

Abstract
Mumps is caused by the mumps virus (MuV), a member of the Paramyxoviridae family of enveloped, non-segmented, negative-sense RNA viruses. Mumps is characterized by painful inflammatory symptoms, such as parotitis and orchitis. The virus is highly neurotropic, with laboratory evidence of central nervous system (CNS) infection in approximately half of cases. Symptomatic CNS infection occurs less frequently; nonetheless, prior to the introduction of routine vaccination, MuV was a leading cause of aseptic meningitis and viral encephalitis in many developed countries. Despite being one of the oldest recognized diseases, with a worldwide distribution, surprisingly little attention has been given to its study. Cases of aseptic meningitis associated with some vaccine strains and a global resurgence of cases, including in highly vaccinated populations, has renewed interest in the virus, particularly in its pathogenesis and the need for development of clinically relevant models of disease. In this review we summarize the current state of knowledge on the virus, its pathogenesis and its clinical and pathological outcomes.
Keywords: mumps, mumps virus, pathogenesis, neurovirulence, vaccine
Introduction
Before routine mumps vaccination programmes were introduced, 95% of adults had serological markers of exposure, with peak acquisition during childhood [1–4]. Following the use of mumps vaccine in the USA in the late 1960s, disease incidence declined dramatically, and by the 1980s very few cases were reported. By 2001 the disease was on the verge of elimination, with <0.1 cases/100,000 population reported [5], representing a 99.9% decrease in disease incidence compared to the prevaccination era. Similar success in mumps control was achieved in other countries through vaccination. However, within a few years of these historic lows, large, sporadic mumps outbreaks began to appear globally, involving a high percentage of persons with a history of vaccination [6–20]. The reason for the lower than expected efficacy of mumps vaccines is a subject of much debate, ranging from waning immunity to the emergence of virus strains that might be capable of escaping immunity engendered by the vaccine. In addition to questions over vaccine efficacy, safety concerns have come to light following reports of meningitis linked to some vaccine strains used outside the USA. This has led to withdrawal of some vaccine strains and, in some cases, complete cessation of mumps vaccination. In Japan, for example, mumps vaccination was removed from the national immunization programme. Japan now has one of the highest rates of mumps among developed countries, with over a million cases reported annually [21,22]. Considering the numerous issues surrounding mumps, including ongoing outbreaks, a review of MuV pathogenesis is timely.
Humans are the only natural host of MuV. The disease is characterized by painful swelling of the parotid glands, but can involve numerous other tissues and organs, resulting in a wide array of inflammatory reactions, including encephalitis, meningitis, orchitis, myocarditis, pancreatitis and nephritis [23]. Mumps is self-limiting, often with complete recovery within a few weeks of symptom onset; however, long-term sequelae, such as paralysis, seizures, cranial nerve palsies, hydrocephalus and deafness, can occur. The disease is rarely fatal and the lack of autopsy tissue limits opportunities to examine disease pathogenesis and pathology. Our current knowledge of MuV pathogenesis is therefore mostly based on animal studies, often following unnatural routes of infection. Consequently, the pathogenesis of the virus in humans remains in question. This review will summarize the current knowledge of the virus, its inferred pathogenesis, clinical manifestations, and the importance of an understanding of disease pathogenesis as a prerequisite to the development of safer and more efficacious vaccines.
Mumps virus
Mumps was first described by Hippocrates in the fifth century BC, in his first Book of Epidemics, but a viral aetiology was not demonstrated until the 1930s, when Johnson and Goodpasture fulfilled Koch’s postulates by transferring the disease from experimentally infected rhesus macaques (Macaca mulatta), to children in his neighbourhood, using a bacteria-free, filter-sterilized preparation of macerated monkey parotid tissue [24,25].
The virus, a member of the family Paramyxoviridae, is an enveloped particle containing a non-segmented negative strand RNA molecule of 15,384 nucleotides. Other significant paramyxoviruses that infect humans and livestock include measles virus, canine distemper virus, parainfluenza virus, Newcastle disease virus, respiratory syncytial virus and metapneumovirus. The encapsidated genome (Figure 1) contains seven tandemly linked transcription units, in the order: nucleo- (N), V/P/I (V/phospho-/I proteins), matrix (M), fusion (F), small hydrophobic (SH), haemagglutinin-neuraminidase (HN) and large (L) proteins [26,27]. The template for viral replication and transcription is the ribonucleoprotein (RNP) complex, which is composed of the negative-strand viral RNA encapsidated by N protein. The RNA-dependent RNA polymerase, a complex of the L and P proteins, acts as a replicase to copy the negative sense (−) RNA to a positive sense (+) RNA and as a transcriptase to generate mRNAs from the (−) RNP by entering at a single promoter at the 3′ end of the genome.
Figure 1.

MuV virion structure. (A) Thin sectioned transmission electron micrograph showing a typical MuV particle alongside (B) a schematic of the particle. The enveloped particles are pleomorphic, in the size range 100–600 nm. Within this structure lies the long, coiled electron-dense ribonucleoprotein (RNP), containing the MuV genome. Small spikes can be observed on the surface of the particle, corresponding to the viral HN and F glycoproteins. The same general features of the MuV particle are shown in the schematic (B). The envelope (blue lines) is studded with the HN (purple) and F (blue) glycoproteins and encases the viral RNP, made up of the RNA genome (3′–5′) in complex with N (yellow), P (orange) and L (gold) proteins. The M protein (red) interacts with the envelope, glycoproteins and the RNP. The V, I and SH proteins are expressed in infected cells, but are not thought to be incorporated within the virion. Photomicrograph courtesy of CDC/A Harrison and FA Murphy (http://phil.cdc.gov/phil/details.asp)
In infected cells the HN and F glycoproteins are transported through the endoplasmic reticulum and Golgi complex to the cell surface. The M protein is involved in localizing the viral RNP to regions of the host cell membrane expressing the F and HN glycoproteins, facilitating budding of the infectious virions from the infected cells [28,29]. The HN glycoprotein is responsible for attachment of the newly budded virus to neighbouring cells via its receptor, sialic acid, which is abundantly present on the surface of most animal cells. The HN glycoprotein, in concert with the F glycoprotein, mediates virus-to-cell fusion and cell-to-cell membrane fusion, facilitating virus spread. The SH protein is thought to play a role in evasion of the host antiviral response by blocking the TNFα-mediated apoptosis pathway [30,31]. This protein is not essential for virus replication, as demonstrated in studies with recombinant (r) MuVs engineered to lack the open reading frame encoding this protein [32]. The V and I proteins are encoded by the same transcriptional unit that encodes the P protein [27,33]. Like the SH protein, the V protein is also involved in evasion of the host antiviral response, where it inhibits IFN production and signalling [34–36]. The role of the I protein is unknown.
Clinical features, pathogenesis and pathology
Given the incidence of mumps in the pre-vaccine era, comparatively little is known about the pathogenesis of the disease. Much is inferred by comparison with related viruses, from experimental infection of laboratory animals, and from the clinical features and pathology of the disease in humans.
Initial infection: targeting of the upper respiratory tract epithelium?
MuV is transmitted via the respiratory route by inhalation or oral contact with infected respiratory droplets or secretions, as suggested by the aforementioned John-son and Goodpasture study, and in a subsequent study by Henle and colleagues, who transmitted the disease to children by both oral and nasal routes of inoculation [37]. MuV has been isolated from children with respiratory disease without parotitis [38] and has been detected by RT–PCR in nasal samples [39]. The spread of mumps among persons in close contact also suggests this mode of transmission. Based on studies of other respiratory viruses, it is assumed that, following exposure, MuV infects the upper respiratory tract, but this has not yet been formally demonstrated. For measles virus, a related paramyxovirus, it was also assumed that the virus initially infects the respiratory epithelium, but this assumption proved to be incorrect when macrophages and dendritic cells in the lung tissue were shown to be the primary target cells infected with this virus [40].
Systemic spread: from epitheliotropic to lymphotropic?
Given the array of symptoms, it is clear that MuV is able to disseminate systemically in the body, which has led to the assumption that, following infection of the upper respiratory mucosa, the virus spreads to regional lymph nodes, resulting in viraemia during the early acute phase (Figure 2). However, despite the high frequency of extra-respiratory symptoms, virus has only rarely been detected in blood [41–43], even in experimentally infected animals [44]. One possible explanation is the coincident development of MuV-specific humoral antibodies.
Figure 2.

