

Abstract
The cellular requirements for Dicer, an essential miRNA processing enzyme, and the consequences of altered levels of its expression on tumorigenesis are incompletely understood. We review the effects of Dicer loss in cells of different origin and whether loss of p53 permits cell survival and growth in the absence of Dicer.
Keywords: Dicer, lymphoma, p53, tumorigenesis
microRNAs (miRNAs) are small non-coding RNAs that post-transcriptionally regulate gene expression. Several miRNAs are linked to biological processes associated with cancer initiation and progression.1 In tumorigenesis, the requirements for miRNAs and the enzymes that process them remain unresolved. While tumor development may be facilitated by the gain or loss of specific miRNAs, an overall reduction in miRNA has been reported in human malignancies.1 Reduced activity or expression of Dicer, an enzyme crucial for miRNA biogenesis, causes downregulation of miRNA in some cancers.1 These observations led to the hypothesis that reduced Dicer levels facilitate tumorigenesis.
In support of this concept, Dicer was reported to function as a haploinsufficient tumor suppressor in mouse models of soft-tissue sarcoma, lung adenocarcinoma, and retinoblastoma.2-4 However, Dicer hypomorphic mice, which express Dicer at 20% of normal levels, do not have an increased incidence of cancer.5 Moreover, we demonstrated that Dicer does not function as a haploinsufficient tumor suppressor in B-cell lymphoma, as loss of one Dicer allele did not affect the rate of B-cell lymphomagenesis regardless of the p53 tumor suppressor status. Additionally, heterozygosity of Dicer did not alter Dicer protein levels or mature miRNA production in B-cell lymphomas, indicating that one Dicer allele is sufficient for normal Dicer function in B cells.6,7 Therefore, reduced levels of Dicer appear to have differential effects in different tissues during transformation.
There is mounting evidence that Dicer, and thus miRNA, is required for tumorigenesis. Although it was initially reported that deletion of Dicer in the lung led to tumor development, it was later determined the lung cancer cells in these mice actually retained one Dicer allele.2 Additionally, attempts at homozygous Dicer deletion in other mouse models, including data generated in our laboratory with a Myc oncogene-driven B-cell lymphoma model, have revealed selection against complete ablation of Dicer in cancers.2,3,6,7 We showed that Myc-induced lymphomas in Cre-expressing B cells were delayed in Dicer conditional knockout mice as a result of B-cell apoptosis, and none of the lymphomas that did develop had deleted both Dicer alleles. Moreover, biallelic deletion of Dicer in established lymphomas resulted in apoptosis. Screening of 1,303 lymphoma clones that survived Dicer deletion from two studies revealed that all had retained at least one Dicer allele.6,7 In addition, although homozygous deletion of Dicer in sarcoma cells was possible, the cells had impaired proliferation.4 Thus, a certain level of Dicer expression appears to be necessary for tumor cell development, survival, and growth.
Inactivation of the p53 pathway is a prerequisite to cancer as it allows cells to evade apoptotic and growth arrest signals. Loss of miRNA biogenesis can lead to cellular stress and activation of p53 (Fig. 1). Ablation of Dicer in untransformed murine fibroblasts or epidermal cells led to p53 activation and premature senescence or apoptosis, respectively.8,9 Deleting p53 or Arf (a tumor suppressor and regulator of p53) delayed the premature senescence in fibroblasts,9 and loss of p53 inhibited the apoptosis in epidermal cells8 (Fig. 1). Additionally, a small fraction of p53-null, mutant K-Ras–expressing sarcoma cells and SV40-immortalized (p53 and Rb inactivated) mesenchymal stem cells can survive deletion of Dicer4 (Fig. 1). We detected an elevated frequency of p53 inactivation in B-cell lymphomas upon Dicer deletion, indicating an increase in p53 activation during transformation.7 These data suggest that p53 inactivation may extend cell survival or be required for survival when Dicer is deleted. However, a p53 deficiency did not rescue the B-cell apoptosis caused by Dicer deletion (Fig. 1), and did not prevent the resultant delay in lymphomagenesis. p53 deletion was also insufficient to allow established B-cell lymphomas to survive Dicer deletion in vitro or in vivo.6 Arf deletion also did not confer a survival advantage to B-cell lymphomas with deletion of Dicer. Furthermore, B-cell lymphomas frequently overexpress the Bcl-2 anti-apoptotic protein, which may partially protect developing B cells, but not lymphomas, from apoptosis induced by deletion of Dicer.6,10 Additionally, synthetic lethality occurred when p53 was inactivated in Dicer- and Rb-deficient retinal cells.1 We observed that p53 deficiency restored development of the characteristic B-cell lymphoma in our Dicer conditional knockout mouse model in comparison to the early precursor B-cell lymphoma that arose in 40% of the mice born with both p53 alleles.6,7 Together, the data reveal tissue-specific requirements for Dicer and that p53 pathway inactivation is unlikely to rescue, or may only delay, the effects of Dicer deletion. Moreover, in contrast to non-hematopoietic cells, miRNAs appear to have an irreplaceable function in B-cell survival, regardless of cellular transformation or p53 status.
Figure 1.
