

Abstract
Adenylyl cyclases are widely distributed across all kingdoms whereas guanylyl cyclases are generally thought to be restricted to eukaryotes. Here we report that the α–proteobacterium Rhodospirillum centenum secretes cGMP when developing cysts and that a guanylyl cyclase deletion strain fails to synthesize cGMP and is defective in cyst formation. The R. centenum cyclase was purified and shown to effectively synthesize cGMP from GTP in vitro, demonstrating that it is a functional guanylyl cyclase. A homolog of the Escherichia coli cAMP receptor protein (CRP) is linked to the guanylyl cyclase and when deleted is deficient in cyst development. Isothermal calorimetry (ITC) and differential scanning fluorimetry (DSF) analyses demonstrate that the recombinant CRP homolog preferentially binds to, and is stabilized by cGMP, but not cAMP. This study thus provides evidence that cGMP has a crucial role in regulating prokaryotic development. The involvement of cGMP in regulating bacterial development has broader implications as several plant-interacting bacteria contain a similar cyclase coupled by the observation that Azospirillum brasilense also synthesizes cGMP when inducing cysts.
Introduction
The cyclic nucleotide second messengers cAMP and cGMP regulate numerous cellular processes across kingdoms, altering the activities of sensor kinases (Francis and Corbin, 1994), ion-channels (Finn et al., 1996), phosphodiesterases (PDEs) (Heikaus et al., 2009), guanine nucleotide exchange factors (Quilliam et al., 1995) and transcription factors (Harman, 2001). Their synthesis from ATP and GTP is catalyzed by six classes of nucleotidyl cyclases grouped by differences in primary amino acid sequence of catalytic domains. Class I, II, V and VI cyclases are restricted to bacteria, whereas class IV cyclases are also present in archaea (Cotta et al., 1998; Danchin, 1993; Iyer and Aravind, 2002; Sismeiro et al., 1998; Tellez-Sosa et al., 2002). Class III enzymes comprise the majority of adenylyl, and the entirety of guanylyl cyclases, and are present in eukaryotes, bacteria and archaea (Shenroy and Visweswariah, 2004; Shaap, 2005). While numerous cAMP regulated signaling pathways have been elucidated in eukaryotes and bacteria, cGMP regulated processes have only been well defined in eukaryotes.
Even though early research indicated the presence of cGMP in several bacteria, many of these these studies are discredited due to difficulty in reproducibility, failure to isolate and biochemically characterize guanylyl cyclases, and the inability to definitively identify examples of cGMP regulated processes. Nevertheless, intracellular cGMP pools have been reported in several bacteria, with fluctuations often coinciding with changes in cellular growth states. For example, the existence of cGMP in Escherichia coli was reported in 1974 (Bernlohr et al., 1974), and later demonstrated to periodically increase from 0.02 to 0.2 pmol mg−1 protein per cell division in synchronized cultures (Cook et al., 1980). Intracellular cGMP in Caulobacter crescentus was also detected and found to decrease from 0.67 to 0.16 pmol mg−1 protein between exponential and stationary growth (Kurn et al., 1977). Similar low levels of cGMP have been reported in an assortment of other bacteria, including Bacillus megaterium (Setlow and Setlow, 1978), Bacillus licheniformis (Bernlohr et al., 1974) Myxococcus Xanthus (Ho and McCurdy, 1980), Mycobacterium smegmatis (Bhatnagar et al., 1984) and Bradyrhizobium japonicium (Lim et al., 1979). Genetic evidence appeared to identify the source of cGMP in E. coli as an unintended side activity of adenylyl cyclase (Shibuya et al., 1977). However, partially purified protein fractions exhibiting guanylyl cyclase activity have been reported from cell lysates of E. coli (Macchia et al., 1975; Macchia et al., 1981), C. crescentus (Sun et al., 1974), B. licheniformis (Clark and Bernlohr, 1972), M. smegmatis (Bhatnagar et al., 1984), M. Xanthus (Devi et al., 1984) and Nocardia sp. strain NRRL (Sun and Rosazza, 2000. In every instance, the identity of the enzyme in question has remained undetermined, precluding genetic and biochemical analyses that could prove the existence of a bona fide guanylyl cyclase in any of the above mentioned species. While intriguing, such studies left significant doubts as to the physiological significance of cGMP in bacteria.
In contrast to early studies that provided inconclusive evidence for cGMP in bacteria, higher intracellular concentrations of cGMP have recently been reported in several cyanobacterial species including Synechocystis PCC 6308 (17–35 pmol mg−1 protein), Plectonema (4.0 pmol mg−1 protein) and Nostoc PCC 8009 (1.0–3.5 pmol mg−1 protein) (Herdman and Elmorjani, 1988). In Synechocystis PCC 6803 cGMP levels are dependent on culture conditions, being relatively low during autotrophic growth (3.5–6.0 pmol mg−1 protein) as compared to mixotrophic and photoheterotrophic growth (~10 pmol mg−1 protein) or growth in media lacking combined nitrogen (20 pmol mg−1 protein) (Herdman and Elmorjani, 1988). A membrane bound class III cyclase (Cya2) was implicated in cGMP synthesis in Synechocystis PCC 6803 when a cya2 disrupted strain exhibited attenuated intracellular levels of cGMP, but not cAMP (Ochoa De Alda et al., 2000). Biochemical and structural studies of a Cya2 truncated catalytic domain demonstrated that it is indeed a highly specific guanylyl cyclase (Rauch et al., 2008). cGMP has been proposed to regulate UV-B induced photoacclimation in Synechocystis (Cadoret et al., 2005), although additional genetic and biochemical analyses are needed to verify this hypothesis, as well as to uncover regulators that actually interact with cGMP.
Recently we have undertaken genetic analysis of the complex developmental life-cycle present in Rhodospirillum centenum that features three morphologically distinct cell types; swim cells, swarm cells and resting cyst cells (Favinger et al., 1989; Ragatz et al., 1995, Berelman et al., 2004; Berelman and Bauer, 2004 and 2005). Metabolically dormant cysts are thought to form as a means of surviving environmental stresses such as desiccation, and have been reported in a wide range of proteobacterial genera. Ultra-structural and morphological characteristics of cyst cell formation have been well-studied in the Azotobacter and Azospirillum genera (Sadasivan and Neyra 1987; Stevenson and Socolofsky, 1966), the latter to which R. centenum is closely related (Fani et al., 1995; Stofels et al., 2001). R. centenum encystment typically involves nutrient starvation triggering a multi-stage developmental process that includes rounding of cells, production of large intracellular storage granules of poly-hydroxybutyrate (PHB) and the excretion of highly refractive lipopolysaccharide and lipoprotein protective coatings that eventually envelop dormant cyst cells (Berleman and Bauer, 2004). While descriptive features of encystment have been reported for several species, molecular mechanisms governing encystment are poorly understood, certainly not as well understood as regulators that govern Bacillus endospore formation or myxospore development in Myxococcus (Koos, 2008). In an effort to identify encystment regulatory elements, we undertook a screen for R. centenum mutants that suppressed a previously described hyper-cyst phenotype back to vegetative growth (Berleman et al., 2004). Two insertions fell within a single gene predicted to encode a class III cyclase domain that is shown to be involved in cGMP production required for encystment. Linked to the cyclase are two genes encoding proteins of unknown function that are also required for cGMP production, as well as a homolog of the E. coli transcription factor CRP that preferentially binds cGMP over cAMP, and is essential for inducing cyst formation in R. centenum.
Results
Identification of Cyst Regulatory Elements by Hyper-Cyst Suppression
Our previous screen for regulatory elements that control encystment centered on the isolation of hyper-cyst mutants displaying derepressed cyst formation (Berleman et al., 2004). The hyper-cyst screen successfully uncovered several regulatory elements, including a chemotaxis-like gene cluster (che3) involved in regulating encystment (Berleman et al., 2004; Berelman and Bauer 2005). However, this screen did not identify downstream DNA binding elements, prompting the development of an alternative screening strategy. In this study, we undertook mini-Tn5 transposon mutagenesis of a Δche3 hyper-cyst strain and screened for mutants that suppressed the hyper-cyst phenotype. Disrupted loci were identified by cloning and sequencing chromosomal segments that flanked the inserted transposon, and then searching against the sequenced R. centenum genome (Lu et al., 2010).
