

PAX5 encodes for a transcription factor essential for B-lymphoid cell commitment and is rearranged in 30% of B-cell precursor acute lymphoblastic leukemia pediatric patients (BCP-ALL).1 Since the first characterization of PAX5/ETV6,2 an increasing number of PAX5 fusion genes have been described in both adult and pediatric BCP-ALL patients. As reported in our recent review,3 PAX5 fusion partners encode a variety of proteins involved in cell structure (ELN,4 POM1215), transcriptional regulation (ETV6,1,2,6,7 PML,5 FOXP1,1,7 MLLT38), phosphorylation (JAK25), and unknown functions (C20orf112,7 AUTS29).
Here we report the characterization of three new PAX5 partner genes in BCP-ALL: CHFR (a novel transcript isoform), SOX5 and POM121C. Moreover, we report cases displaying PAX5/AUTS2 and PAX5/MLLT3 fusions.8,9
Among 237 BCP-ALL pediatric patients enrolled in the AIEOP-BFM ALL2000 protocol for whom standard cytogenetic karyotyping was successfully performed, 35 (14.7%) presented visible abnormalities on the 9p potentially involving PAX5, and 19 of 35 had material available to be considered for this study. An additional adult BCP-ALL patient (Patient 20), carrying both a 9p abnormality in proximity of the PAX5 region and a t(9;22) translocation, was also included. Patients’ clinical and cytogenetic features can be found in the Online Supplementary Table S1.
A detailed fluorescence in situ hybridization (FISH) analysis10 using selected BAC and fosmid probes (Online Supplementary Appendix) identified a large deletion affecting PAX5 in 14 patients (Online Supplementary Table S2 and Figure S1) and, more importantly, a translocation involving PAX5 in 5 cases (Patients 5, 7, 10, 15 and 19) (Figure 1A and Online Supplementary Table S3).
Figure 1.
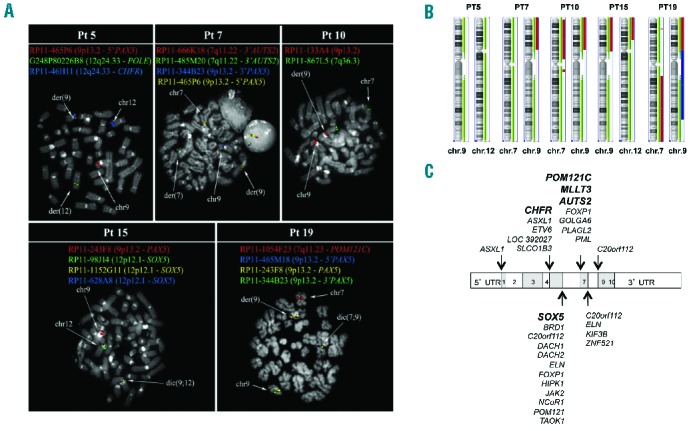
PAX5 translocated patients. (A) FISH analysis of PAX5 translocations. BAC probes, shown on the top of each panel, are colored according to their corresponding signals in the merged image. (B) Karyoview of chromosomes involved in the translocation event in each patient. The Chromosome Analysis Suite software was used (Affymetrix, Santa Clara, CA, USA). We considered as relevant gains and losses involving a minimum number of 20 markers and a minimal size of 100 Kb and 50 Kb, respectively. Red and Blue colors correspond to loss and gain of chromosome regions, respectively. Light and dark green are indicators of markers coverage (both for Copy Number and SNP probes). (C) Map of the PAX5 breakpoints in translocations generating in-frame or out-of-frame fusions, or truncated isoforms.3 The names of the partner genes described in this manuscript are in bold.
To determine whether these PAX5 alterations might be involved in leukemogenesis as driving genetic lesions, we further characterized PAX5-fused cases (Patients 5, 7, 10, 15, 19 and 20), as well as their corresponding remissions using copy number variation (CNV) analysis, by Affymetrix Cytogenetics Whole Genome Array. Data are available in the ArrayExpress database (available from: www.ebi.ac.uk/arrayexpress, accession number E-MTAB-2416).
A characterization of each novel fusion gene is given below and the expected fusion proteins are described in Figures 2 and 3.
Figure 2.
