

Abstract
The evolutionary trajectories of reproductive systems, including both male and female multiple mating and hence polygyny and polyandry, are expected to depend on the additive genetic variances and covariances in and among components of male reproductive success achieved through different reproductive tactics. However, genetic covariances among key components of male reproductive success have not been estimated in wild populations. We used comprehensive paternity data from socially monogamous but genetically polygynandrous song sparrows (Melospiza melodia) to estimate additive genetic variance and covariance in the total number of offspring a male sired per year outside his social pairings (i.e. his total extra-pair reproductive success achieved through multiple mating) and his liability to sire offspring produced by his socially paired female (i.e. his success in defending within-pair paternity). Both components of male fitness showed nonzero additive genetic variance, and the estimated genetic covariance was positive, implying that males with high additive genetic value for extra-pair reproduction also have high additive genetic propensity to sire their socially paired female's offspring. There was consequently no evidence of a genetic or phenotypic trade-off between male within-pair paternity success and extra-pair reproductive success. Such positive genetic covariance might be expected to facilitate ongoing evolution of polygyny and could also shape the ongoing evolution of polyandry through indirect selection.
Keywords: life-history trade-off, multiple mating, paternity success, polyandry, polygyny, quantitative genetics, reproductive strategy, sexual selection
Introduction
In general, the evolutionary dynamics of reproductive systems and associated traits are expected to be shaped by negative and positive genetic covariances among the life-history components that define the reproductive system and hence by genetic trade-offs and synergies acting within and across the sexes (Lande, 1982; Roff, 2002). Such genetic covariances, rather than solely phenotypic covariances, therefore need to be quantified in order to understand and predict ongoing evolution (Stearns, 1989; Roff, 2002; Kruuk et al., 2008; Robinson & Beckerman, 2013).
One specific ambition is to understand the evolution and persistence of complex reproductive systems where males and females mate with multiple opposite-sex individuals within single reproductive episodes (termed polygyny and polyandry, respectively, Pizzari & Birkhead, 2002; Parker, 2006; Evans & Simmons, 2008; Slatyer et al., 2012). Polyandry has proved difficult to explain, particularly in situations where direct selection on female multiple mating appears likely to be negative (i.e. a negative female Bateman gradient beyond a single mating, Jennions & Petrie, 2000; Slatyer et al., 2012; Parker & Birkhead, 2013). One potential explanation is that polyandry is positively genetically correlated with components of male fitness and consequently evolves or is maintained through indirect selection (Halliday & Arnold, 1987; Keller & Reeve, 1995; Evans & Simmons, 2008; Forstmeier et al., 2011). In contrast, polygyny, which often occurs alongside polyandry, often seems easy to explain because multiple mating is widely expected to increase a male's total reproductive success and hence to experience positive direct selection (i.e. a positive male Bateman gradient, Halliday & Arnold, 1987; Parker, 2006; Forstmeier et al., 2011; Kvarnemo & Simmons, 2013). However, it is not always emphasized that these explanations for the ongoing evolution and persistence of polygyny and polyandry both depend critically on the direction and magnitude of genetic covariances among different and potentially conflicting components of male fitness (Halliday & Arnold, 1987; Evans, 2010; Kvarnemo & Simmons, 2013).
For example, one widespread but evolutionarily puzzling polygynandrous reproductive system is social monogamy with extra-pair reproduction. Here, some offspring are sired by extra-pair males rather than by a female's socially paired male, whereas the female's socially paired male commonly also sires offspring of other females with whom he is not socially paired (Jennions & Petrie, 2000; Griffith et al., 2002; Parker & Birkhead, 2013). More extra-pair or extra-group paternity than expected from observed parental or territorial behaviour also commonly occurs in socially polygamous populations (Jones et al., 2001; Kvarnemo & Simmons, 2013). Such polygynandry creates an opportunity for individual socially paired males to increase their total reproductive success through reproduction with extra-pair or extra-group females (Webster et al., 1995; Lebigre et al., 2012). However, these same males simultaneously risk losing the paternity of offspring produced by their socially paired female(s) (Westneat & Stewart, 2003; Vedder et al., 2011; Kvarnemo & Simmons, 2013). Substantial variation in male fitness, and consequent selection and evolution, could therefore stem from the direction and magnitude of genetic and environmental covariation in males' success in defending the paternity of their socially paired female's offspring vs. accumulating extra-pair paternity elsewhere (Webster et al., 1995).
Specifically, successful extra-pair sires might have relatively low within-pair paternity success due to negative genetic covariance (i.e. a genetic trade-off) between the two routes to reproductive success. Negative covariance might arise if within-pair and extra-pair reproduction imposed conflicting demands on resource allocation, for example across mate guarding vs. mate searching or across sperm allocations to within-pair vs. extra-pair matings, or if there were directly antagonistic genetic effects on such traits (Parker, 2006). Resulting trade-offs could erode male Bateman gradients and mean that the net evolutionary response to selection on male multiple mating could be small or negative, thereby negating standard evolutionary explanations for polygyny. At the same time, negative genetic covariance between male fitness components would also complicate hypotheses that explain ongoing evolution and persistence of polyandry as a function of positive genetic covariance with individual components of male fitness (e.g. Halliday & Arnold, 1987; Keller & Reeve, 1995; Evans & Simmons, 2008), because negative genetic covariance with other components of male fitness might then arise.
Conversely, successful extra-pair sires might have relatively high within-pair paternity success due to positive genetic covariance (i.e. genetic synergy) between the two routes to reproductive success. Positive covariance might arise if there were genetic variation in resource acquisition, allowing individual males to make correlated investments in within-pair and extra-pair reproduction, or if there were directly pleiotropic effects on components of both within-pair and extra-pair success such as sperm competitiveness or mating frequency. Such synergy might be expected to increase male Bateman gradients and hence facilitate evolution of both polygyny (through direct selection) and polyandry (through indirect selection, see Discussion). Quantifying the direction and magnitude of genetic covariance between male within-pair paternity and extra-pair reproduction specifically, and among other forms of ‘defensive’ vs. ‘offensive’ paternity success more generally, is therefore prerequisite to understanding the (co)evolution and persistence of multiple mating by both sexes (Evans, 2010; Fricke et al., 2010; Engqvist, 2011; Droge-Young et al., 2012; Kvarnemo & Simmons, 2013).
