

Abstract
The purpose of this study was to investigate the genetic features of canine coronavirus (CCV) strains detected in Korea. M gene sequences obtained for isolates from 22 dogs with enteritis over a 5-year period were evaluated. Sequence comparison revealed that the 22 Korean CCV strains had an 87.2 to 100% nucleotide homology. Comparing to the typical reference CCV strains (type II), the nucleotide sequence of Korean strains had homology ranged from 86.3% to 98.3% (89.1% to 99.2% for the amino acid sequence) and 87.7% to 97.8% (92.4% to 100% for the amino acid sequence) when compared to FCoV-like CCV strains (type I). Three amino acid variations in the M gene were characteristic for the Korean CCV strains. Phylogenetic analysis demonstrated that the 22 Korean CCV strains belonged to four typical CCV clusters (i.e., a unique Korean CCV cluster, a type II and transmissible gastroenteritis virus cluster, an intermediate cluster between type I and II, and a type I cluster). This study was the first to identify genetic differences of the M gene from Korean CCV strains and provided a platform for molecular identification of different Korean CCV strains.
Keywords: canine coronavirus, M gene, phylogenetic analysis
Introduction
Canine coronavirus (CCV) is an enveloped RNA virus with a positive-sense single-stranded genome of 27~32 kb in length [21]. This virus belongs to group I of the Coronaviridae family that includes transmissible gastroenteritis virus (TGEV) of swine, porcine epidemic diarrhea virus, porcine respiratory coronavirus, feline infectious peritonitis virus, and human coronavirus 229E [23,30]. The close antigenic relationship between feline coronavirus (FCoV) and CCV facilitates a potential for cross-species infection [3,13,16,20]. For both FCoV and CCV, interspecies recombinations have been found by sequence analysis in different parts of the viral genome (e.g., the spike [S] gene encoding the 'spike' on the viral surface and the membrane [M] gene encoding the 'membrane' or 'integral membrane' protein) [10,18,19].
A new FCoV type, designated as FCoV type II, has emerged in the field due to recombination (in the S protein gene region) of FCoV type I and CCV [10]. Epidemiological investigations have shown that FCoV type I is the predominant field virus type in the UK, Austria, and Japan [1,5,11]. Furthermore, other studies have demonstrated that FCoV type I strains bind to feline aminopeptidase N (fAPN), a cell surface receptor in the lung, spleen, kidney, and gut while FCoV type II strains do not. The possible clinical and epidemiological impact of these findings is still unclear [5,32]. In a recent study of S gene sequences obtained from field isolates, two types of CCV were identified. Pratelli et al. [24,25,26,27] analyzed M gene sequences of CCV strains and found that the isolates belonged to two novel genetic clusters. The newly discovered FCoV-like CCV strain was different from typical CCV strains, and more closely related to FCoV strains than CCV strains [7]. Divergent FCoV-like CCV strains have emerged in particular from dog kennels and animal shelters. One study found that some FCoV-like CCV strains are more virulent than typical CCV strains and cause severe hemorrhagic diarrhea [4]. FCoV-like CCV stains have been found in Austria, Italy, and China [4,26,34]. Accurate genotyping for field CCV strains is undoubtedly important. However, only one molecular genetic study of Korean CCV strains has been performed to date [2]. Therefore, the present investigation was conducted to assess the molecular genetic features of CCV strains isolated in Korea.
