

Abstract
Electrical stimulation and exercise are treatments to enhance recovery from peripheral nerve injuries. Brain-derived neurotrophic factor and androgen receptor signaling are requirements for the effectiveness of these treatments. Increased neuronal activity is adequate to promote regeneration in injured nerves, but the dosing of activity and its relationship to neurotrophins and sex steroid hormones is less clear. Translation of these therapies will require principles associated with their cellular mechanisms.
Following traumatic injury to nerves in the peripheral nervous system (PNS), full functional recovery is poor, despite the well documented ability of axons to regenerate and reinnervate peripheral targets. This poor recovery is an important public health concern. Among the 200,000+ victims of new peripheral nerve injuries in the U.S. each year (72), only ∼10% ever recover full function (65). The slowness and inefficiency of axon regeneration is most often blamed for this poor outcome. Regenerating axons from the proximal segment of a cut peripheral nerve must enter a pathway in the distal segment of the nerve and then elongate in that pathway to reach their targets (11). Elongation in the pathway is slow so that if the distance needed to be traversed is long, as they sometimes can be in human patients, a prolonged recovery time will be expected. Elongation of the regenerating axons requires the presence of growth-promoting molecules in the pathway, and over time the ability of supporting cells in the pathway diminishes, which also contributes to poor recovery (34). Enhancing axon regeneration has emerged as a therapeutic target for improving functional recovery after PNS injuries.
Based on earlier hypotheses that neuronal activity might contribute to the process of axon survival and regeneration (e.g., Ref. 40), Gordon and colleagues undertook a systematic evaluation of the effect of electrical stimulation (ES) of cut peripheral nerves on subsequent axon regeneration (3). They discovered that both motor (3) and sensory (29) axon regeneration were enhanced by as little as 1 h of continuous (20 Hz) supramaximal stimulation of the proximal stump of a cut nerve. Blocking propagation of the evoked action potentials from reaching the cell bodies of these neurons by injecting the sodium channel blocker tetrodotoxin into the nerve proximal to the stimulation site resulted in a complete loss of the enhancement induced by ES (3), indicating the activity dependence of the treatment. A number of laboratories have now used ES to enhance axon regeneration in different models of peripheral nerve injury (reviewed in Ref. 76).
On the basis of these results, we showed that moderate daily exercise for 2 wk also enhanced axon regeneration, perhaps exceeding the effects of ES (FIGURE 1) (62). Others have used slightly different exercise protocols to achieve the same end (7, 51, 54, 80, 81), also including swimming (38, 39, 44, 74), and rhythmic limb movements in anesthetized animals (77). Navarro and colleagues even studied the effects of 1 h of ES followed by daily exercise and found greater enhancement of regeneration than was found with using either treatment alone (7).
FIGURE 1.
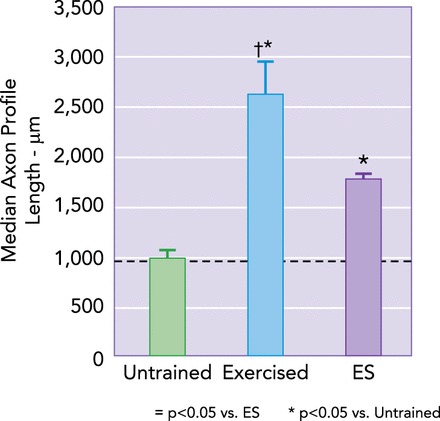
Electrical stimulation and exercise promote axon regeneration in cut peripheral nerves
The common fibular nerves of mice were cut and repaired by end-to-end anastomosis. Lengths of profiles of regenerating axons were measured 2 wk later. Average median axon profile lengths (±SE) are shown for untrained mice, mice that were exercised for 1 h daily during the recovery period, and mice that were exposed to a single hour of electrical stimulation (ES) at the time of nerve repair. Note that both of these therapies result in enhancement of axon regeneration, but exercise is ∼50% more potent. Data from Refs. 24, 62.
A common feature to all of these approaches has been assumed to be an increase in the activity of the neurons whose axons are regenerating (76). Increased activity in both sensory neurons and motoneurons is well established with ES. Increased activity of axotomized motoneurons is anticipated during therapies involving walking or swimming, and these activities are driven by the outputs of spinal circuits. Indeed, rhythmic activity in cut ventral roots, presumed to reflect the outputs of the central pattern generators for locomotion, has been described in reduced preparations (e.g., Ref. 52). Activity during treadmill training of motoneurons whose axons have been cut and are regenerating has not been studied specifically. However, it seems unlikely that the outputs of spinal circuitry controlling those behaviors will be suppressed by peripheral nerve transection, especially because the forelimbs and higher centers that drive lumbar central pattern generation (37, 66) are left intact and animals are trained quadrupedally. Similarly, activity of sensory neurons whose axons have been cut and are regenerating has not been studied. However, it has been proposed that the central pattern generator for locomotion controls the efficacy of transmission of some primary afferent pathways via primary afferent depolarization (53), but whether these effects might promote increased activity of axotomized sensory neurons during walking/swimming is not known.
The details of the approaches to enhancing axon regeneration using the different experimental therapies described above vary considerably, which complicates any potential translation of these findings to treatments of the wide variety of different peripheral nerve injuries encountered clinically. Approaches involving exercise have the advantage of being low tech, and thus low cost, and they enable patients to participate in and take responsibility for their own recovery. Unfortunately, the large number of variables associated with even the simplest exercise protocol makes the application of exercise to any patient a daunting one. Indeed, even within the experimental studies cited above, differences in the intensity of exercise, the duration of exercise, the pattern of training, the frequency of training sessions, and the time after injury when the exercise was begun are profound.
A number of recent studies have begun to investigate the cellular mechanisms underlying the effectiveness of these activity-associated therapies. In an effort to begin to establish principles for the application of exercise to enhance axon regeneration after injuries to the PNS, in this review we will consider three aspects of the cellular basis for the efficacy of exercise as a therapy that might advance its translational potential: neurotrophins, sex steroid hormones, and neural activity.