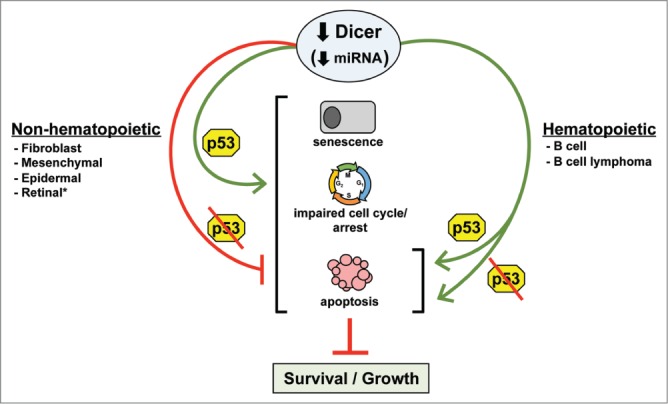
p53 inactivation protects some cells from the negative consequences of deletion of Dicer. The p53 tumor suppressor responds to impaired miRNA processing by inducing senescence, cell cycle arrest, or apoptosis. Non-hematopoietic cells that harbor inactivated p53 can survive and grow (albeit possibly more slowly) in the absence of Dicer. In contrast, hematopoietic cells are extremely sensitive to Dicer loss and rapidly undergo apoptosis, regardless of p53 status. *Co-inactivation of the Rb pathway with p53 results in synthetic lethality.
Hematopoietic cells preferentially undergo apoptosis, whereas non-hematopoietic cells favor senescence, when stressful situations such as impaired miRNA processing are encountered (Fig. 1). We demonstrated that inactivation of the p53 pathway neither protected against nor cooperated with Dicer loss during B-cell lymphomagenesis.6,7 Although it remains to be determined whether specific genetic alterations would permit B-cell lymphomas to survive without Dicer, our data establish Dicer as a possible therapeutic target for the treatment of B-cell lymphomas, and likely other hematopoietic malignancies. A concern with moving forward on such a strategy is that Dicer appears to function as a haploinsufficient tumor suppressor for lung, muscle, and retina, but this has only been shown in the context of p53 inactivation and after prolonged Dicer deficiency.2-4 B-cell lymphomas are profoundly sensitive to loss of Dicer and rapidly undergo apoptosis,6,7 suggesting that short-term inactivation of Dicer would result in the death of B-cell lymphoma cells. Moreover, standard DNA damage-inducing chemotherapeutics can result in cancer development, whereas transient inactivation of Dicer is unlikely to be tumorigenic.
Disclosure of Potential Conflict of Interest
No potential conflicts of interest were disclosed.
Funding
Our studies on Dicer were supported by F31CA165728 (CMA) and R01CA148950 (CME).
References
- 1. Jansson MD, Lund AH. MicroRNA and cancer. Mol Oncol 2012; 6:590-610; PMID:23102669; http://dx.doi.org/ 10.1016/j.molonc.2012.09.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2. Kumar MS, Pester RE, Chen CY, Lane K, Chin C, Lu J, Kirsch DG, Golub TR, Jacks T. Dicer1 functions as a haploinsufficient tumor suppressor. Genes Dev 2009; 23:2700-4; PMID:19903759; http://dx.doi.org/ 10.1101/gad.1848209 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3. Lambertz I, Nittner D, Mestdagh P, Denecker G, Vandesompele J, Dyer MA, Marine JC. Monoallelic but not biallelic loss of Dicer1 promotes tumorigenesis in vivo. Cell death Differ 2010; 17:633-41; PMID:20019750; http://dx.doi.org/ 10.1038/cdd.2009.202 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Ravi A, Gurtan AM, Kumar MS, Bhutkar A, Chin C, Lu V, Lees JA, Jacks T, Sharp PA. Proliferation and tumorigenesis of a murine sarcoma cell line in the absence of DICER1. Cancer Cell 2012; 21:848-55; PMID:22698408; http://dx.doi.org/ 10.1016/j.ccr.2012.04.037 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5. Morita S, Hara A, Kojima I, Horii T, Kimura M, Kitamura T, Ochiya T, Nakanishi K, Matoba R, Matsubara K, et al. . Dicer is required for maintaining adult pancreas. PloS One 2009; 4:e4212; PMID:19148298; http://dx.doi.org/ 10.1371/journal.pone.0004212 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6. Adams CM, Eischen CM. Inactivation of p53 is insufficient to allow B cells and B-cell lymphomas to survive without dicer. Cancer Res 2014; 74:3923-34; PMID:24840646; http://dx.doi.org/ 10.1158/0008-5472.CAN-13-1866 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7. Arrate MP, Vincent T, Odvody J, Kar R, Jones SN, Eischen CM. MicroRNA biogenesis is required for Myc-induced B-cell lymphoma development and survival. Cancer Res 2010; 70:6083-92; PMID:20587524; http://dx.doi.org/ 10.1158/0008-5472.CAN-09-4736 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Lyle S, Hoover K, Colpan C, Zhu Z, Matijasevic Z, Jones SN. Dicer cooperates with p53 to suppress DNA damage and skin carcinogenesis in Mice. PloS One 2014; 9:e100920; PMID:24979267; http://dx.doi.org/ 10.1371/journal.pone.0100920 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. Mudhasani R, Zhu Z, Hutvagner G, Eischen CM, Lyle S, Hall LL, Lawrence JB, Imbalzano AN, Jones SN. Loss of miRNA biogenesis induces p19Arf-p53 signaling and senescence in primary cells. J Cell Biol 2008; 181:1055-63; PMID:18591425; http://dx.doi.org/ 10.1083/jcb.200802105 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Koralov SB, Muljo SA, Galler GR, Krek A, Chakraborty T, Kanellopoulou C, Jensen K, Cobb BS, Merkenschlager M, Rajewsky N, et al. . Dicer ablation affects antibody diversity and cell survival in the B lymphocyte lineage. Cell 2008; 132:860-74; PMID:18329371; http://dx.doi.org/ 10.1016/j.cell.2008.02.020 [DOI] [PubMed] [Google Scholar]