Materials and Methods
Clinical specimens
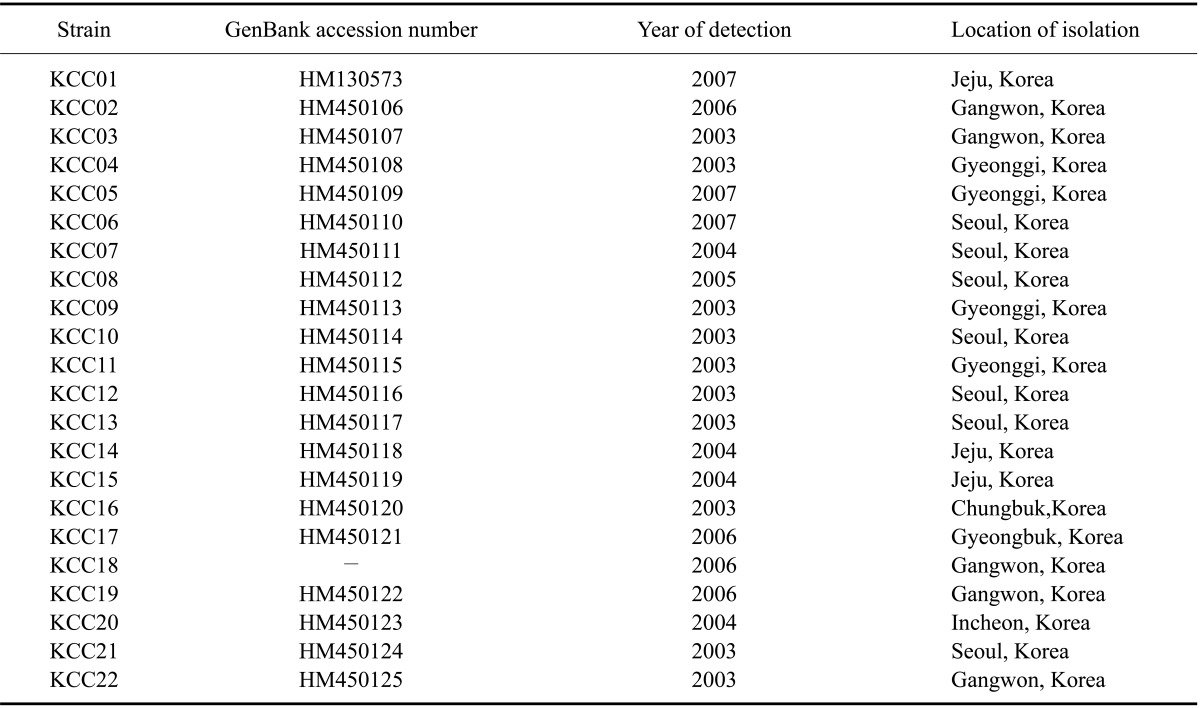
A total of 67 fecal samples of diarrheic dogs from various parts of Korea were collected by rectal swab from 2003 to 2007. The samples were transported on ice as quickly as possible and stored at -80℃ before use. CCV was detected by reverse transcription (RT)-PCR and canine parvovirus was detected with a SNAP Parvo Antigen Test Kit (IDEXX Laboratories, USA). CCVs were detected in 22 dogs (Table 1). Eight animals were male and 14 were female. The age of the dogs ranged from 5 to 31 weeks. Only 1 dog had received a CCV vaccination. Eleven dogs were vomiting and four had symptoms of respiratory disease such as coughing and nasal discharge. Canine parvovirus (CPV) was detected in all fecal samples in which CCV was detected.
Table 1.
Korean canine coronavirus (CCV) strains used for sequence comparison and phylogenetic analysis
PCR amplification
Viral RNA was extracted from fecal samples using an RNeasy mini kit (Qiagen, Germany) according to the manufacturer's protocol for animal cells. The target sequence for amplification was located in a segment of the gene encoding the transmembrane protein M of CCV. The following primers were prepared [22,28]: primer CCV1 (sense) 5'-TCC AGA TAT GTA ATG TTC GG-3' (337-356 nucleotides) and primer CCV2 (antisense) 5'-TCT GTT GAG TAA TCA CCA GCT-3' (726-746 nucleotides). The CCV1/CCV2 primer pair was used to amplify a 409-bp fragment [10]. cDNA was synthesized with the commercial kit (Accupower RT PreMix; Bioneer, Korea) as instructed by the manufacturer. cDNA synthesis was performed at 37℃ for 30 min with a final incubation at 94oC for 5 min. PCR was carried out using the commercial kit (Accupower PCR PreMix; Bioneer, Korea) with the following conditions: 35 cycles of denaturation at 94℃ for 30 sec, annealing at 55℃ for 30 sec, and extension at 72℃ for 1 min followed by a final extension at 72℃ for 10 min. The amplicons were separated on a 2% agarose gel and visualized by staining with ethidium bromide (10 µL/mL; Sigma, USA).