Neurotrophins
The effectiveness of ES and other activity-associated experimental therapies for treating peripheral nerve injury, such as exercise, requires the ability of the regenerating axons to produce brain-derived neurotrophic factor (BDNF). This neurotrophin has been considered an important promoter of axon elongation during regeneration of peripheral nerves for some time, but in a slightly different cellular context than that invoked to explain the effectiveness of therapies such as ES or exercise. Transformed Schwann cells in the distal segment of cut nerves are known to express BDNF (27, 36), which then acts as a retrograde signal, via trkB receptors on regenerating axons, to stimulate axon regeneration. Treatments of cut and repaired nerves with recombinant BDNF (24, 48) or small molecule trkB agonists (23) promote axon regeneration. Blocking the effect of BDNF with function-blocking antibodies (86) or by using mouse genetics to knock out the BDNF gene selectively in Schwann cells (83) markedly reduces axon outgrowth. Expression of BDNF and its receptor trkB increases transiently in motor and sensory neurons during the first 2–3 days following peripheral nerve transection (33). One hour of ES produces a rapid and marked increase in expression of BDNF and trkB in motoneurons that lasts for at least a week (2). Increased expression of BDNF mRNA in motoneurons is well established following voluntary exercise (32) or treadmill training (59, 83). In mice null for BDNF or trkB specifically in neurons, the effect of exercise is lost completely (23, 83) (FIGURE 2). Expression of the BDNF gene is known to be driven by neuronal activity (41) but also by the downstream effects of activity-stimulated increases in expression of the transcription factor Sox11 (55, 64, 70).
FIGURE 2.
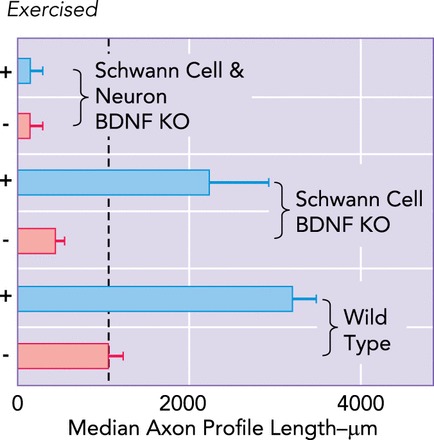
Exercise-based enhancement of axon regeneration requires neuronal BDNF
Common fibular nerves of wild-type and cell-type-specific BDNF knockout mice were cut and repaired. Some were exercised daily for 2 wk and others were not. Average (±SE) median lengths of profiles of regenerating axons are shown. Values for all groups differ significantly (P < 0.05) from the value found in untrained wild-type mice (bottom bar, vertical dashed line). Knocking out BDNF in Schwann cells surrounding regenerating axons inhibits axon regeneration, but this can be reversed by exercise. Knocking out BDNF in both Schwann cells and the regenerating axons also reduces axon regeneration, but it cannot be reversed by exercise. Data are from Ref. 83.
These results all are compatible with the hypothesis first put forth by Gordon and coworkers (4) that therapies such as ES or exercise promote axon regeneration in the PNS by an autocrine or paracrine BDNF signaling mechanism. Secretion of BDNF by growth cones of regenerating axons binds to trkB receptors on the same or adjacent axons to promote growth. Any paracrine stimulation of axon elongation would require BDNF to be secreted from one axon and cross the barriers of at least two endoneurial tubes to stimulate the growth of adjacent axons. Because in mice in which the gene for BDNF is selectively knocked out in only a subset of peripheral axons we found no promotion of regeneration of BDNF-deficient axons from adjacent BDNF-expressing axons (83), we favor an autocrine signaling pathway. This mechanism compliments the traditional role of BDNF as a retrograde signal to promote regeneration.
Sensitive assays have now been developed to measure BDNF in serum, and they have been used to show that BDNF protein levels increase in serum with exercise protocols similar to those we have shown to result in enhanced axon regeneration (61). Whether these assays could be applied clinically to screen for the effectiveness of different types of exercise as a therapy for axon regeneration in the PNS is problematic, since the role of BDNF may be so localized to the regenerating axons that its effect might not be detected in serum. However, it would be of considerable interest to know whether measures of serum BDNF concentration would form a reasonable predictor of the success of any exercise protocol in promoting axon regeneration in cut nerves.
An additional impediment to the translation of these findings might be the presence of single nucleotide polymorphisms (SNPs) in the BDNF gene. Several SNPs of this gene have been described, but the best known of these (rs6265) results in a single amino acid (Val66Met) substitution and is found in >25% of the human population (6). The Val66Met substitution is in the prodomain of the BDNF molecule (6), which is cleaved to form mature BDNF either intracellularly, in the trans-Golgi network or secretory vesicles (73), or by plasmin or matrix metalloproteinases after secretion (49, 85). Regulated release of BDNF is impaired in cells containing the Val66Met SNP (12, 14, 22), and in a mouse model of this SNP, the mutant prodomain stimulated growth cone retraction via the common neurotrophin receptor p75NTR (6). Thus individuals with the Val66Met SNP might not be able to respond appropriately to therapies for peripheral nerve injury such as ES or exercise, and their application might even result in an inhibition of axon regeneration. A mouse model of the Val66Met SNP has been developed (13), and its phenotype is remarkably similar to that observed in individuals with this SNP. Evaluating the effectiveness of these experimental therapies for treatment of peripheral nerve injuries in this mouse model should provide important insights as to their efficacy in the human population.
Sex Steroid Hormones
In the course of evaluating the effect of exercise on axon regeneration in mice, we discovered a marked sex difference. In male mice, 1 h of slow treadmill walking daily (continuous training protocol) results in a marked increase in the length of regenerating axons 2 wk later, but the same exercise protocol has no enhancing effect in females. In female mice exposed to daily interval training at a faster treadmill speed, a pattern of activity that is comparable to that observed during voluntary wheel running (20), the enhancement of axon regeneration is impressive, but no enhancement is found in male mice exposed to the same interval training protocol (84).
Sex steroid hormones likely mediate this sex difference. Levels of serum testosterone are significantly increased in continuously exercised male mice but not in continuously exercised females or interval-exercised mice of either sex. Castrating males blocked the enhancing effect of exercise completely and did not influence the ineffectiveness of the interval-training protocol, suggesting a role for gonadally derived androgens. Although no increase in serum testosterone was found in interval-trained females, treatments of unexercised female mice with an inhibitor of P450 aromatase, an enzyme that catalyzes the conversion of testosterone and its precursors into estradiol, resulted in a striking enhancement of the lengths of regenerating axons (84). In subsequent experiments, we found that treating mice with the androgen receptor blocker flutamide blocked the effect of both exercise and ES in both males and females (75) (FIGURE 3).