MuV clinical presentation and pathogenesis. Mumps is a respiratory-spread, acute, inflammatory disease in humans, which causes a range of systemic symptoms. The incubation period is 2–4 weeks. Approximately one-third of infections are asymptomatic. The prodromal phase is characterized by non-specific, often mild symptoms, such as low-grade fever, headache and malaise. An early acute phase follows, likely representing spread of the virus from the respiratory tract and development of systemic symptoms, typically parotitis, which lasts from a few days to 1 week. During the established acute phase, orchitis, meningitis or encephalitis may appear. Symptoms usually resolve within 2 weeks, coincident with the development of a MuV-specific humoral response. Long-term complications and death are rare
Approximately one-third to one-half of MuV infections are asymptomatic or result in only mild respiratory symptoms, sometimes accompanied by fever [45–49]. The hallmark of mumps is salivary gland swelling, typically the parotid glands, which forms the basis of a clinical diagnosis. Parotitis is usually bilateral, developing 2–3 weeks after exposure and lasting for 2–3 days, but it may persist for a week or more in some cases [37,50,51]. Submaxillary, submandibular and sublingual glands can be involved, but rarely as the only manifestation of mumps. Viral replication in the parotid gland results in perivascular and interstitial mononuclear cell infiltration, haemorrhage, oedema and necrosis of acinar and epithelial duct cells [52]. Serum and urine amylase levels may be elevated as a result of inflammation and tissue damage in the parotid gland [53]. Virus is excreted in the saliva from approximately 1 week before to 1 week after the onset of salivary gland swelling [37,54,55]. MuV has also been identified in the saliva of asymptomatic persons [37]. Coupled with excretion of virus up to 1 week before symptom appearance, this may explain some of the difficulties in controlling mumps outbreaks.
Orchitis, which is typically unilateral, is the most common extra-salivary gland manifestation of mumps. It occurs in approximately 10–20% of infections in post-pubertal men [46,56,57]. MuV has been recovered from semen and the testis, suggesting that epididymo-orchitis is the result of direct infection of testicular cells [58,59]. However, an indirect immune-mediated mechanism has also been postulated [60]. Both Leydig and germ cells are involved, associated with reduced levels of testosterone production [61–63]. Necrosis of acinar and epithelial duct cells is evident in the germinal epithelium of the seminiferous tubules of the testes. Orchitis is almost always accompanied by epididymitis and fever, all resolving within 1 week. Atrophy of the involved testicle occurs in approximately half of cases and can be associated with oligospermia and hypofertility, but rarely sterility [58,62,64,65]. Mastitis and oophoritis (manifesting as pelvic pain) occurs in 5–10% of mumps cases in postpubertal women [12,46,66]. Oophoritis has been associated with infertility [67] and premature menopause [68], but such cases are extremely rare.
Virus frequently disseminates to the kidneys, as suggested by the frequency of viruria during the established acute phase of the disease (Figure 2) [69,70]. Epithelial cells of the distal tubules, calyces and ureters appear to be primary sites of virus replication [52]. Kidney involvement in mumps is almost always benign, but cases of severe interstitial nephritis have been reported. In such cases, renal biopsy or postmortem necropsy show evidence of immune complex deposition, interstitial mononuclear cell infiltration and fibrosis, oedema and focal tubular epithelial cell damage [71–73]. Pancreatitis, diagnosed as severe epigastric pain and tenderness, has been reported in approximately 4% of cases [6,12,74]. There are conflicting reports on the association between mumps pancreatitis and diabetes mellitus [75–79] and it is unclear whether there is a causal link.
CNS involvement: from lymphotropic to neurotropic
MuV is highly neurotropic, with evidence of central nervous system (CNS) involvement in up to half of all cases of infection, based on pleocytosis of the cerebrospinal fluid [48,80–82]. Symptomatic CNS infection is less common, but significant. Meningitis occurs in approximately 5–10% of cases and encephalitis in <0.5%. Although these are small percentages, MuV was the leading cause of encephalitis in the USA until 1975, when mumps-containing vaccine gained widespread use [83]. In unvaccinated populations, mumps continues to account for a high percentage of viral encephalitis cases [46,84,85]. Little is known of the CNS pathology, since the disease is rarely fatal. Of the few postmortem cases examined, the pathology includes oedema and congestion throughout the brain with haemorrhage, lymphocytic perivascular infiltration, perivascular glio-sis and demyelination, with relative sparing of neurons. These latter observations suggest that in some cases of mumps encephalitis the inflammation stems from a para-infectious process. However, virus can be recovered from CSF early in the course of meningitis [86,87], as well as from brain tissue in some cases of mumps encephalitis.
Experimental infection in rodents suggests the virus enters the CSF through the choroid plexus, or possibly via transiting mononuclear cells during viraemia. Based on animal data, once in the CSF, virus appears to be carried throughout the ventricular system, resulting in virus replication within ependymal cells that line the ventricles (Figure 3A) [44]. From these locations, virus can penetrate into the brain parenchyma, often infecting pyramidal cells in the cerebral cortex and hippocampus [88]. The infected ependymal epithelia become inflamed, lose their cilia, degenerate and collapse into the CSF (Figure 3B), a postulated cause of the aqueductal stenosis that is believed to be responsible for the occurrence of hydrocephalus, typically of the lateral and third ventricles, a common outcome in intracerebrally injected animals [89–93]. Mumps hydrocephalus has been reported in humans, most often presenting as obstruction of the cerebral aqueduct with dilatation of the lateral and third ventricles. However, obstruction of the foramen of Monro between the lateral and third ventricles, or obstruction of the foramina of Magendie and Luschka between the fourth ventricle and the sub-arachnoid space, have also been reported [94–98]. The finding of ependymal cell debris in the CSF of mumps patients [94,96,99] suggests that the pathogenesis of hydrocephalus in experimentally infected animals is similar to the mechanism of hydrocephalus in humans. However, hydrocephalus has been observed before, or in the total absence of, canal obstruction [88,91,100,101], indicating that such events could be a secondary consequence of external compression by surrounding oedematous tissue and not causally related to the pathogenesis of hydrocephalus.
Figure 3.
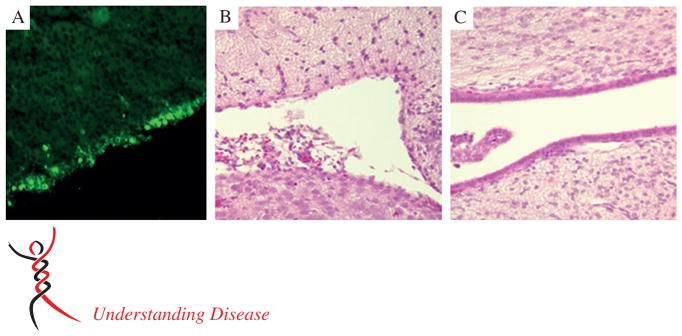
MuV infection of the rat brain. The most prominent neuropathological outcome following MuV intracranial inoculation in small animal models (hamsters, rats) is enlargement of the lateral and third ventricles, ie hydrocephalus, which has also been reported in cases in humans. The cause of hydrocephalus is postulated to be denuding of virus-infected ependymal cells lining the ventricles. (A) Sagittal section of rat brain tissue immunohistochemically stained for the MuV nucleoprotein, showing extensive infection of the ventricular ependymal cells (green foci). (B) Approximately 3 weeks later, ependymal cell loss is evident in comparison to the well-preserved ependymal cell architecture in rats injected with the Jeryl Lynn vaccine strain (C); haematoxylin and eosin (H&E) stain
Deafness has been reported in approximately 4% of mumps cases and is the most frequent cause of acquired unilateral sensorineural hearing loss in children. Hearing loss is typically unilateral and transient, but can be permanent [102–106]. Pathological findings include lesions and degeneration of the stria vascularis, tectorial membrane and organ of Corti [107,108]. MuV infection of the CSF has been implicated in the pathogenesis of deafness in mumps, given the detection of the virus in perilymph, which freely communicates with the CSF [109]. This is also supported by animal studies, where instillation of the virus into the CSF has resulted in infection of the cochlea [110]. However, deafness does not occur any more frequently in patients with meningitis or encephalitis than it does in patients lacking signs of CNS infection, suggesting that CSF may in fact not be involved in the pathogenesis of deafness. An alternative explanation could be that virus infects the inner ear via a haematogenous route, ie that mumps labyrinthitis occurs as a consequence of viraemia. This is supported by studies in guinea pigs following intravascular inoculation of the virus [111], and clinical findings by Lindsay [107] and Mizushima and Murakami [112] suggesting ’viral endolymphatic labyrinthitis’ in the pathogenesis of mumps deafness in humans. Hearing loss caused by indirect effects of virus infection (eg immune-mediated damage) have also been suggested [113]. MuV was also identified in the vestibular ganglia in experimentally infected animals [110], which likely also occurs in humans and explains vestibular symptoms, such as vertigo, which often present in cases of mumps deafness [114,115].