Our hyper-cyst suppression screen generated several mutants that had mini-Tn5 insertions within ORFs encoding putative regulatory elements (Table 1), including two histidine kinase response regulator hybrids (rc1_0896, rc1_3465), a serine/threonine phosphatase (rc1_3466) and a σ70 transcription factor (rc1_1638). Additionally, three insertions were in a region containing a class III nucleotidyl cyclase (ORF rc1_3783) of indeterminate substrate specificity (key substrate discrimination residues are not conserved as indicated in Fig. 1c), multiple ORFs of unknown function, and a homolog of the E. coli cAMP receptor protein (CRP) (ORF rc1_3788) (Fig. 1a, 1b, 1d). CRP is a founder of the CRP/FNR superfamily of prokaryotic transcription factors that possess an N-terminal cyclic nucleotide-monophosphate binding domain (cNMP) and a C-terminal helix-turn-helix (HTH) DNA-binding motif. CRP homologs have been shown to directly or indirectly regulate processes such as energy metabolism, development, virulence, quorum sensing and motility in diverse bacteria. All determinant residues of cyclic nucleotide binding to CRP (Passner et al., 2000; Weber and Steitz, 1987; Youn et al., 2007) are identically or functionally conserved in RC1_3788 (residues G90, E91, R100, T101, R141, T145, and S146), indicating that this homolog likely binds a cyclic nucleotide (Fig. 1d). Because of the shared genomic context of the CRP-like transcription factor with the class III nucleotidyl cyclase, we speculated that these proteins may have congruent regulatory roles in controlling encystment. This potentiality is the focus of this study.
Table 1.
ORFs disrupted by transposition screening for hypo-cyst suppressor mutants.
| HSM Strain | Locus Tag | Description |
|---|---|---|
| Metabolic Class | ||
| 44 | rc1_0155 | Purine biosynthesis |
| 41, 52, 142 | rc1_2014 | Pyrimidine biosynthesis |
| 67 | rc1_1525 | Pyrimidine biosynthesis |
| 43, 66, 94, 98, 138 | rc1_2543 | Undecaprenyl-phosphate galactose phosphotransferase |
| 28, 118 | rc1_2544 | UDP-glucose-4-epimerase |
| 26, 30, 68 | rc1_3992 | NAD dependant epimerase/dehydratase |
| 119 | rc1_2536 | Glycosyltransferase |
| 121, 123, 131, 146 | rc1_2537 | Glycosyltransferase |
| 101 | rc1_1410 | Polysaccharide biosynthesis/export protein |
| Regulatory Class | ||
| 69, 114, 115, 122 | rc1_3465 | Sensor kinase response regulator hybrid |
| 117 | rc1_3466 | Serine/threonine phosphatase |
| 90, 93, 97, 105, 108 | rc1_0896 | Sensor kinase response regulator hybrid |
| 70 | rc1_1638 | Sigma 70 transcription factor |
| 55, 76 | rc1_3783 | Class III nucleotidyl cyclase |
| 57 | rc1_3784/85 | Intergenic Space |
Figure 1.

Genetic context of the guanylyl cyclase-CRP gene cluster. (a) Mini-Tn5 insertion sites are denoted by black triangles (b) Identified domains in RC1_3783 include an adenylyl/guanylyl cyclase catalytic domain (CYCc) (aa 14–197), a predicted partial ATPase (aa 221–595), followed by seven tetratricopeptide repeats (TPR). Domains in RC1_3788 include a cyclic nucleotide-monophosphate binding domain (cNMP) (aa 21–139), and a Helix-turn-Helix (HTH) domain (aa 176–226). (c) Amino acid sequence alignment of the RC1_3783 cyclase domain with representative class III cyclase domains. Species abbreviations are: Rhodospirillum centenum (Rc) RC1_3783 (YP_002299938); Synechocystis sp. PCC 6803 (Sy) Cya2 (NP_440289); Homo sapiens (Hs) GUCY2C (NP_004954); Chlamydomonas reinhardtii (Cr) CYG12 (XP_001700847); Dictyostelium discoideum (Dc) ACRA (AAD50121); Arthrospira platensis (Ap) CyaC (BAA22997) & CyaG (BAB19924); Mycobacterium tuberculosis H37Rv (Mt) Rv1264 (NP_215780). Positions of residues involved in metal binding (Yellow), substrate specification (Blue) and transition state stabilization (Red) are indicated by asterisks (*), inverted triangles (▼) and arrows (↓). Shading indicates identically and functionally conserved residues at 100% (Black), 87.5 or 75% (Dark gray shading with white letters), and 62.5% (Light gray shading with dark letters) conservation. (d) Amino acid sequence alignment of RC1_3788 and representative CRP homologous. Sequences and species abbreviations used are as follows: Mycobacterium tuberculosis (Mt) Rv3676 (NP_218193); Escherichia coli (Ec) CRP (NP_417816), Streptomyces coelicolor (Sc) CRPSco (NP_627768), Synechocystis PCC 6803 (Sy) SyCRP1 (NP_440289). The alignment was constructed in MEGA v.4.0 utilizing a BLOSUM matrix, with respective pair-wise and multiple alignment gap opening and extension penalties of 20 and 0.2. Intra-subunit E. coli CRP residues which directly contact cAMP are shaded red and denoted by an asterisk (*) and a single inter-subunit contact (S128) is shaded blue and marked by an inverted triangle (▼). Residues which make direct contacts with DNA shaded yellow and denoted by arrows (↓). Additional shading indicates identically and functionally conserved residues at 100% (Black) and 80% (Dark gray) conservation.
Deletion of the Nucleotidyl Cyclase and CRP Homolog Disrupts Cyst Formation
To address the function of the nucleotidyl cyclase (rc1_3783) and CRP homolog (rc1_3788) in cyst cell development, we constructed in-frame deletions of these genes in a wild-type background. Wild-type and deletion mutants were then phenotyped with respect to encystment by plating on vegetative (CENS) and cyst inducing (CENS-8xN) culture media. When grown on CENS medium, wild-type and deletion strains (Δrc1_3783, Δrc1_3788) developed shiny, convex colonies that contained vegetative cells (Fig. 2a, 2b). However, when grown on cyst inducing CENS-8xN medium the wild-type strain rapidly underwent encystment, as evidenced by the formation of dry intricately ridged colonies that contain many round, refractive cyst cells (Fig. 2a, 2b). In stark contrast, strains deleted for the nucleotidyl cyclase (Δrc1_3783) and CRP homolog (Δrc1_3788) exhibited shiny vegetative colonies and vibroid shaped cells on cyst inducing medium (Fig. 2a, 2b). We also quantified encystment by assaying the percentage of viable cells that exhibit desiccation resistance, a trait only exhibited by cyst cells (Berleman and Bauer, 2004). When grown for 72 hours on vegetative cyst inducing CENS-8xN medium ~1% of viable wild-type cells were desiccation resistant, while cells of strains deleted for the nucleotidyl cyclase or the CRP homolog exhibited no assayable desiccation resistance (Fig. 2c).
Figure 2.
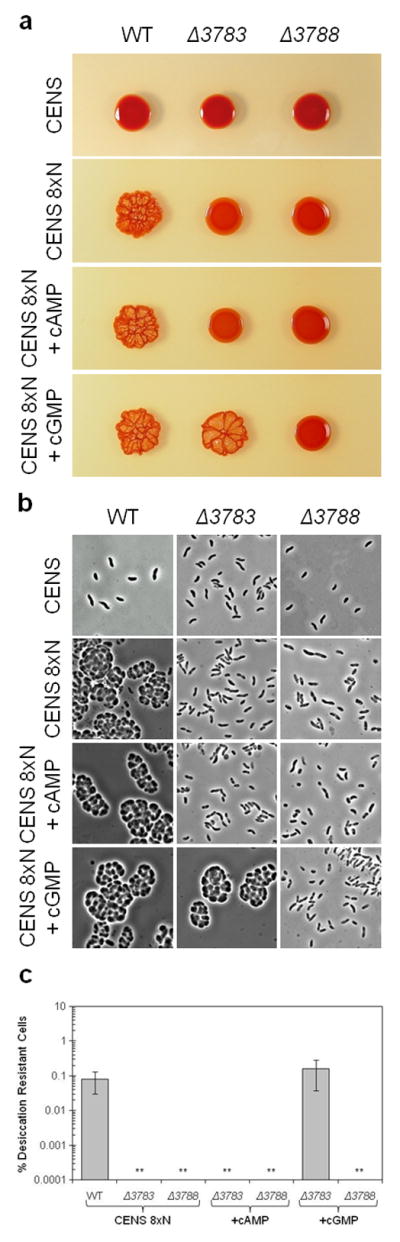
Analyses cyst cell formation in wild-type, Δrc1_3783 (nucleotidyl cyclase) and Δrc1_3788 (CRP homolog) strains after 3 days growth on agar solidified cyst-inducing CENS-8xN media. (a) Wild-type colonies formed dry/ridged cyst containing colonies under all growth conditions whereas a strain deleted for the cyclase (Δ rc1_3783) only formed cyst colonies in the presence of 50 μM cGMP. A strain deleted for the CRP homolog (Δrc1_3788) only formed shiny vegetative colonies (b) Microscopic observations of cyst and vegetative cells within the above shown colonies. (c) Desiccation resistant quantitation of cyst cells in the above colonies. The double asterisk (**) indicates that no cysts were detected in any cell dilution assayed.
cGMP is a Signal Controlling Encystment
We assessed whether R. centenum is capable of producing cGMP when undergoing cyst cell differentiation by assaying for the presence of cGMP during growth in butyrate minimal liquid medium that effectively induces cyst formation (Statwald-Demchick et al., 1990). The results show significant extracellular accumulation of cGMP in wild-type cultures that are undergoing cyst formation up to 33.0 nM (Fig. 3). This is contrasted by strains deleted for the nucleotidyl cyclase (Δrc1_3783) or the CRP homolog (Δrc1_3788) that fail to form cysts and have <1 nM cGMP production (Fig. 3). Similar measurements also showed that cAMP levels did not significantly accumulate during cyst induction in either the wild-type or deletion strains (data not shown). We also determined whether cGMP accumulates intracellularly by assaying cell free extracts derived from washed cell pellets. Intracellular cGMP levels in cells from 11 day-old cultures were measured at 13.2, 0.4 and 5.2 pmol/mg of total protein in respective cell pellets of the wild-type, Δrc1_3783 and Δrc1 _3788 strains.
