

Significance
Fire is a key ecological process affecting ecosystem dynamics and services. Fire frequency, intensity, and size reflect complex climate–vegetation–human interactions and their evolution through time. The long-term history of these interactions provides insights into the variability of the ecosystem and a context for future environmental change. We use paleoenvironmental data from the Patagonian forest/steppe border and introduce generalized additive models as trend-detection tools in paleoecology to assess the relative influence of climate, vegetation, and humans in shaping the fire regime. Our results suggest that for the last 18,000 y, fires have been predominantly limited by fuel discontinuity rather than by suitable climate conditions. In contrast to extensive present-day anthropogenic impact, Patagonian ecosystems evolved with minimal human influence prior to European settlement.
Keywords: charcoal, fire, climate, anthropogenic impact, Patagonia
Abstract
Forest/steppe boundaries are among the most dynamic ecosystems on Earth and are highly vulnerable to changes in climate and land use. In this study we examine the postglacial history of the Patagonian forest/steppe ecotone (41–43°S) to better understand its sensitivity to past variations in climate, disturbance, and human activity before European colonization. We present regional trends in vegetation and biomass burning, as detected by generalized additive models fitted to seven pollen and charcoal records, and compare the results with other paleoenvironmental data, as well as archeological and ecological information to (i) estimate postglacial fire trends at regional scales, (ii) assess the evolution of climate–vegetation–fire linkages over the last 18,000 calibrated (cal) years B.P., and (iii) evaluate the role of humans in altering pre-European landscapes and fire regimes. Pollen and charcoal data indicate that biomass burning was relatively low during warm/dry steppe-dominated landscapes in the late glacial/Early Holocene transition and increased as more humid conditions favored forest development after ca. 10,000 cal years B.P. Postglacial fire activity was thus limited by fuel availability associated with sparse vegetation cover rather than by suitable climate conditions. In contrast to extensive burning by European settlers, variations in indigenous population densities were not associated with fluctuations in regional or watershed-scale fire occurrence, suggesting that climate–vegetation–fire linkages in northern Patagonia evolved with minimal or very localized human influences before European settlement.
Fire is a significant disturbance agent in most terrestrial ecosystems. The characteristic frequency, size, and intensity of fire (i.e., the fire regime) are driven by interactions between top–down drivers, such as average climatic conditions, weather variability and frequency of ignition, and bottom–up processes related to fuel load, topography, and soil. These interactions operate from local to global scales and over time scales of hours to millennia (1, 2). Disentangling large-scale from local-scale controls on particular fire events, as well as climatic from nonclimatic drivers of fire regimes, are grand challenges in Earth systems science (3, 4) and are topics where paleoecology, with its long-time perspective, has made substantial contributions (5–7).
Currently, natural fire regimes around the world are being altered by deliberate burning, fire elimination, and changes in fuel distribution associated with land-use practices. Human–fire relationships, nonetheless, have evolved as economies shifted from hunting–gathering to pastoralism, farming, and industrialization (8). Thus, the changing role of humans has to be factored into an understanding of past ecosystem dynamics, recognizing that their influence is also scale, time, and location dependent.
Woodland/grassland borders are areas of particular interest in this regard because they are highly sensitive to climatic variability and disturbance and have been occupied and used by humans for millennia (9). The Patagonian forest/steppe ecotone along the eastern flank of the Andes is among the most striking vegetation borders in the world. Its composition and location are influenced by the position and strength of the Southern Westerlies (10) and the strong orographic barrier created by the Andes, as well as by the interplay between fire and a complex fuel mosaic.
Humans have been present in Patagonia for the last 14,000 y and have used the region for hunting, gathering, and foraging. Intensifying land-use practices, such as deforestation, livestock herbivory, and cultivation, occurred with European colonization in the 17th century and modified fuel conditions, postfire vegetation recovery, and the timing and severity of fire occurrence (11). Recent population growth along the forest/steppe ecotone has only exacerbated natural environmental stresses, and now this region is facing unprecedented fires, water and nutrient shortages, and threats to biodiversity with consequences for economic sustainability (12, 13).
In this study we examine the postglacial fire history of the forest/steppe border to better understand its sensitivity to past variations in climate, vegetation composition, and human activity before European colonization. We present regional composite pollen and charcoal records developed from a network of lakes along the forest/steppe border and compare the results with paleoclimatic data, as well as archeological and ecological information. Our objectives are to (i) estimate postglacial fire trends at regional scales, (ii) assess the evolution of climate–vegetation–fire linkages over the last 18,000 calibrated (cal) years B.P., and (iii) evaluate the role of humans in altering pre-European landscapes and fire regimes.
Setting
The climate of northern Patagonia is largely driven by the interplay between westerly storm tracks, guided by the Southern Westerlies, and the north–south-trending Andes. The latitudinal position of storm tracks fluctuates seasonally in response to shifts in the strength and position of the southeastern Pacific high-pressure system and a subpolar low-pressure trough centered along the Antarctic Circle (10). The uplift of low-level winds over the western slope of the Andes produces continental orographic precipitation. Forced subsidence on the Argentine flanks causes adiabatic warming of the air masses and dry conditions in eastern Patagonia (14). Mechanisms for interannual-to-decadal climate variability are related to tropospheric forcing of midlatitude sea surface temperatures, indirect effects of coupled ocean–atmosphere phenomena rooted in the tropical Pacific [El Niño Southern Oscillation (ENSO)] (15), and pressure anomalies resulting from the fluctuations in strength and position of the polar vortex [Southern Annular Mode (SAM)] (16).
Vegetation composition in the study region is largely explained by the decline in effective moisture from Andean crest to the eastern flanks and plains. The montane slopes of the Andes are dominated by tall forests of the evergreen Nothofagus pumilio (>1,500 m elevation) and by Nothofagus dombeyi (1,500–1,200 m elevation). Between 1,200 and 600 m elevation, N. dombeyi and Austrocedrus chilensis codominate in low-elevation dry forests. These forests become more open and support diverse shrubs (e.g., Colletia, Discaria, Maytenus) in progressively drier settings to the east. At the eastern flank of the Andes, forests give way to the sparse xerophytic taxa of the northern Patagonian steppe (e.g., Stipa, Festuca, Mulinum; Fig. 1) (17).
Fig. 1.