FIGURE 3.
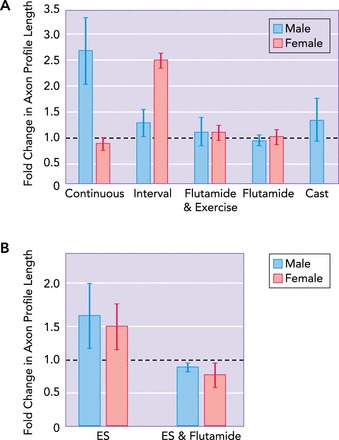
Androgens are required for enhancement of regeneration by exercise or ES
A: average (+95% confidence limits) fold changes in median lengths of profiles of regenerating axons 2 wk after the common fibular nerve was cut and repaired are shown for male and female mice. Animals were exercised using either slow continuous walking for 1 h or a more intense interval training strategy. Some animals were treated with a sustained release dosage form of the androgen receptor blocker flutamide, with either sex-appropriate exercise or no exercise. Animals in one group of male mice were castrated at the time of nerve repair. Data from Refs. 72, 81. B: similar flutamide treatment blocked the enhancing effect of a single application of ES in both male and female mice. Data are from Ref. 75.
Based on these findings, we proposed that the effects of therapies such as ES or exercise in promoting axon regeneration in the PNS had an androgenic component in both males and females. Although androgen receptors are found in all motoneurons (26), the identity of cells in which this critical androgen receptor signaling takes place and the relationship between this signaling and ES or exercise on the one hand and increased BDNF/trkB expression on the other are not clear at this time. It is likely there is also an estrogenic component to the enhancement. Estrogen receptors have been shown to increase expression in axotomized lumbar motoneurons (46), and exogenous estrogen administration has been shown to enhance axon regeneration (45, 67, 71) and prevent axotomy-induced cell death (43) following nerve crush. The source of these endogenous sex hormones is not clear; the hormones could be gonadally derived or locally synthesized by neurons or glia. Although the expression of neuronal androgen and estrogen receptors and the effects of gonadal hormones after nervous injuries have been studied extensively, only recently has de novo neuronal steroid synthesis in injury and disease begun to be investigated. Application of findings regarding the role of steroid hormones may be difficult since it is reasonable to assume that neuronal sex steroids will have differential roles intrinsic to the state of the nervous system (developing, aging, injured, sedentary, exercising, male, female, etc.). For example, the androgen receptor blocker flutamide has no effect on normal hippocampal cell proliferation but blocks exercise-induced increases in neurogenesis (60).
Glia in the central nervous system respond to peripheral axotomy and are also known to be responsive to androgens (8, 15, 16, 19, 42) and estrogens (1, 69). Astrocytes express estrogen and androgen receptors (42), and microglia express androgen receptors (28). After brain lesion, reactive astrocytes and microglia may be targets of androgens and estrogens (28). These reactive astroctyes and microglia can also secrete BDNF (57, 68). In addition, microglia express steroid-converting enzymes (35) such as 5 alpha-reductase, which converts testosterone into dihydrotestosterone. The presence of these enzymes suggest that glia may be able to locally produce androgens and could stimulate the release of BDNF in an autocrine and/or paracrine manner. Although microglial-secreted BDNF has been implicated in the maintenance of neuropathic pain after peripheral nerve injury (17, 78) and astrocyte-secreted BDNF increases dramatically after spinal cord injury (21), whether these glia assist or hinder neural regeneration after injury is still not clear.
Expression of BDNF and trkB is stimulated by estrogens (9) and androgens (67, 82) in neurons as well. This hormone-dependent expression appears to have a slower onset but a longer time course of expression than found after ES (67). If exercise is enhancing axon regeneration by both promoting neuronal activity similar to ES and stimulating the release of sex steroids, then exercise may result in both a rapid onset and a prolonged duration of increased expression of BDNF and trkB in axotomized neurons and/or surrounding reactive glia, but this has not been studied. Determining the time dependence of these mechanisms will be important to create appropriate exercise-based therapies for patients.
Although it is clear that neuronal BDNF as well as androgen receptor signaling is necessary for axon regeneration, the details of how exercise increases androgens is less clear. Axotomized neurons likely synthesize androgens de novo or are supplied androgens or androgenic precursors by neighboring glia. We hypothesize that exercise- or ES-induced neuronal activity increases local neuronal androgen levels by stimulating expression of the enzymes (17β-hydroxysteriod dehydrogenase and 5α-reductase) that catalyze the conversion of androgenic precursors into testosterone and dihydrotestosterone. We also hypothesize that neuronal activity can modulate the expression of the enzyme P450 aromatase, which converts testosterone or its precursors into estradiol.
The source of estrogen as well as the specific mechanism by which estrogen signaling may be altered by exercise also is not clear. Estrogen and exercise interact to promote the expression of BDNF. Berchtold and colleagues (9) found that ovariectomizing female rats immediately reduced voluntary exercise levels and dramatically reduced BDNF mRNA levels in the hippocampus. Restoring estrogen to intact control levels immediately restored voluntary exercise levels and slowly returned BDNF mRNA expression to control levels (9). This interaction of estrogen and BDNF could be due to genomic regulation of the BDNF gene via the activation of the estrogen response element on the BDNF gene or various nongenomic mechanisms (56).
Discovering how expression of these two pathways is affected by injury to the PNS and neuronal activity may reveal new pharmacological targets and will aid in the translation of experimental activity-associated therapies. The two hormone pathways may be acting independently or synergistically to promote axon regeneration (67, 71). Because androgen (and likely estrogen) receptor signaling is required in both males and females for the effectiveness of therapies such as ES or exercise, their application in patients with steroid hormone deficiency, a history of prostate cancer, or postmenopausal women should be investigated more thoroughly.