Figure 3.

Accumulation of exogenous cGMP in cyst inducing liquid CENBA medium by wild-type cells (grey bars), the cyclase deletion strain Δrc1_3783 (black bars), and the CRP homolog strain Δrc1_3788 (white bars). While cGMP accumulates to appreciable levels in the wild type strain, in the Δrc1_3783 and Δrc1_3788 strains levels remain <1 nM.
Previous studies have shown that addition of exogenous cAMP is capable of complementing disruptions of microbial adenylyl cyclases (Roberts et al., 2007; Terauchi and Ohmori, 1999). We therefore attempted to complement the cyst deficient (hypo-cyst) phenotype exhibited by deletions of the nucleotidyl cyclase and CRP homologs by growing cells on CENS-8xN encystment media supplemented with cAMP or cGMP. When grown on cyst inducing CENS-8xN medium, the wild-type strain formed cysts after three days growth under all assayed conditions (Fig. 2). In contrast, the strain deleted for the cyclase (Δrc1_3783) failed to encyst, except when supplemented with cGMP at a final concentration of 50 μM, which allowed production of wild-type levels of cysts (Fig. 2). Although Fig. 2 shows complementation with 50 μM cGMP, effective complementation of the cyclase deletion also occurs with as little as 5 μM cGMP (data not shown). Addition of 50 μM cAMP, or any other adenine or guanine nucleotide (AMP, ADP, ATP, GMP, GDP, and GTP), did not promote induction of cysts in the cyclase mutant (Fig. 2; Supplementary Fig. S1). With respect to the CRP homolog mutant (Δrc1_3788), there was no discernable response to the addition of cGMP nor to any other nucleotide (cAMP, AMP, ADP, ATP, GMP, GDP, and GTP), as colonies from these plates were identical in appearance to those grown in the absence of nucleotides and contained only vegetative cells (Fig. 2; Supplementary Fig. S1). Interestingly, cyst formation is also rapidly induced in both the wild-type and cyclase mutant strains by the addition of 200 μM cGMP to vegetative CENS media, which is a growth medium that normally does not induce cyst formation (Fig. 4). Again, the strain deleted for the CRP homolog (Δrc1_3788) remained unable to induce development of cysts (Fig. 4).
Figure 4.
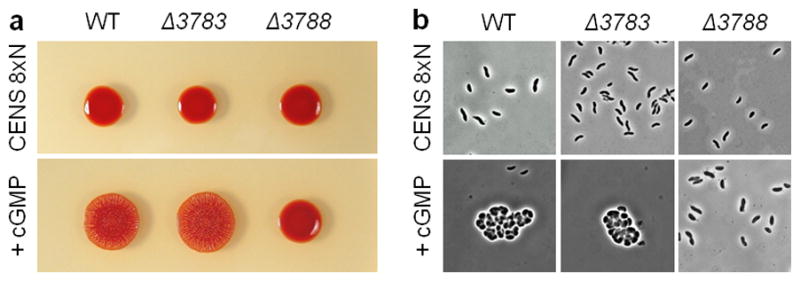
Analyses of cyst cell formation in wild-type, Δrc1_3783 (nucleotidyl cyclase) and Δrc1_3788 (CRP homolog) strains after 3 days growth on agar solidified CENS vegetative media. Media supplemented with cGMP had a final concentration of 200 μM. (a) Top panel shows shiny vegetative colonies forming on CENS media whereas the bottom panel shows dry ridged cyst forming colonies from wild-type and Δrc1_3783 strains grown on CENS medium containing 200 μM cGMP (b) Microscopic observations of cyst and vegetative cells from the above panels.
ORF rc1_3783 Encodes a Guanylyl Cyclase
An alignment of the catalytic core of the R. centenum cyclase with previously characterized eukaryotic and prokaryotic cyclases shows that several key catalytic amino acids are not present in the R. centenum cyclase (Fig. 1c). Specifically, while two critical metal-ion binding aspartates are conserved (D41, D85), the transition state stabilizing asparagine and arginine residues are substituted by histidine (H157) and lysine (K161). In addition, glutamine (Q81) and proline (P157) residues occupy substrate specifying positions, neither of which are similar to residues that are classic predictors of purine selectivity (Linder, 2005). The observed cellular response to cGMP, coupled with the presence of ambiguous residues at substrate specifying positions, prompted us to ascertain substrate specificity and product formation by the R. centenum cyclase coded by ORF rc1_3783. For this analysis we expressed and purified a His-tagged version of this cyclase using an E. coli expression strain, and then used high pressure liquid chromatography to resolve adenine and guanine nucleotide substrates and products from enzymatic assays. The ability of the recombinant cyclase to utilize either ATP or GTP as a substrate was tested in the presence of either Mg2+ or Mn2+, with substrate and product retention times compared with nucleotide standards (Fig. 5a, 5e). Synthesis of cAMP was not observed in reactions containing ATP and Mg2+ (Fig. 5c), whereas synthesis of cGMP was observed in an identical assay using GTP as a substrate (Fig. 5g). In reactions containing Mn2+ the production of cGMP was greatly enhanced (Fig. 5h), with cGMP increasing concomitantly with GTP substrate reduction over time. In a reaction containing ATP and Mn2+ a small peak corresponding to cAMP was observed (Fig. 5d), though cAMP formation was significantly less than the amount of cGMP produced either in the presence of Mg2+ or Mn2+. Small amounts of nucleotide mono and diphosphates observed in all assays were attributed to substrate degradation. Prolonged incubation of GTP at elevated temperatures has been reported to cause non-enzymatic formation of cGMP (Kimura and Murad, 1974). Since our reactions were terminated by heat denaturation, we addressed whether heat significantly contributed to spontaneous cGMP formation with a control reaction lacking enzyme, which produced no detectable cGMP (Fig. 5d).
Figure 5.

HPLC separation of products and substrates from assays using purified recombinant guanylyl cyclase (RC1_3783). (a) Separation of adenine nucleotide standards AMP (1), ADP (2), cAMP (3) and ATP (4). (e) Separation of guanine nucleotide standards GMP (1), GDP (2), cGMP (3) and GTP (4). All reactions were incubated for 2 hr and contained 0.5 mM ATP (b–d) or GTP (f–h), 10 mM MnCl2 (b, d, f, h) or MgCl2 (c, g) and 0.5 μg/μl purified RC1_3783 (c, d, g, h). Control reactions (b, f) had buffer added in place of enzyme. A small amount of cGMP indicated by a black arrow (↓) is produced in reaction g containing GTP, Mg2+ and recombinant cyclase, with greater activity occurring in reaction h when Mn2+ is substituted for Mg2+. (i) Purified recombinant cyclase elutes as a dimer in the absence of NaCl (Blue line) but as a monomer when chromatographed on a Superose 6 column equilibrated in 100, 50 or 25 mM NaCl (Black, Green & Red lines, respectively).
We observed that cyclase activity was salt inhibited, as reactions lacking NaCl produced ~3x more cGMP than those containing 100 mM NaCl (data not shown). Since class III cyclases are active as dimers, we assessed the effect of salt on oligomerization by size exclusion chromatography, which showed that purified cyclase eluted as a monomer in columns equilibrated with 25, 50 and 100 mM NaCl, and as a dimer in the absence of NaCl (Fig. 5i).
The CRP homolog RC1_3788 Preferentially Binds cGMP
We also characterized the cyclic nucleotide binding abilities of the CRP-like homolog coded by ORF rc1_3788. For this analysis the CRP homolog was purified after overexpression from an E. coli expression strain and assayed for binding of cyclic nucleotides using both differential scanning fluorimetry (DSF) and isothermal titration calorimetry (ITC). DSF measures protein thermal denaturation, or melting temperature (Tm), which is often stabilized upon binding to a substrate (Matulis et al., 2005; Niessen et al., 2007). Using this technique the Tm of the CRP homolog in the absence of a ligand was measured at 67°C. The thermal stability of RC1_3788 was significantly and maximally enhanced by the addition of 40 μM cGMP, which increased the overall Tm to 82°C (Supplementary Fig. S2; Table 2). In stark contrast, an equivalent concentration of cAMP increased the overall Tm by only 1.5°C (Supplementary Fig. S2; Table 2).
Table 2.