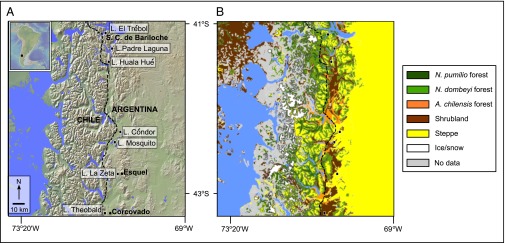
(A) Study area and location of Laguna El Trébol, Laguna Padre Laguna, Lago Huala Hué, Lago Cóndor, Lago Mosquito, Laguna La Zeta, and Lago Theobald, and the main vegetation types. The dashed line shows the position of the modern forest/steppe ecotone. (B) Main vegetation types in the region. Modified with permission from ref. 21.
Climate also plays a significant role in shaping present-day fire dynamics. Tree-ring records spanning the last 500 y point to moisture deficits during the fire season (November–March) linked to the positive phase of ENSO as an important driver of fire activity (18, 19). Similarly, area burned has been attributed to warmer and drier than average springs associated with the positive phase of SAM (20). Such linkages suggest a strong correlation between regional biomass burning and interannual climate variability.
Results
Lithology, Pollen, and Charcoal Data.
Age–depth relations for each record were developed from a sequence of accelerator MS (AMS) radiocarbon dates on terrestrial plant macrofossils within the cores. The shortest record (L. Padre Laguna) was ca. 5,000 cal years long, and the oldest site was L. La Zeta, extending back >18,000 y. The chronology, lithology, and vegetation and fire history for each site are discussed in detail by Iglesias et al. (21) and will only be described briefly.
The sediment cores at all of the sites featured similar sequential lithologic transitions from glacial laminated clay to Early Holocene organic silt and then Middle and Late Holocene fine-detritus gyttja. At L. Huala Hué, cores were taken from fen margins adjacent to the open lake, so the uppermost sediments were peat. Lithologic transitions are coupled with a long-term trend toward increased sedimentation rates and organic matter content, suggesting a shift in sediment source from glacial outwash to Holocene autochthonous biogenic sediment associated with increased lake productivity.
The charcoal and pollen stratigraphy was also broadly similar among sites (Figs. 2–8). In the late glacial period (>16,000 cal years B.P.), the pollen data from L. El Trébol and L. La Zeta indicate that steppe dominated by Poaceae (>32%), Asteraceae (<18%), and other steppe taxa (<38%) prevailed on the eastern flanks of the Andes (22). A charcoal accumulation rate (CHAR) <0.03 particles per cm2⋅y−1 suggests negligible biomass burning at this time. Between ca. 16,000 and 8,000 cal years B.P., percentages of N. dombeyi-type pollen increased at all sites (>70%). Higher percentages of arboreal taxa indicate development of late glacial/Early Holocene parkland (23). CHAR values increased up to 20 particles per cm2⋅y−1 at all sites at ca. 15,000 cal years B.P., implying that biomass burning was higher than before.
Fig. 2.

Pollen percentages of forest taxa and CHAR time series for L. El Trébol. Expected CHAR values for each site based on the GAM fitted to the charcoal data are provided for reference (in red). Watershed-scale anomalies in biomass burning as inferred from outliers in the residuals of the CHAR model are indicated with a “+”. Positive anomalies represent periods of higher local biomass activity than the estimated regional trend. Inferred human population densities near the watershed are also shown.
Fig. 8.

Pollen percentages of forest taxa and CHAR time series for L. Theobald. Expected CHAR values for each site based on the GAM fitted to the charcoal data are provided for reference (in red). Watershed-scale anomalies in biomass burning as inferred from outliers in the residuals of the CHAR model are indicated with a “+”. Positive anomalies represent periods of higher local biomass activity than the estimated regional trend. Inferred human population densities near the watershed are also shown.
Fig. 5.

Pollen percentages of forest taxa and CHAR time series for L. Cóndor. Expected CHAR values for each site based on the GAM fitted to the charcoal data are provided for reference (in red). Watershed-scale anomalies in biomass burning as inferred from outliers in the residuals of the CHAR model are indicated with a “+”. Positive anomalies represent periods of higher local biomass activity than the estimated regional trend. Inferred human population densities near the watershed are also shown.
Fig. 6.

Pollen percentages of forest taxa and CHAR time series for a L. Mosquito. Expected CHAR values for each site based on the GAM fitted to the charcoal data are provided for reference (in red). Watershed-scale anomalies in biomass burning as inferred from outliers in the residuals of the CHAR model are indicated with a “+”. Positive anomalies represent periods of higher local biomass activity than the estimated regional trend. Inferred human population densities near the watershed are also shown.
Fig. 7.

Pollen percentages of forest taxa and CHAR time series for L. La Zeta. Expected CHAR values for each site based on the GAM fitted to the charcoal data are provided for reference (in red). Watershed-scale anomalies in biomass burning as inferred from outliers in the residuals of the CHAR model are indicated with a “+”. Positive anomalies represent periods of higher local biomass activity than the estimated regional trend. Inferred human population densities near the watershed are also shown.
During the Middle Holocene (ca. 8,000–45,00 cal years B.P.), Cupressaceae pollen percentages rose at all sites (>10%), suggesting a regional increase in Austrocedrus abundance. High percentages of Rhamnaceae (<8%) and Maytenus (<7%) at all sites indicate that the study area featured shrubland vegetation (22). CHAR values ranged from up to 20 particles per cm2⋅y−1 at L. El Trébol, L. Huala Hué, and L. Mosquito to >0.8 particles per cm2⋅y−1 in the remaining watersheds; spatial variations in abundance suggest relatively heterogeneous fire patterns at this time.
After 4,500 cal years B.P., pollen records indicate an increase of N. dombeyi-type (<80%) and Cupressaceae pollen (<58%) and marks the establishment of mixed Nothofagus–Austrocedrus forest along the ecotone (23). Levels of biomass burning increased at L. Huala Hué (CHAR > 0.3 particles per cm2⋅y−1), L. Mosquito (CHAR > 2 particles per cm2⋅y−1), and L. Theobald (CHAR > 0.4 particles per cm2⋅y−1) at the beginning of the period, but CHAR values varied little at L. El Trébol and L. Padre Laguna and decreased at L. Cóndor (from 1.3 to 1 particles per cm2⋅y−1) at 4,500 cal years B.P. This comparison suggests relatively high fire activity in the study area, especially toward the west. Although human settlement at/near the study sites occurred in the Late Holocene (specially after ca. 2,000 cal years B.P.) (24), the probability of anomalous fire activity at the watershed scale during this period was not different from that of the preoccupation period (Table S1). The similarity implies that anthropogenic burning was not pushing regional fire activity beyond a level predicted by climate/fuel conditions.