Based on electrocardiographic abnormalities in mumps patients, MuV likely infects cardiac tissue [116]. While this is rarely symptomatic, interstitial lymphocytic myocarditis and pericarditis have been reported [117], which can lead to endocardial fibroelastosis [118]. MuV has also been identified in cardiac muscle from patients with these disorders. Clinically apparent cardiac complications are rare, but can be serious [116,119,120].
Other rare complications include cerebellar ataxia [121,122], transverse myelitis [123,124], ascending polyradiculitis [125], a poliomyelitis-like disease [126,127], arthropathy [128,129], autoimmune haemolytic anaemia [130,131], thyroiditis [132,133], thrombocytopenia [134,135], hepatitis [136,137] and retinitis and corneal endotheliitis [138–140].
Transplacental transmission of the virus has been demonstrated in non-human primates [141] and is suggested by the isolation of virus from the human fetus following spontaneous or planned abortion during maternal mumps [142–144]. Aborted fetal tissue from such cases has been found to exhibit a proliferative necrotizing villitis with decidual cells containing intracytoplasmic inclusions [145]. Virus has also been isolated at birth from infants born to women with mumps [146] and from breast milk [147], but few cases of perinatal mumps have been described [146,148] and it is not clear whether breast milk was responsible for these cases. Mumps virus does not appear to cause congenital malformations [149]. The major morbidity from mumps is from complications of meningitis, encephalitis and orchitis. The case fatality ratio is 1.6–3.8/10,000 [150,151], with most fatalities occurring in persons with encephalitis.
Animal models and molecular determinants of MuV pathogenesis
Historically, the most widely used animal models of mumps have been the hamster and the monkey, and information from these models serves as the basis for much of our current understanding of MuV pathogenesis and disease. However, the relevance of findings in these models for humans is questionable, given the use of unnatural routes of inoculation (eg intracranial, intraperitoneal or intravenous) and the inability of these models to clearly and reliably discriminate strains that are attenuated for humans from wild-type, virulent strains [152–158]. In the one study where monkeys (rhesus macaques) were inoculated via natural routes (intranasal and intratracheal), only wild-type virus was tested [159]. Nonetheless, this study demonstrated the potential to identify sites of early and late MuV replication, which supports further evaluation of this model. Mice and ferrets were also explored as model systems; however, virus replication in these species is self-limiting, making them poor candidates for pathogenesis studies [159–164].
A key advance in the study of MuV pathogenesis was the development of a rat model predictive of the neurovirulence potential of MuV strains for humans. In this model, 1 month after intracerebral injection of virus into newborn Lewis rats, brains are removed and evaluated for virus-induced hydrocephalus (Figure 4), the severity of which correlates with the neurovirulence potential of the virus for humans [165]. With the advent of plasmid-based reverse genetics systems for MuV, it became possible to examine molecular determinants of virulence, and thus gain a better understanding of virus factors that influence pathogenicity. The first such study was published by Lemon et al [166], who generated Jeryl Lynn vaccine strain-based viruses expressing genes derived from the Kilham MuV strain, a hamster brain-adapted laboratory strain. Of the single gene replacements assessed (M, F, SH and HN), only the F gene was found to significantly increase the neurovirulence potential of the highly attenuated Jeryl Lynn strain [166]. However, in a subsequent study using a different virulent MuV strain, 88-1961, Sauder et al [167] found the F gene to have no biologically meaningful effect on the neurovirulence potential of Jeryl Lynn. MuV strain-specific molecular determinants of virulence are also highlighted in other studies. For example, Xu et al [168] identified the SH gene as a virulence factor for the wild-type IA MuV strain, whereas Malik et al [32] found no such role for the SH gene in 88-1961 virulence. Of additional interest, in the Sauder et al study, the Jeryl Lynn genes found to neuro-attenuate the 88-1961 strain (eg M, HN and L), when derived from the 88-1961 strain, did not meaningfully increase the neurovirulence of the Jeryl Lynn virus. Thus, not only do genes that influence virulence of one strain often not affect virulence of another strain, but genes involved in neurovirulence are not necessarily involved in neuroattenuation. Taken together, these results raise doubt as to the prospect of identifying broadly applicable genetic determinants of virulence and attenuation. An alternative, or complementary, approach may be to examine differences between virulent and attenuated strains in terms of sites of primary replication and spread, following infection via natural routes in disease-relevant animal models. To this end, new in vitro and in vivo model systems that recapitulate the diverse features of a natural infection are needed.
Figure 4.
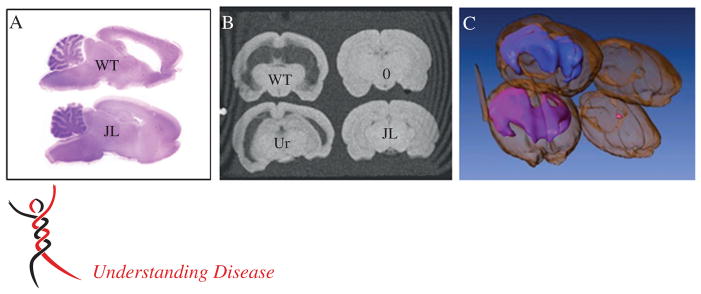
Hydrocephalus severity in MuV-infected rats. (A) H&E-stained sagittal sections of brain from a representative 30 day-old rat injected with a wild-type (WT) MuV isolate as a newborn (top), compared to a rat similarly injected with the highly attenuated Jeryl Lynn (JL) vaccine strain (bottom). (B) T1 weighted gradient-echo image from MRI of the same brains as in (A) (upper left corner and lower right corner), compared to brain from a rat injected with an insufficiently attenuated vaccine strain, Urabe-AM9 (Ur, lower left corner) and an uninfected rat brain (0, upper right corner). Note that the severity of hydrocephalus tracks with the virus strain’s neurovirulence potential for humans. (C) Assembled three-dimensional rendering of MRI slices represented in (B), showing ventricular volume (blue, wild-type; pink, Urabe-AM9)
Conclusion
As a re-emerging pathogen, with concerns over vaccine safety and efficacy, elucidation of mechanisms of MuV pathogenesis is of paramount importance, as this information will help direct the development of improved vaccines. The utilization of existing reverse genetic systems alongside the generation of new, clinically relevant systems and the development of robust animal models for other aspects of the disease will allow a more complete understanding of disease. This review has summarized our current understanding of MuV clinical disease, pathology, and how this relates to viral pathogenesis. However, a number of areas are evidently poorly understood and important questions remain (Figure 5). These include the elucidation of the target cell tropism throughout an infection, the mechanisms by which MuV establishes a systemic infection and the basis of neurotropism. Determination of these likely requires the establishment of a primate model of MuV infection, similar to what has been achieved with measles. The measles system, exploiting a fluorescent reporter-expressing wild-type MV in a clinically-relevant macaque (Macaca fascicularis) model has facilitated the elucidation of key features of measles virus pathogenesis relevant to disease, transmission and immunity [40]. Applying what has been learned from the measles model to mumps will provide an ideal basis to understanding determinants of MuV pathogenesis.
Figure 5.

A number of important questions remain unresolved regarding MuV pathogenesis. This is of particular relevance to renewed efforts towards development of a more efficacious MuV vaccine, in light of the resurgence of mumps in vaccinated populations. The classic method of virus attenuation is extensive blind passage in vitro. While this often leads to the desired effect of a loss of virulence and reactogenicity, it can also lead to loss of immunogenicity and efficacy. Clearly, a more rational approach to virus attenuation is needed, and understanding the natural pathogenesis of the infectious agent is a prerequisite to any such endeavour. This figure highlights this issue, showing our current assumptions of pathogenesis (black text) and unresolved questions (red text)
Acknowledgments
This work was supported by the NIH R01 AI105063 to WPD and by the Oak Ridge Institute for Science and Education through an interagency agreement between the US Department of Energy and the US Food and Drug Administration. We wish to thank Christian Sauder and Laurie Ngo (FDA, Silver Spring, MD) for critical reading of the manuscript, and Jan Johannessen (FDA, Silver Spring, MD) for providing the MRI rat brain images. We dedicate this work to the memory of the late Dr Philip Snoy, DVM, Director, CBER Veterinary Services, respected scientist, admired leader and treasured friend, who laid much of the groundwork towards development of improved preclinical MuV neurotoxicity tests.
Footnotes
No conflicts of interest were declared.
