

Abstract
In a screening of about 500 lines of Tartary buckwheat, we identified lines that contained no detectable rutinosidase isozymes using an in-gel detection assay. We confirmed that seeds of these individuals had only a trace level of in-vitro rutinosidase activity. To investigate the heritability of the trace-rutinosidase characteristic, we analyzed the progeny of crosses between rutinosidase trace-lines, ‘f3g-162’, and the ‘Hokkai T8’. The F2 progeny clearly divided into two groups: those with rutinosidase activity under 1.5 nkat/g seed (trace-rutinosidase) and those with activity over 400 nkat/g seed (normal rutinosidase). The segregation pattern of this trait in F2 progeny exhibited 1 : 3 ratio (trace-rutinosidase : normal rutinosidase), suggesting that the trace-rutinosidase trait is conferred by a single recessive gene; rutinosidase-trace A (rutA). In addition, sensory panelists evaluated the bitterness of flour from trace-rutinosidase individuals and did not detect bitterness, whereas flour from normal rutinosidase individuals was found to have strong bitterness. Although at least three bitter compounds have been reported in Tartary buckwheat seeds, our present findings indicate that rutin hydrolysis is the major contributing factor to bitterness. In addition, the trace-rutinosidase line identified here, ‘f3g-162’, is a promising material for generating a non-bitter Tartary buckwheat variety.
Keywords: Tartary buckwheat, bitterness, rutinosidase, genetic resources, quality
Introduction
Rutin is a type of flavonoid that has numerous biological functions, including the strengthening of blood capillaries (Griffith et al. 1944, Shanno 1946), antioxidative (Awatsuhara et al. 2010, Jiang et al. 2007), antihypertensive (Matsubara et al. 1985) and anti-inflammatory activities, and alpha-glucosidase inhibitory activity (Li et al. 2009). The clinical effects of rutin were investigated in a double-blind crossover study that showed that treatment with rutin led to reductions in serum myeloperoxidase and cholesterol levels (Wieslander et al. 2011), mucosal symptoms, headache, and tiredness (Wieslander et al. 2012). Due to these beneficial properties, plants with high levels of rutin are considered to have great therapeutic potential. Although rutin is widely distributed in the plant kingdom (Couch et al. 1946, Fabjan et al. 2003, Haley and Bassin 1954, Sando and Lloyd 1924), buckwheat is the only known cereal to contain high levels of rutin in seeds. For this reason, buckwheat has been utilized as a rutin-rich material for numerous food products (Ikeda et al. 2012, Kreft et al. 2006).
Among buckwheat species, Tartary buckwheat (Fagopyrum tataricum Gaertn.) is a particularly rich source of rutin, containing approximately 100-fold higher concentrations of rutin in seeds than that of common buckwheat. However, Tartary buckwheat seeds also have markedly high rutinosidase activity, which is sufficient to hydrolyze the rutin present in buckwheat flour (approximately 1%–2% [w/w]) to quercetin and rutinose (Fig. 1) within a few minutes after the addition of water (Suzuki et al. 2002, Yasuda et al. 1992, Yasuda and Nakagawa 1994). Therefore, the identification of Tartary buckwheat varieties with low rutinosidase activity is desirable for the production of flour with a high rutin content.
Fig. 1.
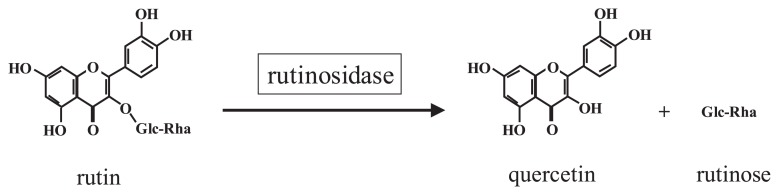
Rutinosidase in Tartary buckwheat seeds converts rutin to quercetin and the disaccharide rutinose.