Time-Series Analysis Results.
About 18,000 y of data encompassing 553 pollen samples and 5,873 charcoal samples were used to construct regional composite records and estimate regional trends in vegetation and fire activity. The relative abundance of forest taxa was inferred from the counts of forest taxa pollen types (forest taxai). A generalized additive model (GAM) (Eq. 1) that assumed forest taxai to be negative binomial distributed with mean μI × terrestrial pollen sumi−1 and variance μi × terrestrial pollen sumi−1 + (μi × terrestrial pollen sumi−1)2/2.3 outperformed all of the other models [Δ Akaike’s information criterion (AIC) >22; Table S2 and Fig. 9]:
| [1] |
where the ith ratio of forest pollen types to terrestrial pollen sum in the time series was modeled as a smoothing function of the concatenated time series of all sites [f(timei) =(timeij), with n = total number of covariates and s(timeij) = smoothing function], and the nominal variable, sitei. log(1/terrestrial pollen sum)i is the offset and εi is the residual error.
Fig. 9.

Glacier advances (25–27), oxygen isotope ratios from Taylor Dome (28), precipitation off shore in Chile at 41°S (29), and ENSO (30). These independent paleoclimate records are compared with regional trends in forest taxa percentages and CHAR, which have been estimated with GAMs and converted to the appropriate units (i.e., pollen percentages and charcoal particles cm2⋅y−1; this study). The gray area represents 95% confidence intervals. Inferred human population densities along the transect are also shown.
L. Padre Laguna was arbitrarily chosen as the baseline site, which thus defines the base intercept, alpha (α). Alpha was estimated to be 11.09 (SE = 0.15, z = 72.85, P < 0.001). The intercepts for the other sites (i.e., sitei) were 11.79 (SE = 0.19, z = −3.77, P = 3.54) for L. El Trébol, 11.55 (SE = 0.21, z = 2.22, P = 0.02) for L. Huala Hué, 11.51 (SE = 0.19, z = −2.21, P = 0.02) for L. Cóndor, 10.91 (SE = 0.17, z = −1.07, P = 0.28) for L. Mosquito, 10.83 (SE = 0.19, z = −1.36, P = 0.57) for L. La Zeta, and 10.97 (SE = −0.57, z = −0.56, P = 0.57) for L. Theobald. The model explained 55.5% of the deviance.
The postglacial trend in regional biomass burning as inferred from charcoal data were best detected by a GAM (Eq. 2) that assumed charcoali to be negative binomial distributed with mean μi × accumulation ratei and variance μi × accumulation ratei + (μi × accumulation ratei)2/10.5. The GAM explained 59.4% of the deviance (ΔAIC > 153; Table S2 and Fig. 9):
| [2] |
where the ith charcoal count per unit of accumulation rate is a function of time [f(timei) = (timeij), with n = total number of covariates and s(timeij) is a smoothing function], log(1/accumulation rate)i is the offset, and εi is the residual error.
The intercept for the baseline site (i.e., L. Padre Laguna), α, was estimated to be 6.05 (SE = 0.02, z = 302, P < 0.001). The intercepts for the remaining sites, sitei, were 7.83 (SE = 0.02, z = 74, P < 0.001) for L. El Trébol, 6.15 (SE = 0.02, z = 4.23, P < 0.001) for L. Huala Hué, 6.95 (SE = 0.02, z = 34.56, P < 0.001) for L. Cóndor, 5.00 (SE = 0.02, z = −47.19, P < 0.001) for L. Mosquito, 7.11 (SE = 0.02, z = 71.80, P < 0.001) for L. La Zeta, and 5.30 (SE = 0.02, z = −30.54, P < 0.001) for L. Theobald. The model results are expressed on a logarithmic scale.
Discussion
Regional Linkages Between Climate, Vegetation, and Fire History.
Present-day plant communities are just one realization from a suite of ecological and climate conditions and historical events. Both moisture availability and fire have been proposed to limit forest expansion into grasslands (31). Support for the moisture-availability hypothesis comes from the African savanna and the Australian Northern Territories, where Bond (32) found a strong positive correlation between mean annual precipitation and woody cover, such that the ecotone was a pronounced transition from Eucalyptus spp. forest to savanna. In contrast, the ecotone from mixed forest to Great Plains prairie in central North America also exhibits sharp ecosystem boundaries despite its location along a gradual precipitation gradient. It has been proposed that North American prairie/forest border does not respond linearly to climate but rather the dynamics are governed by feedbacks involving vegetation and fire (33).
The results from northern Patagonia are consistent with the savanna model, inasmuch as they show strong linkages among climate, vegetation, and fire that track well with pronounced past changes in atmospheric circulation. During the Last Glacial Maximum, annual insolation was at the minimum value of the last 23,000 y. Temperatures in Antarctica, inferred from oxygen isotopes ratios from the Taylor Dome ice core (28), were low, leading to a strengthening and/or shift of the Southern Westerlies northward of their present-day position (29). As a result, cold dry conditions prevailed throughout Patagonia (34–36). Sparse steppe-dominated vegetation and negligible fire activity characterized the eastern flanks of the Andes (Figs. 2–9). Under the prevailing arid conditions, lack of fuel continuity and possibly ignitions were the primary constraint on fire occurrence.
After 16,000 cal years B.P., winter and annual insolation began to increase (37) and so too did Antarctic temperatures (28), favoring a gradual weakening and/or southward shift of the Southern Westerlies (36, 38). This trend toward warmer conditions and longer growing seasons peaked in the Early Holocene (ca. 10,000 cal years B.P.) and was associated with the expansion of tree populations (rising arboreal pollen percentages) and increasing fire activity (rising CHAR). High fire activity in the Early Holocene was promoted by increasing fuel load and amplified by the low-amplitude seasonal cycle of insolation coupled with high annual insolation, which may have created dry warm winters and early onset of the fire season (38).
