [36] | ↓ in NSCLC, pathological stage, lymph node metastasis [36] |
| TARID | Activates TCF21 via GADD45A [37] | Unknown | ↓ in LAC and LSCC [37] |
| TUG1 | Induced by p53, represses HOXB7 via PRC2 [38] | Inhibits proliferation & growth in vivo [38] | ↓ in NSCLC, advanced TNM stage, poor survival [38] |
A summary of the lung cancer-associated lncRNAs and the molecular pathways, cell processes, and clinical features that are linked to these lncRNAs. See text for details. CARLo-5: Cancer-associated region long non-coding RNA; CCAT2: colon cancer-associated transcript 2; HOTAIR: HOX transcript antisense RNA; LCAL1: lung cancer associated lncRNA 1; MALAT1: Metastasis associated in lung adenocarcinoma transcript 1; MVIH: microvascular invasion in hepatocellular carcinoma; SCAL1: smoke and cancer-associated lncRNA-1; SOX2ot: Sox2 overlapping transcript; BANCR: BRAF activated non-coding RNA; GAS6-AS1: GAS6 antisense RNA 1; MEG3: Maternally expressed gene 3; SPRY4-IT1: SPRY4 intronic transcript 1; TARID: TCF21 antisense RNA inducing demethylation; TUG1: taurine-upregulated gene 1; NSCLC: non-small cell lung cancer; SCLC: small cell lung cancer; LAC: lung adenocarcinoma; LSCC: lung squamous cell carcinoma.
↑ and ↓ indicate increase and decrease, respectively.
The lncRNA HOX Transcript Antisense RNA (HOTAIR) has attracted intense investigation in lung cancer (Table 1) [19]-[24],[39]. Herein we review the literature of HOTAIR in lung cancer with an emphasis on the molecular mechanisms underlying its regulation of lung cancer. To obtain comprehensive insight of HOTAIR in lung cancer, we integrate mechanistic studies of HOTAIR in other types of cancer in our review.
Discovery of the HOTAIR gene
HOTAIR was discovered by Howard Chang’s group as a lncRNA that recruits Polycomb Repressive Complex 2 (PRC2), a transcriptional co-repressor, to repress the expression of the homeobox gene D cluster (HOXD) [39]. The human HOTAIR gene resides within the intergenic region between HOXC11 and HOXC12 in the HOXC cluster on chromosome 12. The HOTAIR gene is transcribed in an antisense direction relative to its flanking HOXC11 and HOXC12 genes. Its principal transcript (RefSeq NR_003716) is a 2364 bp RNA transcribed from a 6449 bp gene locus and composed of 6 exons (Figure 1, marked by an red open rectangle). An 89 bp fragment in the 5′ end of HOTAIR (221–300 bp in RefSeq NR_003716) binds to PRC2, and a 646 bp fragment in its 3′ end binds to the LSD1/CoREST/REST complex (Figure 2) [40],[41]. PRC2 contains Enhancer of Zeste Homolog 2 (EZH2), a histone methyltransferase that marks a gene for transcriptional repression via tri-methylation of histone H3 Lys27 (H3K27me3) [42]. HOTAIR appears to bind to GA-rich motifs in the genome to nucleate broad domains of PRC2 occupancy and consequent H3K27me3 [43]. The LSD1/CoREST/REST complex contains Lysine-Specific Demethylase 1 (LSD1), a histone demethylase that inactivates gene expression via demethylation of the di-methylated histone H3 Lys4 (H3K4me2), a histone modification that is critical for transcriptional activation [44]. Methylation of C1683 in HOTAIR’s principal transcript (RefSeq NR_003716) at the boundary of the LSD1-binding motif is thought to be critical for the HOTAIR-LSD1 physical interaction [45]. Acting as a bridging scaffold for PRC2 and LSD1/CoREST/REST, HOTAIR represses gene expression by coupling an increase of the repression code H3K27me3 with a decrease of the activation code tri-methylation of histone H3 lysine 4 (H3K4me3) on its target promoters [41]. In accordance, deletion of the mouse Hotair gene results in de-repression of the HOXD cluster that is coupled with decreased occupancy of H3K27me3 and increased occupancy of H3K4me3 on the HOXD gene promoters due to a loss of HOTAIR-mediated recruitment of PRC2 and LSD1 [46]. Consequently, HOTAIR null mice exhibit homeotic transformation of the spine and malformation of metacarpal-carpal bones [46].
Figure 1.
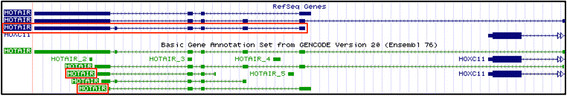
Isoforms of human HOTAIR transcripts. The USCS Genome Browser track of the human HOTAIR gene is used to illustrate isoforms of the human HOTAIR transcript [47]. The principal transcript RefSeq NR_003716 and two multi-exon HOTAIR variants in the GENCODE catalogue that lack the PRC2-interacting domain and the LSD1-interacting domain are marked by red open rectangles. See text for details.
Figure 2.
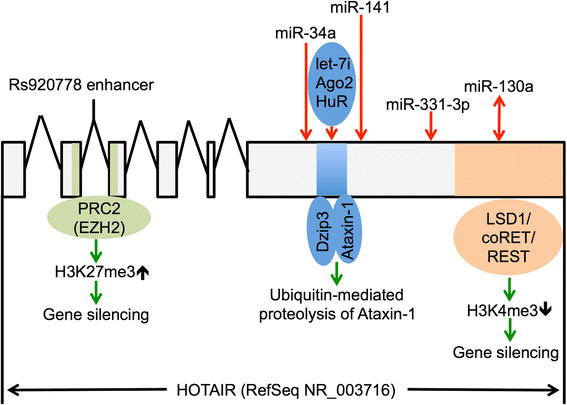
Molecular mechanisms of the tumor-promoting actions of HOTAIR. The interactions between HOTAIR and its partners are summarized. The length of each exon and positions of the interacting region for each partner are proportional to their length and positions in the principal transcript RefSeq NR_003716 of the human HOTAIR gene. The introns are not drawn proportionally to their length. A green arrow indicates positive regulation of the processes or substrates targeted by the arrow. A red arrow indicates negative regulation of the targeted processes or substrates by the arrow. A red bi-directional arrow is used to illustrate the reciprocal negative regulation between HOTAIR and miR-130a. HOTAIR’s interaction with E3 ubiquitin ligase Mex3b and its substrate Snurportin-1 is not included in the figure because the interaction is mediated through the region that overlaps with the Dzip3-Ataxin-1 interacting domain in HOTAIR. See text for details. PRC2: Polycomb Repressive Complex 2; EZH2: enhancer of zeste homolog 2; HuR: Human antigen R; LSD1: Lysine-Specific Demethylase 1.
The human HOTAIR gene can be transcribed into several variants via alternative splicing as illustrated in the GRCh38/hg38 Assembly on UCSC Genome Browser (Figure 1). The RefSeq catalogue includes three HOTAIR variants (Figure 1). The GENCODE v20 catalogue includes nine HOTAIR variants and four of them are single exon transcripts (Figure 1). A recent study using a targeted RNA capture and sequencing strategy identified six major HOTAIR splicing variants and proposed one alternative splice site, when active, can eliminate the PRC2 binding domain [48]. Consistently, two multi-exon HOTAIR variants in the GENCODE catalogue lack the PRC2-interacting domain and the LSD1-interacting domain (Figure 1, marked by red open rectangles). It is a worthy cause to determine whether alternative splicing of HOTAIR is regulated in any physiological or pathological context and whether the splicing variants exert different functions due to their different structures.
Since its first link to metastasis in breast cancer, elevated expression of HOTAIR has been reported in at least 16 types of malignancies [19],[22]-[24],[49]-[82]. Dysregulated expression of HOTAIR has not yet been reported in hematological malignancies, although the protein-coding HOX genes play a critical role in those disorders [83].
Expression of HOTAIR in lung cancer
HOTAIR exhibits significantly higher expression in the tumor tissue than the adjacent non-tumor tissue in patients with small cell lung cancer (SCLC) and non-small cell lung cancer (NSCLC) (Table 1) [19]-[24]. In SCLC, elevated expression of HOTAIR is linked to lymphatic invasion and relapse (Table 1) [21]. In NSCLC, elevated expression of HOTAIR is linked to lymph node metastasis and poor survival in patients with lung adenocarcinoma (LAC) and squamous cell carcinoma (LSCC) (Table 1) [19],[20],[22]-[24]. Moreover, elevated expression of HOTAIR is correlated with brain metastasis in NSCLC [24].