Rutinosidase is a β-glycosidase that releases the disaccharide rutinose from rutin. Tartary buckwheat seeds contain at least two rutinosidase isozymes, which have similar characteristics with respect to substrate specificity (Km) and optimum temperature and pH (Suzuki et al. 2002, Yasuda et al. 1992, Yasuda and Nakagawa 1994). Therefore, for a variety of Tartary buckwheat to have low rutinosidase activity in seeds, the expression of both isozymes must be suppressed. In addition to rutinosidase activity, Tartary buckwheat flour has strong bitterness, a property that has limited its use in food products. Kawakami et al. (1995) reported that Tartary buckwheat dough contains at least three bitter compounds: quercetin, which is the hydrolyzed moiety of rutin, and unidentified compounds ‘F3’ and ‘F4’. Therefore, many researchers have suggested that suppression of rutinosidase activity may contribute to reduced bitterness. However, it remains uncertain which one of these three compounds is the major cause of bitterness in Tartary buckwheat. To resolve this issue, development of a variety with low rutinosidase activity is necessary to reduce quercetin production and allow the contribution of ‘F3’ and ‘F4’ to bitterness generation to be determined. To date, however, no reports have described the development of non-bitter or low rutinosidase lines of Tartary buckwheat.
In this report, we describe the identification of lines contained individual of Tartary buckwheat with only trace-rutinosidase activity. In addition, we performed genetic analysis to clear the heredity of this trait and discuss the effects of rutin hydrolysis on bitterness.
Materials and Methods
Plant materials, cultivation, and screening of low rutinosidase individuals
Tartary buckwheat seeds were sown in late May at a density of 67 seeds/m2 and 60-cm row spacing in a 1.2 m2 plot of the experimental field at the Memuro Upland Farming Research Station of the National Agriculture and Food Research Organization, Hokkaido Agricultural Research Center (Shinsei, Memuro, Kasai-Gun, Japan; longitude, 143°03′E; latitude, 42°55′N). The fertilizer applied contained 0.6 kg N, 1.8 kg P2O5, 1.4 kg K2O, and 0.5 kg MgO per 10 are. After cultivation for about 10–16 weeks, the Tartary buckwheat plants were harvested, threshed using a threshing machine, and the seeds were stored at 4°C. We screened approximately 200 genetic resources and 300 ethyl methanesulfonate (EMS) mutant lines (about 8,400 plants) for rutinosidase activity using an in-gel detection method, as previously described (Suzuki et al. 2004). EMS mutant lines were developed using the ‘Hokkai T8’ variety by suspending seeds in 2% (v/v) EMS for 4 h before seeding. In Tartary buckwheat seeds, rutinosidase activity is distributed in testa, a thin layer-like organ that covers the embryo (Suzuki et al. 2002). The generation of testa is the same as a plant having the testa. Therefore, M3 seeds, whose testa is derived from the M2 generation, were used for screening of rutinosidase activity. Genetic resources were obtained from the National Institute of Agrobiological Sciences (NIAS) Genebank (NIAS, 2-1-2 Kannondai, Tsukuba, Ibaraki 305-8602, Japan). Genetic resources that were originally obtained from Nepal, such as the parent of ‘f3g-162’, and are stored at our institute were also used. The trace-rutinosidase individuals were sown, grown for approximately 12–16 weeks, and individually harvested. After harvesting seeds, the isozyme composition of rutinosidase was again investigated using the in-gel detection method. Individuals in which no examined seeds contained rutinosidase isozymes were selected as trace-rutinosidase lines, and included ‘f3g-162’ (Table 1).
Table 1.
Results of screening for trace rutinosidase activity
| Conservation Lot No.a | Numbers of individuals | Rate of rutinosidase-trace individuals (%) | Collection country, region | Collection altitude (m) | |
|---|---|---|---|---|---|
|
| |||||
| Normal rutinosidaseb | Trace-rutinosidasec | ||||
| 3027881 | 30 | 26 | 46.4 | Nepal, Phidim | 1,700 |
| 3027884 | 52 | 4 | 7.1 | Nepal, Phidim | 1,900 |
| 94182 | 39 | 17 | 30.4 | Nepal, Chamaita | 2,000 |
| 94184 | 31 | 25 | 44.6 | Nepal, Chamaita | 2,100 |
Stored at the National Institute of Agrobiological Sciences; Genebank; 2-1-2 Kannondai, Tsukuba, Ibaraki 305-8602, Japan.
Individuals with rutinosidase activity higher than 400 nkat/g seed.
Individuals with rutinosidase activity lower than 1.5 nkat/g seed.