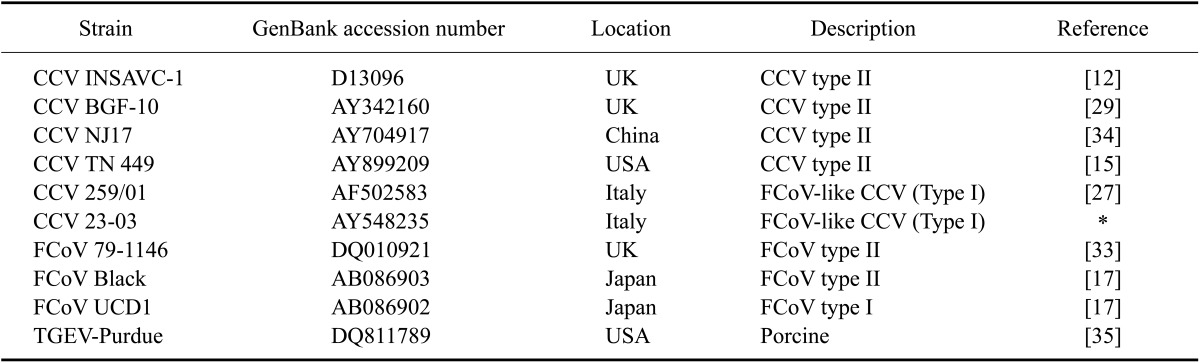
The PCR amplicons were purified with a commercial kit (QIAquick Gel Extraction Kit; Qiagen), and sequenced using the dideoxynucleotide chain termination method with the CCV1/CCV2 primer pair and a commercial kit (Big Dye terminator cycle sequencing kit; Applied Biosystems, USA). The nucleotide and deduced amino acid sequences of the M gene from 22 Korean CCV strains were determined and compared to those for 10 reference coronaviruses obtained from GenBank (National Center for Biotechnology Information, USA), and aligned by the Clusteral W multiple sequence alignment algorithm using commercial software (DNASTAR 5.0, MegAlign; DNASTAR, USA) for phylogenetic analysis (Tables 1 and 2).
Table 2.
Reference coronavirus strains used for sequence comparison and phylogenetic analysis
*Unpublished data.
Results
Analysis of the M gene revealed that the nucleotide sequences of the 22 Korean strains had an 87.2 to 100% homology. Furthermore, the Korean strains had an 86.3% to 98.3% sequence homology with the CCV type II reference strains (i.e., CCV INSAVC-1, BGF-10, NJ17, and TN 449) as well as an 87.7% to 97.8% sequence homology with the FCoV-like CCV type I reference strains (i.e., CCV 259/01 and 23-03). M gene sequences of seven Korean strains (i.e., KCC06, KCC07, KCC08, KCC09, KCC10, KCC11, and KCC12) shared a complete identity at the nucleotide level (Table 2 and 3).
Table 3.
Sequence homology of nucleotides and deduced amino acids of the M gene in the Korean CCV isolates and reference strains
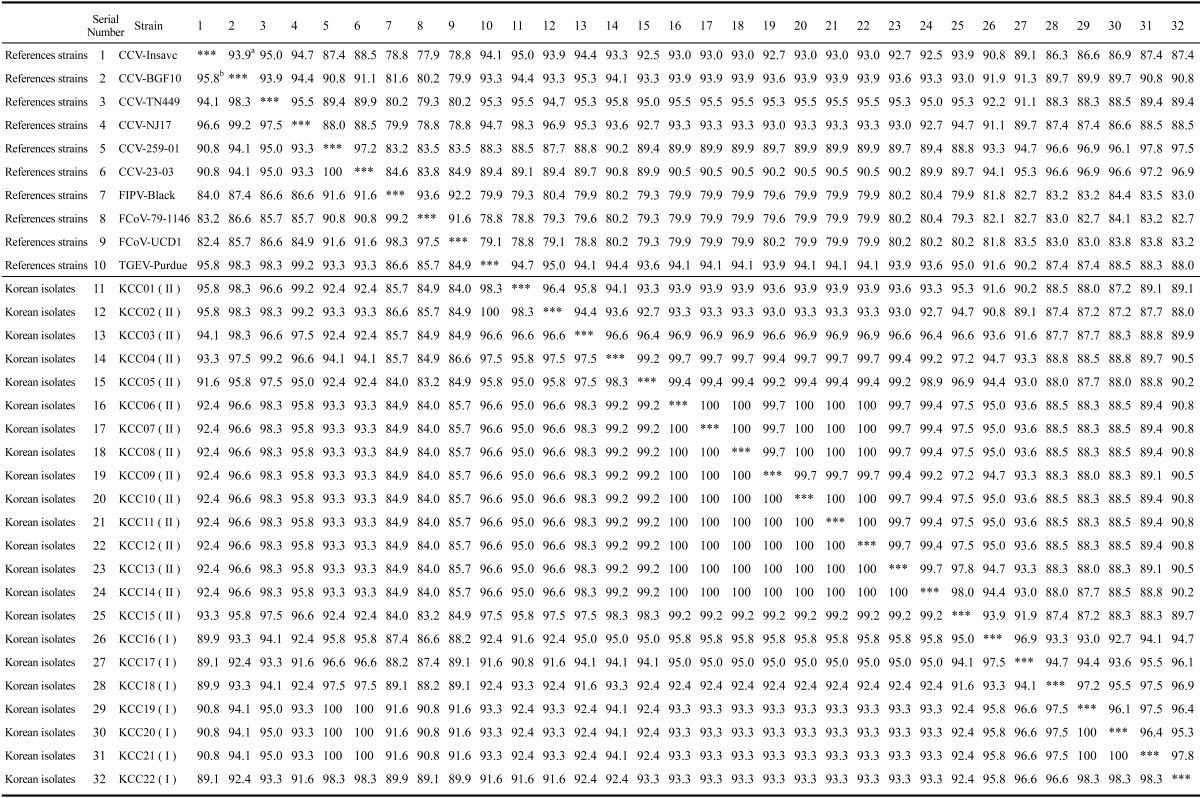
aPercent nucleotide similarity is indicated in the upper right triangle. bPercent amino acid similarity is indicated in the lower left triangle. cNucleotide and deduced amino acid sequences of the M genes obtained from GenBank. CCV-II: INSAVC (accession no. D13096). CCV-II: BGF-10 (AY342160), CCV-II: TN 449 (AY899209), CCV-II: NJ17: (AY704917), FCoV-like CCV: 259/01 (AF502583), FCoV-like CCV: 23-03 (AY548235), FCoV-I: 79-1146 (DQ010921), FCoV-II: Black (AB086903), FCoV-II: UCD1 (AB086902), and TGEV: Purdue (DQ811789).