However, such genetic covariances have not been estimated in wild populations where males (and females) experience natural variation in reproductive success. This is due to the difficulty of accurately measuring within-pair paternity and extra-pair reproduction across sufficient males in polygynandrous populations where relatedness is sufficiently high, and measured sufficiently accurately, to support quantitative genetic analyses. It also reflects the difficulty of fitting appropriate quantitative genetic models across components of male reproductive success that have intrinsically non-Gaussian distributions, complex covariances and among-male dependencies. We used 20 years of paternity data from socially monogamous but genetically polygynandrous song sparrows (Melospiza melodia, Wilson) to estimate the additive genetic covariance between a male's liability to sire an offspring produced by his socially paired female (his liability for within-pair paternity success, WPPS) and his total number of extra-pair offspring sired per year (his extra-pair reproductive success, EPRS) and thereby elucidate one key genetic covariance that could shape ongoing evolution of extra-pair reproduction and underlying (co)evolution of polygyny and polyandry. We highlight methodological challenges presented by estimating this covariance and discuss the implications of estimates for understanding the (co)evolution of male and female multiple mating.
Materials and methods
Study system
Mandarte Island, BC, Canada (ca. 6 hectares), holds a resident and primarily socially monogamous song sparrow population which has been studied intensively since 1975 (Smith et al., 2006) and numbered ca. 10–50 breeding pairs during 1993–2012. Both song sparrow sexes can breed from age 1 year, with a median reproductive lifespan of 2 years (interquartile range 1–4 years, Smith et al., 2006; Lebigre et al., 2012). Pairs typically rear up to three broods of offspring during April–July each year, but females can lay up to six clutches given repeated nest failure (Smith et al., 2006). First and last laying dates, and hence breeding season duration, vary substantially among years (Wilson & Arcese, 2003). Females incubate clutches (typically 3–4 eggs), whereas both socially paired parents defend the breeding territory and provision hatched offspring (Smith et al., 2006). Both sexes can form new social pairs between years, and sometimes between breeding attempts within a single year, given mortality or divorce of their previous mate.
Each year, all nests on Mandarte were located and monitored, and all offspring surviving to ca. 6 days post-hatch were marked with unique combinations of metal and coloured rings to allow subsequent identification (Keller, 1998; Smith et al., 2006; Lebigre et al., 2012). All adult immigrants to Mandarte (1.1 per year on average, which is sufficient to prevent inbreeding from rapidly accumulating) were mist-netted and ringed. All adult (≥1 year old) song sparrows alive in each year were identified with annual resighting probability of ca. 1, meaning that all surviving individuals on Mandarte were observed in each year (Wilson et al., 2007). These included all socially paired adults and hence the social parents of all offspring and also included any males that remained socially unpaired due to the typically male-biased adult sex ratio (Sardell et al., 2010; Lebigre et al., 2012).
During 1993–2012, 99.6% of ringed offspring and adults were blood sampled and genotyped at 13 polymorphic microsatellite loci to allow assignment of genetic parents. Bayesian full probability models that incorporated genetic and spatial information assigned genetic sires to 99.7% of sampled offspring with ≥95% individual-level confidence (Sardell et al., 2010; Reid et al., 2014b). Moreover, paternities were subsequently verified using ≥120 polymorphic microsatellite loci and were therefore assigned with extremely high confidence. Overall, ca. 28% of offspring were assigned to males other than a female's observed socially paired mate and hence were classified as extra-pair offspring (Sardell et al., 2010; Reid et al., 2014b; compared with 24% in a nearby mainland song sparrow population, Hill et al., 2011). Genotypes of all observed mothers and offspring were congruent, confirming that mothers were correctly identified from maternal behaviour (Sardell et al., 2010).
For all adult males alive during 1993–2012, the genetic paternity data were used to quantify each male's observed WPPS as the number of offspring sired out of the total offspring ringed in each brood that the male reared (i.e. offspring produced by the male's socially paired female(s)). Phenotypic WPPS was therefore unobservable for socially unpaired males that did not rear any offspring and unobserved for socially paired males whose breeding attempts failed prior to offspring genotyping and paternity assignment at ca. 6 days post-hatch. Each male's EPRS was quantified as the total number of ringed extra-pair offspring sired per year (i.e. offspring sired in broods produced by females other than the male's socially paired mate(s)) and was observed for all adult male song sparrows alive in each year, including males that were socially unpaired. There were therefore no missing phenotypic data for EPRS or WPPS measured as numbers of ringed offspring.
Quantitative genetic analyses
A bivariate animal model was fitted to estimate the additive genetic variances in male EPRS and liability for WPPS and the additive genetic covariance between the two. WPPS was treated as a binomial threshold trait, thereby estimating additive genetic variance in a male's underlying liability to retain rather than lose the paternity of an offspring he reared (e.g. Bennewitz et al., 2007; Reid et al., 2014a). EPRS was assumed to follow an overdispersed Poisson distribution and was not substantially zero-inflated compared with expectation given additive overdispersion. There was therefore little requirement, or power, to estimate parameters pertaining to a distinct zero-inflation process (see Reid et al., 2011a).
The animal model included a variance–covariance matrix of additive genetic random effects derived from pairwise kinship (k) coefficients calculated from pedigree data (Kruuk, 2004). As the phenotypic data spanned 1993–2012 and many males bred in multiple years (see Results), the model also included random year and individual effects on both EPRS and liability for WPPS and hence estimated year and ‘permanent individual’ (co)variances, where the latter are assumed to comprise permanent environmental and nonadditive genetic (co)variances (Kruuk, 2004). The model also included linear regressions on individual coefficient of inbreeding (f), thereby estimating inbreeding depression in male EPRS and liability for WPPS and ensuring that estimated additive genetic (co)variances could not be biased by unmodelled inbreeding depression (Reid & Keller, 2010). The model also included appropriate male age effects specified based on preliminary analyses, namely a linear regression of liability for WPPS on age and three-level effects on EPRS corresponding to age classes 1, 2–5 and ≥6 years.
Multivariate animal models must specify appropriate residual covariance structures to account for instantaneous random effects that influence multiple traits expressed by individuals, otherwise estimated genetic (co)variances could be biased. However, appropriate model specification can be difficult when focal traits are measured on different but overlapping and interacting timescales and sets of individuals, meaning that the form of covariance may be unknown and complex. In the current analysis, male WPPS is most usefully measured per brood rather than per year (i.e. summed across multiple broods that a male reared within a single breeding season). This is because WPPS observed per brood reflects a male's performance in defending paternity during the days when each brood was conceived and equals the observed degree of extra-pair reproduction by his socially paired female. A male's total WPPS observed per year would also incorporate among-brood variation in environmental and female effects on paternity, particularly for males that socially paired with multiple females within a single year. Variance in male liability for WPPS could then stem from among-brood rather than within-brood variation in paternity, which is less directly relevant to understanding the magnitude and mechanisms of selection on male and female multiple mating within single reproductive episodes.