References
- 1.Wagenvoort JH, Harmsen M, Boutahar-Trouw BJ, et al. Epidemiology of mumps in The Netherlands. J Hyg (Lond) 1980;85:313–326. doi: 10.1017/s0022172400063385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Anderson RM, Crombie JA, Grenfell BT. The epidemiology of mumps in the UK: a preliminary study of virus transmission, herd immunity and the potential impact of immunization. Epidemiol Infect. 1987;99:65–84. doi: 10.1017/s0950268800066875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Cox MJ, Anderson RM, Bundy DA, et al. Seroepidemiological study of the transmission of the mumps virus in St. Lucia, West Indies. Epidemiol Infect. 1989;102:147–160. doi: 10.1017/s0950268800029770. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Centers for Disease Control and Prevention. Current trends mumps – United States, 1984–1985. Morb Mortal Wkly Rep. 1986;35:216–219. [PubMed] [Google Scholar]
- 5.McNabb SJ, Jajosky RA, Hall-Baker PA, et al. Summary of notifiable diseases – United States, 2005. Morb Mortal Wkly Rep. 2007;54:1–92. [PubMed] [Google Scholar]
- 6.Boxall N, Kubinyiova M, Prikazsky V, et al. An increase in the number of mumps cases in the Czech Republic, 2005–2006. Euro-surveillance. 2008;13:Article 8. [PubMed] [Google Scholar]
- 7.Bangor-Jones RD, Dowse GK, Giele CM, et al. A prolonged mumps outbreak among highly vaccinated Aboriginal people in the Kimberley region of Western Australia. Med J Aust. 2009;191:398–401. doi: 10.5694/j.1326-5377.2009.tb02850.x. [DOI] [PubMed] [Google Scholar]
- 8.Park DW, Nam MH, Kim JY, et al. Mumps outbreak in a highly vaccinated school population: assessment of secondary vaccine failure using IgG avidity measurements. Vaccine. 2007;25:4665–4670. doi: 10.1016/j.vaccine.2007.04.013. [DOI] [PubMed] [Google Scholar]
- 9.Centers for Disease Control and Prevention. Mumps epidemic – United Kingdom, 2004–2005. Morb Mortal Wkly Rep. 2006;55:173–175. [PubMed] [Google Scholar]
- 10.Watson-Creed G, Saunders A, Scott J, et al. Two successive outbreaks of mumps in Nova Scotia among vaccinated adolescents and young adults. Can Med Assoc J. 2006;175:483–488. doi: 10.1503/cmaj.060660. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Schmid D, Pichler AM, Wallenko H, et al. Mumps outbreak affecting adolescents and young adults in Austria, 2006. Eurosurveillence. 2006;11:E060615. [PubMed] [Google Scholar]
- 12.Dayan GH, Quinlisk MP, Parker AA, et al. Recent resurgence of mumps in the United States. N Engl J Med. 2008;358:1580–1589. doi: 10.1056/NEJMoa0706589. [DOI] [PubMed] [Google Scholar]
- 13.Sartorius B, Penttinen P, Nilsson J, et al. An outbreak of mumps in Sweden, February–April 2004. Eurosurveillence. 2005;10:191–193. [PubMed] [Google Scholar]
- 14.Vandermeulen C, Roelants M, Vermoere M, et al. Outbreak of mumps in a vaccinated child population: a question of vaccine failure? Vaccine. 2004;22:2713–2716. doi: 10.1016/j.vaccine.2004.02.001. [DOI] [PubMed] [Google Scholar]
- 15.Castilla J, Garcia CM, Barricarte A, et al. Mumps outbreak in Navarre region, Spain, 2006–2007. Eurosurveillence. 2007;12:E070215. doi: 10.2807/esw.12.07.03139-en. [DOI] [PubMed] [Google Scholar]
- 16.Centers for Disease Control and Prevention. Brief report: update: mumps activity –United States, January 1–October 7, 2006. Morb Mortal Wkly Rep. 2006;55:1152–1153. [PubMed] [Google Scholar]
- 17.Cortese MM, Jordan HT, Curns AT, et al. Mumps vaccine performance among university students during a mumps outbreak. Clin Infect Dis. 2008;46:1172–1180. doi: 10.1086/529141. [DOI] [PubMed] [Google Scholar]
- 18.Marin M, Quinlisk P, Shimabukuro T, et al. Mumps vaccination coverage and vaccine effectiveness in a large outbreak among college students – Iowa, 2006. Vaccine. 2008;26:3601–3607. doi: 10.1016/j.vaccine.2008.04.075. [DOI] [PubMed] [Google Scholar]
- 19.Roberts C, Porter-Jones G, Crocker J, et al. Mumps outbreak on the island of Anglesey, North Wales, December 2008–January 2009. Eurosurveillence. 2009;14:Article 1. doi: 10.2807/ese.14.05.19109-en. [DOI] [PubMed] [Google Scholar]
- 20.Rota JS, Turner JC, Yost-Daljev MK, et al. Investigation of a mumps outbreak among university students with two measles–mumps–rubella (MMR) vaccinations, Virginia, September–December 2006. J Med Virol. 2009;81:1819–1825. doi: 10.1002/jmv.21557. [DOI] [PubMed] [Google Scholar]
- 21.Nagai T, Okafuji T, Miyazaki C, et al. A comparative study of the incidence of aseptic meningitis in symptomatic natural mumps patients and monovalent mumps vaccine recipients in Japan. Vaccine. 2007;25:2742–2747. doi: 10.1016/j.vaccine.2005.11.068. [DOI] [PubMed] [Google Scholar]
- 22.Baba K, Okuno Y, Tanaka-Taya K, et al. Immunization coverage and natural infection rates of vaccine-preventable diseases among children by questionnaire survey in 2005 in Japan. Vaccine. 2011;29:3089–92. doi: 10.1016/j.vaccine.2010.09.022. [DOI] [PubMed] [Google Scholar]
- 23.Litman N, Baum SG. Mumps Virus. In: Mandell GL, Bennett JE, Donlin R, editors. Principles and Practice of Infectious Diseases. 7. Churchill Livingstone; Philadelphia, PA: 2010. pp. 2201–2206. [Google Scholar]
- 24.Johnson CD, Goodpasture EW. An investigation of the etiology of mumps. J Exp Med. 1934;59:1–19. doi: 10.1084/jem.59.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Johnson CD, Goodpasture EW. The etiology of mumps. Am J Hyg. 1935;21:46–57. [Google Scholar]
- 26.Elango N, Varsanyi TM, Kovamees J, et al. Molecular cloning and characterization of six genes, determination of gene order and intergenic sequences and leader sequence of mumps virus. J Gen Virol. 1988;69:2893–2900. doi: 10.1099/0022-1317-69-11-2893. [DOI] [PubMed] [Google Scholar]
- 27.Elliott GD, Yeo RP, Afzal MA, et al. Strain-variable editing during transcription of the P gene of mumps virus may lead to the generation of non-structural proteins NS1 (V) and NS2. J Gen Virol. 1990;71:1555–1560. doi: 10.1099/0022-1317-71-7-1555. [DOI] [PubMed] [Google Scholar]
- 28.Li M, Schmitt PT, Li Z, et al. Mumps virus matrix, fusion, and nucleocapsid proteins cooperate for efficient production of virus-like particles. J Virol. 2009;83:7261–7272. doi: 10.1128/JVI.00421-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Harrison MS, Sakaguchi T, Schmitt AP. Paramyxovirus assembly and budding: building particles that transmit infections. Int J Biochem Cell Biol. 2010;42:1416–1429. doi: 10.1016/j.biocel.2010.04.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.He B, Leser GP, Paterson RG, et al. The paramyxovirus SV5 small hydrophobic (SH) protein is not essential for virus growth in tissue culture cells. Virology. 1998;250:30–40. doi: 10.1006/viro.1998.9354. [DOI] [PubMed] [Google Scholar]