It remains unknown whether elevated expression of HOTAIR in lung cancer is caused by genetic alterations, such as amplification, deletion, or point mutations. One recent study reported that the human HOTAIR gene harbors an enhancer-like region between +1719 bp and +2353 bp downstream of its transcription start site in intron 2 [84]. The enhancer contains a risk SNP rs920778 for esophageal squamous cell carcinoma, and the rs920778T allele containing the enhancer drives higher expression of a reporter gene than the rs920778C allele (Figure 2) [84]. More importantly, the rs920778TT allele is correlated with higher expression of HOTAIR in the esophageal tissue than the rs920778CC allele, and the HOTAIR rs920778TT carriers are at a higher risk of esophageal squamous cell carcinoma than the HOTAIR rs920778CC carriers [84].
One emerging mechanism underlying up-regulation of HOTAIR in cancer cells is direct transcriptional activation of HOTAIR by classical oncogenes. For instance, HOTAIR is transcriptionally activated by the oncogene Myc through an E-box located at 1053 bp upstream of the transcription start site of the human HOTAIR gene in gallbladder cancer cells [85]. Because Myc is also a well-documented oncogene in lung cancer, this mechanism needs to be explored in lung cancer [86].
Transcriptional up-regulation of the human HOTAIR gene in cancer involves epigenetic mechanisms. An intriguing observation in breast cancer tissues is that increased DNA methylation in an intergenic CpG island located between HOXC12 and HOTAIR is positively correlated with HOTAIR expression in breast cancer [69]. It is proposed by the authors that the methylated intergenic CpG island acts as a barrier to prevent repressive heterochromatin from spreading from the HOXC12 gene into the neighboring HOTAIR gene [69]. On the other hand no CpG insland is predicted in the human HOTAIR promoter (2 kb upstream of HOTAIR’s transcription start site) using MethPrimer [87]. Besides DNA methylation histone modifications regulate the expression of HOTAIR. In breast cancer cells, estradiol activates the expression of HOTAIR via recruitment of histone methyltransferases mixed lineage leukemia proteins (MLL) to the HOTAIR promoter [50]. Consequently, MLL poises the HOTAIR promoter for transcription via H3K4me3.
Similar to protein-coding genes, lncRNAs have emerged as targets of microRNAs in a base-pairing fashion [88]. In exon 6, the HOTAIR transcript harbors a target site for miR-34a (902–923 bp in RefSeq NR_003716) (Figure 2) [54]. miR-34a reduces the expression of HOTAIR and a reporter gene that is controlled by the miR-34a target site from HOTAIR in prostate cancer cells [54]. A target site for miR-141 is identified in exon 6 of the HOTAIR transcript (1287–1308 bp in RefSeq NR_003716) (Figure 2) [53]. miR-141 reduces the expression of HOTAIR and a reporter gene that is controlled by the miR-141 target site from HOTAIR in renal carcinoma cells [53]. It is noteworthy that miR-141 is a member of the miR-200 family, one of the most potent miRNA inhibitors of epithelial-mesenchymal transition (EMT), a pathological process that is promoted by HOTAIR in cancer [49],[89]. HOTAIR is also predicted to harbor a let-7i target site in its exon 6 (2120–2141 bp in RefSeq NR_003716) although its binding to let-7i has not been experimental validated (Figure 2) [90]. Nevertheless the RNA levels of HOTAIR can be reduced by overexpression of let-7i and increased by introduction of a let-7i-specific antagomir [90]. let-7i-mediated decay of HOTAIR appears to rely on formation of a hetero-tetramer that consists of HOTAIR, let-7i, Ago2, and a RNA binding protein human antigen R (HuR). The HuR binding domain in HOTAIR is mapped to exon 6 (~1,028–1,272 bp in RefSeq NR_003716). Although it remains unclear how let-7i, Ago2, and HuR coordinate decay of HOTAIR, HuR’s binding to HOTAIR appears to recruit the let-7i/Ago2 complex to HOTAIR for decay (Figure 2) [90]. In summary, the tumor suppressive miRNA-mediated decay of HOTAIR, although established in other cancer types, warrants further investigation in lung cancer because let-7, miR-34, and miR-141 act as critical tumor suppressors in lung cancer [91]-[94].
An intriguing phenomenon observed in the seminal study of HOTAIR in breast cancer is that established breast cancer cell lines exhibit a much lower expression of HOTAIR than breast cancer tissues [60]. This apparent discrepancy might be attributed to activation of HOTAIR expression by several metastasis-promoting signals that are aberrantly enriched in the tumor microenvironment but absent in routine cell culture. For instance, transforming growth factor-β1 (TGF-β1) activates the expression of HOTAIR in breast and colon cancer cells, and such an induction is required for acquisition of EMT and cancer stem cell phenotypes [49],[95]. Prolonged exposure of human breast cancer MCF-7 cells to tumor necrosis factor-α (TNF-α) induces the expression of HOTAIR and EMT [96]-[98]. Moreover, type 1 collagen transcriptionally up-regulates the expression of HOTAIR in lung adenocarcinoma cells [19]. Interestingly, all three stimuli are potent inducers of EMT in lung cancer cells and can up-regulate expression of several tumor-promoting miRNAs, such as miR-21 and the miR-17 ~ 92 cluster [95],[96],[99]-[102].
Functions of HOTAIR in lung cancer
Elevated expression of HOTAIR is correlated with invasion, metastasis, and poor survival in patients with lung cancer (Table 1) [19]-[24]. In lung cancer cells HOTAIR regulates genes and signaling pathways that are pivotal to differentiation, proliferation, and invasion. Among the HOTAIR-regulated genes in lung cancer cells, HOXA5 is of particular interest because of its established roles in lung development and tumorigenesis [23]. HOXA5 is essential to morphogenesis of the embryonic respiratory tract and postnatal lung development [103]. Interestingly, HOXA5 is also down-regulated by another HOX cluster derived non-coding RNA, miR-196a, whose expression is inversely correlated with HOXA5 in lung cancer [104]. It is plausible that HOTAIR and miR-196a act in concert to repress the expression of HOXA5 and thereby promote dedifferentiation of lung epithelial cells during lung tumorigenesis. Another HOTAIR-repressed gene is p21WAF1/CIP1, a mediator of p53-induced growth arrest and apoptosis in response to DNA damage [22]. HOTAIR promotes proliferation, survival, and resistance to cisplatin through repression of p21WAF1/CIP1 in lung adenocarcinoma cells [22]. Thus HOTAIR can promote dedifferentiation and proliferation in lung cancer.
In addition to proliferative phenotype, HOTAIR mediates invasive phenotype of lung cancer cells through its promotion of EMT. EMT is defined as a series of events through which epithelial cells lose many of their epithelial characteristics and acquire property that is typical of mesenchymal cells, which leads to invasiveness and stemness of cancer cells [105]. During EMT, HOTAIR represses the expression of cell adhesion-related genes that are characteristic of epithelial cells in SCLC cells [21]. HOTAIR also mediates EMT via repression of EMT inhibitors. For instance, HOTAIR represses the expression of Wnt inhibitory factor 1 (WIF-1), an inhibitor of the Wnt/β-catenin pathway that mediates EMT in esophageal cancer cells [58]. In addition HOTAIR represses the expression of phosphatase and tensin homolog (PTEN), an inhibitor of EMT, in laryngeal squamous cell carcinoma cells [65]. Besides repression of EMT inhibitors, HOTAIR also mediates the expression of EMT effectors. For example, HOTAIR is required for the expression of matrix metalloproteinases that break down the extracellular matrix to pave the path for invasion in lung cancer cells [20],[59],[76],[79],[106]. Taken together, HOTAIR is induced by EMT stimuli, and such an induction in turn promotes the gene expression program that results in EMT.
The prevailing mechanism of HOTAIR-mediated regulation of cancer is that elevated expression of HOTAIR shifts PRC2-mediated gene repression from tumorigenic genes to tumor-suppressive genes [60],[61],[63],[64]. This mode of action is supported by studies on HOTAIR’s partners in lung cancer. The components of PRC2 are overexpressed in lung cancer and exert tumorigenic effects in lung cancer. EZH2 is overexpressed in SCLC and represses the expression of cell adhesion-related genes, which resembles the effects of overexpression of HOTAIR in SCLC cells [21],[107]. Another PRC2 component, SUZ12, promotes proliferation and metastasis of NSCLC cells via repression of E2F1, ROCK1, and ROBO1 [108]. Besides PRC2, HOTAIR may promote lung cancer through LSD1. LSD1 mediates proliferation and EMT in lung cancer cells, and its overexpression is associated with shorter overall survival of patients with SCLC and NSCLC [109],[110].