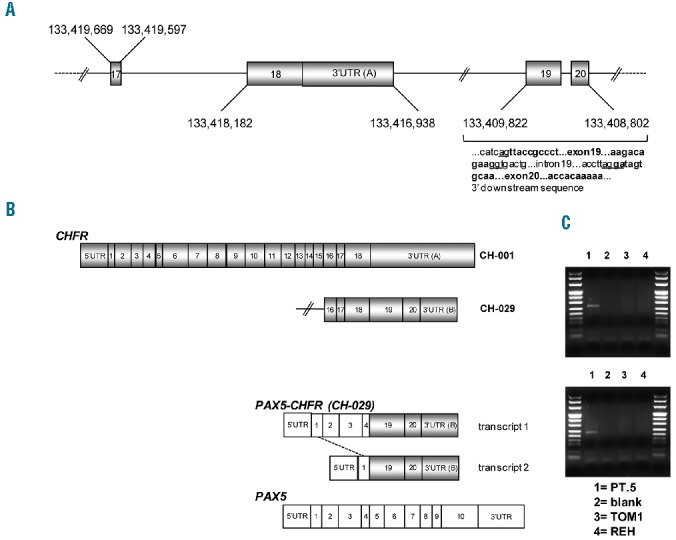
PAX5/CHFR fusion transcript and novel CHFR RNA isoform. (A) Partial genomic organization of CHFR on chromosome 12, and sequence of the newly identified exons (in bold), according to GRCh37/hasg19. Splicing donor/acceptor sites are indicated as underlined. (B) Schematic representation of CH-001 and the novel CH-029 CHFR transcript isoforms. In addition, a schematic representation of wild-type PAX5 and of the two alternative PAX5/CHFR fusion transcripts is reported. (C) PAX5-CH-029 transcript 1 (upper right panel) and transcript 2 (lower right panel) are exclusively expressed in Patient 5 (lane 1) and not in control BCP-ALL cell lines (TOM1, lane 3; REH, lane 4). Lane 2: no template control.
Figure 3.
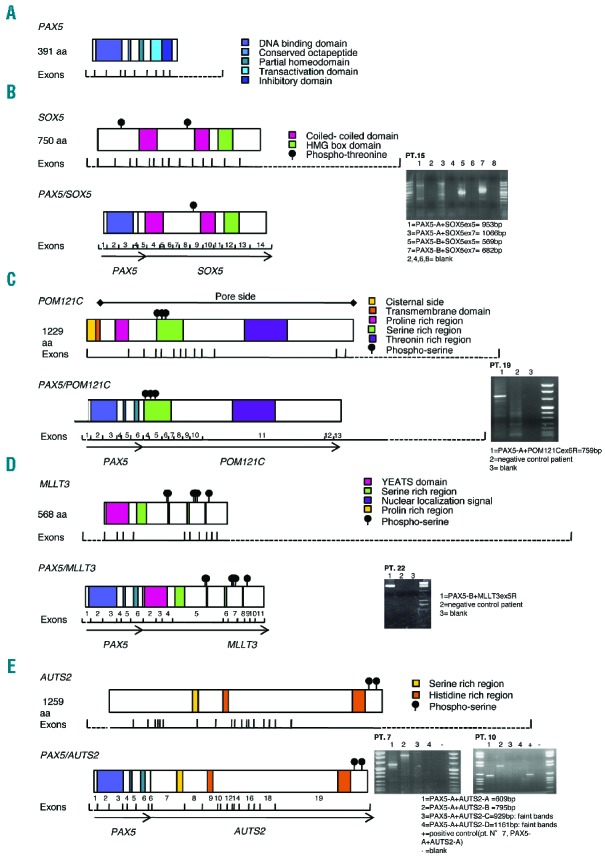
Schematic representation of novel PAX5 fusion proteins and mRNA expression levels. For each novel fusion gene (B–E), the RT-PCR gel electrophoresis (the specific primers reported in the Online Supplementary Table S4 are indicated) and the scheme of the predicted fusion protein are represented on the right and on the left, respectively. The conserved functional domains of the wild-type protein counterparts and exon numbers of fused transcripts are also indicated. (A) Normal PAX5. (B) PAX5/SOX5 in Patient 15. (C) PAX5/POM121C in Patient 19. (D) PAX5/MLLT3 in Patient 20. (E) PAX5/AUTS2 in Patients 7 and 10.