- 31.He B, Lin GY, Durbin JE, et al. The SH integral membrane protein of the paramyxovirus simian virus 5 is required to block apoptosis in MDBK cells. J Virol. 2001;75:4068–4079. doi: 10.1128/JVI.75.9.4068-4079.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Malik T, Shegogue CW, Werner K, et al. Discrimination of mumps virus small hydrophobic gene deletion effects from gene translation effects on virus virulence. J Virol. 2011;85:6082–6085. doi: 10.1128/JVI.02686-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Paterson RG, Lamb RA. RNA editing by G-nucleotide insertion in mumps virus P-gene mRNA transcripts. J Virol. 1990;64:4137–4145. doi: 10.1128/jvi.64.9.4137-4145.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Andrejeva J, Childs KS, Young DF, et al. The V proteins of paramyxoviruses bind the IFN-inducible RNA helicase, mda-5, and inhibit its activation of the IFN-β promoter. Proc Natl Acad Sci USA. 2004;101:17264–17269. doi: 10.1073/pnas.0407639101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Puri M, Lemon K, Duprex WP, et al. A point mutation, E95D, in the mumps virus V protein disengages STAT3 targeting from STAT1 targeting. J Virol. 2009;83:6347–6356. doi: 10.1128/JVI.00596-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Kubota T, Yokosawa N, Yokota S, et al. C terminal CYS-RICH region of mumps virus structural V protein correlates with block of interferon-α and -γ signal transduction pathway through decrease of STAT 1-α. Biochem Biophys Res Commun. 2001;283:255–259. doi: 10.1006/bbrc.2001.4764. [DOI] [PubMed] [Google Scholar]
- 37.Henle G, Henle W, Wendell K, et al. Isolation of mumps virus from human being with induced apparent or inapparent infections. J Exp Med. 1948;88:223–232. doi: 10.1084/jem.88.2.223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Foy HM, Cooney MK, Hall CE, et al. Isolation of mumps virus from children with acute lower respiratory tract disease. Am J Epidemiol. 1971;94:467–472. doi: 10.1093/oxfordjournals.aje.a121343. [DOI] [PubMed] [Google Scholar]
- 39.Zajac BA, Henle W, Henle G. The effect of mumps virus on the resistance of Burkitt lymphoma cell lines to various viruses. Cancer Res. 1969;29:481–488. [PubMed] [Google Scholar]
- 40.Lemon K, de Vries RD, Mesman AW, et al. Early target cells of measles virus after aerosol infection of non-human primates. PLoS Pathog. 2011;7:e1001263. doi: 10.1371/journal.ppat.1001263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Kilham L. Isolation of mumps virus from the blood of a patient. Proc Soc Exp Biol Med. 1948;69:99–100. doi: 10.3181/00379727-69-16629. [DOI] [PubMed] [Google Scholar]
- 42.Overman JR. Viremia in human mumps virus infections. Arch Intern Med. 1958;102:354–356. doi: 10.1001/archinte.1958.00030010354002. [DOI] [PubMed] [Google Scholar]
- 43.Mishra B, Pujhari SK, Dhiman V, et al. Genotyping and subtyping of mumps virus isolates from the Indian subcontinent. Arch Virol. 2013;158:2359–2363. doi: 10.1007/s00705-013-1717-4. [DOI] [PubMed] [Google Scholar]
- 44.Wolinsky JS, Klassen T, Baringer JR. Persistance of neuroadapted mumps virus in brains of newborn hamsters after intraperitoneal inoculation. J Infect Dis. 1976;133:260–267. doi: 10.1093/infdis/133.3.260. [DOI] [PubMed] [Google Scholar]
- 45.Reed D, Brown G, Merrick R, et al. A mumps epidemic on St. George Island, Alaska. J Am Med Assoc. 1967;199:113–117. doi: 10.1001/jama.199.13.113. [DOI] [PubMed] [Google Scholar]
- 46.Philip RN, Reinhard KP, Lachman DB. Observations on a mumps epidemic in a ‘virgin’ population. Am J Hyg. 1959;69:91–111. doi: 10.1093/oxfordjournals.aje.a119992. [DOI] [PubMed] [Google Scholar]
- 47.Brunell PA, Brickman A, O’Hare D, et al. Ineffectiveness of isolation of patients as a method of preventing the spread of mumps. Failure of the mumps skin-test antigen to predict immune status. N Engl J Med. 1968;279:1357–1361. doi: 10.1056/NEJM196812192792502. [DOI] [PubMed] [Google Scholar]
- 48.Falk WA, Buchan K, Dow M, et al. The epidemiology of mumps in southern Alberta, 1980–1982. Am J Epidemiol. 1989;130:736–749. doi: 10.1093/oxfordjournals.aje.a115395. [DOI] [PubMed] [Google Scholar]
- 49.Cooney MK, Fox JP, Hall CE. The Seattle Virus Watch. VI. Observations of infections with and illness due to parainfluenza, mumps and respiratory syncytial viruses and Mycoplasma pneumoniae. Am J Epidemiol. 1975;101:532–551. doi: 10.1093/oxfordjournals.aje.a112125. [DOI] [PubMed] [Google Scholar]
- 50.Hope-Simpson RE. Infectiousness of comminicable diseases in the household (measles, chickenpox, and mumps) Lancet. 1952;2:549–554. doi: 10.1016/s0140-6736(52)91357-3. [DOI] [PubMed] [Google Scholar]
- 51.Henle G, Enders JF. Mumps virus. In: Horstfall F, Tamm I, editors. Viral and Rickettsial Infections of Man. Lippincott; Philadelphia, PA: 1965. pp. 755–768. [Google Scholar]
- 52.Weller TH, Craig JR. Isolation of mumps virus at autopsy. Am J Pathol. 1949;25:1105–1115. [PMC free article] [PubMed] [Google Scholar]
- 53.Scully C, Eckersall PD, Emond RT, et al. Serum α-amylase isozymes in mumps: estimation of salivary and pancreatic isozymes by isoelectric focusing. Clin Chim Acta. 1981;113:281–291. doi: 10.1016/0009-8981(81)90281-3. [DOI] [PubMed] [Google Scholar]
- 54.Chiba Y, Horino K, Umetsu M, et al. Virus excretion and antibody response in saliva in natural mumps. Tohoku J Exp Med. 1973;111:229–238. doi: 10.1620/tjem.111.229. [DOI] [PubMed] [Google Scholar]
- 55.Ennis FA, Jackson D. Isolation of virus during the incubation period of mumps infection. J Pediatr. 1968;72:536–537. doi: 10.1016/s0022-3476(68)80347-6. [DOI] [PubMed] [Google Scholar]
- 56.Postovit VA. Epidemic parotitis in adults. Voen Med Zh. 1983;3:38–41. [PubMed] [Google Scholar]
- 57.McGuiness AC, Gall EA. Mumps at army camps in 1943. War Med. 1944;5:95–104. [Google Scholar]
- 58.Bjorvatn B. Mumps virus recovered from testicles by fine-needle aspiration biopsy in cases of mumps orchitis. Scand J Infect Dis. 1973;5:3–5. doi: 10.3109/inf.1973.5.issue-1.02. [DOI] [PubMed] [Google Scholar]
- 59.Jalal H, Bahadur G, Knowles W, et al. Mumps epididymo-orchitis with prolonged detection of virus in semen and the development of anti-sperm antibodies. J Med Virol. 2004;73:147–150. doi: 10.1002/jmv.10544. [DOI] [PubMed] [Google Scholar]
- 60.Clifford V, Wadsley J, Jenner B, et al. Mumps vaccine associated orchitis: evidence supporting a potential immune-mediated mechanism. Vaccine. 2010;28:2671–2673. doi: 10.1016/j.vaccine.2010.01.007. [DOI] [PubMed] [Google Scholar]
- 61.Ternavasio-de la Vega HG, Boronat M, Ojeda A, et al. Mumps orchitis in the post-vaccine era (1967–2009): a single-center series of 67 patients and review of clinical outcome and trends. Medicine (Baltimore) 2010;89:96–116. doi: 10.1097/MD.0b013e3181d63191. [DOI] [PubMed] [Google Scholar]
- 62.Dejucq N, Jegou B. Viruses in the mammalian male genital tract and their effects on the reproductive system. Microbiol Mol Biol Rev. 2001;65:208–231. doi: 10.1128/MMBR.65.2.208-231.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Le GR, Mouchel T, Ruffault A, et al. Mumps virus decreases testosterone production and γ-interferon-induced protein 10 secretion by human Leydig cells. J Virol. 2003;77:3297–3300. doi: 10.1128/JVI.77.5.3297-3300.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Candel S. Epididymitis in mumps, including orchitis: further clinical studies and comment. Ann Intern Med. 1951;34:20. doi: 10.7326/0003-4819-34-1-20. [DOI] [PubMed] [Google Scholar]