During the last 10,000 y, winter insolation decreased, the Southern Westerlies gradually shifted to their current position, and ENSO variability strengthened in the region, especially after ca. 5,000 cal years B.P. (38). The long-term trend from Early Holocene aridity to Late Holocene cool wet and variable climate conditions favored gradual tree population establishment and expansion. Comparison of Holocene regional trends in tree abundance, fire activity, temperature (28), and precipitation (29) suggests that biomass burning was relatively low during warm/dry steppe-dominated landscapes in the late glacial/Early Holocene transition, and increased as more-humid-than-before conditions allowed forest development and higher vegetation cover during the Middle and Late Holocene. The close link between fuel load (forest taxa pollen) and biomass burning (CHAR) suggests that conditions in the late glacial and Early Holocene were conducive for fires, but discontinuous fuel cover limited their occurrence along the ecotone (39). We propose that gradual Holocene moisture-controlled expansion of forest taxa into steppe provided increased biomass and fuel continuity and allowed a progressive rise in fire activity.
The Late Holocene increase in fire activity was associated with the strengthening of ENSO frequency/intensity inasmuch as maximum fire occurrence (highest CHAR) followed the increased frequency of ENSO events after ca. 3,000 cal years B.P. (Fig. 9). In northern Patagonia, present-day El Niño conditions induce positive anomalies in winter precipitation that enhance fine fuel production during the growing season. Widespread fires tend to follow the switching from El Niño to La Niña conditions by favoring desiccation of fuels accumulated in previous years (20, 39). Thus in the last three millennia a combination of increased vegetation cover and heightened interannual variability allowed build up of fuels, regular desiccation of fuels, and ultimately high fire activity.
Our results suggest that climate variability drove regional trends in vegetation productivity over the last 18,000 y. For most of that time span, vegetation was the proximal control of fire activity, and long-term trends in climate served as a large-scale distal control of regional fire activity. Nonetheless it is important to note that our GAMs explain 55.5% and 59.4% of the deviance in the pollen and charcoal time series, respectively. We interpret the unexplained deviance as noise and site-specific anomalies in vegetation and fire. Microclimate conditions and nonclimatic factors such as topography, soil, and biophysical feedbacks therefore also had a significant influence on the vegetation and fire history at watershed scales.
The Role of Humans in Altering the Fire Regime.
The role of prehistoric humans in shaping the composition and location of the forest/steppe border rests on spatial extrapolation of very limited archeological data and a few observations by European explorers. In northern Patagonia, early immigrants and missionaries reported fires set by aboriginal peoples to hunt guanacos (Lama guanicoe) and rheas (Rhea spp.) at lower treeline (40, 41). Archeological data indicate that humans were present north of the study area (Huemul Cave, lat 36°56′S) (42) and at L. El Trébol (43) as early as ca. 10,600 cal years B.P.; near L. Huala Hué, L. Padre Laguna, L. La Zeta, and L. Theobald at ca. 3,200 cal years B.P. (44); and in the L. Mosquito and L. Cóndor area at ca. 1,750 cal years B.P. (45, 46). The low density of archeological findings and limited range of artifact types, however, suggest that the ecotone was not densely populated until ca. 2,000 cal years B.P. (24). At the regional scale, human presence before this date was limited to the steppe and hunter–gatherers who accessed the forest sporadically (24, 47, 48).
The scarcity of archeological data makes it difficult to infer the extent to which indigenous people used fire to manipulate the vegetation. In other parts of the world, hunter–gatherers have used fire to facilitate travel, hunting, and sometimes warfare, as well as to gather and maintain important plant resources (49, 50). Although it is possible that fire was required for the same purposes in northern Patagonia, there is no evidence that anthropogenic burning altered the broad patterns of fire (inferred from the generalized CHAR trends) inasmuch as shifts in biomass burning seem well explained by changes in fuel availability and climate (Fig. 9).
Regional fire activity was highest between 3,000 and 1,900 cal years B.P., peaking at ca. 2,500 cal years B.P. Human and animal bones, projectile points, stone artifacts, mineral pigments, and unidentified fragments at occupation sites nonetheless indicate that human populations did not increase until ca. 2,000 cal years B.P. (24, 44–46). The mismatch between highest regional fire activity occurring when human groups were still relatively small suggests a limited role for broad-scale anthropogenic burning. At the least a lack of synchrony between maximum fire activity and the onset of population expansion implies that human use of fire was not linearly dependent on population density.
Analysis of watershed trends in vegetation and fire anomalies (i.e., pollen data and local departures from regional biomass burning, respectively) does not reveal shifts in vegetation or anomalous fire activity specifically associated with changes in human populations (Figs. 2 and 4–8). The only exception is L. Padre Laguna (Fig. 3), where higher levels of biomass burning after ca. 2,000 cal years B.P. coincide with archeological evidence of rapid population growth in the vicinity of the lake. The other sites do not show persistent positive residual charcoal pattern (Figs. 2 and 4–8) or higher-than-expected probability of fire (Table S1) to suggest that local fire was elevated as a consequence of human manipulation. Thus, although it is possible that deliberate burning altered the landscape, climate–vegetation–fire linkages in northern Patagonia probably evolved with limited human influence before European settlement.
Fig. 4.

Pollen percentages of forest taxa and CHAR time series for L. Huala Hué. Expected CHAR values for each site based on the GAM fitted to the charcoal data are provided for reference (in red). Watershed-scale anomalies in biomass burning as inferred from outliers in the residuals of the CHAR model are indicated with a “+”. Positive anomalies represent periods of higher local biomass activity than the estimated regional trend. Inferred human population densities near the watershed are also shown.
Fig. 3.

Pollen percentages of forest taxa and CHAR time series for L. Padre Laguna. Expected CHAR values for each site based on the GAM fitted to the charcoal data are provided for reference (in red). Watershed-scale anomalies in biomass burning as inferred from outliers in the residuals of the CHAR model are indicated with a “+”. Positive anomalies represent periods of higher local biomass activity than the estimated regional trend. Inferred human population densities near the watershed are also shown.
The weak relation between fire and prehistoric humans is in contrast to the influence that European settlement had on fire regimes. By altering the probability of ignition through accidental and deliberate burning, and converting large areas of native forest to fire-prone communities (e.g., pine and eucalyptus plantations), Europeans have gradually increased the risk of fire in Patagonia (12). This trend is likely to continue into the future with drier climates, threatening the regeneration of fire-sensitive keystone species like A. chilensis (51).