Analysis of the M gene deduced amino acid sequence revealed that the Korean strains had a 90.8% to 100% similarity. Furthermore, the Korean strains showed an 89.1% to 99.2% sequence homology with the CCV type II reference strains (i.e., CCV INSAVC-1, BGF-10, NJ17, and TN 449) along with a 92.4% to 100% sequence homology with the FCoV-like CCV type I reference strains (i.e., CCV 259/01 and 23-03). Amino acid sequences of the M gene obtained for nine Korean strains (i.e. KCC06, KCC07, KCC08, KCC09, KCC10, KCC11, KCC12, KCC13, and KCC14) shared a complete identity while those obtained for three Korean strains (i.e. KCC19, KCC20, and KCC21) shared a complete identity with the reference FCoV-like CCV strains (i.e. CCV 259/01 and CCV23-03; Tables 2 and 3). There were characteristic sequence variations of the M gene among the Korean strains (i.e., Ser165Asn, Trp232Cys/Leu, and Ala233Ser). Although most nucleotide substitutions observed in the Korean strains were synonymous, some substitutions led to amino acid changes (i.e., Ile123Ile/Val, Val124Ile, Ile127Ala, Ser154Asn, Val173Thr, Cys187Tyr, Ile193Met, Asp200Glu, Asn201His, Val212Ser, Lys223Gln, and Asp242Gly; Table 4).
Table 4.
Amino acid variations of the M gene in the CCV strains analyzed for this study
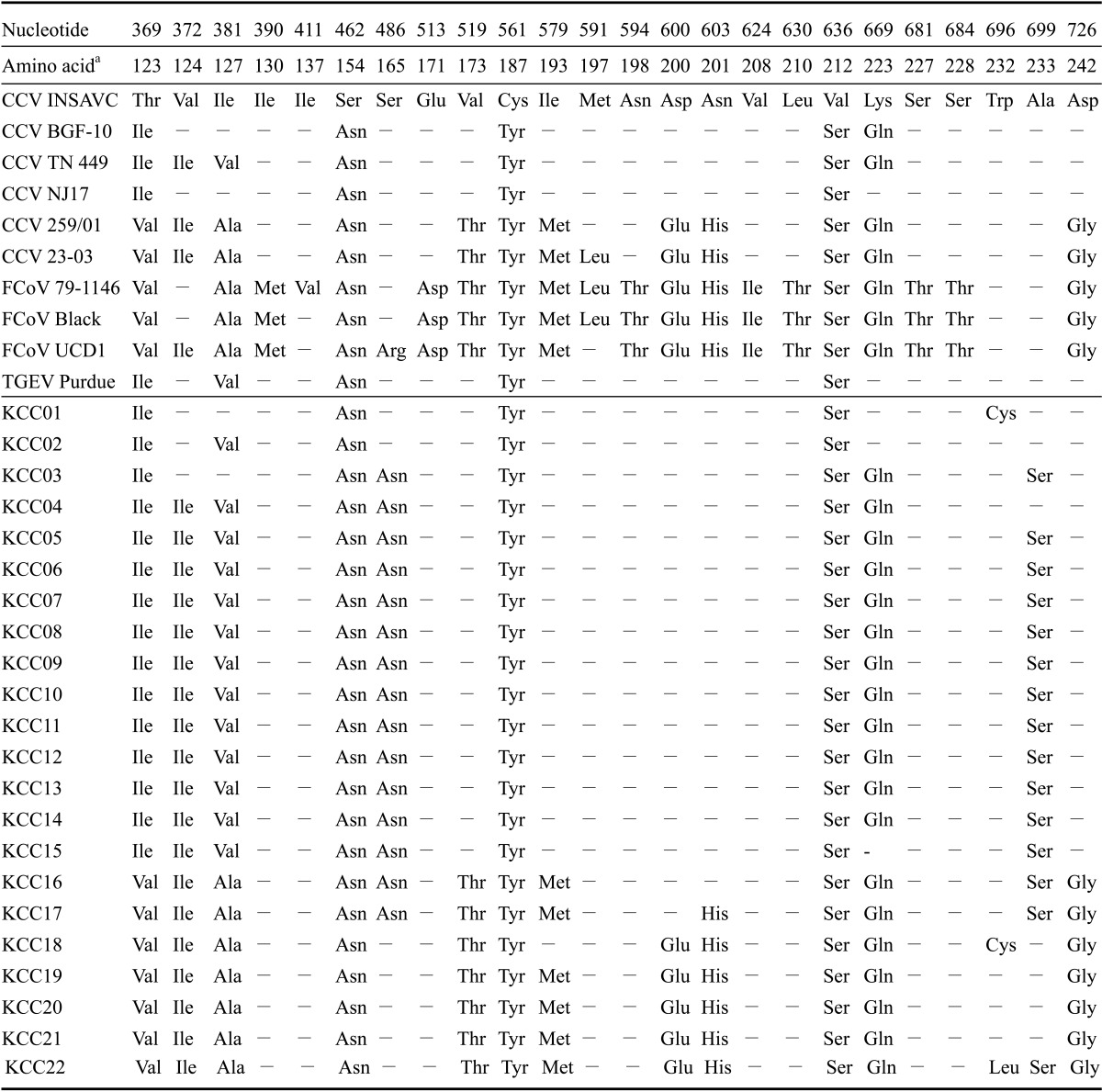
aAmino acid sequences of the M genes obtained from GenBank. CCV-II: INSAVC (accession no. D13096). CCV-II: BGF-10 (AY342160), CCV-II: TN 449 (AY899209), CCV-II: NJ17: (AY704917), FCoV-like CCV: 259/01 (AF502583), FCoV-like CCV: 23-03 (AY548235), FCoV-I: 79-1146 (DQ010921), FCoV-II: Black (AB086903), FCoV-II: UCD1 (AB086902), and TGEV: Purdue (DQ811789). bAmino acids identical to those in the reference strain are indicated by dashes while amino acids different from those in the reference strain are indicated by letters.