Neuronal Activity
The consensus reached in the earliest studies of the effects of exercise in enhancing recovery from spinal cord injury was that the effects of exercise were the result of increased neuronal activity. In more recent papers, other explanations have been put forth, including environmental enrichment (31), increased caloric utilization (30), and intermittent hypoxia (18). Using mice in which the light-sensitive cation channel channelrhodopsin (ChR2) is expressed in some but not all axons in peripheral nerves, we have been able to show that increasing neuronal activity using light results in a selective enhancement of regeneration of motor axons expressing ChR2 and not axons that do not contain the transgene (FIGURE 4) (47). These results are consistent with the hypothesis that increased neuronal activity alone is sufficient to enhance axon regeneration. However, a number of important questions remain unanswered. The nature of the increase in neural activity, the minimal amount of increase that is sufficient, and whether sensory and motor axons have the same requirements for increased activity all are not now known but are amenable to future study using this optogenetics approach. Indeed, although it is clear that exercise leads to an enhancement of regeneration of motor axons (see below), its effect on the regeneration of sensory axons is relatively unknown. The use of retrograde labeling to study sensory axon regeneration after different forms of exercise should be a future priority.
FIGURE 4.
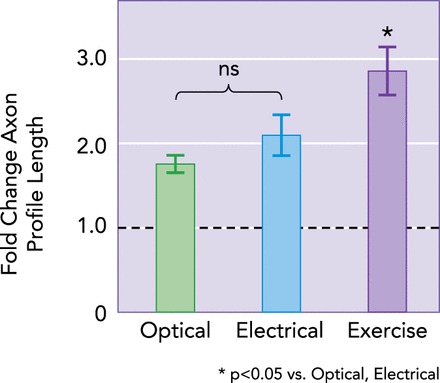
Increased neuronal activity may be sufficient to promote axon regeneration
In male thy-1-ChR2/YFP mice (optical; n = 4), cut axons were activated by blue light for 1 h before nerve repair. In control mice, nerves were cut and repaired with no treatment (untreated; n = 8), 1 h of ES (n = 5), or 2 wk of daily treadmill exercise (n = 7). Means ± SE fold changes in lengths of regenerating axons relative to untreated mice are shown. The horizontal dashed line at 1.0 reflects the amount of regeneration observed in untreated animals. Mean fold changes for all three groups shown are significantly greater than for untrained mice. Data are from Refs. 47, 62.
In addition to any requirements for increased activity, establishing the appropriate timing of that increase will be important. In rats that were treadmill trained daily for 2 wk beginning immediately (3 days) after sciatic nerve transection and repair, we studied the restoration of the compound muscle action potential (M response) recorded from reinnervated muscles and that of the H reflex, the muscle potential evoked in response to sensory axon stimulation (10). These results were compared with a series of rats in which the same 2-wk exercise protocol was applied but delayed until the first sign of muscle reinnervation, ∼3 wk after injury, and to a group of untrained controls. By 8–10 wk after injury, the amplitudes of the restored M responses in both the immediately trained rats and those whose training was delayed were significantly greater than those found in untrained controls (FIGURE 5A). Using a slightly different immediate training protocol, Navarro and colleagues (7) also found an enhancement of motor axon regeneration leading to a larger M response. In contrast, the amplitude of the restored H reflexes differed in the different groups. In untreated control rats, a transient increase in the amplitude of the restored H reflex is followed by a significant decrease, relative to that found in intact rats (10, 58). In immediately trained rats, the exaggerated reflex response is retained for at least 8–10 wk (FIGURE 5B) (10). If the application of exercise is delayed, the size of the restored H reflex at this recovery time is the same as that found in untreated rats (FIGURE 5B). Whether these different effects of the timing of initiation of exercise on the plasticity of spinal circuits following injury to the PNS are the result of differences in the enhancing effects of exercise on sensory vs. motor axon regeneration or whether they are influenced by the rearrangements of synaptic inputs onto motoneurons that accompany peripheral axotomy (e.g., Ref. 5) is not clear at this time. A marked difference in the outcome of the effect of exercise on synaptic plasticity may depend on when that treatment is applied. More studies are needed to define the therapeutic window for application of exercise.
FIGURE 5.
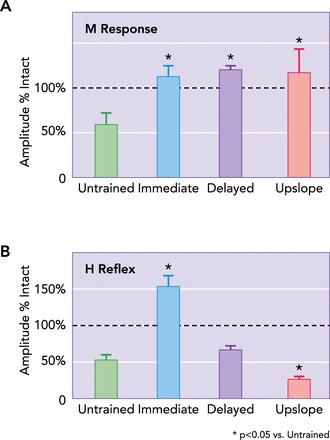
Outcomes of exercise on axon regeneration and functional recovery may differ for different kinds of axons and the treatment applied
A: in rats treated with 2 wk of daily moderate treadmill training, the amplitude of the direct muscle (M) response to stimulation of regenerated axons is increased markedly 8–10 wk after injury, whether the exercise was begun immediately (3 days after transection), delayed for 3 wk, or conducted using an upslope inclined treadmill. Mean amplitudes (±SE; n = 6 for each group) of M responses, scaled to the pre-transection M response amplitude (horizontal dashed line) are shown. B: similar data for the amplitude of the H reflex in these three groups. In animals of the immediate group, H-reflex amplitude is increased by exercise. In animals in the upslope group, the training actually decreases the amplitude of the H reflex. Some data are from Ref. 62, 63.
We also have investigated the effects of changing the intensity or amplitude of the applied exercise. Rats and mice were exercised for 1 h daily, 5 days/wk, for 2 wk by walking slowly on an upward-inclined treadmill. Training was begun on day 3 after sciatic nerve transection and repair. This immediate upslope training was expected to result in the recruitment of more motor units into activity than level training, since the amplitude of EMG activity in both flexor and extensor muscles is increased with slope (63). Since during upslope walking muscles undergo very little lengthening (50), we expected upslope training to result in decreased activity in sensory axons encoding stretch of intact synergist muscles that feed back onto the axotomized motoneurons.
In mice, we found that upslope training resulted in the successful regeneration of significantly more motor axons during the first 2 post-transection weeks than either untrained or level-trained animals (25). More sciatic motoneurons could be retrogradely labeled from tracer application 4 mm distal to the injury site in upslope-trained mice (240.98 ± 9.64, mean ± SE) than in untrained animals (38.20 ± 6.76), level-trained animals (151.09 ± 21.51), or animals treated with 1 h of ES (109.33 ± 27.22). Recruiting more motoneurons into activity during exercise produced the anticipated enhancement of motor axon regeneration.