HOTAIR can potentially regulate lung cancer through physical interactions with E3 ubiquitin ligases and their corresponding substrates. For instance, E3 ubiquitin ligase Dzip3 and its substrate Ataxin-1 bind tandem to a ~250 nucleotide region in exon 6 (~1,028–1,272 bp in RefSeq NR_003716) through their respective RNA binding domains [90]. On the other hand, E3 ubiquitin ligase Mex3b and its substrate Snurportin-1 bind to HOTAIR in two far apart regions at ~125–250 bp and ~1,142–1,272 bp (RefSeq NR_003716), respectively [90]. Thus HOTAIR serves as an assembly scaffold that facilitates the interactions of the bound E3 ubiquitin ligases and their corresponding substrates, which leads to proteolysis of Ataxin-1 and Snurportin-1 [90]. Intriguingly, Ataxin-1, Snurportin-1, and HuR appear to compete for the same region in HOTAIR (~1,028–1,272 bp in RefSeq NR_003716) that mediates decay of HOTAIR upon HuR binding (Figure 2) [90]. It is plausible that HuR-mediated decay of HOTAIR and HOTAIR-mediated ubiquitination of Ataxin-1 and Snurportin-1 are mutually exclusive because of their competition for the same region in HOTAIR. The intertwining of HOTAIR decay and proteolysis may play a role in cell senescence. Induction of HOTAIR in senescent cells prevents premature senescence via interaction with Dzip3 and Mex3b and the consequent rapid proteolysis of Ataxin-1 and Snurportin-1 [90]. HOTAIR-mediated regulation of senescence is potentially important in lung cancer because evasion of senescence is proposed as a critical step in lung tumorigenesis [111]. Moreover, HOTAIR-mediated ubiquitination and degradation of Ataxin-1 is of particular interest to lung cancer because Ataxin-1 is essential to lung alveolization [112]. Thus HOTAIR may promote dedifferentiation of lung epithelial cells through two distinct mechanisms, i.e., transcriptional repression of HOXA5 and ubiquitin-mediated proteolysis of Ataxin-1 [23],[90].
An emerging theme in the non-coding RNA world is the crosstalk between miRNAs and lncRNAs [88]. As discussed above, the expression of HOTAIR is regulated by several tumor suppressive miRNAs, such as miR-34a and miR-141 in cancer cells (Figure 2) [53],[54]. On the other hand, HOTAIR antagonizes several tumor suppressive miRNAs. In gastric cancer cells, HOTAIR acts as a competitive endogenous RNA (ceRNA) to trap miR-331-3p through a complementary target site (1451–1471 bp in RefSeq NR_003716) and thereby increases the expression of the miR-331-3p-targeted oncogene HER2 (Figure 2) [68]. In gall bladder cancer, HOTAIR’s oncogenic activity requires its binding to and neutralization of miR-130a (1805–1826 bp in RefSeq NR_003716) (Figure 2) [85]. Reciprocally, miR-130a represses the expression of HOTAIR in a target site-dependent manner (Figure 2) [85]. Despite its discovery in other types of cancer, a crosstalk between HOTAIR and miRNAs is worth exploring in lung cancer because miR-331 and miR-130a are tumor suppressors in lung cancer [113],[114].
Clinical potentials of HOTAIR in lung cancer
HOTAIR can be explored as a biomarker in lung cancer because its elevated expression in lung tumor tissues is correlated with metastasis, drug resistance, and poor survival in patients with lung cancer (Table 1). For instance, in a cohort of 42 patients with NSCLC, 5-year post-operative survival in 21 patients with high expression of HOTAIR is at only 20% vs a 45% survival rate in 21 patients with low expression of HOTAIR [23]. In another cohort of 35 patients with SCLC average disease-free survival is at 30.8 months in 12 patients with high expression of HOTAIR vs average survival of 46.3 months in 23 patients with low expression of HOTAIR [21].
HOTAIR’s feasibility as a biomarker is enhanced by the findings that lncRNAs are stable and measurable in body fluids and thereby suitable for measurement via non-invasive procedures [7]. HOTAIR along with several other lncRNAs can be quantitatively measured in plasma samples collected from patients with gastric cancer [115]. HOTAIR’s power as a biomarker is further enhanced when it is measured in combination with other critical regulators of lung cancer. A combined measurement of exosomal miR-21 and HOTAIR yields greater sensitivity and specificity in distinguishing laryngeal squamous cell carcinoma from benign polyps than each individual measurement alone [77]. This approach can be readily applied to lung cancer because miR-21 is a miRNA signature of NSCLC and co-upregulated by Col-1 in lung cancer cells [19],[99],[116]. Another approach to increase the predictive power of HOTAIR is simultaneous measurement of HOTAIR and its protein partners, e.g., EZH2. As exemplified in a breast cancer study simultaneous increase of HOTAIR and PRC2 has a greater correlation with poor survival than the increase of each marker alone [52].
HOTAIR is an appealing therapeutic target because inhibition of HOTAIR exhibits promising anti-tumor efficacy in preclinical models of lung cancer (Table 1). Moreover, pharmacological inhibitors of PRC2 exhibit convincing anti-tumor efficacy in preclinical models of NSCLC and SCLC [117],[118]. It is important to specifically disrupt the interaction between HOTAIR and PRC2 in cancer cells upon successful molecular and biochemical resolution of the interaction between HOTAIR and PRC2. This approach can potentially spare any HOTAIR-independent physiological functions of PRC2. Another appeal of HOTAIR as a therapeutic target arises from its critical role in resistance to chemotherapy drugs in lung cancer cells [22]. A combination of traditional chemotherapy and inhibition of HOTAIR can potentially overcome drug resistance and increase tolerance to traditional chemotherapy.
Challenges and future directions
HOTAIR has emerged as a promising diagnostic and therapeutic target for lung cancer (Table 1). However, several challenges hinder realization of HOTAIR’s potential in intervention of lung cancer. One challenge is our limited understanding of the interaction between HOTAIR and its protein partners [41]. A high-resolution map of HOTAIR-PRC2 and HOTAIR-LSD1 interactions is essential to develop compounds that can effectively and specifically disrupt their interaction in lung cancer cells. This is highlighted by the fact that PRC2 physically interacts with thousands of lncRNAs, and its function is tightly regulated by these interacting lncRNAs [119],[120]. It is conceivable that PRC2 forms a pool of functional units as defined by their lncRNA partners, and this pool of PRC2-lncRNA units is dynamically fine-tuned to maintain an appropriate gene expression program to meet the cell’s needs in a particular cellular context. How an increased expression of HOTAIR disturbs this fine-tuned pool of PRC2-lncRNA units and promotes cancer is a daunting question to answer. One can speculate that increased HOTAIR binding to PRC2 can interfere with formation of other PRC2-lncRNA units through competitive binding or alteration of PRC2 conformation. This is critical to lung cancer because TUG1, also a PRC2-interacting lncRNA, exerts its tumor suppressive action through PRC2-mediated repression of HOXB7 [38].
Another challenge arises from EZH2-mediated methylation of non-histone proteins. Undoubtedly, inhibition of either HOTAIR or EZH2 hinders progression of lung cancer (Table 1) [117],[118]. However, the experimental designs in these studies are not able to exclude the possibility that the altered gene expression and cell behaviors can be, at least in part, attributed to altered methylation of transcription factors and other non-histone proteins methylated by EZH2. For instance, EZH2 directly methylates transcription factor GATA4 and diminishes GATA4’s transcriptional activity [121],[122]. HOTAIR-regulated EZH2-dependent methylation of non-histone substrates in lung cancer cells can be explored using a proteomic survey of the methylated proteins with or without inhibition of HOTAIR and/or EZH2.
It is also naive to conclude that PRC2, LSD1, and the newly discovered E3 ubiquitin ligases are the sole protein partners of HOTAIR to mediate its functions in lung cancer [41],[90]. lncRNAs reside in every subcellular compartment and act in a wide range of cell processes, such as signaling transduction, RNA splicing, and ligand-receptor engagement [4]. The versatility of HOTAIR function in lung cancer needs to be explored with a thorough screening of HOTAIR-bound protein partners using HOTAIR as bait in lung cancer cells.
Summary
HOTAIR has emerged as a novel master regulator of lung cancer. HOTAIR possesses tremendous diagnostic and therapeutic potentials in intervention of lung cancer. Materialization of HOTAIR’s clinical potential requires further investigation of the molecular mechanisms underlying the tumor-promoting actions of HOTAIR in lung cancer.
Acknowledgements
This work is supported by Washington State University Startup Fund awarded to BS and NIH R01GM097571 awarded to JJ.
Abbreviations
- lncRNA
Long non-coding RNA
- HOTAIR
HOX transcript antisense RNA
- PRC2
Polycomb repressive complex 2
- EZH2
Enhancer of zeste homolog 2
- LSD1
Lysine-specific demethylase 1
- PTEN
Phosphatase and tensin homolog
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
{kind=link}
{kind=link}
Footnotes
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
All authors have contributed to the preparation of this manuscript. All authors have read and approved the manuscript.