Results of the phylogenetic analysis of the deduced amino acid sequences obtained for the 22 Korean strains and 10 reference coronaviruses are summarized in Fig. 1. The 22 Korean strains belonged to four genetic clusters based on the description of the reference CCV strains of Table 2. These included a unique Korean CCV cluster (cluster I), CCV type II and TGEV cluster (cluster II), intermediate cluster between CCV type I and II (cluster III), and CCV type I cluster (cluster IV).
Fig. 1.

Phylogenetic tree of the CCVs based on a comparison of partial M gene amino acid sequences obtained for 22 Korean strains and 10 reference coronaviruses. CI, cluster I; CII, cluster II; CIII, cluster III; and CIV, cluster IV.
Discussion
The M glycoprotein of CCV can elicit strong immune responses [8,21]. The mutations in the M gene might confer some sort of growth advantage or avoiding host immunity [27]. The main purposes of this study were to determine the genetic diversity among CCV strains detected in Korea and examine their relationships to previously characterized strains. Twenty-two CCV strains from domestic dogs with enteritis in Korea were evaluated. These isolates shared high nucleotide and amino acid identities with each other. The nucleotide and amino acid sequence homologies indicated that most of the Korean strains were more closely related to the CCV type II strain than the FCoV-like CCV (type I) strain. This result is similar to findings from investigations conducted in other countries. Escutenaire et al. [9] showed that Swedish isolates share a higher identity with typical type II CCV based on the deduced amino acid sequences of the 5' region of the M gene.