Standard assay for in-vitro rutinosidase activity and determination of rutin concentration
A standard assay for rutinosidase activity was performed by measuring the quercetin concentration in a reaction mixture using high-pressure liquid chromatography (HPLC). The standard assay mixture consisted of 50 mM acetate-NaOH buffer (pH 5.0 at 4°C), 20% (v/v) methanol, and 0.2% (w/v) rutin in a final volume of 0.05 ml. The reaction was performed at 37°C and was stopped by the addition of 0.2 ml methanol (Suzuki et al. 2002). Rutin concentrations in seeds and dough were measured by HPLC as previously described (Suzuki et al. 2002).
Detection of rutinosidase isozymes by native PAGE with a rutin-copper complex
Tartary buckwheat has at least two rutinosidase isozymes. To accurately screen for rutinosidase activity, an assay that can detect all isozymes is needed. In the present study, rutinosidase isozymes were detected using native-PAGE gels and a rutin-copper complex, as previously described (Suzuki et al. 2004). Briefly, one Tartary buckwheat seed was homogenized with 2 ml extraction buffer containing 10% (v/v) glycerol and 0.01% (w/v) Bromophenol blue (BPB) in 50 mM acetate-NaOH buffer (pH 5.0). The resulting crude enzyme solution was loaded onto a 7.5% (w/v) native-PAGE gel and separated under non-denaturing conditions using a slab gel apparatus (Bio-Rad). After electrophoresis, the gel was equilibrated with 50 mM acetate- NaOH buffer (pH 5.0 at 4°C) contained 20% (v/v) methanol for 10 min. The equilibrated gel was stained using 50 mM acetate-NaOH buffer (pH 5.0 at 4°C), 20% (v/v) methanol, 0.6% (w/v) rutin, and 5 mM CuSO4 for 20 min (Suzuki et al. 2004). The gel was removed from the staining buffer and washed thoroughly with water.
Analysis of progeny from hybridization between ‘f3g-162’ (trace-rutinosidase line) and ‘Hokkai T8’ (normal rutinosidase variety)
The F1 seeds of a cross between ‘f3g-162’ and ‘Hokkai T8’ were planted in pots, and F2 seeds were harvested from each F1 plant. A total of 151 seeds of the F2 population were planted, and segregation of rutinosidase activity, isozyme composition of rutinosidase, rutin concentration, and bitterness in F3 seeds (testa is derived from F2 progeny) were investigated. The bitterness of the dough prepared from flour made from F3 seeds was evaluated by four expert panelists, as described below. Chi-square analysis was used to examine the fit of the observed segregations to the expected ratios.
Bitterness evaluation of Tartary buckwheat dough
To prepare dough, 250 μL water was thoroughly mixed with 250 mg Tartary buckwheat flour. Three minutes after the addition of water, four expert panelists placed a sample of the prepared dough in their mouth and assessed the degree of bitterness over a period of approximately 1 min. Based on this sensory evaluation, the panelists classified the dough samples as ‘bitter’ or ‘not bitter’.
Results
Screening for low rutinosidase lines of Tartary buckwheat
We identified four genetic resources of Tartary buckwheat; each genetic resource contained both normal rutinosidase and trace-rutinosidase individuals (Table 1). We could not detect any rutinosidase isozymes using an in-gel detection method in the trace-rutinosidase individuals in these four genetic resources (Fig. 2). The in-vitro rutinosidase activity in the seeds of individuals within these resources was also measured and used to divide seeds into two groups: those with high enzyme activity (>400 nkat/g seed) and those will trace enzyme activity (<1.5 nkat/g seed). Among the examined genetic resources, the individuals with trace-rutinosidase characteristics in seeds ranged from 7.1% to 46.4%, whereas the other individuals had normal rutinosidase levels. The geographic location where each genetic resource was collected by Namai and Gotoh (1994) is indicated on the map of eastern Nepal (Fig. 3). The genetic resources with trace-rutinosidase activity were collected from the foot of the mountain located near Chamaita, south of the Hewa River (Fig. 3).
Fig. 2.

Detection of rutinosidase isozymes using an in-gel detection method. A–D: Typical native-PAGE results for the crude protein extracts from trace-rutinosidase individuals. E: ‘Hokkai T8’ (normal rutinosidase variety; pollen parent). Rutinosidase isozymes in seeds were detected using an in-gel detection method involving the native-PAGE separation of crude protein extracts.