- 65.Shulman A, Shohat B, Gillis D, et al. Mumps orchitis among soldiers: frequency, effect on sperm quality, and sperm antibodies. Fertil Steril. 1992;57:1344–1346. doi: 10.1016/s0015-0282(16)55099-7. [DOI] [PubMed] [Google Scholar]
- 66.Maynard JE, Shramek G, Noble GR, et al. Use of attenuated live mumps virus vaccine during a ‘virgin soil’ epidemic of mumps on St. Paul Island, Alaska. Am J Epidemiol. 1970;92:301–306. doi: 10.1093/oxfordjournals.aje.a121211. [DOI] [PubMed] [Google Scholar]
- 67.Prinz W, Taubert HD. Mumps in pubescent females and its effect on later reproductive function. Gynaecologia. 1969;167:23–27. doi: 10.1159/000302156. [DOI] [PubMed] [Google Scholar]
- 68.Morrison JC, Givens JR, Wiser WL, et al. Mumps oophoritis: a cause of premature menopause. Fertil Steril. 1975;26:655–659. [PubMed] [Google Scholar]
- 69.Utz JP, Szwed CF. Mumps III. Comparison of methods for detection of viruria. Proc Soc Exp Biol Med. 1962;110:841–844. doi: 10.3181/00379727-110-27666. [DOI] [PubMed] [Google Scholar]
- 70.Utz JP, Houk VN, Alling DW. Clinical and laboratory studies of mumps. IV. Viruria and abnormal renal function. N Engl J Med. 1964;270:1283–1286. doi: 10.1056/NEJM196406112702404. [DOI] [PubMed] [Google Scholar]
- 71.Lin CY, Chen WP, Chiang H. Mumps associated with nephritis. Child Nephrol Urol. 1990;10:68–71. [PubMed] [Google Scholar]
- 72.Kabakus N, Aydinoglu H, Bakkaloglu SA, et al. Mumps interstitial nephritis: a case report. Pediatr Nephrol. 1999;13:930–931. doi: 10.1007/s004670050731. [DOI] [PubMed] [Google Scholar]
- 73.Hughes WT, Steigman AJ, Delong HF. Some implications of fatal nephritis associated with mumps. Am J Dis Child. 1966;111:297–301. doi: 10.1001/archpedi.1966.02090060107013. [DOI] [PubMed] [Google Scholar]
- 74.Nishino H, Engel AG, Rima BK. Inclusion body myositis: the mumps virus hypothesis. Ann Neurol. 1989;25:260–264. doi: 10.1002/ana.410250309. [DOI] [PubMed] [Google Scholar]
- 75.Dacou-Voutetakis C, Constantinidis M, Moschos A, et al. Diabetes mellitus following mumps. Insulin reserve. Am J Dis Child. 1974;127:890–891. doi: 10.1001/archpedi.1974.02110250116018. [DOI] [PubMed] [Google Scholar]
- 76.Hyoty H, Leinikki P, Reunanen A, et al. Mumps infections in the etiology of type 1 (insulin-dependent) diabetes. Diabetes Res. 1988;9:111–116. [PubMed] [Google Scholar]
- 77.Sultz HA, Hart BA, Zielezny M, et al. Is mumps virus an etiologic factor in juvenile diabetes mellitus. J Pediatr. 1975;86:654–656. doi: 10.1016/s0022-3476(75)80206-x. [DOI] [PubMed] [Google Scholar]
- 78.Otten A, Helmke K, Stief T, et al. Mumps, mumps vaccination, islet cell antibodies and the first manifestation of diabetes mellitus type I. Behring Inst Mitt. 1984;75:83–88. [PubMed] [Google Scholar]
- 79.Goto A, Takahashi Y, Kishimoto M, et al. A case of fulminant type 1 diabetes associated with significant elevation of mumps titers. Endocr J. 2008;55:561–564. doi: 10.1507/endocrj.k07e-126. [DOI] [PubMed] [Google Scholar]
- 80.Bang HO, Bang J. Involvement of the central nervous system in mumps. Acta Med Scand. 1943;113:487–505. [Google Scholar]
- 81.Brown JW, Kirkland HB, Hein GE. Central nervous system involvement during mumps. Am J Med Sci. 1948;215:434–441. doi: 10.1097/00000441-194804000-00012. [DOI] [PubMed] [Google Scholar]
- 82.Bruyn HB, Sexton HM, Brainerd HD. Mumps meningoencephalitis. A clinical review of 119 cases with one death. Calif Med. 1957;86:153–160. [PMC free article] [PubMed] [Google Scholar]
- 83.Centers for Disease Control and Prevention. Measles, mumps, and rubella – vaccine use and strategies for the elimination of measles, rubella, and congenital rubella syndrome and control of mumps: recommendations of the Advisory Committee on Immunization Practices (ACIP) Morbid Mortal Weekly Rep. 1998;47:1–48. [PubMed] [Google Scholar]
- 84.Ponka A, Pettersson T. The incidence and aetiology of central nervous system infections in Helsinki in 1980. Acta Neurol Scand. 1982;66:529–535. doi: 10.1111/j.1600-0404.1982.tb03139.x. [DOI] [PubMed] [Google Scholar]
- 85.Mallewa M, Vallely P, Faragher B, et al. Viral CNS infections in children from a malaria-endemic area of Malawi: a prospective cohort study. Lancet Glob Health. 2013;1:e153–160. doi: 10.1016/S2214-109X(13)70060-3. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Azimi PH, Cramblett HG, Haynes RE. Mumps meningoencephalitis in children. J Am Med Assoc. 1969;207:509–512. [PubMed] [Google Scholar]
- 87.Kilham L. Mumps meningoencephalitis with and without parotitis. Am J Dis Child. 1949;78:324–333. doi: 10.1001/archpedi.1949.02030050337006. [DOI] [PubMed] [Google Scholar]
- 88.Takano T, Mekata Y, Yamano T, et al. Early ependymal changes in experimental hydrocephalus after mumps virus inoculation in hamsters. Acta Neuropathol. 1993;85:521–525. doi: 10.1007/BF00230492. [DOI] [PubMed] [Google Scholar]
- 89.Johnson RT, Johnson KP, Edmonds CJ. Virus-induced hydrocephalus: development of aqueductal stenosis in hamsters after mumps infection. Science. 1967;157:1066–1067. doi: 10.1126/science.157.3792.1066. [DOI] [PubMed] [Google Scholar]
- 90.Johnson RT, Johnson KP. Hydrocephalus following viral infection: the pathology of aqueductal stenosis developing after experimental mumps virus infection. J Neuropathol Exp Neurol. 1968;27:591–606. [PubMed] [Google Scholar]
- 91.Wolinsky JS, Baringer JR, Margolis G, et al. Ultrstructure of mumps virus replication in newborn hamster central nervous system. Lab Invest. 1974;31:403–412. [PubMed] [Google Scholar]
- 92.London WT, Kent SG, Palmer AE, et al. Induction of congenital hydrocephalus with mumps virus in rhesus monkeys. J Infect Dis. 1979;139:324–328. doi: 10.1093/infdis/139.3.324. [DOI] [PubMed] [Google Scholar]
- 93.Rubin SA, Pletnikov M, Carbone KM. Comparison of the neurovirulence of a vaccine and a wild-type mumps virus strain in the developing rat brain. J Virol. 1998;72:8037–8042. doi: 10.1128/jvi.72.10.8037-8042.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Timmons GD, Johnson KP. Aqueductal stenosis and hydrocephalus after mumps encephalitis. N Engl J Med. 1970;283:1505–1507. doi: 10.1056/NEJM197012312832707. [DOI] [PubMed] [Google Scholar]
- 95.Pongener I, Nandan D, Bhatt GC, et al. Hydrocephalus due to aqueductal stenosis in a case of mumps meningoencephalitis: a rare complication. J Pediatr Neurosci. 2013;8:179–180. doi: 10.4103/1817-1745.117871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Ogata H, Oka K, Mitsudome A. Hydrocephalus due to acute aqueductal stenosis following mumps infection: report of a case and review of the literature. Brain Dev. 1992;14:417–419. doi: 10.1016/s0387-7604(12)80352-4. [DOI] [PubMed] [Google Scholar]
- 97.Lahat E, Aladjem M, Schiffer J, et al. Hydrocephalus due to bilateral obstruction of the foramen of Monro: a possible late complication of mumps encephalitis. Clin Neurol Neurosurg. 1993;95:151–154. doi: 10.1016/0303-8467(93)90011-5. [DOI] [PubMed] [Google Scholar]
- 98.Tastan Y, Cam H, Yilmaz I, et al. Reversible acute tetraventricular hydrocephalus complicating possible mumps meningoencephalitis. Pediatr Int. 2007;49:100–102. doi: 10.1111/j.1442-200X.2007.02300.x. [DOI] [PubMed] [Google Scholar]