In contrast, a male's total EPRS is most appropriately measured per year rather than in relation to any single breeding event, thereby quantifying a male's total annual extra-pair offspring sired and hence fitness gained through extra-pair reproduction. Breeding attempts are asynchronous across the song sparrow population within any year, because individuals vary in first clutch laying date and subsequent clutch laying dates depend on the timing and success of earlier breeding attempts (Wilson & Arcese, 2003; Smith et al., 2006). As is typical for populations with multibrooded life-histories, there are therefore no clearly distinct population-wide breeding events within individual years within which male EPRS could be measured.
Although only one observation of annual EPRS exists per male per year (hereafter ‘male-year’), many males reared multiple broods per year (see Results), providing multiple observations of WPPS per male-year. Residual covariance therefore cannot be simply estimated as if there were single paired observations of EPRS and WPPS within each male-year. Furthermore, for males that were socially unpaired or otherwise failed to rear any broods of offspring in a particular year, EPRS was observed but WPPS was not. Such males therefore contributed to estimates of additive genetic (co)variances in and among EPRS and WPPS with no possible residual covariance within years. We therefore fitted a model designed to robustly estimate residual covariance across all observations of WPPS and EPRS within each male-year. Specifically, we fitted random male-year effects on male liability for WPPS, thereby accounting for any correlation in WPPS observed across multiple broods reared by a male within a single year. We also fitted random male-year effects on EPRS even though, as there is exactly one observation of EPRS per male-year, male-year variance in EPRS is synonymous with residual variance. By fixing the residual variance in EPRS to a small value, we then forced all additional residual variance to be estimated as male-year variance (while allowing residual variance in male liability for WPPS to be freely estimated). Male-year covariance between EPRS and liability for WPPS was then estimable, thereby accounting for any covariance between EPRS and WPPS within male-years that was not due to additive genetic, permanent individual or year effects (Appendix S1).
Further covariances among observations of WPPS and EPRS for different males within individual years could potentially stem from numerical dependencies among these traits. Specifically, as all offspring have exactly one father, one male's EPRS will depend partly on other males' realized WPPS and vice versa, meaning that observed phenotypes are not entirely independent. However, such dependencies and any consequent biases are likely to be small in the current analysis (see Discussion and Appendix S2).
The males whose WPPS and EPRS were observed were the offspring of numerous different mothers and fathers, meaning that there was little expectation that estimated additive genetic (co)variances could be biased by common parental effects on male phenotypes. Indeed, estimated additive genetic (co)variances remained similar when random parental effects were additionally modelled (Appendix S3).
Analysis implementation
Standard algorithms were used to compute f, k and the inverse relationship matrix from comprehensive pedigree data spanning 1975–2012 (Reid et al., 2011a,b, 2014b; Appendix S4). Kinship between new immigrants and existing Mandarte-hatched natives, and hence f of offspring of immigrant-native pairings, was defined as zero (Reid et al., 2006). Phenotypic data for immigrant males were excluded because f is undefined for immigrants (as opposed to their offspring). As additive genetic (co)variances were estimated across numerous related males whose EPRS and WPPS were observed across numerous years and females, estimates should be relatively unbiased by specific interactions between individual males and females. Interpretation may therefore be less ambiguous than in studies where phenotypes of sets of closely related males are only observed in environments posed by small numbers of females (e.g. García-González & Evans, 2010).
The animal model was fitted using Bayesian methods implemented in package MCMCglmm 2.17 in R v2.15.2 (Hadfield, 2010; R Development Core Team, 2012), using logit and log link functions for WPPS and EPRS, respectively. The model for EPRS estimated additive overdispersion as additional residual variance to that defined by the mean. Fixed effect priors were normally distributed with mean zero and large variance. The model was rerun using a variety of relatively uninformative priors on the (co)variance components and/or genetic correlation, and posterior distributions were robust to such prior variation. The pedigree was pruned to males whose EPRS and WPPS were observed and all their known ancestors. Analyses used 3005000 iterations, burn-in 5000 and thinning interval 3000, ensuring low autocorrelation among thinned samples (< 0.05). Mixing and model convergence were verified by inspecting posterior traces and by qualitative comparison and Gelman-Rubin diagnostics of posterior metrics generated from multiple independent chains.
The posterior distribution of the latent-scale heritability of male liability for WPPS was estimated as VA/(VTotal + π2/3) given logistic variance proportional to π2/3, where VA is the additive genetic variance and VTotal is the sum of all estimated variance components (Nakagawa & Schielzeth, 2010; Reid et al., 2011b). The posterior distribution of the latent-scale heritability of male EPRS was estimated as VA/(VTotal + log(1/exp(xP) + 1)) with exp(xP) taken as the raw mean EPRS (Reid et al., 2011a). Posterior means and 95% highest posterior density credible intervals (95% CI, which are appropriate for skewed posterior distributions) for estimated effects, (co)variances, heritabilities and genetic correlations were estimated across thinned samples, as was the percentage of posterior density for the genetic covariance that exceeded zero.
Further environmental, individual or social effects that could influence WPPS or EPRS were not modelled because the current aim was to partition rather than explain phenotypic variation. Total phenotypic covariance between EPRS and liability for WPPS is not directly observable across all males, because liability for WPPS exists on an underlying scale rather than as a directly observed phenotype, and furthermore, phenotypic WPPS was not observable for all males whose EPRS was observed in each year (because some males were socially unpaired or failed to rear any offspring). However, the total covariance between EPRS and liability for WPPS can be inferred by summing all estimated covariance components. Raw means are presented ± 1SD. Data are available on Dryad.
Results
Distributions of EPRS and WPPS
Male EPRS, defined as the total number of ringed extra-pair offspring sired per year, was observed for 368 individual male song sparrows encompassing 892 male-years (mean 2.4 ± 1.8 years per male, range 1–10). EPRS was zero in 588 (66%) of these male-years. However, there was substantial variation, with up to 11 extra-pair offspring sired (mean 0.9, variance 2.7).
Male WPPS, defined as the number of ringed offspring that a male sired out of each brood produced by his socially paired female(s), was observed for 998 broods reared by 273 individual male song sparrows (mean 3.7 ± 3.0 broods per male, median 3, range 1–19). These 998 broods were reared across 578 male-years, comprising means of 2.1 ± 1.4 years per male (median 2, range 1–7) and 1.7 ± 0.9 (median 2, range 1–6) broods per male-year. There were therefore 578 male-years where EPRS and WPPS were both observed (comprising 256 and 322 male-years when WPPS was observed for one and multiple broods, respectively), and 314 male-years where EPRS was observed but WPPS was not (because males were socially unpaired or failed to rear any offspring). Mean brood size across all 998 observed broods was 2.8 ± 1.0 offspring (median 3, range 1–4). The mean proportion of offspring that a male sired within a brood that he reared was 0.72 ± 0.37 (range 0–1).