Changes in melting temperature (Tm) of the CRP homolog RC1_3788 in the presence of increasing cAMP or cGMP relative to a sample lacking ligand, as determined by DSF.
| [Ligand] μM | ΔTm (°C) RC1_3788+cGMP |
ΔTm (°C) RC1_3788+cAMP |
|---|---|---|
| 0 | 0 | 0 |
| 10 | 8.5 | 0.5 |
| 20 | 12.0 | 0.9 |
| 30 | 14.0 | 1.5 |
| 40 | 14.4 | 1.5 |
| 50 | 14.4 | 2.0 |
Using ITC analysis for substrate binding, we observed a biphasic titration curve by the addition of cGMP (Supplementary Fig. S3; Table 3). The initial binding event is characterized by exothermic enthalpy (ΔH = − 0.9 ± 0.2) followed by an endothermic event (ΔH = 7.2 ± 0.2). The observed raw data was best fitted to a two-site sequential model, described in Experimental Procedures. Binding of cGMP exhibited a negative cooperativity (Kd1 < Kd2) with a stoichiometry of 2 cGMP per peptide dimer and apparent Ka1 (2.6 ± 0.6 × 107 M−1) and Ka2 (0.8 ± 0.2 × 107 M−1) (Table 3) similar to those observed for the binding of cAMP to E. coli CRP (Lin and Lee, 2002a). Indeed the exothermic and endothermic binding events, negative cooperativity and Ka values are surprisingly similar to those previously reported for cAMP binding to CRP from E. coli (Gorshkova et al., 1995).
Table 3.
Binding affinities of CRP homolog RC1_3788 for cAMP and cGMP, as determined by ITC.
| Ligand | Model | n | ΔH (kcal mol−1) | Kobs (×107 M−1) | ΔS (kcal mol−1 K−1) | ΔG (kcal mole−1) |
|---|---|---|---|---|---|---|
| cAMP | One-site binding | 0.5 ± 0.03 | 3.3 ± 0.4 | 0.03 ± 0.01 | 0.04 ± 0.4 | − 7.5 ± 0.2 |
| cGMP | Two-site sequential binding* | 1** 1** |
−0.9 ± 0.2 7.2 ± 0.2 |
2.6 ± 0.6 0.8 ± 0.2 |
0.03 ± 0.2 0.1 ± 0.3 |
− 10.1 ± 0.1 − 9.4 ± 0.1 |
Assumes an integral stoichiometry value of n = 1;
According to this model, n1=n2=1.
Interestingly, the R. centenum CRP homolog exhibits slight, albeit much poorer, binding to cAMP with a binding constant (Ka) of 0.03 ± 0.01 × 107 M−1 (Supplementary Fig. S3; Table 3). E. coli CRP also exhibits the capability of binding to cGMP, but at a much reduced affinity to that of cAMP (Gorshkova et al., 1995; Lin and Lee, 2002b). Therefore, the R. centenum CRP-like homolog and E. coli CRP bind both cAMP and cGMP, with the caveat that R. centenum preferentially binds cGMP over that of cAMP while E. coli CRP preferentially binds cAMP over cGMP. Thus, these homologs appear to have “swapped” cyclic nucleotide substrate specificities.
The Nucleotidyl Cyclase Gene Cluster, and the ability to produce cGMP, Is Conserved In Other Soil Inhabiting Species
Searching the NCBI database, we discovered homologs of the R. centenum guanylyl cyclase, and of ORF rc1_3786 and ORF rc1_3787, clustered in several plant associated soil bacteria (Fig. 6a). Interestingly, Azospirillum sp. B510, which has a very similar life cycle as R. centenum (Berleman and Bauer, 2004), has linked homologs of the R. centenum cyclase, ORFs rc1_3786 and rc1_3787 as well as a homolog to the CRP family transcription factor coded by ORF rc1_3788 (Fig. 6a). Conservation of this gene cluster suggests that other soil inhabiting species may also produce cGMP and that ORFs rc1_3786 and rc1_3787 may be involved in regulating the production, transport or stability of cGMP. To test these possibilities we constructed in frame deletions of ORFs rc1_3786 and rc1_3787 in R. centenum and assayed these mutants for their ability to produce cysts. Deletions of either rc1_3786 or rc1_3787 results in a phenotype that is indistinguishable from the strain deleted for the cyclase. Specifically, these additional deletion strains are defective in forming cysts on CENS-8xN cyst inducing medium, and this defect can be complemented by exogenous addition of cGMP at a final concentration of 50 μM (Fig. 7a, 7b). Furthermore, deletion of either of these ORFs also results in defects in cGMP production when grown in liquid cyst inducing CENBA media (Fig. 7c). Thus, in vivo production of cGMP in R. centenum requires three gene products coded by ORFs rc1_3783, rc1_3786 and rc1_3787.
Figure 6.

(a) Species containing cyclases with similar genetic contexts as the R. centenum guanylyl cyclase coded by gcyA (rc1_3783), the downstream ORFs needed for cyclase activity (rc1_3786 and rc1_3787) and the CRP homolog coded by cgrA (rc1_3788) in Azosprillum sp. B510. Red (nucleotidyl cyclase), green (COG1944), orange (COG3482) and blue (CRP homolog). (b) Amino acid sequence alignment of the RC1_3783 cyclase domain with putative guanylyl cyclase domains. Species are: Rhodospirillum centenum, RC1_3783 (YP_002299938); Azospirillum sp. B510, AZL_d00190 (YP_003452389); Sinorhizobium meliloti 1021, SMc01491 (NP_386201); Mesorhizobium loti MAFF303099, mll0576 (NP_102350); Rhizobium leguminosarum bv. viciae 3841, pRL110256 (YP_771288); Rhizobium etli CIAT 652, RHECIAT_PA0000121 (YP_001985730); Rhizobium sp. NGR234, NGR_c20030 (YP_002826519); Sinorhizobium medicae WSM419, Smed_1991 (YP_001327660). Domains were identified using the SMART database and aligned in MEGA v.4.0 utilizing a PAM matrix with respective pair-wise and multiple alignment gap opening and extension penalties of 10 and 0.1. Relative positions of residues involved in metal binding (Yellow), substrate specification (Blue) and transition state stabilization (Red) are additionally indicated by asterisks (*), inverted triangles (▼) and arrows (↓). Further shading indicates identically and functionally conserved residues at 100% (Black), 80% (Dark gray, White letters), and 60% (Light gray, Dark letters) conservation.
Figure 7.

Analyses of cyst cell formation and cGMP production in wild-type, Δrc1_3786 and Δrc1_3787 deletion strains. (a) After 3 days growth on cyst inducing CENS-8xN media the wild-type strain displayed ridged colony morphology indicative of cyst cell formation, while both deletion strains remained vegetative in appearance. In contrast, on a plate supplemented to 50 μM cGMP, all three strains displayed encysted colony morphologies. (b) Microscopic observations of vegetative and cyst cells from colonies shown in panel a. (c) Exogenous cGMP levels present in culture supernatants after 11 days growth in liquid CENBA media.
The presence of linked homologs of rc1_3783, rc1_3786 and rc1_3787 in the genomes of several species that interact with plant roots suggest that the ability to produce cGMP may be a conserved trait of this bacterial niche. To test this possibility, we assayed for cGMP production in Azospirillum brasilense cultures that were grown in cyst inducing MSM flocculation medium (Neyra and Vasnberkum, 1977). We observed accumulation of up to ~4 nM of cGMP in the culture supernatant that was concurrent with the formation of cysts in this species (Fig. 8).
Figure 8.

Accumulation of exogenous cGMP in culture supernatants of wild-type Azospirillum brasilense grown in cyst inducing MSM medium. Cultures were grown aerobically in MSM media and the culture supernatant was measured for cGMP levels as described in Materials and Methods. Error bars represent standard deviation derived from three replicate assays.
Discussion
While cAMP is common in all three kingdoms, the existence of cGMP has only been reported for a few bacteria (Bernlohr et al., 1974; Kurn et al., 1977; Setlow and Setlow, 1978; Ho and McCurdy 1980; Bhatnagar et al., 1984; Lim et al., 1979). There is also scant biochemical evidence as to any role cGMP has in controlling a prokaryotic cellular process, thus the importance of cGMP in bacteria has remained controversial. The only other example of a biochemically characterized bona fide prokaryotic guanylyl cyclase is a class III enzyme from Synechocystis PCC 6803 that has been crystallized, but as yet has an ill defined role (Rauch et al., 2008). Our study of the role of cGMP in controlling R. centenum encystment thus represents the first example of prokaryote that produces cGMP with a clearly defined role in controlling a prokaryotic process. As such, we designate ORF rc1_3783 as gycA for guanyl cyclase and ORF rc1_3788 as cgrA for cyclic GMP regulator.
R. centenum secretes large quantities of cGMP into the extracellular environment as it transitions to cyst formation. Interestingly, cAMP is secreted by several bacterial species; for example, E. coli secretes >99.9% of produced cAMP under certain conditions (Matin and Matin, 1982), the reason why this occurs remains unclear. Cyanobacteria also secrete cAMP, including Spirulina platensis (Sakamoto et al., 1991), which displays increased mat formation and gliding motility in response to exogenously added cAMP (Ohmori et al., 1992). In Streptomyces coelicolor, extracellular cAMP peaks during spore germination and again during aerial mycelium and actinorhodin production suggesting that cAMP may be a diffusible signal that coordinates production of this antibiotic (Susstrunk et al., 1998).