Materials and Methods
Rationale.
Understanding ecotones is challenging because their dynamics result from the interactions of several components (e.g., species, soil, climate, disturbance) at multiple scales. A useful theoretical model for testing hypotheses regarding complex systems is that of a hierarchy of factors acting at different levels, wherein the higher levels of interconnections constrain lower ones to various degrees (52). In this framework, the relative importance of a given process is scale dependent (53–56).
Based on this approach, climate, vegetation, and anthropogenic impact are expected to be the large-scale controls of fire regimes (5). If conditions at higher levels of the hierarchy are satisfied to create flammable fuel conditions, site-specific factors (e.g., ignition frequency and fuel load) are thought to govern fire occurrence at the watershed scale. We assessed the relative role of climate, vegetation, and anthropogenic impact in driving regional biomass burning by (i) reconstructing the postglacial fire and vegetation history of the forest/steppe ecotone at seven sites, (ii) estimating regional trends in vegetation and fire activity so as to smooth site-specific variability that may reflect local controls, and (iii) visually comparing the estimated trends in biomass burning and vegetation with independent paleoclimate and archeological evidence from the region.
Deciphering the impact of hunter–gatherer groups on Patagonian fire regimes is not straightforward because pre-European use of fire is not well known. Present-day human societies alter the environment at global scales. Nonetheless, pre-European anthropogenic burning could have been negligible at the regional scale (i.e., >20,000 km2) but locally significant (i.e., watershed scale). We evaluated the role of people in altering fire activity at the watershed scale by (i) visually correlating local anomalies in fire activity with the local archeological record and (ii) comparing the probability of local fire anomalies before and after human populations established near each site.
Local anomalies were inferred from the residuals of the model used to estimate regional trends in biomass burning. The residuals of the model are expected to result from a combination of natural and statistical noise and site-specific departures from regional biomass burning. Because the threshold between noise and locally produced charcoal should occur in the right tail of the residual distribution (57), we focused on the positive residuals whose absolute value was larger than 1 SD of the residual population. Local departures from regional biomass burning were also compared with shrub taxa percentages at each site to assess the response of fire to changes in the local abundance of shrubs, which constitute an important source of fuel in present-day ecosystems (11).
Time series of the local departures from regional biomass burning at each site were divided in two periods defined by the time of human occupation of each watershed. The probability of local fire anomalies for both periods was calculated for all sites as
| [3] |
with s being the site (i.e., L. El Trébol, L. Padre Laguna, L. Huala Hué, L. Cóndor, L. Mosquito, L. La Zeta, and L. Theobald) and t being the time period (i.e., before human occupation and after human occupation). Differences in probability of fire in the pre- and postoccupation periods were tested for significance at a 0.05 alpha level by calculating a z value for proportions and adjusting the P values for multiple testing (58).
Field and Laboratory.
We reconstructed the fire and vegetation history of northern Patagonia from pollen and charcoal data from seven sites (21–461 ha in size with water depths of 488–1,050 cm) located at or near the forest/steppe boundary between the city of San Carlos de Bariloche in the north and Corcovado in the south [i.e., L. El Trébol (59), L. Padre Laguna (60), L. Huala Hué (60), L Cóndor (61), L. Mosquito (61), L. La Zeta (21, 62) and L. Theobald (21); Table S3 and Fig. 1]. Sediment cores were collected with a modified Livingstone piston sampler and extruded in the field. All necessary permits for coring were obtained from the Administración de Parques Nacionales (Argentine National Park Service, San Carlos de Bariloche, Argentina) and Estancia Leleque (Chubut, Argentina).
In the laboratory, cores were split longitudinally into a working half and an archive. The working half was described, photographed, and analyzed. The archival half and other materials are stored at the Paleoecology Laboratory at Montana State University.
We submitted bulk sediment, charcoal, and pollen samples to National Ocean Sciences Accelerator Mass Spectrometry Facility–Woods Hole Oceanographic Institution (Woods Hole, MA) and National Science Foundation–Arizona Accelerator Mass Spectrometry Facility (University of Arizona, Tucson, AZ) for radiocarbon analyses. Radiocarbon ages were calibrated using the Southern Hemisphere radiocarbon calibration dataset for samples <10,000 cal years B.P. and the Northern Hemisphere radiocarbon dataset for samples >10,000 cal years B.P. Chronologies were developed from modeling sediment age as a function of sediment depth (Table S4).
Sediment samples of 0.5 cm3 were taken at <250-y intervals for pollen analysis and prepared according to standard techniques (63). Pollen identification was based on a reference collection and published atlases (64, 65) and performed at 400× and 1,000× magnification. In all cases, counts exceeded 300 terrestrial pollen grains. Terrestrial pollen percentages were based on the sum of trees, shrubs, and herbs. Pollen data have been deposited in the Neotoma Paleoecology Database (www.neotomadb.org).
Fire histories were reconstructed from charcoal analysis performed on 2-cm3-volume samples taken at contiguous 0.5-cm intervals (66). The material was wet-screened through a 125-mm-mesh sieve, and charcoal particles were identified under a stereomicroscope. Particles within this relatively large-size class (i.e., macrocharcoal) are expected to be the product of fires near the lake (67) and are widely used as a proxy of local biomass burning (57, 66). Charcoal concentrations and deposition times were calculated and converted to CHARs (particles per cm2⋅y−1) so as to override the effects of changing sedimentation rates (57). Charcoal data have been deposited in the Global Charcoal Database (www.gpwg.org).
Estimation of Regional Trends in Vegetation and Fire.
Several approaches have been used to estimate regional trends in vegetation development and fire activity. One method involves visually comparing percentages of similar pollen types at each site, recognizing that they have different temporal resolutions (e.g., ref. 68). A second approach, used in global reconstructions of charcoal time series, is to standardize and interpolate stratigraphic data into evenly spaced time intervals and fit cubic splines in order obtain regional composite records (6, 69, 70).
In this study we construct regional composite records of vegetation and fire activity by fitting GAM to forest taxa, based on the ratio of forest pollen abundance to total terrestrial pollen, and CHAR time series. We focus on GAMs because they allow specification of the distribution of the response variable and the link function and do not assume linearity (71). The trends provided by the GAMs overcome the limitations of compounded single-site interpretations, do not require data interpolation or standardization, and account for autocorrelation, thus avoiding a common violation in regression modeling, namely assuming independent residuals.