Contributor Information
Gregory Loewen, Email: gregory.loewen@gmail.com.
Janarthanan Jayawickramarajah, Email: jananj@tulane.edu.
Ying Zhuo, Email: zhuoying1@yahoo.com.
Bin Shan, Email: bin.shan@wsu.edu.
References
- 1.Djebali S, Davis CA, Merkel A, Dobin A, Lassmann T, Mortazavi A, Tanzer A, Lagarde J, Lin W, Schlesinger F, Xue C, Marinov GK, Khatun J, Williams BA, Zaleski C, Rozowsky J, Roder M, Kokocinski F, Abdelhamid RF, Alioto T, Antoshechkin I, Baer MT, Bar NS, Batut P, Bell K, Bell I, Chakrabortty S, Chen X, Chrast J, Curado J, et al. Landscape of transcription in human cells. Nature. 2012;489(7414):101–108. doi: 10.1038/nature11233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Rinn JL, Chang HY. Genome regulation by long noncoding RNAs. Annu Rev Biochem. 2012;81:145–166. doi: 10.1146/annurev-biochem-051410-092902. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Derrien T, Johnson R, Bussotti G, Tanzer A, Djebali S, Tilgner H, Guernec G, Martin D, Merkel A, Knowles DG, Lagarde J, Veeravalli L, Ruan X, Ruan Y, Lassmann T, Carninci P, Brown JB, Lipovich L, Gonzalez JM, Thomas M, Davis CA, Shiekhattar R, Gingeras TR, Hubbard TJ, Notredame C, Harrow J, Guigo R. The GENCODE v7 catalog of human long noncoding RNAs: analysis of their gene structure, evolution, and expression. Genome Res. 2012;22(9):1775–1789. doi: 10.1101/gr.132159.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Wang KC, Chang HY. Molecular mechanisms of long noncoding RNAs. Mol Cell. 2011;43(6):904–914. doi: 10.1016/j.molcel.2011.08.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Booton R, Lindsay MA. Emerging role of MicroRNAs and long noncoding RNAs in respiratory disease. Chest. 2014;146(1):193–204. doi: 10.1378/chest.13-2736. [DOI] [PubMed] [Google Scholar]
- 6.Zhang H, Chen Z, Wang X, Huang Z, He Z, Chen Y. Long non-coding RNA: a new player in cancer. J Hematol Oncol. 2013;6:37. doi: 10.1186/1756-8722-6-37. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Prensner JR, Chinnaiyan AM. The emergence of lncRNAs in cancer biology. Canc Discov. 2011;1(5):391–407. doi: 10.1158/2159-8290.CD-11-0209. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.White NM, Cabanski CR, Silva-Fisher JM, Dang HX, Govindan R, Maher CA. Transcriptome sequencing reveals altered long intergenic non-coding RNAs in lung cancer. Genome Biol. 2014;15(8):429. doi: 10.1186/s13059-014-0429-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Yang J, Lin J, Liu T, Chen T, Pan S, Huang W, Li S. Analysis of lncRNA expression profiles in non-small cell lung cancers (NSCLC) and their clinical subtypes. Lung Cancer. 2014;85(2):110–115. doi: 10.1016/j.lungcan.2014.05.011. [DOI] [PubMed] [Google Scholar]
- 10.Xu G, Chen J, Pan Q, Huang K, Pan J, Zhang W, Chen J, Yu F, Zhou T, Wang Y. Long noncoding RNA expression profiles of lung adenocarcinoma ascertained by microarray analysis. PLoS One. 2014;9(8):e104044. doi: 10.1371/journal.pone.0104044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Wang Y, Chen W, Chen J, Pan Q, Pan J. LncRNA expression profiles of EGFR exon 19 deletions in lung adenocarcinoma ascertained by using microarray analysis. Med Oncol. 2014;31(9):137. doi: 10.1007/s12032-014-0137-y. [DOI] [PubMed] [Google Scholar]
- 12.Yang Y, Li H, Hou S, Hu B, Liu J, Wang J. The noncoding RNA expression profile and the effect of lncRNA AK126698 on cisplatin resistance in non-small-cell lung cancer cell. PLoS One. 2013;8(5):e65309. doi: 10.1371/journal.pone.0065309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Luo J, Tang L, Zhang J, Ni J, Zhang HP, Zhang L, Xu JF, Zheng D. Long non-coding RNA CARLo-5 is a negative prognostic factor and exhibits tumor pro-oncogenic activity in non-small cell lung cancer. Tumour Biol. 2014;35(11):11541–11549. doi: 10.1007/s13277-014-2442-7. [DOI] [PubMed] [Google Scholar]
- 14.Qiu M, Xu Y, Yang X, Wang J, Hu J, Xu L, Yin R. CCAT2 is a lung adenocarcinoma-specific long non-coding RNA and promotes invasion of non-small cell lung cancer. Tumour Biol. 2014;35(6):5375–5380. doi: 10.1007/s13277-014-1700-z. [DOI] [PubMed] [Google Scholar]
- 15.Liu F, Killian JK, Yang M, Walker RL, Hong JA, Zhang M, Davis S, Zhang Y, Hussain M, Xi S, Rao M, Meltzer PA, Schrump DS. Epigenomic alterations and gene expression profiles in respiratory epithelia exposed to cigarette smoke condensate. Oncogene. 2010;29(25):3650–3664. doi: 10.1038/onc.2010.129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Kaplan R, Luettich K, Heguy A, Hackett NR, Harvey BG, Crystal RG. Monoallelic up-regulation of the imprinted H19 gene in airway epithelium of phenotypically normal cigarette smokers. Cancer Res. 2003;63(7):1475–1482. [PubMed] [Google Scholar]
- 17.Kondo M, Suzuki H, Ueda R, Osada H, Takagi K, Takahashi T, Takahashi T. Frequent loss of imprinting of the H19 gene is often associated with its overexpression in human lung cancers. Oncogene. 1995;10(6):1193–1198. [PubMed] [Google Scholar]
- 18.Chen B, Yu M, Chang Q, Lu Y, Thakur C, Ma D, Yi Z, Chen F. Mdig de-represses H19 large intergenic non-coding RNA (lincRNA) by down-regulating H3K9me3 and heterochromatin. Oncotarget. 2013;4(9):1427–1437. doi: 10.18632/oncotarget.1155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Zhuang Y, Wang X, Nguyen HT, Zhuo Y, Cui X, Fewell C, Flemington EK, Shan B. Induction of long intergenic non-coding RNA HOTAIR in lung cancer cells by type I collagen. J Hematol Oncol. 2013;6:35. doi: 10.1186/1756-8722-6-35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zhao W, An Y, Liang Y, Xie XW. Role of HOTAIR long noncoding RNA in metastatic progression of lung cancer. Eur Rev Med Pharmacol Sci. 2014;18(13):1930–1936. [PubMed] [Google Scholar]
- 21.Ono H, Motoi N, Nagano H, Miyauchi E, Ushijima M, Matsuura M, Okumura S, Nishio M, Hirose T, Inase N, Ishikawa Y. Long noncoding RNA HOTAIR is relevant to cellular proliferation, invasiveness, and clinical relapse in small-cell lung cancer. Cancer Med. 2014;3(3):632–642. doi: 10.1002/cam4.220. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Liu Z, Sun M, Lu K, Liu J, Zhang M, Wu W, De W, Wang Z, Wang R. The long noncoding RNA HOTAIR contributes to cisplatin resistance of human lung adenocarcinoma cells via downregualtion of p21(WAF1/CIP1) expression. PLoS One. 2013;8(10):e77293. doi: 10.1371/journal.pone.0077293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Liu XH, Liu ZL, Sun M, Liu J, Wang ZX, De W. The long non-coding RNA HOTAIR indicates a poor prognosis and promotes metastasis in non-small cell lung cancer. BMC Cancer. 2013;13:464. doi: 10.1186/1471-2407-13-464. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Nakagawa T, Endo H, Yokoyama M, Abe J, Tamai K, Tanaka N, Sato I, Takahashi S, Kondo T, Satoh K. Large noncoding RNA HOTAIR enhances aggressive biological behavior and is associated with short disease-free survival in human non-small cell lung cancer. Biochem Biophys Res Commun. 2013;436(2):319–324. doi: 10.1016/j.bbrc.2013.05.101. [DOI] [PubMed] [Google Scholar]
- 25.Schmidt LH, Gorlich D, Spieker T, Rohde C, Schuler M, Mohr M, Humberg J, Sauer T, Thoenissen NH, Huge A, Voss R, Marra A, Faldum A, Muller-Tidow C, Berdel WE, Wiewrodt R. Prognostic impact of Bcl-2 depends on tumor histology and expression of MALAT-1 lncRNA in Non-small-cell lung cancer. J Thorac Oncol. 2014;9(9):1294–1304. doi: 10.1097/JTO.0000000000000243. [DOI] [PubMed] [Google Scholar]