Cytogenetic analysis of Patient 5 revealed a t(9;12) translocation (Figure 1A), with breakpoints mapping respectively within PAX5 and POLE genes. By 5′ RACE-and RT-PCR, PAX5 was found as joined to a novel CHFR (Checkpoint with fork-head associated and ring finger) transcript isoform (12q24.33), herein indicated as CH-029 (Figure 2). Since CHFR is located 200 Kb distally to POLE (Online Supplementary Figure S2A) and shows an opposite transcriptional orientation to PAX5, we speculate the occurrence of a complex genomic rearrangement leading to an in-frame PAX5/CHFR fusion transcript (Online Supplementary Figure S2F). The newly identified CHFR coding sequence contained two additional exons, herein named as exons 19 and 20, and overlapping with previously mapped ESTs (GRCh37/hg19) located after the 3′ UTR of the CH-001 isoform (Figure 2A). The two transcript isoforms (CH-001 and CH-029) used alternative 3′UTR, defined as 3′UTR (A) and (B), respectively (Figure 2B). Two alternative fusion transcripts, joining CHFR exons 19–20 to either PAX5 exons 1–4 (transcript 1, 952nt) or exon 1 (transcript 2, 522nt) were identified (Figure 2B). As expected, both PAX5/CHFR transcripts were exclusively expressed in Patient 5 (Figure 2C). Conversely, the CH-029 was expressed in Patient 5 as well as in several cell lines (Online Supplementary Figure S2B-D). Unfortunately, any effort to amplify the reciprocal CHFR/PAX5 transcript was unsuccessful (data not shown). Both CH-001 and CH-029 transcripts were broadly expressed in non-tumor samples by RT-qPCR, with the highest expression found in normal peripheral blood (Online Supplementary Figure S2E).
Copy number variation (CNV) analysis of Patient 5 detected dispersed copy number neutral loss of heterozygosity (LOH) as well as genome losses, although not flanking the t(9;12) translocation breakpoints (Figure 1B and Online Supplementary Table S5).
In Patient 15, FISH analysis identified SOX5 [SRY (sex determining region y)-Box 5] (12p12.1) as fused to PAX5 on a dic(9;12)(p13;p12.1) dicentric chromosome (Figure 1A), being accompanied by losses at breakpoint regions, as confirmed by CNV analysis (Figure 1B and Online Supplementary Table S5). RACE-PCR defined the junction as located between PAX5 exon 5 and SOX5 exon 4, confirmed by RT-PCR (Figure 3B). Since SOX5 and PAX5 are in oppo site transcriptional orientation, we hypothesize that such a fusion is the result of a complex genomic event, similarly to that described for Patient 5 (see above). The occurrence of an intragenic deletion of SOX5, verified by FISH and SNP array, strongly supports this view. SOX5 encodes for a transcription factor involved in neuron development through the specific repression of target genes in the central nervous system (CNS). Intriguingly, this PAX5/SOX5 patient experienced a CNS relapse. Thus, it can be hypothesized that the chimeric protein might act as an aberrant transcription factor affecting both B-cell (through PAX5 pathway alteration) and CNS features (by SOX5 pathway alteration). Interestingly, SOX5 has already been found fused to P2RY8 in primary splenic follicular lymphoma.11
In Patient 19, CNV analysis detected deletions at 9p13.2 and 7q11.23, involving PAX5 and POM121C (POM121 transmembrane nucleoporin C), respectively (Figure 1B and Online Supplementary Table S5). RNA-sequencing identified reads with PAX5 exon 6 fused to POM121C exon 4 (Online Supplementary Figure S3), verified by RT-PCR (Figure 3C). FISH analysis confirmed the occurrence of the two copies of the fusion gene, respectively on a dicentric chromosome dic(7;9)(q11.23;p13.2) and on a second chromosome that might be the result of a further rearrangement of a second copy of the same dic(7;9) (Figure 1A and Online Supplementary Figure S4). Notably, POM121C is a member of the same nucleoporin family as POM121C, already identified as a fusion partner of PAX5.5 The adult B-ALL patient (Patient 20) carried a t(9;22)(q34;q11) translocation together with an additional large deletion at 9p (Online Supplementary Table S1). CNV analysis detected a 16 Mb loss between PAX5 at 9p13 and MLLT3 at 9p21.3 (Online Supplementary Figure S5A). RT-PCR identified a fusion transcript joining PAX5 exon 6 to MLLT3 exon 2 (Online Supplementary Figure S5B). MLLT3 is also known as AF9, which is one of the most recurrent MLL partner genes in ALL and AML. CNV analyses highlighted several genetic lesions, although most of them were located in regions lacking coding genes or sno/miRNAs (Online Supplementary Figure S5C). This fusion gene has already been described in a B-ALL case,8 but the patient’s age, as well as the association with the t(9;22) translocation, makes the genomic profile of this patient unique when compared to the other PAX5-translocated cases.