Distributions of k and f
The pedigree comprising the 368 male song sparrows for whom WPPS and/or EPRS was observed and all their pedigreed ancestors totalled 671 individuals. Mean pairwise k was 0.058 ± 0.044 among all 671 individuals (median 0.055, range 0.000–0.471, 6% zeros) and 0.071 ± 0.037 among the 368 focal males (median 0.064, range 0.005–0.471). Mean f was 0.064 ± 0.053 across the 368 males (median 0.058, range 0.000–0.308).
(Co)variances in EPRS and liability for WPPS
The animal model estimated moderate additive genetic variance and heritability in both EPRS and liability for WPPS; posterior mean heritabilities were 0.14 (95% CI: 0.06–0.26) and 0.07 (95% CI: 0.02–0.15), respectively (Table1, Fig.1). The posterior mean additive genetic covariance was 0.30, equating to a posterior mean genetic correlation of 0.56 (Table1, Fig.1). Although the 95% CI for the additive genetic covariance was wide and marginally overlapped zero, 97.7% of the posterior density exceeded zero, and the 95% CI for the genetic correlation did not quite overlap zero (Table1, Fig.1). These small differences arose because the posterior distributions were slightly asymmetrical (Fig.1).
Table 1.
Posterior mean estimates (and 95% credible intervals) for additive genetic, permanent individual, year, male-year and residual variances (VA, VPI, VY, VMY and VR, respectively), additive genetic, permanent individual, year and male-year covariances (covA, covPI, covY and covMY, respectively), genetic correlation (rA), heritability (h2), inbreeding depression (β) and age effects in male liability for within-pair paternity success (WPPS) and extra-pair reproductive success (EPRS). For EPRS, age effects are levelled at age class ≥6 years, and levels 1 and 2 show the contrasts with age classes 1 year and 2 to 5 years, respectively. For WPPS, the age effect is the regression slope. VR in EPRS was fixed to 0.1 (Appendix S1)
| VA | covA and rA | VPI | covPI | VY | covY | VMY | covMY | VR | Age | β | h2 | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WPPS | 0.68 (0.14 to 1.28) | covA = 0.30 (−0.02 to 0.65) | 0.45 (0.06 to 1.00) | −0.06 (−0.37 to 0.20) | 0.18 (0.04 to 0.38) | −0.04 (−0.23 to 0.17) | 1.01 (0.14 to 1.95) | 0.43 (0.01 to 0.83) | 3.57 (2.34 to 4.83) | 0.14 (−0.02 to 0.27) | −1.25 (−6.76 to 4.34) | 0.07 (0.02 to 0.15) | |
| EPRS | 0.44 (0.14 to 0.79) | rA = 0.56 (0.01 to 0.87) | 0.32 (0.08 to 0.59) | 0.31 (0.07 to 0.60) | 1.11 (0.75 to 1.51) | 0.1 (fixed) | 1: −0.92 (−1.48 to −0.30) | 2: 0.68 (0.16 to 1.23) | −6.99 (−10.79 to −3.19) | 0.14 (0.06 to 0.26) |
Figure 1.
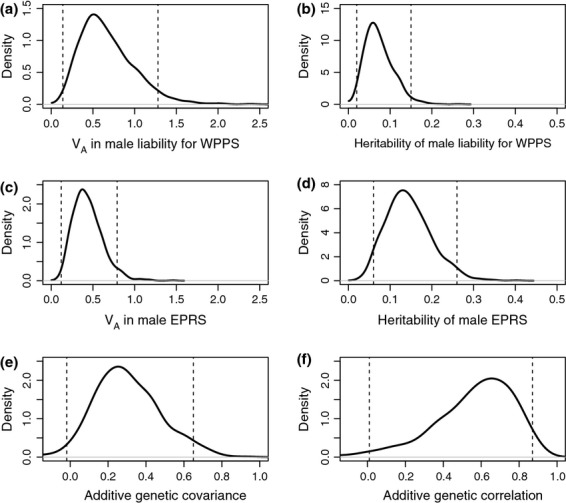
Posterior densities for (a) additive genetic variance (VA) and (b) heritability in male liability for within-pair paternity success (WPPS), (c) additive genetic variance and (d) heritability in male extra-pair reproductive success (EPRS) and the additive genetic (e) covariance and (f) correlation. Dashed lines demarcate 95% credible intervals.
The estimated permanent individual and year variances in EPRS and liability for WPPS were moderate, but the posterior mean covariances were small (Table1). The estimated male-year variances in both traits were substantial (where ‘male-year variance’ in EPRS was forced to estimate ‘residual variance’, Appendix S1), and the posterior mean male-year covariance was positive with a 95% CI that did not overlap zero (Table1). There was also substantial residual variance in liability for WPPS (Table1).
The posterior mean slope of the regression of EPRS on f was negative, demonstrating substantial inbreeding depression in EPRS (Table1). The regression of liability for WPPS on f was also negative, but the 95% CI was wide and overlapped zero (Table1). Liability for WPPS tended to increase with male age, whereas EPRS averaged higher in males aged 2–5 years and lower in 1 year old males than in males aged ≥6 years (Table1).
Discussion
Any (co)evolution of male and female reproductive strategies is expected to depend on the direction and magnitude of genetic covariances among sex-specific life-history components, including among different components of male fitness (Jennions & Petrie, 2000; Kvarnemo & Simmons, 2013; Parker & Birkhead, 2013). Key genetic covariances include those between male reproductive success achieved through potentially conflicting reproductive tactics, where either positive or negative covariances might be predicted (Jones et al., 2001; House & Simmons, 2003; Parker, 2006; Evans, 2010; Fricke et al., 2010; Engqvist, 2011; Kvarnemo & Simmons, 2013). Empirical estimates of such covariances are therefore required, ideally across males experiencing natural variation in reproductive success.