- 26.Gutschner T, Hammerle M, Eissmann M, Hsu J, Kim Y, Hung G, Revenko A, Arun G, Stentrup M, Gross M, Zornig M, MacLeod AR, Spector DL, Diederichs S. The noncoding RNA MALAT1 is a critical regulator of the metastasis phenotype of lung cancer cells. Cancer Res. 2013;73(3):1180–1189. doi: 10.1158/0008-5472.CAN-12-2850. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Shen L, Chen L, Wang Y, Jiang X, Xia H, Zhuang Z: Long noncoding RNA MALAT1 promotes brain metastasis by inducing epithelial-mesenchymal transition in lung cancer.J Neuro-Oncol 2014. [Epub ahead of print], [DOI] [PubMed]
- 28.Weber DG, Johnen G, Casjens S, Bryk O, Pesch B, Jockel KH, Kollmeier J, Bruning T. Evaluation of long noncoding RNA MALAT1 as a candidate blood-based biomarker for the diagnosis of non-small cell lung cancer. BMC Res Notes. 2013;6:518. doi: 10.1186/1756-0500-6-518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Nie FQ, Zhu Q, Xu TP, Zou YF, Xie M, Sun M, Xia R, Lu KH. Long non-coding RNA MVIH indicates a poor prognosis for non-small cell lung cancer and promotes cell proliferation and invasion. Tumour Biol. 2014;35(8):7587–7594. doi: 10.1007/s13277-014-2009-7. [DOI] [PubMed] [Google Scholar]
- 30.Thai P, Statt S, Chen CH, Liang E, Campbell C, Wu R. Characterization of a novel long noncoding RNA, SCAL1, induced by cigarette smoke and elevated in lung cancer cell lines. Am J Respir Cell Mol Biol. 2013;49(2):204–211. doi: 10.1165/rcmb.2013-0159RC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hou Z, Zhao W, Zhou J, Shen L, Zhan P, Xu C, Chang C, Bi H, Zou J, Yao X, Huang R, Yu L, Yan J. A long noncoding RNA Sox2ot regulates lung cancer cell proliferation and is a prognostic indicator of poor survival. Int J Biochem Cell Biol. 2014;53:380–388. doi: 10.1016/j.biocel.2014.06.004. [DOI] [PubMed] [Google Scholar]
- 32.Zhang L, Zhou XF, Pan GF, Zhao JP. Enhanced expression of long non-coding RNA ZXF1 promoted the invasion and metastasis in lung adenocarcinoma. Biomed Pharmacother. 2014;68(4):401–407. doi: 10.1016/j.biopha.2014.03.001. [DOI] [PubMed] [Google Scholar]
- 33.Sun M, Liu XH, Wang KM, Nie FQ, Kong R, Yang JS, Xia R, Xu TP, Jin FY, Liu ZJ, Chen JF, Zhang EB, De W, Wang ZX. Downregulation of BRAF activated non-coding RNA is associated with poor prognosis for non-small cell lung cancer and promotes metastasis by affecting epithelial-mesenchymal transition. Mol Cancer. 2014;13:68. doi: 10.1186/1476-4598-13-68. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Han L, Kong R, Yin DD, Zhang EB, Xu TP, De W, Shu YQ. Low expression of long noncoding RNA GAS6-AS1 predicts a poor prognosis in patients with NSCLC. Med Oncol. 2013;30(4):694. doi: 10.1007/s12032-013-0694-5. [DOI] [PubMed] [Google Scholar]
- 35.Lu KH, Li W, Liu XH, Sun M, Zhang ML, Wu WQ, Xie WP, Hou YY. Long non-coding RNA MEG3 inhibits NSCLC cells proliferation and induces apoptosis by affecting p53 expression. BMC Cancer. 2013;13:461. doi: 10.1186/1471-2407-13-461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Sun M, Liu XH, Lu KH, Nie FQ, Xia R, Kong R, Yang JS, Xu TP, Liu YW, Zou YF, Lu BB, Yin R, Zhang EB, Xu L, De W, Wang ZX. EZH2-mediated epigenetic suppression of long noncoding RNA SPRY4-IT1 promotes NSCLC cell proliferation and metastasis by affecting the epithelial-mesenchymal transition. Cell Death Dis. 2014;5:e1298. doi: 10.1038/cddis.2014.256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Arab K, Park YJ, Lindroth AM, Schafer A, Oakes C, Weichenhan D, Lukanova A, Lundin E, Risch A, Meister M, Dienemann H, Dyckhoff G, Herold-Mende C, Grummt I, Niehrs C, Plass C. Long noncoding RNA TARID directs demethylation and activation of the tumor suppressor TCF21 via GADD45A. Mol Cell. 2014;55(4):604–614. doi: 10.1016/j.molcel.2014.06.031. [DOI] [PubMed] [Google Scholar]
- 38.Zhang EB, Yin DD, Sun M, Kong R, Liu XH, You LH, Han L, Xia R, Wang KM, Yang JS, De W, Shu YQ, Wang ZX. P53-regulated long non-coding RNA TUG1 affects cell proliferation in human non-small cell lung cancer, partly through epigenetically regulating HOXB7 expression. Cell Death Dis. 2014;5:e1243. doi: 10.1038/cddis.2014.201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Rinn JL, Kertesz M, Wang JK, Squazzo SL, Xu X, Brugmann SA, Goodnough LH, Helms JA, Farnham PJ, Segal E, Chang HY. Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs. Cell. 2007;129(7):1311–1323. doi: 10.1016/j.cell.2007.05.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Wu L, Murat P, Matak-Vinkovic D, Murrell A, Balasubramanian S. Binding interactions between long noncoding RNA HOTAIR and PRC2 proteins. Biochemistry. 2013;52(52):9519–9527. doi: 10.1021/bi401085h. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Tsai MC, Manor O, Wan Y, Mosammaparast N, Wang JK, Lan F, Shi Y, Segal E, Chang HY. Long noncoding RNA as modular scaffold of histone modification complexes. Science. 2010;329(5992):689–693. doi: 10.1126/science.1192002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kirmizis A, Bartley SM, Kuzmichev A, Margueron R, Reinberg D, Green R, Farnham PJ. Silencing of human polycomb target genes is associated with methylation of histone H3 Lys 27. Genes Dev. 2004;18(13):1592–1605. doi: 10.1101/gad.1200204. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Chu C, Qu K, Zhong FL, Artandi SE, Chang HY. Genomic maps of long noncoding RNA occupancy reveal principles of RNA-chromatin interactions. Mol Cell. 2011;44(4):667–678. doi: 10.1016/j.molcel.2011.08.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Shi Y, Lan F, Matson C, Mulligan P, Whetstine JR, Cole PA, Casero RA, Shi Y. Histone demethylation mediated by the nuclear amine oxidase homolog LSD1. Cell. 2004;119(7):941–953. doi: 10.1016/j.cell.2004.12.012. [DOI] [PubMed] [Google Scholar]
- 45.Amort T, Souliere MF, Wille A, Jia XY, Fiegl H, Worle H, Micura R, Lusser A. Long non-coding RNAs as targets for cytosine methylation. RNA Biol. 2013;10(6):1003–1008. doi: 10.4161/rna.24454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Li L, Liu B, Wapinski OL, Tsai MC, Qu K, Zhang J, Carlson JC, Lin M, Fang F, Gupta RA, Helms JA, Chang HY. Targeted disruption of Hotair leads to homeotic transformation and gene derepression. Cell Rep. 2013;5(1):3–12. doi: 10.1016/j.celrep.2013.09.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Kent WJ, Sugnet CW, Furey TS, Roskin KM, Pringle TH, Zahler AM, Haussler D. The human genome browser at UCSC. Genome Res. 2002;12(6):996–1006. doi: 10.1101/gr.229102.ArticlepublishedonlinebeforeprintinMay2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Mercer TR, Gerhardt DJ, Dinger ME, Crawford J, Trapnell C, Jeddeloh JA, Mattick JS, Rinn JL. Targeted RNA sequencing reveals the deep complexity of the human transcriptome. Nat Biotechnol. 2012;30(1):99–104. doi: 10.1038/nbt.2024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Padua Alves C, Fonseca AS, Muys BR, de Barros ELBR, Burger MC, de Souza JE, Valente V, Zago MA, Silva WA., Jr Brief report: the lincRNA hotair is required for epithelial-to-mesenchymal transition and stemness maintenance of cancer cell lines. Stem Cells. 2013;31(12):2827–2832. doi: 10.1002/stem.1547. [DOI] [PubMed] [Google Scholar]