Given that our response variables were based on counts (i.e., counts of forest pollen types × terrestrial pollen sum−1 and CHAR), Poisson rate models were applied to our count data (i.e., pollen grains per sample and number of charcoal particles) and assumed to be Poisson-distributed with means μi × Terrestrial pollen sumi−1 and μi × accumulation ratei. We used a logarithmic link between the means of the pollen count and charcoal data and time to ensure that fitted values were always nonnegative. Because overdispersion of the residuals was observed, we also assessed the performance of models that used a negative binomial distribution (Table S2).
In an attempt to achieve an optimal compromise between goodness of fit and parsimony, GAMs were fitted to the pollen and charcoal time series using penalized likelihood maximization, such that the model likelihood was modified by the addition of a penalty for each smooth function (71). All calculations and figures were made using R-programming language (72) with the package mgcv, Version 1.7–22 (73). By developing these models, we were able to highlight regional trends in vegetation composition and biomass burning, smooth site-specific variability that may reflect edaphic controls or local responses to disturbance, and overcome the limitations of sedimentary data.
We compared the models based on their AIC values (74). Following Burnham and Anderson (75), we considered models within two AIC units to have equal predictive ability. Although data exploration did not show clear linear patterns between initial Poisson models for either the pollen or charcoal data and time, we verified the assumption of nonlinearity by also comparing the performance of our best models to that of GLMs with similar specifications (Table S2).
Supplementary Material
Acknowledgments
We thank B. Gresswell, V. Markgraf, M. M. Bianchi, G. Villarosa, V. Outes, T. Kitzberger, and D. Navarro for participation in fieldwork; W. Browner, J. Giskaas, V. Nagashima, B. Ahearn, A. Peery, and C. Florentine who helped with laboratory analyses; and M. M. Bianchi who analyzed the pollen from L. Mosquito, L. El Trébol, and L. Huala Hué. This paper benefitted from the comments of M. Greenwood, B. Maxwell, K. Pierce, and K. Bennett.
Footnotes
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.
Data deposition: Pollen data have been deposited in Neotoma Paleoecology Database, www.neotomadb.org, and charcoal data have been deposited in the Global Charcoal Database, www.gpwg.org.
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1410443111/-/DCSupplemental.
References
- 1.Christensen NL. 1993. The effects of fire on nutrient cycles in longleaf pine ecosystems. The Longleaf Pine Ecosystem: Ecology, Restoration and Management, Proceedings of the 18th Tall Timbers Fire Ecology Conference (Tall Timbers Research Station, Tallahassee, FL), pp 205–214.
- 2.Heyerdahl EK, Morgan P, Riser JP., 2nd Multi-season climate synchronized historical fires in dry forests (1650-1900), northern Rockies, U.S.A. Ecology. 2008;89(3):705–716. doi: 10.1890/06-2047.1. [DOI] [PubMed] [Google Scholar]
- 3.Flannigan MD, Krawchuck MA, de Groot WJ, Wotton BM, Gowman LM. Implications of changing climate for global wildland fire. Int J Wildland Fire. 2009;18:483–507. [Google Scholar]
- 4.Bowman DMJS, et al. Fire in the Earth system. Science. 2009;324(5926):481–484. doi: 10.1126/science.1163886. [DOI] [PubMed] [Google Scholar]
- 5.Whitlock C, Higuera PE, McWethy DM, Briles CE. Paleoperspectives on fire ecology: Revisiting the fire regime concept. Open Ecol J. 2010;3:6–23. [Google Scholar]
- 6.Ali AA, et al. Control of the multimillennial wildfire size in boreal North America by spring climatic conditions. Proc Natl Acad Sci USA. 2012;109(51):20966–20970. doi: 10.1073/pnas.1203467109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Marlon J, et al. Global biomass burning: A synthesis and review of Holocene paleofire records and their controls. Quat Sci Rev. 2013;65:5–25. [Google Scholar]
- 8.Williams M. Deforesting the Earth: From Prehistory to Global Crisis. Univ of Chicago Press; Chicago: 2006. [Google Scholar]
- 9.Kitzberger T. Ecotones as complex arenas of disturbance, climate and human impacts: The trans-Andean forest-steppe ecotone of northern Patagonia. In: Myster R, editor. Ecotones Between Forest and Grassland. Springer; New York: 2012. pp. 59–88. [Google Scholar]
- 10.Garreaud RD, Vuille M, Compagnucci R, Marengo J. Present-day South American climate. Palaeogeogr Palaeoecol. 2008;281(3-4):229–241. [Google Scholar]
- 11.Mermoz M, Kitzberger T, Veblen TT. Landscape influences on occurrence and spread of wildfires in Patagonian forests and shrublands. Ecology. 2005;86(10):2705–2715. [Google Scholar]
- 12.Veblen TT, et al. Adapting to global environmental change in Patagonia: What role for disturbance ecology? Austral Ecol. 2011;36(8):891–903. [Google Scholar]
- 13.Gowda JH, Kitzberger T, Premoli AC. Landscape responses to a century of land use along the northern Patagonian forest-steppe transition. Plant Ecol. 2012;213(2):259–272. [Google Scholar]
- 14.Villalba R, et al. Large-scale temperature changes across the southern Andes: 20th-century variations in the context of the past 400 years. Clim Change. 2003;59:177–232. [Google Scholar]
- 15.Compagnucci R, Araneo D. Alcances de El Niño como predictor de el caudal de los ríos andinos argentinos. Ing Hidrául Méx. 2007;22(3):23–35. [Google Scholar]
- 16.Miller A. The climate of Chile. In: Schwerdtfeger W, editor. Climates of Central and South America. Elsevier; Amsterdam: 1976. pp. 113–145. [Google Scholar]
- 17.Mazzarino MJ, Bertiller M, Gobbi M. Nutrient cycling in Patagonian ecosystems. Ecología Austral. 1998;8:167–181. [Google Scholar]
- 18.Veblen TT, Kitzberger T, Villalba R, Donnegan J. Fire history in northern Patagonia: The roles of humans and climatic variation. Ecol Monogr. 1999;69(1):47–67. [Google Scholar]
- 19.Kitzberger T, Veblen TT, Villalba R. Climatic influences on fire regimes along a rainforest-to-xeric woodland gradient in northern Patagonia, Argentina. J Biogeogr. 1997;24:35–47. [Google Scholar]
- 20.Holz A. Climatic and Human Influences on Fire Regimes and Forest Dynamics in Temperate Rainforests in Southern Chile. University of Colorado; Boulder, CO: 2009. [Google Scholar]
- 21.Iglesias V, Whitlock C, Markgraf V, Bianchi MM. Postglacial history of the Patagonian forest/steppe ecotone (41–43°S) Quat Sci Rev. 2014;94:120–135. [Google Scholar]
- 22.Páez MM, Schaebitz F, Stutz S. Modern pollen-vegetation and isopoll maps in southern Argentina. J Biogeogr. 2001;28:997–1021. [Google Scholar]
- 23.Markgraf V, D’Antoni HL, Ager TA. Modern pollen dispersal in Argentina. Palynology. 1981;5:43–63. [Google Scholar]
- 24.Fernández PM, Calatayud M, Bellelli C, Pedestá M, Scheinsohn V. 2011. Marcas en la piedra, huellas en la tierra. El Poblamiento del bosque del suroeste de Río Negro-noroeste de Chubut [Human settling in the forests of southwestern Río Negro-northwestern Chubut]. Procesos Históricos, Transformaciones Sociales y Construcciones de Fronteras, eds Valverde S, Maragliano G, Impemba M, Trentini F (Facultad de Filosofía y Letras, Universidad de Buenos Aires, Buenos Aires), pp 195–221.