In the context of socially monogamous but genetically polygynandrous systems, one pertinent genetic covariance is that between a male's propensity to sire an offspring produced by his socially paired female (i.e. his liability for within-pair paternity success, WPPS) and his total extra-pair reproductive success (EPRS) achieved by siring other females' offspring. The interpretation and importance of this covariance perhaps require clarification in the context of the wider literature. It does not equate to the genetic covariance between a male's liability for WPPS and his analogous liability to sire an offspring of any individual extra-pair female with whom he mates (i.e. his ‘defensive’ vs. ‘offensive’ paternity success as could be strictly defined as analogous post-copulatory traits, House & Simmons, 2003; Fricke et al., 2010). Nor does it equate to the genetic covariance between a male's total EPRS and his analogous total within-pair reproductive success (which depends on pairing success and female fecundity as well as WPPS, Webster et al., 1995). Furthermore, it does not equate to the genetic covariance between distinct precopulatory and post-copulatory episodes of sexual selection (i.e. mating success vs. subsequent fertilization success, Hosken et al., 2008; Droge-Young et al., 2012; Pischedda & Rice, 2012; Parker & Birkhead, 2013). Rather, the genetic covariance between male EPRS and liability for WPPS is of interest for three primary reasons.
First, it encapsulates the potential male trade-off between defending paternity of a socially paired female's offspring vs. achieving additional total extra-pair reproductive success elsewhere (Westneat & Stewart, 2003; Akçay et al., 2012; Vedder et al., 2011). It therefore describes the degree of evolutionary conflict or synergy across these male reproductive tactics and will influence the resulting male Bateman gradient.
Second, by affecting a male's ability to sire offspring produced by his socially paired female, a male's liability for WPPS influences his paired female's realized degree of extra-pair reproduction, whereas EPRS measures the male's reproductive success gained through extra-pair reproduction. The genetic covariance between the two components of male fitness therefore encapsulates one dimension of the potential for the observed degree of female extra-pair reproduction to evolve through genetic covariance with male extra-pair reproduction.
Third, a male's additive genetic value for EPRS can be interpreted to indicate his genetic propensity to sire any individual extra-pair offspring that is produced across a population. The genetic covariance between male EPRS and liability for WPPS will therefore shape the overall genetic covariance between female extra-pair reproduction and male liability for WPPS that could arise due to linkage disequilibria given the population-wide pattern of within-pair and extra-pair paternity. Specifically, females with high genetic value for extra-pair reproduction are by definition likely to produce extra-pair offspring with males with high genetic value for EPRS. The genetic covariance between male EPRS and liability for WPPS will therefore shape the covariance between female liability for extra-pair reproduction and male liability for WPPS that emerges across resulting offspring (Reid et al., 2014a). As a substantial proportion of variation in male fitness might stem from variation in WPPS (Webster et al., 1995; Lebigre et al., 2012), this covariance could facilitate ongoing evolution of female extra-pair reproduction through indirect selection.
In summary, the genetic covariance between male EPRS and liability for WPPS, as estimated here, could shape the evolutionary dynamics of both male and female extra-pair reproduction and hence of the overall socially monogamous but genetically polygynandrous reproductive system.
Additive genetic (co)variances
Analyses of song sparrow paternity data estimated nonzero additive genetic variance in male liability for WPPS, with a latent-scale heritability of ca. 0.07. This implies that the paternity status of offspring within broods that a male rears, and hence the observed degree of extra-pair reproduction by the male's socially paired female, is influenced by additive genetic effects of a female's socially paired male as well as by additive genetic effects of the female herself (Reid et al., 2014a). The observed degree of extra-pair reproduction could therefore potentially evolve through selection on males as well as through any selection on females.
There was also nonzero additive genetic variance in male song sparrows' total annual EPRS, with an estimated heritability of ca. 0.14. This estimate contrasts with previous univariate analyses that did not detect such additive genetic variance in EPRS in the same song sparrow population (Reid et al., 2011a). This change stems from four additional years of phenotypic and pedigree data that include nonzero EPRS in related males. Evidence of nonzero heritability of male EPRS suggests one mechanism that could facilitate the evolution of female extra-pair reproduction and underlying polyandry. It implies that polyandrous females, who will by definition produce extra-pair offspring with males who are successful extra-pair sires, will on average produce extra-pair sons who are themselves relatively successful extra-pair sires, potentially creating indirect selection for polyandry and extra-pair reproduction (Wedell & Tregenza, 1999; Jennions & Petrie, 2000; Firman, 2011; Reid et al., 2011a; Klemme et al., 2014).
However, any coevolution of male and female extra-pair reproduction, and underlying polygyny and polyandry, will also depend on the additive genetic covariance between male EPRS and liability for WPPS. The estimated genetic covariance and correlation were positive in song sparrows (posterior means of 0.30 and 0.56, respectively). Although the estimated 95% CI for the genetic covariance was wide and marginally overlapped zero, more than 95% of the posterior density exceeded zero and the estimated 95% CI for the genetic correlation did not quite overlap zero. These positive estimates imply that males with high additive genetic liability to sire offspring produced by their socially paired female also had high additive genetic value for siring extra-pair offspring produced by other females. More conservatively, there was no evidence of substantial negative genetic covariance (i.e. a genetic trade-off) between the two routes to male reproductive success.
However, although the fitted animal model should adequately account for residual covariance between male EPRS and liability for WPPS within male-years (Appendix S1), some difficulties of analysis, inference and interpretation remain. Precise inference might be impeded or biased by numerical dependencies that arise because observations of EPRS and WPPS are not entirely independent across males within years (Appendix S2). Such biases might be best eliminated by fitting models that explicitly consider each individual male's liability to sire each individual offspring (rather than fitting models that consider whether or not an offspring was sired by its mother's socially paired male). However, the resulting high dimensionality is likely to render such models impractical to fit, even if restricted sets of potential sires relevant to each individual offspring were identified within the model structure. Instead, analyses of restricted data sets that minimized among-male dependencies suggested that any such biases in the current analyses are probably small (Appendix S2), supporting the conclusion that the additive genetic covariance between a male's liability to sire an offspring produced by his socially paired female and his reproductive success accrued by siring other females' offspring is most probably positive.
Positive genetic covariance between male EPRS and liability for WPPS could stem from pleiotropic genetic effects on post-copulatory and/or precopulatory processes. For example, common alleles could potentially promote success in sperm competition in the contexts of both socially paired and extra-pair females, or increase mating frequencies with both. However, in common with most field and experimental studies, EPRS and WPPS were measured across offspring that survived to paternity assignment at some point post-hatch or post-birth. Estimated genetic (co)variances might consequently reflect variation in pre-assignment offspring mortality in relation to paternity rather than variation in paternity per se (e.g. García-González, 2008; Droge-Young et al., 2012). The estimated additive genetic variance in male liability for WPPS probably does primarily reflect variation in within-pair fertilization success in song sparrows, because estimates remained quantitatively similar in univariate analyses that were restricted to breeding attempts where paternity was assigned to all conceived offspring (Reid et al., 2014a). In contrast, the estimated additive genetic variance in male EPRS might partly reflect genetic variation in early survival of extra-pair offspring sired by different males rather than in extra-pair mating and/or fertilization success; such effects are difficult to quantify without complete data describing paternity at conception. However, estimating the genetic covariance between male liability for WPPS and EPRS across hatched extra-pair offspring is still valuable in the context of understanding the evolutionary dynamics of extra-pair reproduction, because extra-pair offspring that die prehatch cannot contribute to female or male fitness or contribute to future correlated transmission of alleles underlying WPPS, EPRS or multiple mating.