- 50.Bhan A, Hussain I, Ansari KI, Kasiri S, Bashyal A, Mandal SS. Antisense transcript long noncoding RNA (lncRNA) HOTAIR is transcriptionally induced by estradiol. J Mol Biol. 2013;425(19):3707–3722. doi: 10.1016/j.jmb.2013.01.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Chen FJ, Sun M, Li SQ, Wu QQ, Ji L, Liu ZL, Zhou GZ, Cao G, Jin L, Xie HW, Wang CM, Lv J, De W, Wu M, Cao XF. Upregulation of the long non-coding RNA HOTAIR promotes esophageal squamous cell carcinoma metastasis and poor prognosis. Mol Carcinog. 2013;52(11):908–915. doi: 10.1002/mc.21944. [DOI] [PubMed] [Google Scholar]
- 52.Chisholm KM, Wan Y, Li R, Montgomery KD, Chang HY, West RB. Detection of long non-coding RNA in archival tissue: correlation with polycomb protein expression in primary and metastatic breast carcinoma. PLoS One. 2012;7(10):e47998. doi: 10.1371/journal.pone.0047998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Chiyomaru T, Fukuhara S, Saini S, Majid S, Deng G, Shahryari V, Chang I, Tanaka Y, Enokida H, Nakagawa M, Dahiya R, Yamamura S. Long non-coding RNA HOTAIR is targeted and regulated by miR-141 in human cancer cells. J Biol Chem. 2014;289(18):12550–12565. doi: 10.1074/jbc.M113.488593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Chiyomaru T, Yamamura S, Fukuhara S, Yoshino H, Kinoshita T, Majid S, Saini S, Chang I, Tanaka Y, Enokida H, Seki N, Nakagawa M, Dahiya R. Genistein inhibits prostate cancer cell growth by targeting miR-34a and oncogenic HOTAIR. PLoS One. 2013;8(8):e70372. doi: 10.1371/journal.pone.0070372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Cuenda A, Rouse J, Doza YN, Meier R, Cohen P, Gallagher TF, Young PR, Lee JC. SB 203580 is a specific inhibitor of a MAP kinase homologue which is stimulated by cellular stresses and interleukin-1. FEBS Lett. 1995;364(2):229–233. doi: 10.1016/0014-5793(95)00357-F. [DOI] [PubMed] [Google Scholar]
- 56.Cui L, Xie XY, Wang H, Chen XL, Liu SL, Hu LN. [Expression of long non-coding RNA HOTAIR mRNA in ovarian cancer] Sichuan Da Xue Xue Bao Yi Xue Ban. 2013;44(1):57–59. [PubMed] [Google Scholar]
- 57.Endo H, Shiroki T, Nakagawa T, Yokoyama M, Tamai K, Yamanami H, Fujiya T, Sato I, Yamaguchi K, Tanaka N, Iijima K, Shimosegawa T, Sugamura K, Satoh K. Enhanced expression of long non-coding RNA HOTAIR is associated with the development of gastric cancer. PLoS One. 2013;8(10):e77070. doi: 10.1371/journal.pone.0077070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Ge XS, Ma HJ, Zheng XH, Ruan HL, Liao XY, Xue WQ, Chen YB, Zhang Y, Jia WH. HOTAIR, a prognostic factor in esophageal squamous cell carcinoma, inhibits WIF-1 expression and activates Wnt pathway. Cancer Sci. 2013;104(12):1675–1682. doi: 10.1111/cas.12296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Geng YJ, Xie SL, Li Q, Ma J, Wang GY. Large intervening non-coding RNA HOTAIR is associated with hepatocellular carcinoma progression. J Int Med Res. 2011;39(6):2119–2128. doi: 10.1177/147323001103900608. [DOI] [PubMed] [Google Scholar]
- 60.Gupta RA, Shah N, Wang KC, Kim J, Horlings HM, Wong DJ, Tsai MC, Hung T, Argani P, Rinn JL, Wang Y, Brzoska P, Kong B, Li R, West RB, van de Vijver MJ, Sukumar S, Chang HY. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature. 2010;464(7291):1071–1076. doi: 10.1038/nature08975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Hajjari M, Behmanesh M, Sadeghizadeh M, Zeinoddini M. Up-regulation of HOTAIR long non-coding RNA in human gastric adenocarcinoma tissues. Med Oncol. 2013;30(3):670. doi: 10.1007/s12032-013-0670-0. [DOI] [PubMed] [Google Scholar]
- 62.Ishibashi M, Kogo R, Shibata K, Sawada G, Takahashi Y, Kurashige J, Akiyoshi S, Sasaki S, Iwaya T, Sudo T, Sugimachi K, Mimori K, Wakabayashi G, Mori M. Clinical significance of the expression of long non-coding RNA HOTAIR in primary hepatocellular carcinoma. Oncol Rep. 2013;29(3):946–950. doi: 10.3892/or.2012.2219. [DOI] [PubMed] [Google Scholar]
- 63.Kim K, Jutooru I, Chadalapaka G, Johnson G, Frank J, Burghardt R, Kim S, Safe S. HOTAIR is a negative prognostic factor and exhibits pro-oncogenic activity in pancreatic cancer. Oncogene. 2013;32(13):1616–1625. doi: 10.1038/onc.2012.193. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Kogo R, Shimamura T, Mimori K, Kawahara K, Imoto S, Sudo T, Tanaka F, Shibata K, Suzuki A, Komune S, Miyano S, Mori M. Long noncoding RNA HOTAIR regulates polycomb-dependent chromatin modification and is associated with poor prognosis in colorectal cancers. Cancer Res. 2011;71(20):6320–6326. doi: 10.1158/0008-5472.CAN-11-1021. [DOI] [PubMed] [Google Scholar]
- 65.Li D, Feng J, Wu T, Wang Y, Sun Y, Ren J, Liu M. Long intergenic noncoding RNA HOTAIR is overexpressed and regulates PTEN methylation in laryngeal squamous cell carcinoma. Am J Pathol. 2013;182(1):64–70. doi: 10.1016/j.ajpath.2012.08.042. [DOI] [PubMed] [Google Scholar]
- 66.Li G, Zhang H, Wan X, Yang X, Zhu C, Wang A, He L, Miao R, Chen S, Zhao H. Long noncoding RNA plays a key role in metastasis and prognosis of hepatocellular carcinoma. Biomed Res Int. 2014;2014:780521. doi: 10.1155/2014/780521. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Li Z, Li C, Liu C, Yu S, Zhang Y: Expression of the long non-coding RNAs MEG3, HOTAIR, and MALAT-1 in non-functioning pituitary adenomas and their relationship to tumor behavior.Pituitary 2014. [Epub ahead of print], [DOI] [PubMed]
- 68.Liu XH, Sun M, Nie FQ, Ge YB, Zhang EB, Yin DD, Kong R, Xia R, Lu KH, Li JH, De W, Wang KM, Wang ZX. Lnc RNA HOTAIR functions as a competing endogenous RNA to regulate HER2 expression by sponging miR-331-3p in gastric cancer. Mol Cancer. 2014;13:92. doi: 10.1186/1476-4598-13-92. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Lu L, Zhu G, Zhang C, Deng Q, Katsaros D, Mayne ST, Risch HA, Mu L, Canuto EM, Gregori G, Benedetto C, Yu H. Association of large noncoding RNA HOTAIR expression and its downstream intergenic CpG island methylation with survival in breast cancer. Breast Cancer Res Treat. 2012;136(3):875–883. doi: 10.1007/s10549-012-2314-z. [DOI] [PubMed] [Google Scholar]
- 70.Lv XB, Lian GY, Wang HR, Song E, Yao H, Wang MH. Long noncoding RNA HOTAIR is a prognostic marker for esophageal squamous cell carcinoma progression and survival. PLoS One. 2013;8(5):e63516. doi: 10.1371/journal.pone.0063516. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Milhem MM, Knutson T, Yang S, Zhu D, Wang X, Leslie KK, Meng X. Correlation of MTDH/AEG-1 and HOTAIR expression with metastasis and response to treatment in sarcoma patients. J Canc Sci Ther. 2011;S5(4):pii:004. [PMC free article] [PubMed] [Google Scholar]
- 72.Nie Y, Liu X, Qu S, Song E, Zou H, Gong C. Long non-coding RNA HOTAIR is an independent prognostic marker for nasopharyngeal carcinoma progression and survival. Cancer Sci. 2013;104(4):458–464. doi: 10.1111/cas.12092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Niinuma T, Suzuki H, Nojima M, Nosho K, Yamamoto H, Takamaru H, Yamamoto E, Maruyama R, Nobuoka T, Miyazaki Y, Nishida T, Bamba T, Kanda T, Ajioka Y, Taguchi T, Okahara S, Takahashi H, Nishida Y, Hosokawa M, Hasegawa T, Tokino T, Hirata K, Imai K, Toyota M, Shinomura Y. Upregulation of miR-196a and HOTAIR drive malignant character in gastrointestinal stromal tumors. Cancer Res. 2012;72(5):1126–1136. doi: 10.1158/0008-5472.CAN-11-1803. [DOI] [PubMed] [Google Scholar]