Fig. 3.

Map showing the locations of Tartary buckwheat samples collected in eastern Nepal, as reported by Namai and Gotoh (1994). Solid lines indicate major ridges and broken lines indicate rivers. For each collection area, the total number of Tartary buckwheat samples collected is shown, and the number of samples with trace-rutinosidase activity is indicated in parentheses.
Among the identified individuals with trace-rutinosidase activity in seeds, we selected line ‘f3g-162’ for genetic analysis to identify the loci responsible for this trait.
Progeny analysis of hybridization between ‘f3g-162’ (trace-rutinosidase line) and ‘Hokkai T8’ (normal rutinosidase variety)
To clear heredity of the trace-rutinosidase trait, progeny analysis of the hybrids between ‘f3g-162’ and ‘Hokkai T8’ was performed (Fig. 4). Specifically, the rutinosidase activity, rutin concentration, and bitterness of seeds of the obtained hybrids were investigated. With respect to rutinosidase activity, the progeny could be clearly divided into two groups: those with activity under 1.5 nkat/g seed (trace-rutinosidase individuals) and those with activity over 400 nkat/g seed (normal rutinosidase individuals). No individuals with rutinosidase activities between 1.5 and 400 nkat/g seed were found. Among the 157 F2 progeny, 42 were trace-rutinosidase individuals and 115 were normal rutinosidase individuals, a segregation pattern corresponding to 1 : 3 ratio (trace-rutinosidase : normal rutinosidase, χ2-value for 3 : 1 = 0.26, P = 0.61). This finding suggested that the trace-rutinosidase trait is dominated by a single recessive gene, which we named rutinosidase-trace A (rutA).
Fig. 4.

Progeny analysis of hybrids between ‘f3g-162’ (trace-rutinosidase line) and ‘Hokkai T8’ (normal rutinosidase variety). Rutinosidase activity and bitterness in the F2 population from a cross between ‘f3g-162’ and ‘Hokkai T8’. Chi-square analysis was used to examine the fit of the observed segregations to the expected ratios.
We also investigated the isozyme composition of each F2 individual generated from the ‘f3g-162’ and ‘Hokkai T8’ crossing using the in-gel detection method for rutinosidase activity. Among individuals in the trace-rutinosidase group, no rutinosidase isozymes were detected. In contrast, two isozymes were detected in all the individuals in the normal rutinosidase group. In both the trace- and normal rutinosidase groups, the rutin concentration in seeds ranged from 13 to 19 mg/g.
We evaluated and compared the bitterness of flour prepared from seeds of both rutinosidase groups. Among trace-rutinosidase individuals, none of the panelists detected bitterness in any of the flour samples, whereas all flour samples from normal rutinosidase individuals were uniformly found to have strong bitterness. Finally, we confirmed that almost no rutin had been hydrolyzed in dough prepared from the flour of trace-rutinosidase individuals, whereas rutin was completely hydrolyzed in dough samples from individuals in the normal rutinosidase group (data not shown).
Discussion
Discovery of trace-rutinosidase Tartary buckwheat lines
Before screening for Tartary buckwheat varieties with low rutinosidase activity, we first attempted to identify individuals that lacked at least one of the two known rutinosidase isozymes using an in-gel detection method. Notably, however, only individuals lacking both isozymes were found among the examined genetic resources. As the rutinosidase activity in seeds of these individuals was below 1.5 nkat/g, we considered that seeds with no detectable rutinosidase isozymes in the in-gel detection assay have only trace-rutinosidase activity.