In rats, we studied the amplitudes of the restored M responses and H reflexes in groups of animals exposed to 2 wk of upslope training, level training, or no training. The amplitudes of the restored M responses 8–10 wk after injury was significantly greater in both level- and upslope-trained rats than in untrained controls (FIGURE 5A), but no significant differences were found between level- and upslope-training groups. Whether the mouse-rat difference in the effect of upslope training was the result of differences in the types of outcome measures used in the two species or whether it reflects significant changes in motor unit reinnervation following the cessation of the exercise is not clear at this time. In contrast, the effect of upslope training on the amplitude of the restored H reflex is striking. Unlike the effect of level training, where an exaggerated H reflex is found 8–10 wk after nerve repair, a significant decrease in H reflex amplitude was found in upslope-trained rats relative to both level-trained and untrained animals. Whether this markedly different outcome was obtained simply by changing the exercise conditions reflects an effect on the sensory axon regeneration, the efficacy of the synaptic inputs onto the injured motoneurons, or both awaits further study. The nature of the exercise protocol applied can change the nature of at least this simple spinal circuit. It will be of considerable interest to study the effects of downslope treadmill training on restoration of muscle innervation and this spinal circuitry. Recruitment of fewer motoneurons into activity would be expected to occur during downslope walking than level walking (63), but activity of stretch-sensitive afferent neurons in intact synergists might be expected to be greater during downslope walking where prolonged periods of muscle stretch are encountered (50). Evaluation of the effects of different exercise protocols on postinjury properties of other reflex pathways, such as the crossed extensor reflex (79), will be equally interesting.
Conclusions
Activity-associated therapies such as exercise could be applied widely to enhance functional recovery following peripheral nerve injuries, addressing an important public health issue. They are inexpensive and enable patients to assume responsibility for their own recovery. However, application of these therapies to diverse types of nerve injuries in a diverse human population will require exercise prescriptions that are unique to the injured nerve, the desired regeneration of different types of axons, and the developmental stage and genetic background of the patients. Some principles that might form a basis for developing such prescriptions are emerging from a greater understanding of the cellular mechanisms underlying these therapies, but more studies are needed.
Footnotes
This work was supported by National Institutes of Health Grants NS-057190 and HD-032571 to A. W. English.
No conflicts of interest, financial or otherwise, are declared by the author(s).
Author contributions: A.W.E., J.C.W., and P.J.W. conception and design of research; A.W.E., J.C.W., and P.J.W. performed experiments; A.W.E., J.C.W., and P.J.W. analyzed data; A.W.E., J.C.W., and P.J.W. interpreted results of experiments; A.W.E., J.C.W., and P.J.W. prepared figures; A.W.E., J.C.W., and P.J.W. drafted manuscript; A.W.E., J.C.W., and P.J.W. edited and revised manuscript; A.W.E., J.C.W., and P.J.W. approved final version of manuscript.
References
- 1.Acaz-Fonseca E, Sanchez-Gonzalez R, Azcoitia I, Arevalo MA, Garcia-Segura LM. Role of astrocytes in the neuroprotective actions of 17beta-estradiol and selective estrogen receptor modulators. Mol Cell Endocrinol 389: 48–57, 2014. [DOI] [PubMed] [Google Scholar]
- 2.Al-Majed AA, Brushart TM, Gordon T. Electrical stimulation accelerates and increases expression of BDNF and trkB mRNA in regenerating rat femoral motoneurons. Eur J Neurosci 12: 4381–4390, 2000. [PubMed] [Google Scholar]
- 3.Al-Majed AA, Neumann CM, Brushart TM, Gordon T. Brief electrical stimulation promotes the speed and accuracy of motor axonal regeneration. J Neurosci 20: 2602–2608, 2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Al-Majed AA, Tam SL, Gordon T. Electrical stimulation accelerates and enhances expression of regeneration-associated genes in regenerating rat femoral motoneurons. Cell Mol Neurobiol 24: 379–402, 2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Alvarez FJ, Titus-Mitchell HE, Bullinger KL, Kraszpulski M, Nardelli P, Cope TC. Permanent central synaptic disconnection of proprioceptors after nerve injury and regeneration. I. Loss of VGLUT1/IA synapses on motoneurons. J Neurophysiol 106: 2450–2470, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Anastasia A, Deinhardt K, Chao MV, Will NE, Irmady K, Lee FS, Hempstead BL, Bracken C. Val66Met polymorphism of BDNF alters prodomain structure to induce neuronal growth cone retraction. Nature Comm 4: 2490, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Asensio-Pinilla E, Udina E, Jaramillo J, Navarro X. Electrical stimulation combined with exercise increase axonal regeneration after peripheral nerve injury. Exp Neurol 219: 258–265, 2009. [DOI] [PubMed] [Google Scholar]
- 8.Barreto G, Veiga S, Azcoitia I, Garcia-Segura LM, Garcia-Ovejero D. Testosterone decreases reactive astroglia and reactive microglia after brain injury in male rats: role of its metabolites, oestradiol and dihydrotestosterone. Eur J Neurosci 25: 3039–3046, 2007. [DOI] [PubMed] [Google Scholar]
- 9.Berchtold NC, Kesslak JP, Pike CJ, Adlard PA, Cotman CW. Estrogen and exercise interact to regulate brain-derived neurotrophic factor mRNA and protein expression in the hippocampus. Eur J Neurosci 14: 1992–2002, 2001. [DOI] [PubMed] [Google Scholar]