- 74.Ozgur E, Mert U, Isin M, Okutan M, Dalay N, Gezer U. Differential expression of long non-coding RNAs during genotoxic stress-induced apoptosis in HeLa and MCF-7 cells. Clin Exp Med. 2013;13(2):119–126. doi: 10.1007/s10238-012-0181-x. [DOI] [PubMed] [Google Scholar]
- 75.Schiavo G, D’Anto V, Cantile M, Procino A, Di Giovanni S, Valletta R, Terracciano L, Baumhoer D, Jundt G, Cillo C. Deregulated HOX genes in ameloblastomas are located in physical contiguity to keratin genes. J Cell Biochem. 2011;112(11):3206–3215. doi: 10.1002/jcb.23248. [DOI] [PubMed] [Google Scholar]
- 76.Tang L, Zhang W, Su B, Yu B. Long noncoding RNA HOTAIR is associated with motility, invasion, and metastatic potential of metastatic melanoma. Biomed Res Int. 2013;2013:251098. doi: 10.1155/2013/251098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Wang J, Zhou Y, Lu J, Sun Y, Xiao H, Liu M, Tian L. Combined detection of serum exosomal miR-21 and HOTAIR as diagnostic and prognostic biomarkers for laryngeal squamous cell carcinoma. Med Oncol. 2014;31(9):148. doi: 10.1007/s12032-014-0148-8. [DOI] [PubMed] [Google Scholar]
- 78.Wang L, Zeng X, Chen S, Ding L, Zhong J, Zhao JC, Wang L, Sarver A, Koller A, Zhi J, Ma Y, Yu J, Chen J, Huang H. BRCA1 is a negative modulator of the PRC2 complex. EMBO J. 2013;32(11):1584–1597. doi: 10.1038/emboj.2013.95. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Xu ZY, Yu QM, Du YA, Yang LT, Dong RZ, Huang L, Yu PF, Cheng XD. Knockdown of long non-coding RNA HOTAIR suppresses tumor invasion and reverses epithelial-mesenchymal transition in gastric cancer. Int J Biol Sci. 2013;9(6):587–597. doi: 10.7150/ijbs.6339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Yang Z, Zhou L, Wu LM, Lai MC, Xie HY, Zhang F, Zheng SS. Overexpression of long non-coding RNA HOTAIR predicts tumor recurrence in hepatocellular carcinoma patients following liver transplantation. Ann Surg Oncol. 2011;18(5):1243–1250. doi: 10.1245/s10434-011-1581-y. [DOI] [PubMed] [Google Scholar]
- 81.Zhang JX, Han L, Bao ZS, Wang YY, Chen LY, Yan W, Yu SZ, Pu PY, Liu N, You YP, Jiang T, Kang CS. HOTAIR, a cell cycle-associated long noncoding RNA and a strong predictor of survival, is preferentially expressed in classical and mesenchymal glioma. Neuro-Oncology. 2013;15(12):1595–1603. doi: 10.1093/neuonc/not131. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Loewen G, Zhuo Y, Zhuang Y, Jayawickramarajah J, Shan B. lincRNA HOTAIR as a novel promoter of cancer progression. J Canc Res Updates. 2014;3(3):7. doi: 10.6000/1929-2279.2014.03.03.3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Gutierrez SE, Romero-Oliva FA. Epigenetic changes: a common theme in acute myelogenous leukemogenesis. J Hematol Oncol. 2013;6:57. doi: 10.1186/1756-8722-6-57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Zhang X, Zhou L, Fu G, Sun F, Shi J, Wei J, Lu C, Zhou C, Yuan Q, Yang M. The identification of an ESCC susceptibility SNP rs920778 that regulates the expression of lncRNA HOTAIR via a novel intronic enhancer. Carcinogenesis. 2014;35(9):2062–2067. doi: 10.1093/carcin/bgu103. [DOI] [PubMed] [Google Scholar]
- 85.Ma MZ, Li CX, Zhang Y, Weng MZ, Zhang MD, Qin YY, Gong W, Quan ZW. Long non-coding RNA HOTAIR, a c-Myc activated driver of malignancy, negatively regulates miRNA-130a in gallbladder cancer. Mol Cancer. 2014;13:156. doi: 10.1186/1476-4598-13-156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Zajac-Kaye M. Myc oncogene: a key component in cell cycle regulation and its implication for lung cancer. Lung Cancer. 2001;34(Suppl 2):S43–S46. doi: 10.1016/S0169-5002(01)00343-9. [DOI] [PubMed] [Google Scholar]
- 87.Li LC, Dahiya R. MethPrimer: designing primers for methylation PCRs. Bioinformatics. 2002;18(11):1427–1431. doi: 10.1093/bioinformatics/18.11.1427. [DOI] [PubMed] [Google Scholar]
- 88.Yoon JH, Abdelmohsen K, Gorospe M. Functional interactions among microRNAs and long noncoding RNAs. Semin Cell Dev Biol. 2014;34C:9–14. doi: 10.1016/j.semcdb.2014.05.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Brabletz S, Brabletz T. The ZEB/miR-200 feedback loop–a motor of cellular plasticity in development and cancer? EMBO Rep. 2010;11(9):670–677. doi: 10.1038/embor.2010.117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Yoon JH, Abdelmohsen K, Kim J, Yang X, Martindale JL, Tominaga-Yamanaka K, White EJ, Orjalo AV, Rinn JL, Kreft SG, Wilson GM, Gorospe M. Scaffold function of long non-coding RNA HOTAIR in protein ubiquitination. Nat Commun. 2013;4:2939. doi: 10.1038/ncomms3939. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Osada H, Takahashi T. let-7 and miR-17-92: small-sized major players in lung cancer development. Cancer Sci. 2011;102(1):9–17. doi: 10.1111/j.1349-7006.2010.01707.x. [DOI] [PubMed] [Google Scholar]
- 92.Kasinski AL, Slack FJ. miRNA-34 prevents cancer initiation and progression in a therapeutically resistant K-ras and p53-induced mouse model of lung adenocarcinoma. Cancer Res. 2012;72(21):5576–5587. doi: 10.1158/0008-5472.CAN-12-2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Gallardo E, Navarro A, Vinolas N, Marrades RM, Diaz T, Gel B, Quera A, Bandres E, Garcia-Foncillas J, Ramirez J, Monzo M. miR-34a as a prognostic marker of relapse in surgically resected non-small-cell lung cancer. Carcinogenesis. 2009;30(11):1903–1909. doi: 10.1093/carcin/bgp219. [DOI] [PubMed] [Google Scholar]
- 94.Baffa R, Fassan M, Volinia S, O’Hara B, Liu CG, Palazzo JP, Gardiman M, Rugge M, Gomella LG, Croce CM, Rosenberg A. MicroRNA expression profiling of human metastatic cancers identifies cancer gene targets. J Pathol. 2009;219(2):214–221. doi: 10.1002/path.2586. [DOI] [PubMed] [Google Scholar]
- 95.Shan B, Yao TP, Nguyen HT, Zhuo Y, Levy DR, Klingsberg RC, Tao H, Palmer ML, Holder KN, Lasky JA. Requirement of HDAC6 for transforming growth factor-beta1-induced epithelial-mesenchymal transition. J Biol Chem. 2008;283(30):21065–21073. doi: 10.1074/jbc.M802786200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Antoon JW, Lai R, Struckhoff AP, Nitschke AM, Elliott S, Martin EC, Rhodes LV, Yoon NS, Salvo VA, Shan B, Beckman BS, Nephew KP, Burow ME. Altered death receptor signaling promotes epithelial-to-mesenchymal transition and acquired chemoresistance. Sci Rep. 2012;2:539. doi: 10.1038/srep00539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Zhou C, Nitschke AM, Xiong W, Zhang Q, Tang Y, Bloch M, Elliott S, Zhu Y, Bazzone L, Yu D, Weldon CB, Schiff R, McLachlan JA, Beckman BS, Wiese TE, Nephew KP, Shan B, Burow ME, Wang G. Proteomic analysis of tumor necrosis factor-alpha resistant human breast cancer cells reveals a MEK5/Erk5-mediated epithelial-mesenchymal transition phenotype. Breast Cancer Res. 2008;10(6):R105. doi: 10.1186/bcr2210. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Zhuang Y, Nguyen HT, Burow ME, Zhuo Y, El-Dahr SS, Yao X, Cao S, Flemington EK, Nephew KP, Fang F, Collins-Burow B, Rhodes LV, Yu Q, Jayawickramarajah J, Shan B: Elevated expression of long intergenic non-coding RNA HOTAIR in a basal-like variant of MCF-7 breast cancer cells.Mol Carcinog 2014. [Epub ahead of print], [DOI] [PMC free article] [PubMed]