Although the number of samples in our study is too small to yield firm conclusions, we provided evidence that the circulating CCVs in Korea can be divided into four separate clusters. CCVs in cluster I had unique amino acid changes in the M protein and these isolates were very close to the CCV type II reference strains. However, these strains are divergent from the CCV type II reference strains. CCVs in cluster II were close to the CCV type II reference strains and TGEV.
Members of cluster III were similar to the CCV type II reference strains and FCoV-like CCV strains, and had greater identities with the FCoV-like CCV strains than CCV type II reference strains. In another study, CCV isolates from Austrian dogs were found to belong to both the CCV type II cluster and FCoV-like CCV cluster [4]. A phylogenic analysis by Wang et al. [34] showed that one Chinese strain belonged to a cluster between the FCoV-like CCV and CCV type II branches, suggesting the existence of a novel strain. However, Swedish CCVs were segregated into two distinct subgroups of type II CCV strains [9]. The increasing evidence suggests that recombination of coronaviruses, such as CCV and FCoV, often occur under natural conditions. The potential of interspecies circulation of either CCV in cats or FCoV in dogs is very high. And when co-infection occurs in the host, recombination can happen between two coronaviruses naturally [3,16,21,26,31].
The CCVs in cluster IV consisted of FCoV-like CCV reference strains. This result showed that there are FCoV-like CCV strains in Korea similar those described by Pratelli et al. [22,26,27] in Italy and Benetka et al. [4] in Austria. The FCoV-like CCV strain differs from the CCV type II strain with amino acid changes at 12 positions (Ile123Ile/Val, Val124Ile, Ile127Ala, Ser154Asn, Val173Thr, Cys187Tyr, Ile193Met, Asp200Glu, Asn201His, Val212Ser, Lys223Gln, and Asp242Gly) in the M protein [4,25]. This finding highlights the possibility that FCoVs may be transmitted to dogs, and that there may be frequent recombination between the two types of virus [4]. Moreover, the high frequency of recombinations (in addition to small insertions and deletions) in genome containing both structural and non-structural proteins is the dominant force in the microevolution of positive RNA viruses, and results in the proliferation of different virus strains with different biological and immunological functions [6,27]. Coronavirus genome is predicted to accumulate several base substitutions per round of replication as a result of RNA-dependent RNA polymerase errors [14,27]. Due to the relatively high mutation frequency, CCVs have the potential to rapidly adjust to negative pressures such as those presented by the immune system [21,25]. The possibility that the dogs might have been infected simultaneously with population of genetically distinct CCVs and that one population might have prevailed against the other during the long period of virus shedding cannot be excluded. This hypothesis could explain the stable and identical substitutions observed in the nucleotide sequences of the M gene.
In summary, genetic differences of the M gene from Korean CCV strains were identified. Data from this investigation provide a platform for the molecular identification of different Korean CCV strains. Further sequence analysis needs to be extended to other parts of the genomes of CCV and FCoV field strains to obtain more detailed information about the predilection sites of such recombination and mutation events.
Acknowledgments
This study was supported by 2013 Research Grant from Kangwon National University (no. C1010202-01-01).
Footnotes
There is no conflict of interest.