Our working model for cGMP signaling in R. centenum involves the synthesis of cGMP in response to a signal related to nutrient deprivation that activates the guanylyl cyclase (Fig. 9). The mechanism by which the R. centenum cyclase is activated is not immediately apparent, as the only other predicted domain present is a partial ATPase domain of the COG3899 functional group and several TPR protein-protein interaction domains at the C-terminus. While domains of known function are not present in the products of ORFs rc1_3786 and rc1_3787, it seems likely that they interact and form a complex with the cyclase as disruptions of either of these ORFs gives rise to the same hypo-cyst phenotype that, like the guanylyl cyclase disrupted strain, can be rescued by cGMP. The majority of cGMP produced by the guanylyl cyclase appears to be secreted, and is presumably imported back into R. centenum via an unknown transport mechanism to allow intracellular cGMP levels to rise to a level where it binds to the CRP homolog to activate genes required for encystment. That most of the synthesized cGMP is secreted also indicates that cGMP production likely functions as a signal for a coordinated community wide change in cell type.
Figure 9.
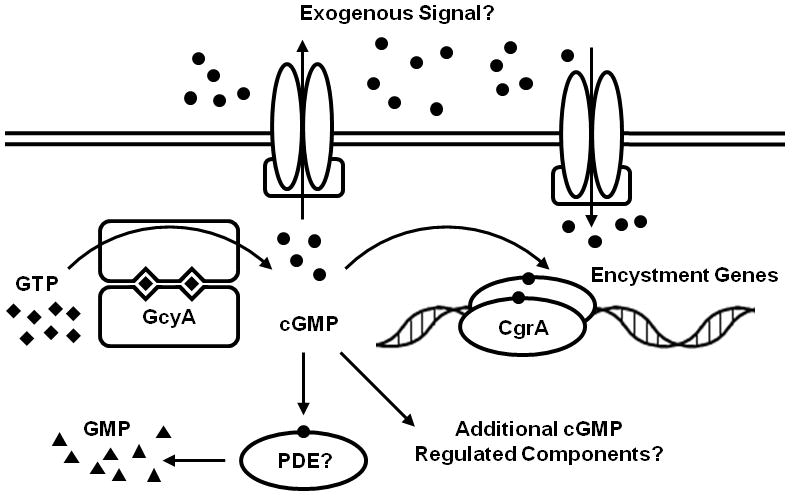
Model of the regulation of R. centenum encystment by cGMP. The guanylyl cyclase coded by gcyA (rc1_3783) synthesizes cGMP in response to an unknown developmental signal. The cNMP binding domain of the CRP homolog coded by cgrA (rc1_3788) binds and is activated by cGMP, resulting in expression of genes required
Bioinformatic approaches have uncovered putative prokaryotic guanylyl cyclases similar to Cya2 of Synechocystis, in other cyanobacteria and more diverse species including Congregibacter litoralis, Mariprofundus ferrooxydans, Roseobacter denitrificans (γ and α–proteobacteria, respectively) and Mycobacterium ulcerans (an actinobacterium) (Rauch et al., 2008; Biswas et al., 2009; Wu et al., 2008). A constraining parameter in the identification of these enzymes is the presence of the substrate specifying glutamate residue, which was a unifying feature of all previously characterized guanylyl cyclases. However, the R. centenum guanylyl cyclase contains a glutamine (Q81) substituted for the substrate specifying glutamate and a proline (P150) at a second substrate binding position. Likewise, asparagine and arginine residues thought to be involved in transition state stabilization are replaced by histidine (H157) and lysine (K161), respectively, in the R. centenum cyclase. That these changes are present in the R. centenum guanylyl cyclase highlights the fact that there is an incomplete understanding of substrate specification in class III cyclases. Indeed, our demonstration that one of these non-canonical cyclases is a second identified bona fide prokaryotic guanylyl cyclase suggests that prokaryotic guanylyl cyclases may be more common than previously believed, especially given that there are abundant uncharacterized microbial cyclases that exhibit diverse substitutions in substrate specifying residues (Shenroy and Visweswariah, 2004).
By performing genomic context searches using the two R. centenum ORFs rc1_3786 and rc1_3787, which are needed to observe in vivo cGMP production, we are intrigued to find that linked homologs of these genes are associated with nucleotidyl cyclases in the genomes of several sequenced Rhizobial species as well as in Azospirillum, that produces cysts (Fig. 6a). An alignment of the catalytic cores from these cyclases revealed a variety of canonical catalytic residue deviations (Fig. 6b), with cyclases from Sinorhizobium meliloti (SMc01491), Sinorhizobium medicae (Smed_1991) and Rhizobium sp. NGR234 having a proline substitution at the second substrate specifying position like the R. centenum cyclase. We have demonstrated that A. brasilense synthesizes and excretes cGMP as cells transition from vegetative to cyst cells (Supplemental Fig. S4), suggesting that the cyst developmental cycle of this related species may be similarly regulated by cGMP. We have also observed that S. meliloti is also capable of synthesizing cGMP (data not shown), and it has been recently reported that this species is capable of making desiccation resistant cysts not unlike that of R. centenum and A. brasilense (Pogorelova et al., 2009). This suggests that the use of cGMP as a signal to control cyst formation may be a conserved trait among several species that interact with plants.
Experimental Procedures
Strains and Growth Conditions
E. coli strains DH5α, DH10B and SCS1101 were used for general cloning, BL21 Rosetta II (Invitrogen) for protein overexpression and S17-1 (λpir) for plasmid conjugation. E. coli strains were grown and cultured on agar-solidified or liquid Luria-Bertani (LB) media at 37 or 16°C with appropriate, antibiotics. The parental R. centenum strain used in this study was a Δche3 derivative of wild-type R. centenum (ATCC 51521). R. centenum strains were cultured aerobically in CENS or CENBA liquid media (Statwald-Demchick et al., 1990) at 37°C, or grown at 42°C on agar-solidified CENS or CENS-8xN, which is CENS supplemented with 8 mM NH4Cl. A. brasilense (ATCC 29145) cells were grown on agar-solidified NB, or cultured aerobically in liquid NB or MSM media (Neyra and Vanberkum, 1977) at 37°C.
Desiccation Resistance Analyses
R. centenum colonies were harvested from CENS-8xN plates after 3 days growth and resuspended in 1 ml phosphate buffer. Cell suspensions were sonicated 2x for 5s at low power to disperse cellular aggregates as needed. Total viable cell counts were enumerated from serial dilutions pipetted onto CENS plates incubated at 42°C for 2 days. Cysts cells were quantified by pipetting identical dilutions onto 0.45 um pore sized filters (Grade: A045FO47A; Advantec), which were dried briefly, then desiccated at 42°C for 3 days. Filters were subsequently transferred to CENS plates and incubated at 42°C for 2 days to allow surviving cell outgrowth. All assays were repeated in triplicate, and desiccation resistance was calculated as total viable cells/ total desiccation resistant cells.
Hypo-cyst Suppression Screen
Transposon mutagenesis and the construction of in-frame deletion mutants of rc1_3783, rc1_3786, rc1_3787 and rc1_3788 were performed as described by Berleman et al, (2004). Mutagenesis of the Δche3 strain was performed as previously described by Berleman and Bauer (2004) and Berleman et al., (2004). Briefly, a modified mini-Tn5 spectinomycin-resistant (Spr) interposon was delivered to a hyper-cyst Δche3 R. centenum strain (Berleman et al, 2004) on plasmid pZJD17 from E. coli S17-1 (λS17) using a filter mating procedure. Overnight R. centenum and E. coli cultures were washed 3x with CENS to remove antibiotics, then were applied in a 5:1 ratio to a 0.45 μm pore sized filter (No. 245-0045; Nalgene). After 4 hrs incubation at 37°C on an antibiotic free CENS plate, cells were resuspended in 5 ml CENS and spread onto CENSKmSp plates in 200 μl aliquots. Spr transconjugates displaying suppression to vegetative growth after 72 hours were selected for further scrutiny of colonial encystment morphologies. Cells from overnight cultures were washed 3x and resuspended in 1/20 volume of phosphate buffer (40 mM KH2PO4/K2HPO4, pH 7), pipetted in 5 μl aliquots onto CENS and CENBA plates that were incubated for 3 and 6 days respectively.
Interposon insertion loci from mutants growing vegetatively on both media were mapped on the chromosome by isolating and digesting mutant genomic DNA with the PstI restriction enzyme and ligating fragments into the corresponding site in pBluescript SK+. Plasmids were electroporated into E. coli DH10B and transformants with a chromosomal DNA fragment containing a Spr interposon were selected for by growth on LBAmpSp. Transformant plasmid DNA was used in sequencing reactions with primers specific to the 5′ and 3′ ends of the interposon (Supplementary Table S1). Resultant DNA sequences were then used in BLAST searches against the R. centenum genome to identify disrupted loci. Domains in RC1_3783 and RC1_3788 were identified by SMART and NCBI Conserved Domain Databases, with the TPR domains in RC1_3783 identified using TPRpred.