Progeny analysis of hybridization between ‘f3g-162’ (trace-rutinosidase line) and ‘Hokkai T8’ (normal rutinosidase variety)
The results of progeny analysis indicate that the trace-rutinosidase characteristic found in ‘f3g-162’ is conferred by a single recessive gene, which we have named rutA. However, Suzuki et al. (2002) and Yasuda and Nakagawa (1994) reported that Tartary buckwheat seeds contain at least two rutinosidase isozymes, and more than two cDNAs corresponding to rutinosidase were cloned and sequenced by Fujino et al. (2012). Our present finding suggests that both isozyme genes are tightly linked and/or controlled at the transcriptional level by a single regulatory gene. To clarify how trace-rutinosidase activity is conferred by the rutA gene, identification and sequencing of the locus is required to understand the mechanism underlying the expression and activation of rutinosidase. In the flour of trace-rutinosidase individuals, none of the sensory panelists perceived bitterness, whereas strong bitterness was detected in all flour samples from normal rutinosidase individuals. Quercetin, the hydrolysis product of rutin by rutinosidase, has been identified as the principle bittering compound in several fruits (Hladik and Bruno 1996, Peterson and Dwyer 1998). However, as described above, Kawakami et al. (1995) reported that Tartary buckwheat dough contains at least three bitter compounds: quercetin, and the unidentified compounds ‘F3’ and ‘F4’. Although it remains unclear which of these three compounds is the major cause of bitterness in Tartary buckwheat, the present findings clearly demonstrate that rutin hydrolysis leads to the generation of bitterness in seeds.
Based on these results, the ‘f3g-162’ of Tartary buckwheat is a promising breeding material because of its non-bitter and trace-rutinosidase characteristics. Unfortunately, the agronomical characteristics of ‘f3g-162’ such as maturing time and yield are not suitable for cultivation in the Hokkaido region, which is the largest production area of Tartary buckwheat in Japan. To develop a non-bitter variety of Tartary buckwheat for large-scale agronomic cultivation, the cross breeding of ‘f3g-162’ with a leading variety of buckwheat is required.
Acknowledgements
We thank Mr. T. Saruwatari, Mr. N. Murakami, Mr. S. Nakamura, Mr. K. Abe, Mr. T. Yamada, Mr. T. Takakura, Mr. T. Fukaya, Mr. M. Oizumi, Mr. T. Hirao, and Mr. K. Suzuki for their assistance in the field. We also thank Ms. K. Fujii, Ms. M. Hayashida, and Ms. T. Ando for technical assistance. Screening of the trace-rutinosidase line was performed as part of the NIAS ‘Gene bank Project’ of the National Institute of Agrobiological Sciences, Japan. Breeding and evaluation of ‘Manten-Kirari’ was partly supported by ‘Research and Development Projects for Application in Promoting New Policy of Agriculture, Forestry and Fisheries’ and ‘grant from the Research Project on Development of Agricultural Products and Foods with Health-promoting benefits (NARO), Japan’.
Literature Cited
- Awatsuhara, R., Harada, K., Maeda, T., Nomura, T. and Nagao, N. (2010) Antioxidative activity of the buckwheat polyphenol rutin in combination with ovalbumin. Mol. Med. Rep. 3: 121–125. [DOI] [PubMed] [Google Scholar]
- Couch, J.F., Naghski, J. and Krewson, C.F. (1946) Buckwheat as a source of rutin. Science 103: 197–198. [DOI] [PubMed] [Google Scholar]
- Fabjan, N., Rode, J., Kosir, I.J., Wang, Z., Zhang, Z. and Kreft, I. (2003) Tartary buckwheat (Fagopyrum tataricum Gaertn.) as a source of dietary rutin and quercitrin. J. Agric. Food Chem. 51: 6452–6455. [DOI] [PubMed] [Google Scholar]
- Fujino, K., Matsui, K., Suzuki, T. and Morishita, T. (2012) Analysis of rutinosidase in the new breeding line of Fagopyrum tartaricum. Jpn. J. Crop Sci. 81: 170–171. [Google Scholar]