References
- 1.Addie DD, Schaap IAT, Nicolson L, Jarrett O. Persistence and transmission of natural type I feline coronavirus infection. J Gen Virol. 2003;84:2735–2744. doi: 10.1099/vir.0.19129-0. [DOI] [PubMed] [Google Scholar]
- 2.An DJ, Jeong W, Yoon SH, Jeoung HY, Kim HJ, Park BK. Genetic analysis of canine group 2 coronavirus in Korean dogs. Vet Microbiol. 2010;141:46–52. doi: 10.1016/j.vetmic.2009.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Barlough JE, Stoddart CA, Sorresso GP, Jacobson RH, Scott FW. Experimental inoculation of cats with canine coronavirus and subsequent challenge with feline infectious peritonitis virus. Lab Anim Sci. 1984;34:592–597. [PubMed] [Google Scholar]
- 4.Benetka V, Kolodziejek J, Walk K, Rennhofer M, Möstl K. M gene analysis of atypical strains of feline and canine coronavirus circulating in an Austrian animal shelter. Vet Rec. 2006;159:170–175. doi: 10.1136/vr.159.6.170. [DOI] [PubMed] [Google Scholar]
- 5.Benetka V, Kübber-Heiss A, Kolodziejek J, Nowotny N, Hofmann-Parisot M, Möstl K. Prevalence of feline coronavirus types I and II in cats with histopathologically verified feline infectious peritonitis. Vet Microbiol. 2004;99:31–42. doi: 10.1016/j.vetmic.2003.07.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Dolja VV, Carrington JC. Evolution of positive-strand RNA viruses. Semin Virol. 1992;3:315–326. [Google Scholar]
- 7.East ML, Moestl K, Benetka V, Pitra C, Höner OP, Wachter B, Hofer H. Coronavirus infection of spotted hyenas in the Serengeti ecosystem. Vet Microbiol. 2004;102:1–9. doi: 10.1016/j.vetmic.2004.04.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Enjuanes L, Spaan W, Snijder E, Cavanach D. Nidovirales. In: van Regenmortel MHV, Fauquet CM, Bishop DHL, Carstens EB, Estes MK, Lemon SM, Maniloff J, Mayo MA, McGeoch DJ, Pringle CR, Wickner RB, editors. Virus Taxonomy. Seventh Report of the International Committee on Taxonomy of Viruses. San Diego: Academic Press; 2000. pp. 835–849. [Google Scholar]
- 9.Escutenaire S, Isaksson M, Renström LHM, Klingeborn B, Buonavoglia C, Berg M, Belák S, Thorén P. Characterization of divergent and atypical canine coronaviruses from Sweden. Arch Virol. 2007;152:1507–1514. doi: 10.1007/s00705-007-0986-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Herrewegh AAPM, Smeenk I, Horzinek MC, Rottier PJM, de Groot RJ. Feline coronavirus type II strains 79-1683 and 79-1146 originate from a double recombination between feline coronavirus type I and canine coronavirus. J Virol. 1998;72:4508–4514. doi: 10.1128/jvi.72.5.4508-4514.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hohdatsu T, Okada S, Ishizuka Y, Yamada H, Koyama H. The prevalence of types I and II feline coronavirus infections in cats. J Vet Med Sci. 1992;54:557–562. doi: 10.1292/jvms.54.557. [DOI] [PubMed] [Google Scholar]
- 12.Horsburgh BC, Brierley I, Brown TDK. Analysis of a 9·6kb sequence from the 3' end of canine coronavirus genomic RNA. J Gen Virol. 1992;73:2849–2862. doi: 10.1099/0022-1317-73-11-2849. [DOI] [PubMed] [Google Scholar]
- 13.Horzinek MC, Lutz H, Pedersen NC. Antigenic relationships among homologous structural polypeptides of porcine, feline, and canine coronaviruses. Infect Immun. 1982;37:1148–1155. doi: 10.1128/iai.37.3.1148-1155.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Jarvis TC, Kirkegaard K. The polymerase in its labyrinth: mechanisms and implications of RNA recombination. Trends Genet. 1991;7:186–191. doi: 10.1016/0168-9525(91)90434-R. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Ma G, Wang Y, Lu C. Molecular characterization of the 9.36 kb C-terminal region of canine coronavirus 1-71 strain. Virus Genes. 2008;36:491–497. doi: 10.1007/s11262-008-0214-4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.McArdle F, Bennett M, Gaskell RM, Tennant B, Kelly DF, Gaskell CJ. Induction and enhancement of feline infectious peritonitis by canine coronavirus. Am J Vet Res. 1992;53:1500–1506. [PubMed] [Google Scholar]