- 99.Herndon RM, Johnson RT, Davis LE, et al. Ependymitis in mumps virus meningitis. Electron microscopical studies of cerebrospinal fluid. Arch Neurol. 1974;30:475–479. doi: 10.1001/archneur.1974.00490360051010. [DOI] [PubMed] [Google Scholar]
- 100.Johnson KP, Johnson RT. Granular ependymitis. Occurrence in myxovirus infected rodents and prevalence in man. Am J Pathol. 1972;67:511–526. [PMC free article] [PubMed] [Google Scholar]
- 101.Takano T, Takikita S, Shimada M. Experimental mumps virus-induced hydrocephalus: viral neurotropism and neuronal maturity. Neuro Report. 1999;10:2215–2221. doi: 10.1097/00001756-199908020-00001. [DOI] [PubMed] [Google Scholar]
- 102.Hall R, Richards H. Hearing loss due to mumps. Arch Dis Child. 1987;62:189–191. doi: 10.1136/adc.62.2.189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Vuori M, Lahikainen EA, Peltonen T. Perceptive deafness in connection with mumps. A study of 298 servicemen suffering from mumps. Acta Otolaryngol. 1962;55:231–236. doi: 10.3109/00016486209127357. [DOI] [PubMed] [Google Scholar]
- 104.Okamoto M, Shitara T, Nakayama M, et al. Sudden deafness accompanied by asymptomatic mumps. Acta Otolaryngol Suppl. 1994;514:45–48. doi: 10.3109/00016489409127557. [DOI] [PubMed] [Google Scholar]
- 105.Bitnun S, Rakover Y, Rosen G. Acute bilateral total deafness complicating mumps. J Laryngol Otol. 1986;100:943–945. doi: 10.1017/s0022215100100362. [DOI] [PubMed] [Google Scholar]
- 106.Hashimoto H, Fujioka M, Kinumaki H. An office-based prospective study of deafness in mumps. Pediatr Infect Dis J. 2009;28:173–175. doi: 10.1097/INF.0b013e31818a8ca8. [DOI] [PubMed] [Google Scholar]
- 107.Lindsay JR, Davey PR, Ward PH. Inner ear pathology in deafness due to mumps. Ann Otol Rhinol Laryngol. 1960;69:918–935. doi: 10.1177/000348946006900318. [DOI] [PubMed] [Google Scholar]
- 108.Everberg G. Deafness following mumps. Acta Otolaryngol. 1957;48:397–403. doi: 10.3109/00016485709126900. [DOI] [PubMed] [Google Scholar]
- 109.Westmore GA, Pickard BH, Stern H. Isolation of mumps virus from the inner ear after sudden deafness. Br Med J. 1979;1:14–15. doi: 10.1136/bmj.1.6155.14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Davis LE, Johnson RT. Experimental viral infections of the inner ear. I. Acute infections of the newborn hamster labyrinth. Lab Invest. 1976;34:349–356. [PubMed] [Google Scholar]
- 111.Tanaka K, Fukuda S, Suenaga T, et al. Experimental mumps virus-induced labyrinthitis. Immunohistochemical and ultrastructural studies. Acta Otolaryngol Suppl. 1988;456:98–105. doi: 10.3109/00016488809125085. [DOI] [PubMed] [Google Scholar]
- 112.Mizushima N, Murakami Y. Deafness following mumps: the possible pathogenesis and incidence of deafness. Auris Nasus Larynx. 1986;13(suppl 1):S55–57. doi: 10.1016/s0385-8146(86)80035-9. [DOI] [PubMed] [Google Scholar]
- 113.Unal M, Katircioglu S, Karatay MC, et al. Sudden total bilateral deafness due to asymptomatic mumps infection. Int J Pediatr Otorhinolaryngol. 1998;45:167–169. doi: 10.1016/s0165-5876(98)00086-x. [DOI] [PubMed] [Google Scholar]
- 114.Tsubota M, Shojaku H, Ishimaru H, et al. Mumps virus may damage the vestibular nerve as well as the inner ear. Acta Otolaryngol. 2008;128:644–647. doi: 10.1080/00016480701646305. [DOI] [PubMed] [Google Scholar]
- 115.Hyden D, Odkvist LM, Kylen P. Vestibular symptoms in mumps deafness. Acta Otolaryngol Suppl. 1979;360:182–183. doi: 10.3109/00016487809123510. [DOI] [PubMed] [Google Scholar]
- 116.Arita M, Ueno Y, Masuyama Y. Complete heart block in mumps myocarditis. Br Heart J. 1981;46:342–344. doi: 10.1136/hrt.46.3.342. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Brown NJ, Richmond SJ. Fatal mumps myocarditis in an 8-month-old child. Br Med J. 1980;281:356–357. doi: 10.1136/bmj.281.6236.356-a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Ni J, Bowles NE, Kim YH, et al. Viral infection of the myocardium in endocardial fibroelastosis. Molecular evidence for the role of mumps virus as an etiologic agent. Circulation. 1997;95:133–139. doi: 10.1161/01.cir.95.1.133. [DOI] [PubMed] [Google Scholar]
- 119.Roberts WC, Fox SM., III Mumps of the heart. Clinical and pathologic features. Circulation. 1965;32:342–345. doi: 10.1161/01.cir.32.3.342. [DOI] [PubMed] [Google Scholar]
- 120.Chaudary S, Jaski BE. Fulminant mumps myocarditis. Ann Intern Med. 1989;110:569–570. doi: 10.7326/0003-4819-110-7-569. [DOI] [PubMed] [Google Scholar]
- 121.Cohen HA, Ashkenazi A, Nussinovitch M, et al. Mumps-associated acute cerebellar ataxia. Am J Dis Child. 1992;146:930–931. doi: 10.1001/archpedi.1992.02160200052025. [DOI] [PubMed] [Google Scholar]
- 122.Nussinovitch M, Prais D, Volovitz B, et al. Post-infectious acute cerebellar ataxia in children. Clin Pediatr (Philadelphia) 2003;42:581–584. doi: 10.1177/000992280304200702. [DOI] [PubMed] [Google Scholar]
- 123.Bansal R, Kalita J, Misra UK, et al. Myelitis: a rare presentation of mumps. Pediatr Neurosurg. 1998;28:204–206. doi: 10.1159/000028651. [DOI] [PubMed] [Google Scholar]
- 124.Gibbons JL, Miller HG, Stanton JB. Para-infectious encephalomyelitis and related syndromes; a critical review of the neurological complications of certain specific fevers. Q J Med. 1956;25:427–505. [PubMed] [Google Scholar]
- 125.Ghosh S. Guillain–Barre syndrome complicating mumps. Lancet. 1967;1:895. doi: 10.1016/s0140-6736(67)91452-3. [DOI] [PubMed] [Google Scholar]
- 126.Kilham L, Levens J, Enders JF. Nonparalytic poliomyelitis and mumps meningoencephalitis. Differential diagnosis. J Am Med Assoc. 1949;140:934–936. doi: 10.1001/jama.1949.02900460004002. [DOI] [PubMed] [Google Scholar]
- 127.Johnstone JA, Ross CA, Dunn M. Meningitis and encephalitis associated with mumps infection. A 10-year survey. Arch Dis Child. 1972;47:647–651. doi: 10.1136/adc.47.254.647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Gordon SC, Lauter CB. Mumps arthritis: a review of the literature. Rev Infect Dis. 1984;6:338–344. doi: 10.1093/clinids/6.3.338. [DOI] [PubMed] [Google Scholar]
- 129.Harel L, Amir J, Reish O, et al. Mumps arthritis in children. Pediatr Infect Dis J. 1990;9:928–929. doi: 10.1097/00006454-199012000-00016. [DOI] [PubMed] [Google Scholar]
- 130.Mulaosmanovic V, Sakic M, Ferhatovic M. Secondary autoimmune haemolytic anaemia caused by epidemic parotitis virus. Med Arh. 2002;56:46–47. [PubMed] [Google Scholar]
- 131.O’Brien PK, Smith DS, Galpin OP. Acute pancreatitis and haemolytic anaemia associated with mumps-virus infection. Br Med J. 1965;2:1529. doi: 10.1136/bmj.2.5477.1529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Eylan E, Zmucky R, Sheba C. Mumps virus and subacute thyroiditis; evidence of a causal association. Lancet. 1957;272:1062–1063. doi: 10.1016/s0140-6736(57)91438-1. [DOI] [PubMed] [Google Scholar]
- 133.Parmar RC, Bavdekar SB, Sahu DR, et al. Thyroiditis as a presenting feature of mumps. Pediatr Infect Dis J. 2001;20:637–638. doi: 10.1097/00006454-200106000-00023. [DOI] [PubMed] [Google Scholar]