Two additional cases, Patients 7 and 10, were both characterized by the presence of a t(7;9) translocation (Online Supplementary Table S1). In both patients, FISH analyses showed a deletion of the 3′ portion of PAX5. The retained segment of RP11-465P6 (5′PAX5) on der(9) was found to co-localize with the splitting signals of both BAC probes specific for the 3′ portion of AUTS2 (RP11-666K18, and RP11-485M20) on der(9) (Figure 1A) indicating the presence of PAX5/AUTS2 fusion gene in both cases, further confirmed by RACE- and RT-PCR (Figure 3E). CNV analysis showed the loss of the 9p with proximal breakpoint within PAX5, and loss of chromosome 7p starting from the AUTS2 gene locus (Figure 1B and Online Supplementary Table S5). PAX5/AUTS2 has been previously reported in BCP-ALL.9
As described in the Online Supplementary Appendix, by whole genome array analysis, we detected specific IG/TCR somatic rearrangements as a marker of the clonal origin of the leukemic cells. Interestingly, besides the translocation events, very few additional lesions were identified, mainly losses (Online Supplementary Table S5), as described for adult PAX5-translocated cases.12
In detail, 3 patients (Patients 7, 10 and 15) out of 5 had only one alteration in addition to the translocation, and 2 out of the 3 CNV events affected a region without known coding genes/miRNAs (UCSC database, GRCh37/hg19). Patient 5 showed 4 alterations, which were linked to two main events, such as LOH of chromosome 5 and mosaicism of chromosome 7. In particular, FISH analysis indicated the presence of two additional clones carrying either loss or gain of chromosome 7 (30% and 20%, respectively) (data not shown).
Patient 19 carried five deletions (3 on chromosome 2). Interestingly, despite the loss of most of the 9p due to translocation, the remaining p arm and most of the q arm (on 9q32, till ZFP37 included) were amplified (Copy Number, CN=3), suggesting that even the fusion gene might be duplicated, as shown by FISH (Figure 1A and Online Supplementary Figure S4).
Intriguingly, our mean value of 2.4 CNVs per patient (Online Supplementary Table S5) is significantly lower than the 4–8 lesions per patient previously reported in childhood BCP-ALL.13 Although the potential presence of point mutations cannot be ruled out, the low number of co-operative genetic lesions would suggest that the PAX5-translocation event itself might play an important role in leukemogenesis.
In all the PAX5 fusions, the breakpoints in PAX5 clustered in two hot-spot regions mapping after exons 5 and 7, respectively, which retained the PAX5 DNA binding domain (Figure 1C). In agreement with previous findings, none of our cases expressed the reciprocal fusion transcript.5
The high heterogeneity of PAX5 fusion genes makes it difficult to understand the contribution of each single partner gene to leukemogenesis and the identification of shared features, except for the disruption of the physiological function of PAX5.3 Although recent studies have indicated a functional role of PAX5 chimeric proteins,14,15 their exact role in leukemogenesis still has to be fully elucidated.
Footnotes
Funding: This work was partially supported by: Associazione Italiana Ricerca sul Cancro (AIRC: IG2009 No.8666 to AB, MFAG2011 No.11405 to CTS; and IG2012 No.13574 to GC) and Federazione Italiana Ricerca sul Cancro (FIRC), Fondazione Cariplo, Progetto InterBioMed Regione Lombardia, MIUR, UNIMIB.
The online version of this article has a Supplementary Appendix.
Information on authorship, contributions, and financial & other disclosures was provided by the authors and is available with the online version of this article at www.haematologica.org.