Quantification of cyclic nucleotides in bacterial cultures
Wild-type, Δrc1_3783, Δrc1_3786, Δrc1_3787 and Δrc1_3788 R. centenum cells taken from agar solidified CENS plates were resuspended in phosphate buffer to an OD600 of ~2.0, 5 μl of which was then subcultured into 25 ml of liquid CENBA medium and cultured at 37°C with shaking at 250 rpm. Measurements of extracellular cGMP produced in wild-type, Δrc1_3783 and Δrc1_3788 cells were from supernatants of liquid cultures. Culture supernatants were treated with 1/100 volume of concentrated HCl, centrifuged at 13,000 rpm at room temperature and directly assayed by EIA. Cell pellets were washed 2x and resuspended in 0.5 ml of phosphate buffer, lysed by sonication and then clarified by centrifugation. The supernatant was mixed with an equal volume of 10% ice-cold trichloroacetic acid (TCA) and kept on ice for 30 min with TCA-insoluble material removed by centrifugation. TCA was then extracted from the supernatant by three successive washes with 2 volumes of diethyl ether. The final aqueous phase was lyophilized and resuspended in 450 μl 0.1M HCl with concentrations of cAMP and cGMP determined with commercially available immunoassay kits (AssayDesigns).
In Frame Deletions of rc1_3783, rc1_3786, rc1_3787 & rc1_3788
Gene flanking 500 bp fragments were PCR amplified using wild-type cells as template for colony PCR and primer sets detailed in Supplementary Table S1. PCR amplified fragments were separately cloned and sequenced in either pTOPO Blunt-II or pGEM-T vectors. The rc1_3783 and rc1_3788 fragments were assembled in pUC18 prior to being sub-cloned into the suicide vector pZJD29a, whereas fragments for the rc1_3786 and rc1_3787 constructs were ligated directly into pZJD29a. Plasmids were mated into wild-type R. centenum from E. coli S17-1 (λpir), and initial recombinants were selected for on CENSGm agar plates. Second recombinants carrying gene deletions were later identified through phenotypic (GmS/SucR) and colony PCR analyses.
Cloning, Overexpression and Purification RC1_3783 & RC1_3788
rc1_3783 was amplified in two separate fragments with primers rc1_3783NdeI-f, rc1_3783SbfI-r, rc1_3783SbfI-f and rc1_3783XhoI-r, whereas rc1_3788 was cloned as one fragment with the primers rc1_3788NdeI-f and rc1_3788XhoI-r (Supplemental Table S1). PCR fragments were gel-purified, cloned and sequenced in pTOPO-Blunt II. Fragments where then sub-cloned into pET28a vectors in ligation reactions using the external NdeI & XhoI restriction sites, and were internally joined in rc1_3783 by a SbfI site common to both fragments. Each construct was sequenced and transformed into E. coli Rosetta II (DE3).
For overexpression of RC1_3783, an overnight culture of the Rosetta II cells containing the RC1_3783 expression plasmid was sub-cultured 1/50 into 2 L LBKmCm, grown to an OD600 ~0.6, then induced at 0.2 mM IPTG and grown overnight at 16°C. Cells were pelleted by centrifugation at 6000 rpm for 5 minutes, washed once with 20 mM Tris 100mM NaCl pH 8.0, then repelleted and resuspended in 40 ml Binding Buffer (20 mM Tris, 100 mM NaCl, 25 mM Imidazole, pH 8). Cells were lysed by four passes through an M-110L Micro Fluidizer Processor at ~2000 p.s.i. (Microfluidics). Insoluble matter was removed by centrifugation at 15,000 rpm for 30 minutes and the decanted supernatant was incubated with 4 ml Ni2+ charged resin (Novagen) with gentle shaking for ~2 hrs. After 5 minutes of centrifugation at 1000 rpm, the pelleted resin was washed 3x with ~45 ml wash buffer (20 mM TrisHCl, 100 mM NaCl, 50 mM Imidazole, pH 8) and then loaded onto a 25 ml disposable column (BioRad). The resin was washed with an additional 20 ml wash buffer and bound protein was then eluted with elution buffer (20 mM TrisHCl, 100 mM NaCl, 200 mM Imidazole, pH 8). Elution fractions were concentrated in a 30k Centriprep Concentrator (Millipore), then was loaded onto a hand-packed Superose 12 size exclusion column (GE Healthcare) equilibrated in binding buffer lacking imidazole. After concentration, dialysis into storage buffer (20 mM Tris-HCl, 100 mM NaCl, 50% glycerol, pH 8) and determination of protein concentration by a Bradford Assay (BioRad), and stored at −20°C until use.
Overexpression and purification conditions for RC1_3788 were essentially identical to those of RC1_3783, with the exception that all buffers had a pH 7.5, and the size exclusion purification step differed. The size exclusion equilibration buffer additionally contained 5% glycerol when RC1_3788 was to be used for DSF analyses, whereas a separate buffer (50 mM KH2PO4/K2HPO4, 200 mM KCl, 0.2 mM Na2EDTA, 0.2 mM DTT, 5% glycerol, pH 7) was used for protein to be used in ITC experiments. RC1_3788 was stored at 4°C and used within one week of purification.
Adenylyl and Guanylyl Cyclase Assays
Adenylyl and guanylyl cyclase reactions were undertaken in 100 μl reactions containing 0.5 μg/μl purified RC1_3783, 20 mM Tris (pH 8.0), 100 mM NaCl, 0.5 mM ATP or GTP and 10 mM MnCl2 or MgCl2. Reactions were incubated at 25°C, stopped by heating at 75°C for 10 min and then clarified by centrifugation at 15,000 rpm for 10 min. HPLC separation of nucleotides was accomplished as described by Gebelein et al., (1992).
ITC and DSF Determination of RC1_3788 Cyclic Nucleotide Binding
Sodium salts of cAMP and cGMP (Sigma) were dissolved in RC1_3788 storage buffer and quantitated using molar coefficients of 1.23 × 104 cm M−1 at 260 nm for cAMP and 1.34 × 104 cm M−1 at 250 nm for cGMP. All reactions contained 10 μM RC1_3788, 2.5x SYPRO Orange dye (Cat. S6650; Invitrogen) and cyclic nucleotide concentrations of 0–80 μM (cAMP) or 0–50 μM (cGMP) tested in at least three independent assays with control reactions of protein alone, protein without ligand and ligand alone. DSF experiments were performed using an Eppendorf Mastercycler EP Realplex thermal cycler, with excitation and emission at 470 and 550 nm, respectively. Denaturation curves were generated as a function of (−R′ (T)) for every °C between 25–95°C, and Tm values.
For ITC analyses, both RC1_3788 and cyclic nucleotide stocks were equilibrated in degassed assay buffer (50 mM KH2PO4/K2HPO4, 200 mM KCl, 0.2 mM Na2EDTA, 0.2 mM DTT, 5% glycerol, pH 7). All experiments were performed at 25°C using a Microcal VP-ITC calorimeter with a stirring speed of 310 rpm. A minimum of two titrations were carried out for each ligand and the experimental values reported are the average of the individual best-fit values collected using a one-site or sequential binding model (ITC Data Analysis in Origin® v.7). The best fit to data produces a binding constant (Ka), enthalpy (ΔH), entropy (ΔS), and reaction stoichiometry (n). ΔG and ΔS were then calculated using thermodynamic definition, ΔG = − RT ln K and ΔG = ΔH − TΔS. The observed raw data was best described by the following reactions:
cAMP + RC1_3788(dimer) ⇌ cAMP·RC1_3788(dimer)
Ka = [cAMP·RC1_3788(dimer)]/([RC1_3788(dimer)][cAMP])
cGMP + RC1_3788(dimer) ⇌ cGMP·RC1_3788(dimer)
Ka1 = [cGMP·RC1_3788(dimer)]/([RC1_3788(dimer)][cGMP])
cGMP + cGMP·RC1_3788(dimer) ⇌ cGMP2·RC1_3788(dimer)
Ka2 = [cGMP2·RC1_3788(dimer)]/([cGMP·RC1_3788(dimer)][cGMP])
2cGMP + RC1_3788(dimer) ⇌ cGMP2·RC1_3788(dimer)
Koverall = [cGMP2·RC1_3788(dimer)]/([ RC1_3788(dimer)][cGMP2])
Supplementary Material
Acknowledgments
We thank Dr. Yves V. Brun for the use of his microscope facility; Dr. Todd Stone, Facility Manager of the Indiana University Physical Biochemistry Instrumentation Facility and Dr. Jonathan A. Karty, Facility Manager of the Indiana University Chemistry Department Mass Spectrometry Facility, for training and assistance with instrumentation. This work was supported by the Metabolomics and Cytomics Initiative (MetaCyte), which is supported by a major grant from the Lilly Endowment, Inc. The second (Q.D.) and third (S.R.) authors contributed equally to this study.