- Griffith, J.Q., Couch, J.F. and Lindauer, A. (1944) Effect of rutin on increased capillary fragility in man. Proc. Soc. Exp. Biol. Med. 55: 228–229. [Google Scholar]
- Haley, T.J. and Bassin, M. (1951) The isolation, purification and derivatives of plant pigments related to rutin. J. Am. Pharm. Assoc. 40: 111–112. [PubMed] [Google Scholar]
- Hladik, C.M. and Sinmen, B. (1996) Taste perception and feeding behavior in nonhuman primates and human populations. Evol. Anthropol. 5: 58–71. [Google Scholar]
- Ikeda, K., Ikeda, S., Kreft, I. and Rufa, L. (2012) Utilization of Tartary buckwheat. Fagopyrum 29: 27–30. [Google Scholar]
- Jiang, P., Burczynski, F., Campbell, C.J., Pierce, G., Austria, J.A. and Briggs, C.J. (2007) Rutin and flavonoid contents in three buckwheat species Fagopyrum esculentum, F. tataricum, and F. homotropicum and their protective effects against lipid peroxidation. Food Res. Int. 40: 356–364. [Google Scholar]
- Kawakami, A., Kayahara, H. and Ujihara, A. (1995) Properties and elimination of bitter components derived from Tartary buckwheat (Fagopyrum tataricum) flour. Nippon Shokuhin Kagaku Kogaku Kaishi 42: 892–898. [Google Scholar]
- Kreft, I., Fabjan, N. and Yasumoto, K. (2006) Rutin content in buckwheat (Fagopyrum esculentum Moench) food materials and products. Food Chem. 98: 508–512. [Google Scholar]
- Li, Y.Q., Zhou, F.C., Gao, F., Bian, J.S. and Shan, F. (2009) Comparative evaluation of quercetin, isoquercetin and rutin as inhibitors of alpha-glucosidase. J. Agric. Food Chem. 57: 11463–11468. [DOI] [PubMed] [Google Scholar]
- Matsubara, Y., Kumamoto, H., Iizuka, Y., Murakami, T., Okamoto, K., Miyake, H. and Yokoi, K. (1985) Structure and hypotensive effect of flavonoid glycosides in Citrus unshiu peelings. Agric. Biol. Chem. 49: 909–914. [Google Scholar]
- Namai, H. and Gotoh, T. (1994) Title in Japanese. Norin Suisan Gijutu Joho Kyokai Norinsuisan Gene Bank no Kisyohseibutsu toh no Idenshigen Chousa Shuushuu Itaku Jigho Seika Hokokusho IV: 1–26. [Google Scholar]
- Peterson, J. and Dwyer, J. (1998) Flavonoids: dietary occurrence and biochemical activity. Nutr. Res. 18: 1995–2018. [Google Scholar]
- Sando, C.E. and Lloyd, J.U. (1924) The isolation and identification of rutin from the flowers of elder (Sambucus canadensis L.). J. Biol. Chem. 58: 737–745. [Google Scholar]
- Shanno, R.L. (1946) Rutin: A new drug for the treatment of increased capillary fragility. Am. J. Med. Sci. 211: 539–543. [PubMed] [Google Scholar]
- Suzuki, T., Honda, Y., Funatsuki, W. and Nakatsuka, K. (2002) Purification and characterization of flavonol 3-glucosidase, and its activity during ripening in Tartary buckwheat seeds. Plant Sci. 163: 417–423. [Google Scholar]
- Suzuki, T., Honda, Y., Funatsuki, W. and Nakatsuka, K. (2004) In-gel detection and study of the role of flavonol 3-glucosidase in the bitter taste generation in tartary buckwheat. J. Sci. Food Agric. 84: 1691–1694. [Google Scholar]
- Wieslander, G., Fabjan, N., Vogrinčič, M., Kreft, I., Janson, C., Spetz-Nyström, U., Vombergar, B., Tagesson, C., Leanderson, P. and Norbäck, D. (2011) Eating buckwheat cookies is associated with the reduction in serum levels of myeloperoxidase and cholesterol: A double blind crossover study in day-care center staffs. Tohoku J. Exp. Med. 225: 123–130. [DOI] [PubMed] [Google Scholar]
- Wieslander, G., Fabjan, N., Vogrinčič, M., Kreft, I., Vombergar, B. and Norbäck, D. (2012) Effect of common and Tartary buckwheat consumption on mucosal symptoms, headache and tiredness: A double-blind crossover intervention study. J. Food Agric. Environ. 10: 107–110. [Google Scholar]
- Yasuda, T., Masaki, K. and Kashiwagi, T. (1992) An enzyme degrading rutin in Tartary buckwheat seeds. J. Jpn. Soc. Food Sci. Technol. 39: 994–1000. [Google Scholar]
- Yasuda, T. and Nakagawa, H. (1994) Purification and characterization of rutin-degrading enzymes in Tartary buckwheat seeds. Phytochem. 37: 133–136. [Google Scholar]