- 10.Boeltz T, Ireland M, Mathis K, Nicolini J, Poplavski K, Rose SJ, Wilson E, English AW. Effects of treadmill training on functional recovery following peripheral nerve injury in rats. J Neurophysiol 109: 2645–2657, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Brushart TM. Nerve Repair. New York: Oxford Univ. Press, 2011, p. 463. [Google Scholar]
- 12.Chen ZY, Ieraci A, Teng H, Dall H, Meng CX, Herrera DG, Nykjaer A, Hempstead BL, Lee FS. Sortilin controls intracellular sorting of brain-derived neurotrophic factor to the regulated secretory pathway. J Neurosci 25: 6156–6166, 2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Chen ZY, Jing D, Bath KG, Ieraci A, Khan T, Siao CJ, Herrera DG, Toth M, Yang C, McEwen BS, Hempstead BL, Lee FS. Genetic variant BDNF (Val66Met) polymorphism alters anxiety-related behavior. Science 314: 140–143, 2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Chen ZY, Patel PD, Sant G, Meng CX, Teng KK, Hempstead BL, Lee FS. Variant brain-derived neurotrophic factor (BDNF) (Met66) alters the intracellular trafficking and activity-dependent secretion of wild-type BDNF in neurosecretory cells and cortical neurons. J Neurosci 24: 4401–4411, 2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Coers S, Tanzer L, Jones KJ. Testosterone treatment attenuates the effects of facial nerve transection on glial fibrillary acidic protein (GFAP) levels in the hamster facial motor nucleus. Metab Brain Dis 17: 55–63, 2002. [DOI] [PubMed] [Google Scholar]
- 16.Conejo NM, Gonzalez-Pardo H, Cimadevilla JM, Arguelles JA, Diaz F, Vallejo-Seco G, Arias JL. Influence of gonadal steroids on the glial fibrillary acidic protein-immunoreactive astrocyte population in young rat hippocampus. J Neurosci Res 79: 488–494, 2005. [DOI] [PubMed] [Google Scholar]
- 17.Coull JA, Beggs S, Boudreau D, Boivin D, Tsuda M, Inoue K, Gravel C, Salter MW, De Koninck Y. BDNF from microglia causes the shift in neuronal anion gradient underlying neuropathic pain. Nature 438: 1017–1021, 2005. [DOI] [PubMed] [Google Scholar]
- 18.Dale-Nagle EA, Hoffman MS, MacFarlane PM, Mitchell GS. Multiple pathways to long-lasting phrenic motor facilitation. Adv Exp Med Biol 669: 225–230, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Day JR, Frank AT, O'Callaghan JP, Jones BC, Anderson JE. The effect of age and testosterone on the expression of glial fibrillary acidic protein in the rat cerebellum. Exp Neurol 151: 343–346, 1998. [DOI] [PubMed] [Google Scholar]
- 20.De Bono JP, Adlam D, Paterson DJ, Channon KM. Novel quantitative phenotypes of exercise training in mouse models. Am J Physiol Regul Integr Comp Physiol 290: R926–R934, 2006. [DOI] [PubMed] [Google Scholar]
- 21.Dougherty KD, Dreyfus CF, Black IB. Brain-derived neurotrophic factor in astrocytes, oligodendrocytes, and microglia/macrophages after spinal cord injury. Neurobiol Dis 7: 574–585, 2000. [DOI] [PubMed] [Google Scholar]
- 22.Egan MF, Kojima M, Callicott JH, Goldberg TE, Kolachana BS, Bertolino A, Zaitsev E, Gold B, Goldman D, Dean M, Lu B, Weinberger DR. The BDNF val66met polymorphism affects activity-dependent secretion of BDNF and human memory and hippocampal function. Cell 112: 257–269, 2003. [DOI] [PubMed] [Google Scholar]
- 23.English AW, Liu K, Nicolini JM, Mulligan AM, Ye K. Small-molecule trkB agonists promote axon regeneration in cut peripheral nerves. Proc Natl Acad Sci USA 110: 16217–16222, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.English AW, Meador W, Carrasco DI. Neurotrophin 4/5 is required for the early growth of regenerating axons in peripheral nerves. Eur J Neurosci 21: 2624–2634, 2005. [DOI] [PubMed] [Google Scholar]
- 25.English AW, Wilhelm JC, Sabatier MJ. Enhancing recovery from peripheral nerve injury using treadmill training. Ann Anat 193: 354–361, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Freeman LM, Padgett BA, Prins GS, Breedlove SM. Distribution of androgen receptor immunoreactivity in the spinal cord of wild-type, androgen-insensitive and gonadectomized male rats. J Neurobiol 27: 51–59, 1995. [DOI] [PubMed] [Google Scholar]
- 27.Funakoshi H, Frisen J, Barbany G, Timmusk T, Zachrisson O, Verge VM, Persson H. Differential expression of mRNAs for neurotrophins and their receptors after axotomy of the sciatic nerve. J Cell Biol 123: 455–465, 1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.García-Ovejero D, Veiga S, García-Segura L, Doncarlo SL. Glial expression of estrogen and androgen receptors after rat brain injury. J Comp Neurol 450: 256–271, 2002. [DOI] [PubMed] [Google Scholar]
- 29.Geremia NM, Gordon T, Brushart TM, Al-Majed AA, Verge VM. Electrical stimulation promotes sensory neuron regeneration and growth-associated gene expression. Exp Neurol 205: 347–359, 2007. [DOI] [PubMed] [Google Scholar]
- 30.Gomez-Pinilla F, Gomez AG. The influence of dietary factors in central nervous system plasticity and injury recovery. Pm R 3: S111–S116, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Gomez-Pinilla F, Vaynman S. A “deficient environment” in prenatal life may compromise systems important for cognitive function by affecting BDNF in the hippocampus. Exp Neurol 192: 235–243, 2005. [DOI] [PubMed] [Google Scholar]
- 32.Gomez-Pinilla F, Ying Z, Roy RR, Molteni R, Edgerton VR. Voluntary exercise induces a BDNF-mediated mechanism that promotes neuroplasticity. J Neurophysiol 88: 2187–2195, 2002. [DOI] [PubMed] [Google Scholar]
- 33.Gordon T. The role of neurotrophic factors in nerve regeneration. Neurosurg Focus 26: E3, 2009. [DOI] [PubMed] [Google Scholar]
- 34.Gordon T, Tyreman N, Raji MA. The basis for diminished functional recovery after delayed peripheral nerve repair. J Neurosci 31: 5325–5334, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Gottfried-Blackmore A, Sierra A, Jellinck PH, McEwen BS, Bulloch K. Brain microglia express steroid-converting enzymes in the mouse. J Steroid Biochem Mol Biol 109: 96–107, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Griesbeck O, Parsadanian AS, Sendtner M, Thoenen H. Expression of neurotrophins in skeletal muscle: quantitative comparison and significance for motoneuron survival and maintenance of function. J Neurosci Res 42: 21–33, 1995. [DOI] [PubMed] [Google Scholar]