References
- Berleman JE, Bauer CE. Characterization of cyst cell formation in the purple photosynthetic bacterium Rhodospirillum centenum. Microbiology. 2004;150:383–390. doi: 10.1099/mic.0.26846-0. [DOI] [PubMed] [Google Scholar]
- Berleman JE, Bauer CE. Involvement of a Che-like signal transduction cascade in regulating cyst cell development in Rhodospirillum centenum. Mol Microbiol. 2005;56:1457–1466. doi: 10.1111/j.1365-2958.2005.04646.x. [DOI] [PubMed] [Google Scholar]
- Berleman JE, Hasselbring BM, Bauer CE. Hypercyst mutants in Rhodospirillum centenum identify regulatory loci involved in cyst cell differentiation. J Bacteriol. 2004;186:5834–5841. doi: 10.1128/JB.186.17.5834-5841.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bernlohr RW, Haddox MK, Goldberg ND. Cyclic guanosine 3′:5′-monophosphate in Escherichia coli and Bacillus lichenformis. J Biol Chem. 1974;249:4329–4331. [PubMed] [Google Scholar]
- Biswas KH, Shenoy AR, Dutta A, Visweswariah SS. The evolution of guanylyl cyclases as multidomain proteins: conserved features of kinase-cyclase domain fusions. J Mol Evol. 2009;68:587–602. doi: 10.1007/s00239-009-9242-5. [DOI] [PubMed] [Google Scholar]
- Bhatnagar NB, Bhatnagar R, Venkitasubramanian TA. Characterization and metabolism of cyclic guanosine 3′5′-monophosphate in Mycobacterium smegmatis. Biochem Biophys Res Comm. 1984;121:634–640. doi: 10.1016/0006-291x(84)90229-8. [DOI] [PubMed] [Google Scholar]
- Blomfield IC, Vaughn V, Rest RF, Eisenstein BI. Allelic exchange in Escherichia coli using the Bacillus subtilis sacB gene and a temperature-sensitive pSC101 replicon. Mol Microbiol. 1991;5:1447–1457. doi: 10.1111/j.1365-2958.1991.tb00791.x. [DOI] [PubMed] [Google Scholar]
- Cadoret JC, Rousseau B, Perewoska I, Sicora C, Cheregi O, Vass I, Houmard J. Cyclic nucleotides, the photosynthetic apparatus and response to a UV-B stress in the Cyanobacterium Synechocystis sp. PCC 6803. J Biol Chem. 2005;280:33935–33944. doi: 10.1074/jbc.M503153200. [DOI] [PubMed] [Google Scholar]
- Clark VL, Bernlohr RW. Guanyl cyclase of Bacillus licheniformis. Biochem Biophys Res Comm. 1972;46:1570–1575. doi: 10.1016/0006-291x(72)90787-5. [DOI] [PubMed] [Google Scholar]
- Cook WR, Kalb VF, Peace AA, Bernlohr RW. Is cyclic guanosine 3′,5′-Monophosphate a cell-cycle regulator? J Bacteriol. 1980;141:1450–1453. doi: 10.1128/jb.141.3.1450-1453.1980. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cotta MA, Whitehead TR, Wheeler MB. Identification of a novel adenylate cyclase in the ruminal anaerobe, Prevotella ruminicola D31d. FEMS Microbiol Lett. 1998;164:257–260. doi: 10.1111/j.1574-6968.1998.tb13095.x. [DOI] [PubMed] [Google Scholar]
- Danchin A. Phylogeny of adenylyl cyclases. Adv Second Messenger Phosphoprotein Res. 1993;27:109–162. [PubMed] [Google Scholar]
- Devi AL, McCurdy HD. Adenylate-Cyclase and Guanylate-Cyclase in Myxococcus xanthus. J Gen Micro. 1984;130:1851–1856. [Google Scholar]
- Fani R, et al. Phylogeny of the genus Azospirillum based on 16S rDNA sequence. FEMS Microbiol Lett. 1995;129:195–200. doi: 10.1111/j.1574-6968.1995.tb07579.x. [DOI] [PubMed] [Google Scholar]
- Favinger J, Stadtwald R, Gest H. Rhodospirillum centenum, sp. nov., a thermotolerant cyst-forming anoxygenic photosynthetic bacterium. Antonie Van Leeuwenhoek. 1989;55:291–296. doi: 10.1007/BF00393857. [DOI] [PubMed] [Google Scholar]
- Finn JT, Grunwald ME, Yau KW. Cyclic nucleotide-gated ion channels: an extended family with diverse functions. Annu Rev Physiol. 1996;58:395–426. doi: 10.1146/annurev.ph.58.030196.002143. [DOI] [PubMed] [Google Scholar]
- Francis SH, Corbin JD. Structure and function of cyclic nucleotide-dependent protein kinases. Annu Rev Physiol. 1994;56:237–272. doi: 10.1146/annurev.ph.56.030194.001321. [DOI] [PubMed] [Google Scholar]
- Gebelein M, Merdes G, Berger MR. Nucleotide preparation from cells and determination of nucleotides by ion-pair high-performance liquid chromatography. J Chromatogr. 1992;577:146–150. doi: 10.1016/0378-4347(92)80610-3. [DOI] [PubMed] [Google Scholar]
- Gorshkova I, Moore JL, McKenney KH, Schwarz FP. Thermodynamics of cyclic nucleotide binding to the cAMP receptor protein and its T127L mutant. J Biol Chem. 1995;270:21679–21683. doi: 10.1074/jbc.270.37.21679. [DOI] [PubMed] [Google Scholar]
- Harman JG. Allosteric regulation of the cAMP receptor protein. Bba-Protein Struct M. 2001;1547:1–17. doi: 10.1016/s0167-4838(01)00187-x. [DOI] [PubMed] [Google Scholar]
- Heikaus CC, Pandit J, Klevit RE. Cyclic nucleotide binding GAF domains from phosphodiesterases: structural and mechanistic insights. Structure. 2009;17:1551–1557. doi: 10.1016/j.str.2009.07.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Herdman M, Elmorjani K. Cyclic nucleotides. Methods in Enzymology. 1988;167:584–591. [Google Scholar]
- Ho J, McCurdy H. Sequential changes in the cyclic nucleotide levels and cyclic nucleotide phosphodiesterase activities during development of Myxococcus xanthus. Cur Micro. 1980;3:197–202. doi: 10.1007/BF02602447. [DOI] [PubMed] [Google Scholar]
- Iyer LM, Aravind L. The catalytic domains of thiamine triphosphatase and CyaB-like adenylyl cyclase define a novel superfamily of domains that bind organic phosphates. BMC Genomics. 2002;3:33. doi: 10.1186/1471-2164-3-33. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kimura H, Murad F. Nonenzymatic formation of guanosine 3′:5′-monophosphate from guanosine triphosphate. J Biol Chem. 1974;249:329–331. [PubMed] [Google Scholar]
- Kroos L. Bacterial development in the fast lane. J Bacteriol. 2008;190:4373–4376. doi: 10.1128/JB.00580-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kurn N, Shapiro L, Agabian N. Effect of carbon source and role of cyclic adenosine-3′,5′-monophosphate on Caulobacter cell-cycle. J Bacteriol. 1977;131:951–959. doi: 10.1128/jb.131.3.951-959.1977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lim ST, Hennecke H, Scott DB. Effect of cyclic guanosine 3′,5′-monophosphate on nitrogen fixation in Rhizobium japonicum. J Bacteriol. 1979;139:256–263. doi: 10.1128/jb.139.1.256-263.1979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lin SH, Lee JC. Communications between the high-affinity cyclic nucleotide binding sites in E. coli cyclic AMP receptor protein: effect of single site mutations. Biochemistry-Us. 2002a;41:11857–11867. doi: 10.1021/bi026099z. [DOI] [PubMed] [Google Scholar]
- Lin SH, Lee JC. Linkage of multiequilibria in DNA recognition by the D53H Escherichia coli cAMP receptor protein. Biochemistry-Us. 2002b;41:14935–14943. doi: 10.1021/bi026756n. [DOI] [PubMed] [Google Scholar]
- Linder JU. Substrate selection by class III adenylyl cyclases and guanylyl cyclases. IUBMB Life. 2005;57:797–803. doi: 10.1080/15216540500415636. [DOI] [PubMed] [Google Scholar]
- Lu YK, et al. Metabolic flexibility revealed in the genome of the cyst-forming alpha-1 proteobacterium Rhodospirillum centenum. BMC Genomics. 2010;11:325. doi: 10.1186/1471-2164-11-325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macchia V, Varrone S, Weissbach H, Miller DL. Guanylate cyclase in Escherichia coli. Purification and properties. J Biol Chem. 1975;250:6214–6217. [PubMed] [Google Scholar]