Environmental effects
Liability for WPPS and EPRS also showed substantial positive male-year covariance across male song sparrows (and therefore, positive total covariance calculated as the sum of all estimated covariance components, Table1). This implies that environmental effects that increased a male's liability to sire his socially paired female's offspring in a particular year also increased his success in siring extra-pair offspring in that year. Such positive covariance could stem from variation in resource acquisition and hence in ‘condition’ or attractiveness and consequent fertilization success (whether due to sperm competition or cryptic female choice) and/or mating success (Kvarnemo & Simmons, 2013). The absence of a phenotypic or genetic trade-off between male EPRS and liability for WPPS may reflect the song sparrow's multibrooded life-history and consequent local asynchrony of breeding attempts (Smith et al., 2006). Guarding or inseminating socially paired female(s) during their fertile period(s) might therefore not preclude males from previously or subsequently mating with fertile extra-pair females (e.g. Yezerinac & Weatherhead, 1997; Griffith et al., 2002; Westneat & Stewart, 2003).
Implications and context
Positive genetic covariance between male EPRS and liability for WPPS might be predicted to facilitate evolution of male extra-pair reproduction, and underlying polygyny, because EPRS will experience both positive direct selection and positive indirect selection stemming from genetic covariance with WPPS. A positive Bateman gradient between mate and offspring numbers is likely to result (Parker & Birkhead, 2013). However, (co)evolution of absolute EPRS and WPPS must ultimately be constrained because all males within a population cannot simultaneously be both successful within-pair sires and successful extra-pair sires. Some form of soft selection and/or a genetic trade-off with some other component(s) of male or female fitness might therefore exist or arise.
Positive genetic covariance between male EPRS and liability for WPPS might also facilitate evolution of female extra-pair reproduction, and underlying polyandry, because extra-pair males with whom polyandrous females produce offspring are likely to have high additive genetic value for both EPRS (by definition) and for WPPS (due to genetic covariance). Positive genetic covariance between female propensity for polyandry and both components of male fitness could result. However, the magnitude of such cross-sex genetic covariances, and their evolutionary consequences, will also depend on the degree to which genetic covariances among female and male fitness components stem from pleiotropy vs. linkage disequilibria, and on any pattern of assortative reproduction with respect to female and male genetic values for polyandry and polygyny and associated fitness components.
Additive genetic (co)variances among male EPRS and liability for WPPS, or other broadly analogous components of male reproductive success, have not previously been explicitly estimated in wild populations. Indeed, the challenge of measuring variation in EPRS, which ideally requires paternity to be assigned to all offspring and males in a population, means that even phenotypic (co)variances between EPRS and observed WPPS have rarely been rigorously estimated (Vedder et al., 2011; Lebigre et al., 2012; see also Shuster, 2009). However, aspects of paternal behaviour and paternity varied with a chromosomal inversion (and associated colour morphs) in white-throated sparrows (Zonotrichia albicollis) implying that paternity success can have a genetic basis (Tuttle, 2003).
Phenotypic covariances among different components of male reproductive success, including those stemming from precopulatory vs. post-copulatory processes, have been estimated in experimental populations. Mating success and fertilization success (or associated traits) can be negatively correlated, indicating that post-copulatory sexual selection stemming from polyandry could decrease the overall opportunity for selection on male traits (Jones et al., 2001; Kvarnemo & Simmons, 2013). However, they can also be uncorrelated (Pischedda & Rice, 2012) or positively correlated, indicating that sequential episodes of sexual selection can be reinforcing (Droge-Young et al., 2012; Parker & Birkhead, 2013).
Although additive genetic variance in male paternity success can be substantial in experimental populations (Evans & Simmons, 2008; Simmons & Moore, 2009; Forstmeier et al., 2011), relatively few studies have explicitly estimated genetic covariances. The estimated genetic correlation between male latency to copulate and paternity success as second male was negative in Drosophila simulans, indicating a positive genetic correlation between male mating success and post-copulatory paternity success (Hosken et al., 2008). In contrast, strong negative genetic covariances among traits associated with mating success and post-copulatory fertilization success were observed in guppies (Poecilia reticulata), implying that the reproductive tactics of ‘courting’ and ‘sneaking’ may be genetically constrained (Evans, 2010). Negative covariance between measures of male attractiveness and nuptial provisioning or sperm viability (and hence expected fertilization success) was also observed across full-sib scorpionfly families (Panorpa cognate, Engqvist, 2011) and half-sib Australian cricket families (Teleogryllus oceanicus, Simmons et al., 2010). Further studies, and methodological developments, are therefore required before robust general conclusions regarding the magnitude of genetic covariances among key components of male reproductive success, the causes of such (co)variances or the consequent implications for (co)evolution of polygyny and polyandry, can be drawn.
Acknowledgments
We thank the Tsawout and Tseycum first nations bands for allowing access to Mandarte, everyone who contributed to long-term data collection, Rebecca Sardell and Pirmin Nietlisbach for their meticulous paternity assignments, Jarrod Hadfield for invaluable analytical advice, and the European Research Council, Marie Curie Actions, the UK Royal Society, Swiss National Science Foundation and the Natural Sciences and Engineering Research Council of Canada for funding.
Supporting Information
Additional Supporting Information may be found in the online version of this article:
Appendix S1 Summary of animal model structure.
Appendix S2 Numerical dependencies among observations of EPRS and WPPS.
Appendix S3 Distributions of focal males across mothers and fathers, and maternal and paternal (co)variances.
Appendix S4 Summary of pedigree data.