- 37.Grillner S. Control of locomotion in bipeds, tetrapods, and fish. In: Handbook of Physiology. The Nervous System. Motor Control. Bethesda, MD: Am. Physiol. Soc., 1981, sect. 1, vol. II, pt. 2, chapt. 26, p. 1179–1236. [Google Scholar]
- 38.Gutmann E, Jakoubek B. Effect of increased motor activity on regeneration of the peripheral nerve in your rats. Physiol Bohemoslov 12: 463–468, 1963. [PubMed] [Google Scholar]
- 39.Herbison GJ, Jaweed MM, Ditunno JF, Scott CM. Effect of overwork during reinnervation of rat muscle. Exp Neurol 41: 1–14, 1973. [DOI] [PubMed] [Google Scholar]
- 40.Hoffer JA, Stein RB, Gordon T. Differential atrophy of sensory and motor fibers following section of cat peripheral nerves. Brain Res 178: 347–361, 1979. [DOI] [PubMed] [Google Scholar]
- 41.Hong EJ, McCord AE, Greenberg ME. A biological function for the neuronal activity-dependent component of Bdnf transcription in the development of cortical inhibition. Neuron 60: 610–624, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Hosli E, Jurasin K, Ruhl W, Luthy R, Hosli L. Colocalization of androgen, estrogen and cholinergic receptors on cultured astrocytes of rat central nervous system. Int J Dev Neurosci 19: 11–19, 2001. [DOI] [PubMed] [Google Scholar]
- 43.Huppenbauer CB, Tanzer L, DonCarlos LL, Jones KJ. Gonadal steroid attenuation of developing hamster facial motoneuron loss by axotomy: equal efficacy of testosterone, dihydrotestosterone, and 17-beta estradiol. J Neurosci 25: 4004–4013, 2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Hutchinson KJ, Gomez-Pinilla F, Crowe MJ, Ying Z, Basso DM. Three exercise paradigms differentially improve sensory recovery after spinal cord contusion in rats. Brain 127: 1403–1414, 2004. [DOI] [PubMed] [Google Scholar]
- 45.Islamov RR, Hendricks WA, Jones RJ, Lyall GJ, Spanier NS, Murashov AK. 17Beta-estradiol stimulates regeneration of sciatic nerve in female mice. Brain Res 943: 283–286, 2002. [DOI] [PubMed] [Google Scholar]
- 46.Islamov RR, Hendricks WA, Katwa LC, McMurray RJ, Pak ES, Spanier NS, Murashov AK. Effect of 17 beta-estradiol on gene expression in lumbar spinal cord following sciatic nerve crush injury in ovariectomized mice. Brain Res 966: 65–75, 2003. [DOI] [PubMed] [Google Scholar]
- 47.Jones LN, Dougherty S, Wilhelm JC, English AW. Enhancement of axon regeneration in cut peripheral nerves using optical stimulation. Program No. 207.01. In: Neuroscience Meeting Planner (Online) New Orleans, LA: Society for Neuroscience, 2012. http://am2012.sfn.org/am2012/pdf/final_program/final_program_b3.pdf. [Google Scholar]
- 48.Lang EM, Schlegel N, Reiners K, Hofmann GO, Sendtner M, Asan E. Single-dose application of CNTF and BDNF improves remyelination of regenerating nerve fibers after C7 ventral root avulsion and replantation. J Neurotrauma 25: 384–400, 2008. [DOI] [PubMed] [Google Scholar]
- 49.Lee R, Kermani P, Teng KK, Hempstead BL. Regulation of cell survival by secreted proneurotrophins. Science 294: 1945–1948, 2001. [DOI] [PubMed] [Google Scholar]
- 50.Maas H, Gregor RJ, Hodson-Tole EF, Farrell BJ, English AW, Prilutsky BI. Locomotor changes in length and EMG activity of feline medial gastrocnemius muscle following paralysis of two synergists. Exp Brain Res 203: 681–692, 2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Marqueste T, Alliez JR, Alluin O, Jammes Y, Decherchi P. Neuromuscular rehabilitation by treadmill running or electrical stimulation after peripheral nerve injury and repair. J Appl Physiol 96: 1988–1995, 2004. [DOI] [PubMed] [Google Scholar]
- 52.Meehan CF, Grondahl L, Nielsen JB, Hultborn H. Fictive locomotion in the adult decerebrate and spinal mouse in vivo. J Physiol 590: 289–300, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Menard A, Leblond H, Gossard JP. The modulation of presynaptic inhibition in single muscle primary afferents during fictive locomotion in the cat. J Neurosci 19: 391–400, 1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Molteni R, Zheng JQ, Ying Z, Gomez-Pinilla F, Twiss JL. Voluntary exercise increases axonal regeneration from sensory neurons. Proc Natl Acad Sci USA 101: 8473–8478, 2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Moore DL, Goldberg JL. Multiple transcription factor families regulate axon growth and regeneration. Dev Neurobiol 71: 1186–1211, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Murashov AK, Islamov RR, McMurray RJ, Pak ES, Weidner DA. Estrogen increases retrograde labeling of motoneurons: evidence of a nongenomic mechanism. Am J Physiol Cell Physiol 287: C320–C326, 2004. [DOI] [PubMed] [Google Scholar]
- 57.Nakajima K, Honda S, Tohyama Y, Imai Y, Kohsaka S, Kurihara T. Neurotrophin secretion from cultured microglia. J Neurosci Res 65: 322–331, 2001. [DOI] [PubMed] [Google Scholar]
- 58.Navarro X, Vivo M, Valero-Cabre A. Neural plasticity after peripheral nerve injury and regeneration. Prog Neurobiol 82: 163–201, 2007. [DOI] [PubMed] [Google Scholar]
- 59.Neeper SA, Gomez-Pinilla F, Choi J, Cotman CW. Physical activity increases mRNA for brain-derived neurotrophic factor and nerve growth factor in rat brain. Brain Res 726: 49–56, 1996. [PubMed] [Google Scholar]