- Macchia V, Caputo G, Mandato E, Rocino A, Adhya S, Pastan I. Guanylate cyclase activity in Escherichia coli mutants defective in adenylate cyclase. J Bacteriol. 1981;147:931–934. doi: 10.1128/jb.147.3.931-934.1981. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Masuda S, Bauer CE. Null mutation of HvrA compensates for loss of an essential relA/spoT-like gene in Rhodobacter capsulatus. J Bacteriol. 2004;186:235–239. doi: 10.1128/JB.186.1.235-239.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matin A, Matin MK. Cellular levels, excretion, and synthesis rates of cyclic AMP in Escherichia coli grown in continuous culture. J Bacteriol. 1982;149:801–807. doi: 10.1128/jb.149.3.801-807.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matulis D, Kranz JK, Salemme FR, Todd MJ. Thermodynamic stability of carbonic anhydrase: Measurements of binding affinity and stoichiometry using ThermoFluor. Biochemistry-Us. 2005;44:5258–5266. doi: 10.1021/bi048135v. [DOI] [PubMed] [Google Scholar]
- Neyra CA, Vanberkum P. Nitrate Reduction and Nitrogenase Activity in Spirillum lipoferum. Can J Microbiol. 1977;23:306–310. doi: 10.1139/m77-045. [DOI] [PubMed] [Google Scholar]
- Niesen FH, Berglund H, Vedadi M. The use of differential scanning fluorimetry to detect ligand interactions that promote protein stability. Nat Protoc. 2007;2:2212–2221. doi: 10.1038/nprot.2007.321. [DOI] [PubMed] [Google Scholar]
- Ochoa de Alda JA, Houmard J. Genomic survey of cAMP and cGMP signalling components in the cyanobacterium Synechocystis PCC 6803. Microbiology. 2000;146:3183–3194. doi: 10.1099/00221287-146-12-3183. [DOI] [PubMed] [Google Scholar]
- Ochoa De Alda JA, Ajlani G, Houmard J. Synechocystis strain PCC 6803 cya2, a prokaryotic gene that encodes a guanylyl cyclase. J Bacteriol. 2000;182:3839–3842. doi: 10.1128/jb.182.13.3839-3842.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohmori K, Hirose M, Ohmori M. Function of Camp as a mat-forming factor in the cyanobacterium Spirulina platensis. Plant Cell Physiol. 1992;33:21–25. [Google Scholar]
- Passner JM, Schultz SC, Steitz TA. Modeling the cAMP-induced allosteric transition using the crystal structure of CAP-cAMP at 2.1 A resolution. J Mol Biol. 2000;304:847–859. doi: 10.1006/jmbi.2000.4231. [DOI] [PubMed] [Google Scholar]
- Pogorelova AY, Loiko NG, Van’kova AA. Effect of alkoxybenzene on formation of dormant cyst-like cells of Sinorhizobium meliloti P221. Izvestiya Timiryazevskoi Sel’skokhozyaistvennoi Akademii. 2009;1:149–154. [Google Scholar]
- Quilliam LA, Khosravi-Far R, Huff SY, Der CJ. Guanine nucleotide exchange factors: activators of the Ras superfamily of proteins. Bioessays. 1995;17:395–404. doi: 10.1002/bies.950170507. [DOI] [PubMed] [Google Scholar]
- Ragatz L, Jiang ZY, Bauer CE, Gest H. Macroscopic phototactic behavior of the purple photosynthetic bacterium Rhodospirillum centenum. Arch Microbiol. 1995;163:1–6. doi: 10.1007/BF00262196. [DOI] [PubMed] [Google Scholar]
- Rauch A, Leipelt M, Russwurm M, Steegborn C. Crystal structure of the guanylyl cyclase Cya2. Proc Natl Acad Sci U S A. 2008;105:15720–15725. doi: 10.1073/pnas.0808473105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roberts DP, McKenna LF, Hu X, Lohrke SM, Kong HS, de Souza JT, Baker CJ, Lydon J. Mutation in cyaA in Enterobacter cloacae decreases cucumber root colonization. Arch Microbiol. 2007;187:101–115. doi: 10.1007/s00203-006-0177-6. [DOI] [PubMed] [Google Scholar]
- Sadasivan L, Neyra CA. Cyst production and brown pigment formation in aging cultures of Azospirillum brasilense ATCC 29145. J Bacteriol. 1987;169:1670–1677. doi: 10.1128/jb.169.4.1670-1677.1987. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sakamoto T, Murata N, Ohmori M. The Concentration of cyclic AMP and adenylate cyclase activity in cyanobacteria. Plant Cell Physiol. 1991;32:581–584. [Google Scholar]
- Schaap P. Guanylyl cyclases across the tree of life. Front Biosci. 2005;10:1485–1498. doi: 10.2741/1633. [DOI] [PubMed] [Google Scholar]
- Setlow B, Setlow P. Levels of cyclic GMP in dormant, germinated, and outgrowing spores and growing and sporulating cells of Bacillus megaterium. J Bacteriol. 1978;136:433–436. doi: 10.1128/jb.136.1.433-436.1978. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shenroy AR, Visweswariah SS. Class III nucleotide cyclases in bacteria and archaebacteria: lineage-specific expansion of adenylyl cyclases and a dearth of guanylyl cyclases. FEBS Lett. 2004;561:11–21. doi: 10.1016/s0014-5793(04)00128-0. [DOI] [PubMed] [Google Scholar]
- Shibuya M, Takebe Y, Kaziro Y. Possible Involvement of cya Gene in Synthesis of Cyclic Guanosine 3′,5′-Monophosphate in Escherichia coli. Cell. 1977;12:521–528. doi: 10.1016/0092-8674(77)90128-3. [DOI] [PubMed] [Google Scholar]
- Sismeiro O, Trotot P, Biville F, Vivares C, Danchin A. Aeromonas hydrophila adenylyl cyclase 2: a new class of adenylyl cyclases with thermophilic properties and sequence similarities to proteins from hyperthermophilic archaebacteria. J Bacteriol. 1998;180:3339–3344. doi: 10.1128/jb.180.13.3339-3344.1998. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Son JK, Rosazza JP. Cyclic guanosine-3′,5′-monophosphate and biopteridine biosynthesis in Nocardia sp. J Bacteriol. 2000;182:3644–3648. doi: 10.1128/jb.182.13.3644-3648.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Statwald-Demchick R, Turner F, Gest H. Physiological properties of the thermotolerant photosynthetic bacterium, Rhodospirillum centenum. FEMS Micro Lett. 1990;67:139–144. [Google Scholar]
- Stevenson LH, Socolofsky MD. Cyst formation and poly-beta-hydroxybutyric acid accumulation in Azotobacter. J Bacteriol. 1966;91:304–310. doi: 10.1128/jb.91.1.304-310.1966. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stoffels M, Castellanos T, Hartmann A. Design and application of new 16S rRNA-targeted oligonucleotide probes for the Azospirillum-Skermanella-Rhodocista-cluster. Syst Appl Microbiol. 2001;24:83–97. doi: 10.1078/0723-2020-00011. [DOI] [PubMed] [Google Scholar]
- Sun IC, Shapiro L, Rosen OM. Purification and characterization of guanylate cyclase from Caulobacter crescentus. Biochem Biophys Res Comm. 1974;61:193–203. doi: 10.1016/0006-291x(74)90552-x. [DOI] [PubMed] [Google Scholar]
- Susstrunk U, Pidoux J, Taubert S, Ullmann A, Thompson CJ. Pleiotropic effects of cAMP on germination, antibiotic biosynthesis and morphological development in Streptomyces coelicolor. Mol Microbiol. 1998;30:33–46. doi: 10.1046/j.1365-2958.1998.01033.x. [DOI] [PubMed] [Google Scholar]
- Tellez-Sosa J, Soberon N, Vega-Segura A, Torres-Marquez ME, Cevallos MA. The Rhizobium etli cyaC product: characterization of a novel adenylate cyclase class. J Bacteriol. 2002;184:3560–3568. doi: 10.1128/JB.184.13.3560-3568.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Terauchi K, Ohmori M. An adenylate cyclase, Cya1, regulates cell motility in the cyanobacterium Synechocystis sp. PCC 6803. Plant Cell Physiol. 1999;40:248–251. doi: 10.1093/oxfordjournals.pcp.a029534. [DOI] [PubMed] [Google Scholar]
- Weber IT, Steitz TA. Structure of a complex of catabolite gene activator protein and cyclic AMP refined at 2.5 A resolution. J Mol Biol. 1987;198:311–326. doi: 10.1016/0022-2836(87)90315-9. [DOI] [PubMed] [Google Scholar]
- Wu J, Bai J, Bao Q, Zhao F. Lineage-specific domain fusion in the evolution of purine nucleotide cyclases in cyanobacteria. J Mol Evol. 2008;67:85–94. doi: 10.1007/s00239-008-9127-z. [DOI] [PubMed] [Google Scholar]
- Youn H, Kerby RL, Koh J, Roberts GP. A C-helix residue, Arg-123, has important roles in both the active and inactive forms of the cAMP receptor protein. J Biol Chem. 2007;282:3632–3639. doi: 10.1074/jbc.M606602200. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.