References
- Akçay Ḉ, Searcy WA, Campbell SE, Reed VA, Templeton CN, Hardwick KM, et al. Who initiates extrapair mating in song sparrows? Behav. Ecol. 2012;23:44–50. [Google Scholar]
- Bennewitz J, Morgades O, Preisinger R, Thaller G, Kalm E. Variance component and breeding value estimation for reproductive traits in laying hens using a Bayesian threshold model. Poult. Sci. 2007;86:823–828. doi: 10.1093/ps/86.5.823. [DOI] [PubMed] [Google Scholar]
- Droge-Young EM, Manier MK, Lüpold S, Belote JM, Pitnick S. Covariance among premating, post-copulatory and viability fitness components in Drosophila melanogaster and their influence on paternity measurement. J. Evol. Biol. 2012;25:1555–1563. doi: 10.1111/j.1420-9101.2012.02540.x. [DOI] [PubMed] [Google Scholar]
- Engqvist L. Male attractiveness is negatively genetically associated with investment in copulations. Behav. Ecol. 2011;22:345–349. [Google Scholar]
- Evans JP. Quantitative genetic evidence that males trade attractiveness for ejaculate quality in guppies. Proc. Biol. Sci. 2010;277:3195–3201. doi: 10.1098/rspb.2010.0826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evans JP, Simmons LW. The genetic basis of traits regulating sperm competition and polyandry: can selection favour the evolution of good- and sexy-sperm? Genetica. 2008;134:5–19. doi: 10.1007/s10709-007-9162-5. [DOI] [PubMed] [Google Scholar]
- Firman RC. Polyandrous females benefit by producing sons that achieve high reproductive success in a competitive environment. Proc. Biol. Sci. 2011;278:2823–2831. doi: 10.1098/rspb.2010.2791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Forstmeier W, Martin K, Bolund E, Schielzeth H, Kempenaers B. Female extrapair mating behaviour can evolve via indirect selection on males. Proc. Natl. Acad. Sci. U.S.A. 2011;108:10608–10613. doi: 10.1073/pnas.1103195108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fricke C, Martin OY, Bretman A, Bussière LF, Chapman T. Sperm competitive ability and indices of lifetime reproductive success. Evolution. 2010;64:2746–2757. doi: 10.1111/j.1558-5646.2010.01022.x. [DOI] [PubMed] [Google Scholar]
- García-González F. Male genetic quality and the inequality between paternity success and fertilization success: consequences for studies of sperm competition and the evolution of polyandry. Evolution. 2008;62:1653–1665. doi: 10.1111/j.1558-5646.2008.00362.x. [DOI] [PubMed] [Google Scholar]
- García-González F, Evans JP. Fertilization success and the estimation of genetic variance in sperm competitiveness. Evolution. 2010;65:746–756. doi: 10.1111/j.1558-5646.2010.01127.x. [DOI] [PubMed] [Google Scholar]
- Griffith SC, Owens IPF, Thuman KA. Extra-pair paternity in birds: a review of interspecific variation and adaptive function. Mol. Ecol. 2002;11:2195–2212. doi: 10.1046/j.1365-294x.2002.01613.x. [DOI] [PubMed] [Google Scholar]
- Hadfield JD. MCMC methods for multi-response generalized linear mixed models: the MCMCglmm R package. J. Stat. Softw. 2010;33:1–22. [Google Scholar]
- Halliday T, Arnold SJ. Multiple mating by females: a perspective from quantitative genetics. Anim. Behav. 1987;35:939–941. [Google Scholar]
- Hill CE, Akçay Ḉ, Campbell SE, Beecher MD. Extra-pair paternity, song and genetic quality in song sparrows. Behav. Evol. 2011;22:73–81. [Google Scholar]
- Hosken DJ, Taylor ML, Hoyle K, Higgins S, Wedell N. Attractive males have greater success in sperm competition. Curr. Biol. 2008;18:553–554. doi: 10.1016/j.cub.2008.04.028. [DOI] [PubMed] [Google Scholar]
- House CM, Simmons LW. Genital morphology and fertilization success in the dung beetle Onthophagus taurus: an example of sexually selected male genitalia. Proc. R. Soc. Lond. B Biol. Sci. 2003;270:447–455. doi: 10.1098/rspb.2002.2266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jennions MD, Petrie M. Why do females mate multiply? A review of the genetic benefits. Biol. Rev. 2000;75:21–64. doi: 10.1017/s0006323199005423. [DOI] [PubMed] [Google Scholar]
- Jones AG, Walker D, Kvarnemo C, Lindström K, Avise JC. How cuckoldry can decrease the opportunity for sexual selection: data and theory from a genetic parentage analysis of the sand goby, Pomoatoschistus minutus. Proc. Natl. Acad. Sci. U.S.A. 2001;98:9151–9156. doi: 10.1073/pnas.171310198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keller LF. Inbreeding and its fitness effects in an insular population of song sparrows (Melospiza melodia. Evolution. 1998;52:240–250. doi: 10.1111/j.1558-5646.1998.tb05157.x. [DOI] [PubMed] [Google Scholar]
- Keller L, Reeve HK. Why do females mate with multiple males? The sexually selected sperm hypothesis. Adv. Stud. Behav. 1995;24:291–315. [Google Scholar]
- Klemme I, Bäumer J, Eccard JA, Ylönen H. Polyandrous females produce sons that are successful at post-copulatory competition. J. Evol. Biol. 2014;27:457–465. doi: 10.1111/jeb.12334. [DOI] [PubMed] [Google Scholar]
- Kruuk LEB. Estimating genetic parameters in natural populations using the ‘animal model’. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2004;359:873–890. doi: 10.1098/rstb.2003.1437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kruuk LEB, Slate J, Wilson AJ. New answers for old questions: the evolutionary quantitative genetics of wild animal populations. Annu. Rev. Ecol. Evol. Syst. 2008;39:525–548. [Google Scholar]
- Kvarnemo C, Simmons LW. Polyandry as a mediator of sexual selection before and after mating. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013;368:20120042. doi: 10.1098/rstb.2012.0042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lande R. A quantitative genetic theory of life-history evolution. Ecology. 1982;63:607–615. [Google Scholar]
- Lebigre C, Arcese P, Sardell RJ, Keller LF, Reid JM. Extra-pair paternity and the variance in male fitness in song sparrows (Melospiza melodia. Evolution. 2012;66:3111–3129. doi: 10.1111/j.1558-5646.2012.01677.x. [DOI] [PubMed] [Google Scholar]
- Nakagawa S, Schielzeth H. Repeatability for Gaussian and non-Gaussian data: a practical guide for biologists. Biol. Rev. 2010;85:935–956. doi: 10.1111/j.1469-185X.2010.00141.x. [DOI] [PubMed] [Google Scholar]