- 99.Li C, Nguyen HT, Zhuang Y, Lin Y, Flemington EK, Guo W, Guenther J, Burow ME, Morris GF, Sullivan D, Shan B. Post-transcriptional up-regulation of miR-21 by type I collagen. Mol Carcinog. 2011;50(7):563–570. doi: 10.1002/mc.20742. [DOI] [PubMed] [Google Scholar]
- 100.Li C, Nguyen HT, Zhuang Y, Lin Z, Flemington EK, Zhuo Y, Kantrow SP, Morris GF, Sullivan DE, Shan B. Comparative profiling of miRNA expression of lung adenocarcinoma cells in two-dimensional and three-dimensional cultures. Gene. 2012;511(2):143–150. doi: 10.1016/j.gene.2012.09.093. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Nguyen HT, Li C, Lin Z, Zhuang Y, Flemington EK, Burow ME, Lin Y, Shan B. The microRNA expression associated with morphogenesis of breast cancer cells in three-dimensional organotypic culture. Oncol Rep. 2012;28(1):117–126. doi: 10.3892/or.2012.1764. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Nguyen HT, Zhuang Y, Sun L, Kantrow SP, Kolls JK, You Z, Zhuo Y, Shan B. Src-mediated morphology transition of lung cancer cells in three-dimensional organotypic culture. Cancer Cell Int. 2013;13(1):16. doi: 10.1186/1475-2867-13-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Mandeville I, Aubin J, LeBlanc M, Lalancette-Hebert M, Janelle MF, Tremblay GM, Jeannotte L. Impact of the loss of Hoxa5 function on lung alveogenesis. Am J Pathol. 2006;169(4):1312–1327. doi: 10.2353/ajpath.2006.051333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Liu XH, Lu KH, Wang KM, Sun M, Zhang EB, Yang JS, Yin DD, Liu ZL, Zhou J, Liu ZJ, De W, Wang ZX. MicroRNA-196a promotes non-small cell lung cancer cell proliferation and invasion through targeting HOXA5. BMC Cancer. 2012;12:348. doi: 10.1186/1471-2407-12-348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Thiery JP, Acloque H, Huang RY, Nieto MA. Epithelial-mesenchymal transitions in development and disease. Cell. 2009;139(5):871–890. doi: 10.1016/j.cell.2009.11.007. [DOI] [PubMed] [Google Scholar]
- 106.Shan B, Zhuo Y, Chin D, Morris CA, Morris GF, Lasky JA. Cyclin-dependent kinase 9 is required for tumor necrosis factor-alpha-stimulated matrix metalloproteinase-9 expression in human lung adenocarcinoma cells. J Biol Chem. 2005;280(2):1103–1111. doi: 10.1074/jbc.M406293200. [DOI] [PubMed] [Google Scholar]
- 107.Coe BP, Thu KL, Aviel-Ronen S, Vucic EA, Gazdar AF, Lam S, Tsao MS, Lam WL. Genomic deregulation of the E2F/Rb pathway leads to activation of the oncogene EZH2 in small cell lung cancer. PLoS One. 2013;8(8):e71670. doi: 10.1371/journal.pone.0071670. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Liu C, Shi X, Wang L, Wu Y, Jin F, Bai C, Song Y. SUZ12 is involved in progression of non-small cell lung cancer by promoting cell proliferation and metastasis. Tumour Biol. 2014;35(6):6073–6082. doi: 10.1007/s13277-014-1804-5. [DOI] [PubMed] [Google Scholar]
- 109.Hayami S, Kelly JD, Cho HS, Yoshimatsu M, Unoki M, Tsunoda T, Field HI, Neal DE, Yamaue H, Ponder BA, Nakamura Y, Hamamoto R. Overexpression of LSD1 contributes to human carcinogenesis through chromatin regulation in various cancers. Int J Cancer. 2011;128(3):574–586. doi: 10.1002/ijc.25349. [DOI] [PubMed] [Google Scholar]
- 110.Lv T, Yuan D, Miao X, Lv Y, Zhan P, Shen X, Song Y. Over-expression of LSD1 promotes proliferation, migration and invasion in non-small cell lung cancer. PLoS One. 2012;7(4):e35065. doi: 10.1371/journal.pone.0035065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Sekido Y, Fong KM, Minna JD. Molecular genetics of lung cancer. Annu Rev Med. 2003;54:73–87. doi: 10.1146/annurev.med.54.101601.152202. [DOI] [PubMed] [Google Scholar]
- 112.Lee Y, Fryer JD, Kang H, Crespo-Barreto J, Bowman AB, Gao Y, Kahle JJ, Hong JS, Kheradmand F, Orr HT, Finegold MJ, Zoghbi HY. ATXN1 protein family and CIC regulate extracellular matrix remodeling and lung alveolarization. Dev Cell. 2011;21(4):746–757. doi: 10.1016/j.devcel.2011.08.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Li X, Shi Y, Yin Z, Xue X, Zhou B. An eight-miRNA signature as a potential biomarker for predicting survival in lung adenocarcinoma. J Transl Med. 2014;12:159. doi: 10.1186/1479-5876-12-159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 114.Acunzo M, Visone R, Romano G, Veronese A, Lovat F, Palmieri D, Bottoni A, Garofalo M, Gasparini P, Condorelli G, Chiariello M, Croce CM. miR-130a targets MET and induces TRAIL-sensitivity in NSCLC by downregulating miR-221 and 222. Oncogene. 2012;31(5):634–642. doi: 10.1038/onc.2011.260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Arita T, Ichikawa D, Konishi H, Komatsu S, Shiozaki A, Shoda K, Kawaguchi T, Hirajima S, Nagata H, Kubota T, Fujiwara H, Okamoto K, Otsuji E. Circulating long non-coding RNAs in plasma of patients with gastric cancer. Anticancer Res. 2013;33(8):3185–3193. [PubMed] [Google Scholar]
- 116.Volinia S, Calin GA, Liu CG, Ambs S, Cimmino A, Petrocca F, Visone R, Iorio M, Roldo C, Ferracin M, Prueitt RL, Yanaihara N, Lanza G, Scarpa A, Vecchione A, Negrini M, Harris CC, Croce CM. A microRNA expression signature of human solid tumors defines cancer gene targets. Proc Natl Acad Sci U S A. 2006;103(7):2257–2261. doi: 10.1073/pnas.0510565103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Kikuchi J, Takashina T, Kinoshita I, Kikuchi E, Shimizu Y, Sakakibara-Konishi J, Oizumi S, Marquez VE, Nishimura M, Dosaka-Akita H. Epigenetic therapy with 3-deazaneplanocin A, an inhibitor of the histone methyltransferase EZH2, inhibits growth of non-small cell lung cancer cells. Lung Cancer. 2012;78(2):138–143. doi: 10.1016/j.lungcan.2012.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Sato T, Kaneda A, Tsuji S, Isagawa T, Yamamoto S, Fujita T, Yamanaka R, Tanaka Y, Nukiwa T, Marquez VE, Ishikawa Y, Ichinose M, Aburatani H. PRC2 overexpression and PRC2-target gene repression relating to poorer prognosis in small cell lung cancer. Sci Rep. 2013;3:1911. doi: 10.1038/srep01911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Zhao J, Ohsumi TK, Kung JT, Ogawa Y, Grau DJ, Sarma K, Song JJ, Kingston RE, Borowsky M, Lee JT. Genome-wide identification of polycomb-associated RNAs by RIP-seq. Mol Cell. 2010;40(6):939–953. doi: 10.1016/j.molcel.2010.12.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Khalil AM, Guttman M, Huarte M, Garber M, Raj A, Rivea Morales D, Thomas K, Presser A, Bernstein BE, van Oudenaarden A, Regev A, Lander ES, Rinn JL. Many human large intergenic noncoding RNAs associate with chromatin-modifying complexes and affect gene expression. Proc Natl Acad Sci U S A. 2009;106(28):11667–11672. doi: 10.1073/pnas.0904715106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Lee ST, Li Z, Wu Z, Aau M, Guan P, Karuturi RK, Liou YC, Yu Q. Context-specific regulation of NF-kappaB target gene expression by EZH2 in breast cancers. Mol Cell. 2011;43(5):798–810. doi: 10.1016/j.molcel.2011.08.011. [DOI] [PubMed] [Google Scholar]
- 122.He A, Shen X, Ma Q, Cao J, von Gise A, Zhou P, Wang G, Marquez VE, Orkin SH, Pu WT. PRC2 directly methylates GATA4 and represses its transcriptional activity. Genes Dev. 2012;26(1):37–42. doi: 10.1101/gad.173930.111. [DOI] [PMC free article] [PubMed] [Google Scholar]