- 25.Porter SC. Onset of neoglaciation in the southern hemisphere. J Quaternary Sci. 2000;15(4):395–408. [Google Scholar]
- 26.Mayewski PA, et al. Holocene climate variability. Quat Res. 2004;62:243–255. [Google Scholar]
- 27.Douglass DC, et al. Evidence of early Holocene glacial advances in southern South America from cosmogenic surface-exposure dating. Geology. 2005;33:237–240. [Google Scholar]
- 28.Grootes PM, Steig EO, Stuiver M, Waddington ED, Morse DL. GISP2_Taylor Dome oxygen isotope ratios. Quat Res. 1999;283:1712–1714. [Google Scholar]
- 29.Lamy F, Hebbeln D, Rohl U, Wefer G. Holocene rainfall variability in southern Chile: A marine record of latitudinal shifts of the southern westerlies. Earth Planet Sci Lett. 2001;185:369–382. [Google Scholar]
- 30.Moy CM, Seltzer GO, Rodbell DT, Anderson DM. Variability of El Niño/Southern Oscillation activity at millennial timescales during the Holocene epoch. Nature. 2002;420(6912):162–165. doi: 10.1038/nature01194. [DOI] [PubMed] [Google Scholar]
- 31.Hirota M, Holmgren M, Van Nes EH, Scheffer M. Global resilience of tropical forest and savanna to critical transitions. Science. 2011;334(6053):232–235. doi: 10.1126/science.1210657. [DOI] [PubMed] [Google Scholar]
- 32.Bond WJ. What limits trees in C4 grasslands and savannas? Annu Rev Ecol Evol Syst. 2008;39:641–659. [Google Scholar]
- 33.Clark JS, Royall PD, Chumbley C. The role of fire during climate change in an eastern deciduous forest at Devil’s Bathtub, New York. Ecology. 1996;77(7):2148–2166. [Google Scholar]
- 34.Heusser CJ. Some pollen profiles from the Laguna de San Rafael area, Chile. In: Cranwell LM, editor. Ancient Pacific Floras. Pacific Press; Honolulu: 1964. pp. 34–56. [Google Scholar]
- 35.Moreno PI. Millennial-scale climate variability in northwest Patagonia over the last 15,000 yr. J Quaternary Sci. 2004;19(1):35–47. [Google Scholar]
- 36.Markgraf V, Whitlock C, Haberle S. Vegetation and fire history during the last 18,000 cal yr BP in Southern Patagonia: Mallín Pollux, Coyhaique, Province Aisén (45 41’30’’ S, 71 50’30’’ W, 840 m elevation) Palaeogeogr Palaeoecol. 2007;257(3):492–507. [Google Scholar]
- 37.Berger A, Loutre MF. Insolation values for the climate of the last 10 million years. Quat Sci Rev. 1991;10(4):297–317. [Google Scholar]
- 38.Whitlock C, Moreno P, Bartlein PJ. Climatic controls of Holocene fire patterns in southern South America. Quat Res. 2007;68(1):28–36. [Google Scholar]
- 39.Kitzberger T, Swetnam TW, Veblen TT. Inter-hemispheric synchrony of forest fires and the El Niño-Southern Oscillation. Glob Ecol Biogeogr. 2001;10(3):315–326. [Google Scholar]
- 40.Musters GC. At Home with The Patagonians: A Year’s Wanderings over Untrodden Ground from the Straits of Magellan to the Río Negro. Murray; London: 1871. [Google Scholar]
- 41.Cox G. Viajes a las regiones septentrionales de Patagonia 1862-1863. Anal Univ Chile. 1963;23:3–239. [Google Scholar]
- 42.Bárbena R, et al. Pleistocene-Holocene transition in northern Patagonia: Evidence from Huenul Cave (Neuquén, Argentina) Curr Res Pleist. 2010;27:4–7. [Google Scholar]
- 43.Hajduk A, Albornoz A, Lezcano M. Levels with extinct fauna in the forest rockshelter El Trébol (North Patagonia, Argentina) Curr Res Pleist. 2006;23:55–57. [Google Scholar]
- 44.Bellelli C, Carballido M, Fernández P, Scheinsohn V. El pasado entre las hojas. Nueva información arqueologica del noroeste de la provincia de Chubut, Argentina. Rev Werken. 2003;4:25–42. [Google Scholar]
- 45.Bellelli C. Arqueología, patrimonio y turismo. Experiencias de investigación, conservacion, manejo y gestión en la Comarca Andina del Paralelo 42° y valle del Manso inferior. Pacarina. 2007;45:309–314. [Google Scholar]
- 46.Podestá M, et al. 2007. Arqueología del valle del río Epuyén (El Hoyo, Chubut, Patagonia argentina). Levantando piedras, desenterrando huesos... y develando arcanos [Archeology in the Epuyén river valley (El Hoyo, Chubut, Argentine Patagonia)]. Arqueología de Fuego-Patagonia, ed Centro de Estudios del Cuaternario de Fuego-Patagonia y Antártica (Universidad de Magallanes, Punta Arenas, Chile), pp 427–442.