References
- 1.Mullighan CG, Goorha S, Radtke I, Miller CB, Coustan-Smith E, Dalton JD, et al. Genome-wide analysis of genetic alterations in acute lymphoblastic leukaemia. Nature. 2007;446(7137):758–64. [DOI] [PubMed] [Google Scholar]
- 2.Cazzaniga G, Daniotti M, Tosi S, Giudici G, Aloisi A, Pogliani E, et al. The paired box domain gene PAX5 is fused to ETV6/TEL in an acute lymphoblastic leukemia case. Cancer Res. 2001;61(12): 4666–70. [PubMed] [Google Scholar]
- 3.Fazio G, Biondi A, Cazzaniga G. The role of PAX5 in ALL. 1st ed. Rijeka: InTech Open Access Publisher, 2011. [Google Scholar]
- 4.Bousquet M, Broccardo C, Quelen C, Meggetto F, Kuhlein E, Delsol G, et al. A novel PAX5-ELN fusion protein identified in B-cell acute lymphoblastic leukemia acts as a dominant negative on the wild type PAX5. Blood. 2007;109(8):3417–23. [DOI] [PubMed] [Google Scholar]
- 5.Nebral K, Denk D, Attarbaschi A, Konig M, Mann G, Haas OA, et al. Incidence and diversity of PAX5 fusion genes in childhood acute lymphoblastic leukemia. Leukemia. 2009;23(1):134–43. [DOI] [PubMed] [Google Scholar]
- 6.An Q, Wright SL, Konn ZJ, Matheson E, Minto L, Moorman AV, et al. Variable breakpoints target PAX5 in patients with dicentric chromosomes: a model for the basis of unbalanced translocations in cancer. Proc Natl Acad Sci U S A. 2008;105(44):17050–4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kawamata N, Ogawa S, Zimmermann M, Kato M, Sanada M, Hemminki K, et al. Molecular allelokaryotyping of pediatric acute lymphoblastic leukemias by high-resolution single nucleotide polymorphism oligonucleotide genomic microarray. Blood. 2008; 111(2):776–84. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Lee ST, Ji Y, Kim HJ, Ki CS, Jung CW, Kim JW, et al. Sequential array comparative genomic hybridization analysis identifies copy number changes during blastic transformation of chronic myeloid leukemia. Leuk Res. 2012;36(4):418–21. [DOI] [PubMed] [Google Scholar]
- 9.Coyaud E, Struski S, Dastugue N, Brousset P, Broccardo C, Bradtke J. PAX5-AUTS2 fusion resulting from t(7;9)(q11.2;p13.2) can now be classified as recurrent in B cell acute lymphoblastic leukemia. Leuk Res. 2010;34(12):e323–5. [DOI] [PubMed] [Google Scholar]
- 10.Storlazzi CT, Albano F, Dencic-Fekete M, Djordjevic V, Rocchi M. Late-appearing pseudocentric fission event during chronic myeloid leukemia progression. Cancer Genet Cytogenet. 2007;174(1):61–7. [DOI] [PubMed] [Google Scholar]
- 11.Storlazzi CT, Albano F, Lo Cunsolo C, Doglioni C, Guastadisegni MC, Impera L, et al. Upregulation of the SOX5 by promoter swapping with the P2RY8 gene in primary splenic follicular lymphoma. Leukemia. 2007;21(10):2221–5. [DOI] [PubMed] [Google Scholar]
- 12.Familiades J, Bousquet M, Lafage-Pochitaloff M, Bene MC, Beldjord K, De Vos J, et al. PAX5 mutations occur frequently in adult B-cell progenitor acute lymphoblastic leukemia and PAX5 haploinsufficiency is associated with BCR-ABL1 and TCF3-PBX1 fusion genes: a GRAALL study. Leukemia. 2009;23(11):1989–98. [DOI] [PubMed] [Google Scholar]
- 13.Bardini M, Spinelli R, Bungaro S, Mangano E, Corral L, Cifola I, et al. DNA copy-number abnormalities do not occur in infant ALL with t(4;11)/MLL-AF4. Leukemia. 2010;24(1):169–76. [DOI] [PubMed] [Google Scholar]
- 14.Fazio G, Cazzaniga V, Palmi C, Galbiati M, Giordan M, te Kronnie G, et al. PAX5/ETV6 alters the gene expression profile of precursor B cells with opposite dominant effect on endogenous PAX5. Leukemia. 2013;27(4):992–5. [DOI] [PubMed] [Google Scholar]
- 15.Fortschegger K, Anderl S, Denk D, Strehl S. Functional Heterogeneity of PAX5 Chimeras Reveals Insight for Leukemia Development. Mol Cancer Res. 2014;12(4):595–606. [DOI] [PubMed] [Google Scholar]