- Parker GA. Sexual conflict over mating and fertilization: an overview. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2006;361:235–259. doi: 10.1098/rstb.2005.1785. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Parker GA, Birkhead TR. Polyandry: the history of a revolution. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013;368:20120335. doi: 10.1098/rstb.2012.0335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pischedda A, Rice WR. Partitioning sexual selection into its mating success and fertilisation success components. Proc. Natl. Acad. Sci. U.S.A. 2012;109:2049–2053. doi: 10.1073/pnas.1110841109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pizzari T, Birkhead TR. The sexually-selected sperm hypothesis: sex-biased inheritance and sexual antagonism. Biol. Rev. 2002;77:183–209. doi: 10.1017/s1464793101005863. [DOI] [PubMed] [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing; 2012. [Google Scholar]
- Reid JM, Keller LF. Correlated inbreeding among relatives: occurrence, magnitude and implications. Evolution. 2010;64:973–985. doi: 10.1111/j.1558-5646.2009.00865.x. [DOI] [PubMed] [Google Scholar]
- Reid JM, Arcese P, Keller LF. Intrinsic parent-offspring correlation in inbreeding level in a song sparrow (Melospiza melodia) population open to immigration. Am. Nat. 2006;168:1–13. doi: 10.1086/504852. [DOI] [PubMed] [Google Scholar]
- Reid JM, Arcese P, Sardell RJ, Keller LF. Additive genetic variance, heritability and inbreeding depression in male extra-pair reproductive success. Am. Nat. 2011a;177:177–187. doi: 10.1086/657977. [DOI] [PubMed] [Google Scholar]
- Reid JM, Arcese P, Sardell RJ, Keller LF. Heritability of female extra-pair paternity rate in song sparrows (Melospiza melodia. Proc. R. Soc. Lond. B Biol. Sci. 2011b;278:1114–1120. doi: 10.1098/rspb.2010.1704. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reid JM, Arcese P, Keller LF, Losdat S. Female and male genetic effects on offspring paternity: additive genetic (co)variances among female extra-pair reproduction and male within-pair paternity success in song sparrows (Melospiza melodia. Evolution. 2014a doi: 10.1111/evo.12424. doi: 10.1111/evo.12424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reid JM, Keller LF, Marr AB, Nietlisbach P, Sardell RJ, Arcese P. Pedigree error due to extra-pair reproduction substantially biases estimates of inbreeding depression. Evolution. 2014b;68:802–815. doi: 10.1111/evo.12305. [DOI] [PubMed] [Google Scholar]
- Robinson MR, Beckerman AP. Quantifying multivariate plasticity: genetic variation in resource acquisition drives plasticity in resource allocation to components of life-history. Ecol. Lett. 2013;16:281–290. doi: 10.1111/ele.12047. [DOI] [PubMed] [Google Scholar]
- Roff DA. Life-history Evolution. Sunderland, MA: Sinauer; 2002. [Google Scholar]
- Sardell RJ, Keller LF, Arcese P, Bucher T, Reid JM. Comprehensive paternity assignment: genotype, spatial location and social status in song sparrows Melospiza melodia. Mol. Ecol. 2010;19:4352–4364. doi: 10.1111/j.1365-294X.2010.04805.x. [DOI] [PubMed] [Google Scholar]
- Shuster SM. Sexual selection and mating systems. Proc. Natl. Acad. Sci. U.S.A. 2009;106:10009–10016. doi: 10.1073/pnas.0901132106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simmons LW. Evolutionary quantitative genetics of sperm. In: Birkhead TR, Hosken D, Pitnick S, Moore AJ, editors. Sperm Biology: An Evolutionary Perspective. London: Academic Press; 2009. pp. 405–434. [Google Scholar]
- Simmons LW, Tinghitella RM, Zuk M. Quantitative genetic variation in courtship song and its covariation with immune function and sperm quality in the field cricket Teleogryllus oceanicus. Behav. Ecol. 2010;21:1330–1336. [Google Scholar]
- Slatyer RA, Mautz BS, Backwell PRY, Jennions MD. Estimating genetic benefits of polyandry from experimental studies: a meta-analysis. Biol. Rev. 2012;87:1–33. doi: 10.1111/j.1469-185X.2011.00182.x. [DOI] [PubMed] [Google Scholar]
- Smith JNM, Keller LF, Marr AB, Arcese P. Conservation and Biology of Small Populations: The Song Sparrows of Mandarte Island. New York, NY: Oxford University Press; 2006. [Google Scholar]
- Stearns SC. Trade-offs in life-history evolution. Func. Ecol. 1989;3:259–268. [Google Scholar]
- Tuttle EM. Alternative reproductive strategies in the white-throated sparrow: behavioural and genetic evidence. Behav. Ecol. 2003;14:425–432. [Google Scholar]
- Vedder O, Komdeur J, van der Velde M, Schut E, Magrath MJL. Polygyny and extra-pair paternity enhance the opportunity for sexual selection in blue tits. Behav. Ecol. Sociobiol. 2011;65:741–752. doi: 10.1007/s00265-010-1078-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Webster MS, Pruett-Jones S, Westneat DF, Arnold SJ. Measuring the effects of pairing success, extra-pair copulations and mate quality on the opportunity for sexual selection. Evolution. 1995;49:1147–1157. doi: 10.1111/j.1558-5646.1995.tb04441.x. [DOI] [PubMed] [Google Scholar]
- Wedell N, Tregenza T. Successful fathers sire successful sons. Evolution. 1999;53:620–625. doi: 10.1111/j.1558-5646.1999.tb03797.x. [DOI] [PubMed] [Google Scholar]
- Westneat DF, Stewart IRK. Extra-pair paternity in birds: causes, correlates and conflict. Annu. Rev. Ecol. Evol. Syst. 2003;34:365–396. [Google Scholar]
- Wilson S, Arcese P. El Niño drives timing of breeding but not population growth in the song sparrow (Melospiza melodia. Proc. Natl. Acad. Sci. U.S.A. 2003;100:11130–11142. doi: 10.1073/pnas.1931407100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wilson S, Norris DR, Wilson AG, Arcese P. Breeding experience and population density affect the ability of a songbird to respond to future climate variation. Proc. R. Soc. Lond. B Biol. Sci. 2007;274:2539–2545. doi: 10.1098/rspb.2007.0643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yezerinac SM, Weatherhead PJ. Reproductive synchrony and extra-pair mating strategy in a socially monogamous bird, Dendroica petechia. Anim. Behav. 1997;54:1393–1403. doi: 10.1006/anbe.1997.0545. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Appendix S1 Summary of animal model structure.
Appendix S2 Numerical dependencies among observations of EPRS and WPPS.
Appendix S3 Distributions of focal males across mothers and fathers, and maternal and paternal (co)variances.
Appendix S4 Summary of pedigree data.