- 17.Motokawa K, Hohdatsu T, Hashimoto H, Koyama H. Comparison of the amino acid sequence and phylogenetic analysis of the peplomer, integral membrane and nucleocapsid proteins of feline, canine and porcine coronaviruses. Microbiol Immunol. 1996;40:425–433. doi: 10.1111/j.1348-0421.1996.tb01089.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Naylor MJ, Harrison GA, Monckton RP, McOrist S, Lehrbach PR, Deane EM. Identification of canine coronavirus strains from feces by S gene nested PCR and molecular characterization of a new Australian isolate. J Clin Microbiol. 2001;39:1036–1041. doi: 10.1128/JCM.39.3.1036-1041.2001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Naylor MJ, Walia CS, McOrist S, Lehrbach PR, Deane EM, Harrison GA. Molecular characterization confirms the presence of a divergent strain of canine coronavirus (UWSMN-1) in Australia. J Clin Microbiol. 2002;40:3518–3522. doi: 10.1128/JCM.40.9.3518-3522.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Pedersen NC, Ward J, Mengeling WL. Antigenic relationship of the feline infections peritonitis virus to coronaviruses of other species. Arch Virol. 1978;58:45–53. doi: 10.1007/BF01315534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Pratelli A. Genetic evolution of canine coronavirus and recent advances in prophylaxis. Vet Res. 2006;37:191–200. doi: 10.1051/vetres:2005053. [DOI] [PubMed] [Google Scholar]
- 22.Pratelli A, Buonavoglia D, Martella V, Tempesta M, Lavazza A, Buonavoglia C. Diagnosis of canine coronavirus infection using nested-PCR. J Virol Methods. 2000;84:91–94. doi: 10.1016/S0166-0934(99)00134-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Pratelli A, Elia G, Decaro N, Tola S, Tinelli A, Martella V, Rocca S, Tempesta M, Buonavoglia C. Cloning and expression of two fragments of the S gene of canine coronavirus type I. J Virol Methods. 2004;117:61–65. doi: 10.1016/j.jviromet.2003.12.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Pratelli A, Elia G, Martella V, Palmieri A, Cirone F, Tinelli A, Corrente M, Buonavoglia C. Prevalence of canine coronavirus antibodies by an enzyme-linked immunosorbent assay in dogs in the south of Italy. J Virol Methods. 2002;102:67–71. doi: 10.1016/S0166-0934(01)00450-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Pratelli A, Elia G, Martella V, Tinelli A, Decaro N, Marsilio F, Buonavoglia D, Tempesta M, Buonavoglia C. M gene evolution of canine coronavirus in naturally infected dogs. Vet Rec. 2002;151:758–761. [PubMed] [Google Scholar]
- 26.Pratelli A, Martella V, Decaro N, Tinelli A, Camero M, Cirone F, Elia G, Cavalli A, Corrente M, Greco G, Buonavoglia D, Gentile M, Tempesta M, Buonavoglia C. Genetic diversity of a canine coronavirus detected in pups with diarrhoea in Italy. J Virol Methods. 2003;110:9–17. doi: 10.1016/S0166-0934(03)00081-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Pratelli A, Martella V, Pistello M, Elia G, Decaro N, Buonavoglia D, Camero M, Tempesta M, Buonavoglia C. Identification of coronaviruses in dogs that segregate separately from the canine coronavirus genotype. J Virol Methods. 2003;107:213–222. doi: 10.1016/S0166-0934(02)00246-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Pratelli A, Tempesta M, Greco G, Martella V, Buonavoglia C. Development of a nested PCR assay for the detection of canine coronavirus. J Virol Methods. 1999;80:11–15. doi: 10.1016/S0166-0934(99)00017-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Sanchez-Morgado JM, Poynter S, Morris TH. Molecular characterization of a virulent canine coronavirus BGF strain. Virus Res. 2004;104:27–31. doi: 10.1016/j.virusres.2004.02.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Siddell SG. The coronaviridae: an introduction. In: Siddell SG, editor. The coronaviridae. New York: Plenum Press; 1995. pp. 1–10. [Google Scholar]
- 31.Stoddart CA, Barlough JE, Baldwin CA, Scott FW. Attempted immunisation of cats against feline infectious peritonitis using canine coronavirus. Res Vet Sci. 1988;45:383–388. doi: 10.1016/S0034-5288(18)30970-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Tresnan DB, Levis R, Holmes KV. Feline aminopeptidase N serves as a receptor for feline, canine, porcine, and human coronaviruses in serogroup I. J Virol. 1996;70:8669–8674. doi: 10.1128/jvi.70.12.8669-8674.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Vennema H, de Groot RJ, Harbour DA, Horzinek MC, Spaan WJ. Primary structure of the membrane and nucleocapsid protein genes of feline infectious peritonitis virus and immunogenicity of recombinant vaccinia viruses in kittens. Virology. 1991;181:327–335. doi: 10.1016/0042-6822(91)90499-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Wang Y, Ma G, Lu C, Wen H. Detection of canine coronaviruses genotype I and II in raised Canidae animals in China. Berl Munch Tierarztl Wochenschr. 2006;119:35–39. [PubMed] [Google Scholar]
- 35.Zhang X, Hasoksuz M, Spiro D, Halpin R, Wang S, Stollar S, Janies D, Hadya N, Tang Y, Ghedin E, Saif L. Complete genomic sequences, a key residue in the spike protein and deletions in nonstructural protein 3b of US strains of the virulent and attenuated coronaviruses, transmissible gastroenteritis virus and porcine respiratory coronavirus. Virology. 2007;358:424–435. doi: 10.1016/j.virol.2006.08.051. [DOI] [PMC free article] [PubMed] [Google Scholar]