- 60.Okamoto M, Hojo Y, Inoue K, Matsui T, Kawato S, McEwen BS, Soya H. Mild exercise increases dihydrotestosterone in hippocampus providing evidence for androgenic mediation of neurogenesis. Proc Natl Acad Sci USA 109: 13100–13105, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Park JS, Hoke A. Treadmill exercise induced functional recovery after peripheral nerve repair is associated with increased levels of neurotrophic factors. PLos One 9: e90245, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Sabatier M, Redmon N, Schwartz G, English A. Treadmill training promotes axon regeneration in injured peripheral nerves. Exp Neurol 211: 489–493, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Sabatier MJ, To BN, Nicolini J, English AW. Effect of slope and sciatic nerve injury on ankle muscle recruitment and hindlimb kinematics during walking in the rat. J Exp Biol 214: 1007–1016, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Salerno KM, Jing X, Diges CM, Davis BM, Albers KM. TRAF family member-associated NF-kappa B activator (TANK) expression increases in injured sensory neurons and is transcriptionally regulated by Sox11. Neuroscience 231: 28–37, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Scholz T, Krichevsky A, Sumarto A, Jaffurs D, Wirth GA, Paydar K, Evans GR. Peripheral nerve injuries: an international survey of current treatments and future perspectives. J Reconstr Microsurg 25: 339–344, 2009. [DOI] [PubMed] [Google Scholar]
- 66.Shah PKGA, G, Choe J, Gad P, Gerasimenko Y, Tillakaratne N, Zhong H, Roy RR, Edgerton VR. Use of quadrupedal step training to re-engage spinal interneuronal networks and improve locomotor function after spinal cord injury. Brain 136: 3362–3377, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Sharma N, Marzo SJ, Jones KJ, Foecking EM. Electrical stimulation and testosterone differentially enhance expression of regeneration-associated genes. Exp Neurol 223: 183–191, 2010. [DOI] [PubMed] [Google Scholar]
- 68.Sohrabji F, Lewis DK. Estrogen-BDNF interactions: implications for neurodegenerative diseases. Front Neuroendocrinol 27: 404–414, 2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Struble RG, Nathan BP, Cady C, Cheng X, McAsey M. Estradiol regulation of astroglia and apolipoprotein E: an important role in neuronal regeneration. Exp Gerontol 42: 54–63, 2007. [DOI] [PubMed] [Google Scholar]
- 70.Sun W, Park KW, Choe J, Rhyu IJ, Kim IH, Park SK, Choi B, Choi SH, Park SH, Kim H. Identification of novel electroconvulsive shock-induced and activity-dependent genes in the rat brain. Biochem Biophys Res Commun 327: 848–856, 2005. [DOI] [PubMed] [Google Scholar]
- 71.Tanzer L, Jones KJ. Gonadal steroid regulation of hamster facial nerve regeneration: effects of dihydrotestosterone and estradiol. Exp Neurol 146: 258–264, 1997. [DOI] [PubMed] [Google Scholar]
- 72.Taylor CA, Braza D, Rice JB, Dillingham T. The incidence of peripheral nerve injury in extremity trauma. Am J Phys Med Rehabil 87: 381–385, 2008. [DOI] [PubMed] [Google Scholar]
- 73.Teng HK, Teng KK, Lee R, Wright S, Tevar S, Almeida RD, Kermani P, Torkin R, Chen ZY, Lee FS, Kraemer RT, Nykjaer A, Hempstead BL. ProBDNF induces neuronal apoptosis via activation of a receptor complex of p75NTR and sortilin. J Neurosci 25: 5455–5463, 2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Teodori RM, Betini J, de Oliveira LS, Sobral LL, Takeda SY, de Lima Montebelo MI. Swimming exercise in the acute or late phase after sciatic nerve crush accelerates nerve regeneration. Neural Plast 2011: 783901, 2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Thompson NJ, Sengelaub DR, English AW. Enhancement of peripheral nerve regeneration due to treadmill training and electrical stimulation is dependent on androgen receptor signaling. Dev Neurobiol 74: 531–540, 2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Udina E, Cobianchi S, Allodi I, Navarro X. Effects of activity-dependent strategies on regeneration and plasticity after peripheral nerve injuries. Ann Anat 193: 347–353, 2011. [DOI] [PubMed] [Google Scholar]
- 77.Udina E, Puigdemasa A, Navarro X. Passive and active exercise improve regeneration and muscle reinnervation after peripheral nerve injury in the rat. Muscle Nerve 43: 500–509, 2011. [DOI] [PubMed] [Google Scholar]
- 78.Ulmann L, Hatcher JP, Hughes JP, Chaumont S, Green PJ, Conquet F, Buell GN, Reeve AJ, Chessell IP, Rassendren F. Up-regulation of P2X4 receptors in spinal microglia after peripheral nerve injury mediates BDNF release and neuropathic pain. J Neurosci 28: 11263–11268, 2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Valero-Cabre A, Navarro X. Changes in crossed spinal reflexes after peripheral nerve injury and repair. J Neurophysiol 87: 1763–1771, 2002. [DOI] [PubMed] [Google Scholar]
- 80.van Meeteren NL, Brakkee JH, Hamers FP, Helders PJ, Gispen WH. Exercise training improves functional recovery and motor nerve conduction velocity after sciatic nerve crush lesion in the rat. Arch Phys Med Rehabil 78: 70–77, 1997. [DOI] [PubMed] [Google Scholar]
- 81.van Meeteren NL, Brakkee JH, Helders PJ, Gispen WH. The effect of exercise training on functional recovery after sciatic nerve crush in the rat. J Peripher Nerv Syst 3: 277–282, 1998. [PubMed] [Google Scholar]
- 82.Verhovshek T, Sengelaub DR. Androgen action at the target musculature regulates brain-derived neurotrophic factor protein in the spinal nucleus of the bulbocavernosus. Dev Neurobiol 73: 587–598, 2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Wilhelm JC, Cucoranu D, Xu M, Chmielewski S, Holmes T, Lau KS, Bassell GJ, English AW. Cooperative roles of BDNF expression in neurons and Schwann cells are modulated by exercise to facilitate nerve regeneration. J Neurosci 32: 5002–5009, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Wood K, Wilhelm JC, Sabatier MJ, English AW. Sex differences in the effects of treadmill training on axon regeneration in cut peripheral nerves. Dev Neurobiol 72: 688–698, 2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Yamamori H, Hashimoto R, Ishima T, Kishi F, Yasuda Y, Ohi K, Fujimoto M, Umeda-Yano S, Ito A, Hashimoto K, Takeda M. Plasma levels of mature brain-derived neurotrophic factor (BDNF) and matrix metalloproteinase-9 (MMP-9) in treatment-resistant schizophrenia treated with clozapine. Neurosci Lett 556: 37–41, 2013. [DOI] [PubMed] [Google Scholar]
- 86.Zhang JY, Luo XG, Xian CJ, Liu ZH, Zhou XF. Endogenous BDNF is required for myelination and regeneration of inured sciatic nerve in rodents. Eur J Neurosci 12: 4171–4180, 2000. [PubMed] [Google Scholar]