- 134.Famma PG, Paton WB, Bostock MI. Thrombocytopenic purpura complicating mumps. Br Med J. 1964;2:1244. doi: 10.1136/bmj.2.5419.1244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Nasab AR. Thrombocytopenic purpura as a complication of mumps. Clin Pediatr (Philadelphia) 1971;10:420–421. doi: 10.1177/000992287101000712. [DOI] [PubMed] [Google Scholar]
- 136.Brent AJ, Hull R, Jeffery KJ, et al. Acute cholecystitis complicating mumps. Clin Infect Dis. 2006;42:302–303. doi: 10.1086/499107. [DOI] [PubMed] [Google Scholar]
- 137.Warren WR. Hepatitis complicating mumps. Arch Intern Med. 1956;98:525–528. doi: 10.1001/archinte.1956.00250280127017. [DOI] [PubMed] [Google Scholar]
- 138.Ando K, Ishihara M, Kusumoto Y, et al. A case of corneal endotheliitis with mumps virus RNA in aqueous humor detected by RT–PCR. Ocul Immunol Inflamm. 2013;21:150–152. doi: 10.3109/09273948.2012.747619. [DOI] [PubMed] [Google Scholar]
- 139.Khubchandani R, Rane T, Agarwal P, et al. Bilateral neuroretinitis associated with mumps. Arch Neurol. 2002;59:1633–1636. doi: 10.1001/archneur.59.10.1633. [DOI] [PubMed] [Google Scholar]
- 140.Singh K, Sodhi PK. Mumps-induced corneal endotheliitis. Cornea. 2004;23:400–402. doi: 10.1097/00003226-200405000-00017. [DOI] [PubMed] [Google Scholar]
- 141.St Geme JW, Jr, Peralta H, Van Pelt LF. Intrauterine infection of the rhesus monkey with mumps virus: abbreviated viral replication in the immature fetus as an explanation for split immunologic recognition after birth. J Infect Dis. 1972;126:249–256. doi: 10.1093/infdis/126.3.249. [DOI] [PubMed] [Google Scholar]
- 142.Kurtz JB, Tomlinson AH, Pearson J. Mumps virus isolated from a fetus. Br Med J (Clin Res Ed) 1982;284:471. doi: 10.1136/bmj.284.6314.471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Ylinen O, Jarniven PA. Parotitis during pregnancy. Acta Obstet Gynecol Scand. 1953;32:121–132. doi: 10.3109/00016345309157568. [DOI] [PubMed] [Google Scholar]
- 144.Yamauchi T, Wilson C, St Geme JW., Jr Transmission of live, attenuated mumps virus to the human placenta. N Engl J Med. 1974;290:710–712. doi: 10.1056/NEJM197403282901304. [DOI] [PubMed] [Google Scholar]
- 145.Garcia AG, Pereira JM, Vidigal N, et al. Intrauterine infection with mumps virus. Obstet Gynecol. 1980;56:756–759. [PubMed] [Google Scholar]
- 146.Jones JF, Ray CG, Fulginiti VA. Perinatal mumps infection. J Pediatr. 1980;96:912–914. doi: 10.1016/s0022-3476(80)80577-4. [DOI] [PubMed] [Google Scholar]
- 147.Kilham L. Mumps virus in human milk and in milk of infected monkey. J Am Med Assoc. 1951;146:1231–1232. doi: 10.1001/jama.1951.63670130011012f. [DOI] [PubMed] [Google Scholar]
- 148.Lacour M, Maherzi M, Vienny H, et al. Thrombocytopenia in a case of neonatal mumps infection: evidence for further clinical presentations. Eur J Pediatr. 1993;152:739–741. doi: 10.1007/BF01953989. [DOI] [PubMed] [Google Scholar]
- 149.Siegel M. Congenital malformations following chickenpox, measles, mumps, and hepatitis. Results of a cohort study. J Am Med Assoc. 1973;226:1521–1524. [PubMed] [Google Scholar]
- 150.Centers for Disease Control and Prevention. Mumps surveillance report. Morbid Mortal Weekly Rep. 1972;21:1–12. [Google Scholar]
- 151.Modlin JF, Orenstein WA, Brandling-Bennett AD. Current status of mumps in the United States. J Infect Dis. 1975;132:106–109. doi: 10.1093/infdis/132.1.106. [DOI] [PubMed] [Google Scholar]
- 152.Ennis FA, Hopps HE, Douglas RD, et al. Hydrocephalus in hamsters: induction by natural and attenuated mumps viruses. J Infect Dis. 1969;119:75–79. doi: 10.1093/infdis/119.1.75. [DOI] [PubMed] [Google Scholar]
- 153.McCarthy M, Jubelt B, Fay DB, et al. Comparative studies of five strains of mumps virus in vitro and in neonatal hamsters: evaluation of growth, cytopathogenicity, and neurovirulence. J Med Virol. 1980;5:1–15. doi: 10.1002/jmv.1890050102. [DOI] [PubMed] [Google Scholar]
- 154.Kilham L, Margolis G. Induction of congenital hydrocephalus in hamsters with attenuated and natural strains of mumps virus. J Infect Dis. 1975;132:462–466. doi: 10.1093/infdis/132.4.462. [DOI] [PubMed] [Google Scholar]
- 155.Wolinsky JS, Stroop WG. Virulence and persistence of three prototype strains of mumps virus in newborn hamsters. Arch Virol. 1978;57:355–359. doi: 10.1007/BF01320075. [DOI] [PubMed] [Google Scholar]
- 156.Buynak EB, Hilleman MR. Live attenuated mumps virus vaccine. 1. Vaccine development. Proc Soc Exp Biol Med. 1966;123:768–775. doi: 10.3181/00379727-123-31599. [DOI] [PubMed] [Google Scholar]
- 157.Rozina EE, Hilgenfeldt M. Comparative study on the neurovirulence of different vaccine strains of parotitis virus in monkeys. Acta Virol. 1985;29:225–230. [PubMed] [Google Scholar]
- 158.Afzal MA, Marsden S, Hull RM, et al. Evaluation of the neurovirulence test for mumps vaccines. Biologicals. 1999;27:43–49. doi: 10.1006/biol.1999.0174. [DOI] [PubMed] [Google Scholar]
- 159.Xu P, Huang Z, Gao X, et al. Infection of mice, ferrets, and rhesus macaques with a clinical mumps virus isolate. J Virol. 2013;87:8158–8168. doi: 10.1128/JVI.01028-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Tsurudome M, Yamada A, Hishiyama M, et al. Replication of mumps virus in mouse: transient replication in lung and potential of systemic infection. Arch Virol. 1987;97:167–179. doi: 10.1007/BF01314419. [DOI] [PubMed] [Google Scholar]
- 161.Kristensson K, Orvell C, Malm G, et al. Mumps virus infection of the developing mouse brain – appearance of structural virus proteins demonstrated with monoclonal antibodies. J Neuropathol Exp Neurol. 1984;43:131–140. doi: 10.1097/00005072-198403000-00003. [DOI] [PubMed] [Google Scholar]
- 162.Hayashi K, Ross ME, Notkins AL. Persistence of mumps viral antigens in mouse brain. Jpn J Exp Med. 1976;46:197–200. [PubMed] [Google Scholar]
- 163.Overman JR, Peers JH, Kilham L. Pathology of mumps virus meningoencephalitis in mice and hamsters. Arch Pathol. 1953;55:457–465. [PubMed] [Google Scholar]
- 164.Parker L, Gilliland SM, Minor P, et al. Assessment of the ferret as an in vivo model for mumps virus infection. J Gen Virol. 2013;94:1200–1205. doi: 10.1099/vir.0.052449-0. [DOI] [PubMed] [Google Scholar]
- 165.Rubin SA, Pletnikov M, Taffs R, et al. Evaluation of a neonatal rat model for prediction of mumps virus neurovirulence in humans. J Virol. 2000;74:5382–5384. doi: 10.1128/jvi.74.11.5382-5384.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 166.Lemon K, Rima BK, McQuaid S, et al. The F gene of rodent brain-adapted mumps virus is a major determinant of neurovirulence. J Virol. 2007;81:8293–8302. doi: 10.1128/JVI.00266-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Sauder CJ, Zhang CX, Ngo L, et al. Gene-specific contributions to mumps virus neurovirulence and neuroattenuation. J Virol. 2011;85:7059–7069. doi: 10.1128/JVI.00245-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Xu P, Li Z, Sun D, et al. Rescue of wild-type mumps virus from a strain associated with recent outbreaks helps to define the role of the SH ORF in the pathogenesis of mumps virus. Virology. 2011;417:126–136. doi: 10.1016/j.virol.2011.05.003. [DOI] [PMC free article] [PubMed] [Google Scholar]