- 47.Méndez CA, Reyes OR. Nuevos datos de la ocupación humana en la transición bosque estepa en Patagonia: Alero Las Quemas (Comuna de LagoVerde, XI Región de Aisén) Magallania. 2006;34(1):161–165. [Google Scholar]
- 48.Méndez CA, Reyes OR. Late Holocene human occupation of the Patagonian forests: A case study in the Cisnes river basin. Antiquity. 2008;82:560–570. [Google Scholar]
- 49.McWethy DB, et al. Rapid landscape transformation in South Island, New Zealand, following initial Polynesian settlement. Proc Natl Acad Sci USA. 2010;107(50):21343–21348. doi: 10.1073/pnas.1011801107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Cronon W. Changes in the Land: Indians, Colonists, and the Ecology of New England. Hill and Wang; New York: 1983. [Google Scholar]
- 51.Kitzberger T. In: Impact of Extreme and Infrequent Events on Terrestrial Ecosystems and Biodiversity. Encyclopedia of Biodiversity. Levin S, editor. Elsevier; Amsterdam: 2013. [Google Scholar]
- 52.Turner MG, Gardner RH, O’Neill RV. Landscape Ecology in Theory and Practice. Springer; New York: 2001. [Google Scholar]
- 53.Nielson RP. Climatic constraints and issues of scale controlling regional biomes. In: Holl MM, Risser PG, Naiman RJ, editors. Ecotones. The Role of Landscape Boundaries in the Management and Restoration of Changing Environments. Chapman and Hall; New York: 1991. pp. 31–51. [Google Scholar]
- 54.Gavin DG, Brubaker LB, Lertzman KP. Holocene fire history of a costal temperate rain forest based on soil charcoal radiocarbon dates. Ecology. 2003;84(1):186–201. [Google Scholar]
- 55.Parisien M-A, Moritz MA. Environmental controls on the distribution of wildfire at multiple spatial scales. Ecol Monogr. 2009;79(1):127–154. [Google Scholar]
- 56.Pechony O, Shindell DT. Driving forces of global wildfires over the past millennium and the forthcoming century. Proc Natl Acad Sci USA. 2010;107(45):19167–19170. doi: 10.1073/pnas.1003669107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Higuera PE, Brubaker LB, Anderson PM, Hu FS, Brown TA. Vegetation mediated the impacts of postglacial climatic change on fire regimes in the south-central Brooks Range, Alaska. Ecol Monogr. 2009;79(2):201–219. [Google Scholar]
- 58.Benjamini Y, Hochberg Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J Roy Stat Soc B Met. 1995;57(1):289–300. [Google Scholar]
- 59.Whitlock C, et al. Postglacial vegetation, climate, and fire history along the east side of the Andes (lat 41-42.5°S), Argentina. Quat Res. 2006;66(2):187–201. [Google Scholar]
- 60.Iglesias V, Whitlock C, Bianchi MM, Villarosa G, Outes V. Climate and local controls of long-term vegetation dynamics in northern Patagonia (lat 41°S) Quat Res. 2012;78(3):502–512. [Google Scholar]
- 61.Iglesias V, Whitlock C, Bianchi MM, Villarosa G, Outes V. Holocene climate variability and environmental history at the Patagonian forest/steppe ecotone: Lago Mosquito (42°29’3.89”S, 71°24’14.57”W) and Laguna del Cóndor (42°20’47.22”S, 71°17’07.62”W) Holocene. 2012;22:1254–1264. [Google Scholar]
- 62.Schaebitz F. Vegetation development and volcanism in the Esquel region, Chubut, Argentina. Quaternary S Am Ant Penin. 1994;10:7–29. [Google Scholar]
- 63.Faegri K, Kaland PE, Krzywinski K. Textbook of Pollen Analysis. Wiley J and Sons; New York: 1989. [Google Scholar]
- 64.Heusser CJ. Pollen and Spores of Chile. Modern Types of the Pteridophyta, Gymnospermae, and Angiospermae. Univ of Arizona Press; Tucson: 1971. [Google Scholar]
- 65.Markgraf V, D’Antoni HL. Pollen Flora of Argentina. Univ. of Arizona Press; Tucson: 1978. [Google Scholar]
- 66.Whitlock C, Larsen CPS. 2001. Terrestrial, algal, and siliceous indicators. Charcoal as a Fire Proxy, eds Smol JP, Birks HJP, Last WM, Tracking Environmental Change Using Lake Sediments (Kluwer Academic Publishers, Dordrecht, The Netherlands), Vol 3, pp 75–97.
- 67.Clark J, Royall PD. Particle size evidence for source areas of charcoal accumulation in late Holocene sediments of eastern North American lakes. Quat Res. 1995;43(1):80–89. [Google Scholar]
- 68.Zhao Y, Zicheng Y, Chen F, Zhang J, Yang B. Vegetation response to Holocene climate change in monsoon-influenced region of China. Earth Sci Rev. 2009;97:242–256. [Google Scholar]
- 69.Marlon JR, et al. Climate and human influences on global biomass burning over the past two millennia. Nat Geosci. 2008;1(10):697–702. [Google Scholar]
- 70.Daniau A-L, et al. Predictability of biomass burning in response to climate changes. Global Biogeochem Cycles. 2012;26(4):12–24. [Google Scholar]
- 71.Wood SN. Stable and efficient multiple smoothing parameter estimation for generalized additive models. J Am Stat Assoc. 2004;99(467):673–686. [Google Scholar]
- 72.R Core Team . R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing; Vienna: 2012. [Google Scholar]
- 73.Wood SN. Fast stable restricted maximum likelihood and marginal likelihood estimation of semiparametric generalized linear models. JR Stat Soc. 2011;73(1):3–36. [Google Scholar]
- 74.Akaike H. Information theory as an extension of the maximum likelihood principle. In: Petrov BN, Csaki F, editors. Second International Symposium on Information Theory. Akademiai Kiado; Budapest: 1973. pp. 267–281. [Google Scholar]
- 75.Burnham KP, Anderson DR. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach. Springer; New York: 2002. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.