

Abstract
The retinoids are a family of compounds that in nature are derived from vitamin A or pro-vitamin A carotenoids. An essential part of the diet for mammals, vitamin A has long been known to be essential for many organ systems in the adult. More recently, however, they have been shown to be necessary for function of the brain and new discoveries point to a central role in processes ranging from neuroplasticity to neurogenesis. Acting in several regions of the central nervous system including the eye, hippocampus and hypothalamus, one common factor in its action is control of biological rhythms. This review summarizes the role of vitamin A in the brain; its action through the metabolite retinoic acid via specific nuclear receptors, and the regulation of its concentration through controlled synthesis and catabolism. The action of retinoic acid to regulate several rhythms in the brain and body, from circadian to seasonal, is then discussed to finish with the importance of retinoic acid in the regular pattern of sleep.
We review the role of vitamin A and retinoic acid (RA) as mediators of rhythm in the brain. In the suprachiasmatic nucleus and hippocampus they control expression of circadian clock genes while in the cortex retinoic acid is required for delta oscillations of sleep. Retinoic acid is also central to a second rhythm that keeps pace with the seasons, regulating function in the hypothalamus and pineal gland.
Keywords: circadian, neural plasticity, nuclear receptor, photoperiod, retinoic acid, vitamin A
Vitamin A is a vital component of the mammalian diet that is delivered to tissues in the form of circulating retinol (Fig.1, structure 2). Retinol is metabolized to the transcriptionally active compound retinoic acid (RA; Fig.1, structure 5). That RA is a key molecule during embryonic development is well established (Ross et al. 2000) and it is particularly crucial during development of the central nervous system (CNS; Maden 2002). More recently, however, results point to an important role for RA in the adult CNS and evidence suggests that RA mediates neuronal plasticity (McCaffery et al. 2006; Chen et al. 2012) as well as neurogenesis in both the hippocampus (Goodman et al. 2012) and hypothalamus (Shearer et al. 2012b). Now, a new homeostatic role is emerging for RA as a regulator of biological rhythms within the CNS (Fig.2).
Fig 1.
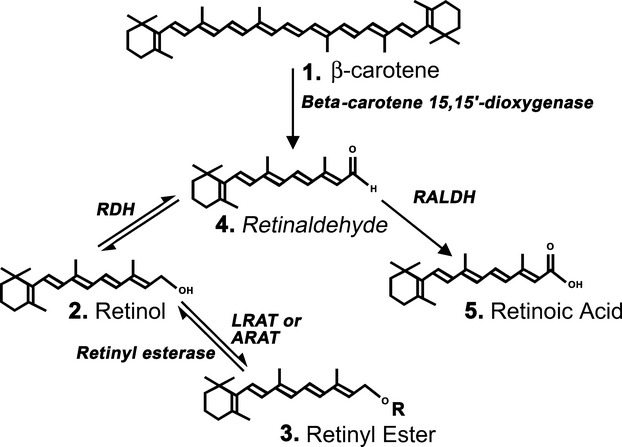
Chemical structures of retinoid family members. β-carotene (structure 1) is cleaved by β-carotene 15,15′-monooxygenase to form two molecules of retinaldehyde (structure 4). The double arrow between retinol (structure 2) and retinaldehyde (structure 4) indicates the interconversion between the two retinoids catalyzed by retinol dehydrogenases (RDHs) primarily RDH1 and RDH10 and dehydrogenase/reductase (SDR family) member 9 (DHRS9). The single arrow between retinaldehyde (structure 4) and retinoic acid (structure 5) indicates the irreversible oxidation catalyzed by retinaldhyde dehydrogenase (RALDH) family members. Esterification of retinol is carried out by lecithin:retinol acyltransferase (LRAT) or acyl-CoA:retinol acyltransferase (ARAT). The predominant retinyl ester (structure 3) is retinyl palmitate (r = C15H31).
Fig 2.
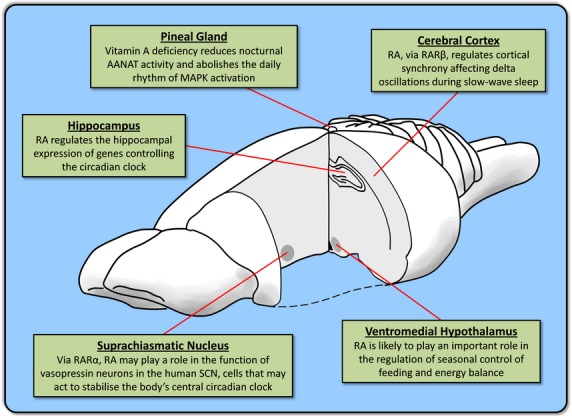
Retinoic acid in the CNS regulates biological rhythms. Retinoic acid (RA) may contribute to the regulation of circadian rhythms in the suprachiasmatic nucleus (SCN), the central clock of the body (Meng et al. 2011), the pineal gland (Herbert and Reiter 1985; Guillaumond et al. 2005), which signals day/night to the rest of the body, and peripheral clocks such as that of the hippocampus (Golini et al. 2012). There is also considerable evidence suggesting that RA plays a crucial role in the seasonal rhythms of energy balance in the ventromedial hypothalamus (Anzano et al. 1979; Ross et al. 2005; Shearer et al. 2010). Rhythmic EEG oscillations in the cerebral cortex during sleep are altered by RA signalling (Maret et al. 2005).
It is now evident that several components of the RA signalling pathway oscillate according to photoperiodic changes in light conditions. Increased day length leads to enhanced retinoid signalling within the hypothalamus of the F344 photoperiodic rat (Shearer et al. 2010). The daily peak in melatonin synthesis by the pineal gland is known to be reduced under vitamin A-deficient conditions in the Japanese quail (Fu et al. 1999). In addition, polymorphisms in the RA receptor β gene can regulate the relative contribution of delta oscillations to slow-wave sleep (Maret et al. 2005). These observations point to roles for RA in both circadian and seasonal rhythms, and also in the cortical regulation of sleep. This review will discuss the mechanisms that link RA with these biological rhythms within the CNS.
The vitamin A signalling pathway
Vitamin A and related retinoid compounds (Fig.1) have a similar chemical structure containing a 6 carbon β-ionone ring and an isoprenoid chain termed a retinyl group. Mammals are not able to generate vitamin A de novo and must instead consume plant or animal sources of this dietary nutrient. Vitamin A is vulnerable to loss through oxidation and in animal tissues is found in the more stable form of fat-soluble retinyl esters. Vitamin A can also be obtained from plant food sources such as β-carotene, which can be cleaved by the enzyme β-carotene 15,15′-monooxygenase to retinaldehyde (Fig.1).
Retinol derived from either dietary source is absorbed by cells of the intestinal mucosa where it is bound by cellular retinol binding protein II (CRBP-II; Noy 2000). CRBP-II then specifically directs retinol substrate to the enzyme lecithin:retinol acyltransferase (Fig.1; Herr and Ong 1992), resulting in the re-esterification of retinol to retinyl esters. These are packaged with other dietary lipids into chylomicrons and secreted from the intestinal mucosa into the general circulation (Blomhoff et al. 1990). The liver is the principal site for the storage and metabolism of retinoids. Here, hydrolysis of retinyl esters results in the formation of retinol which associates with retinol-binding protein (RBP; Newcomer and Ong 2000). Binding of retinol to RBP results in secretion of retinol-RBP into the blood plasma for transportation to peripheral tissues (Ronne et al. 1983).
Retinoic acid synthesis
The transport of retinol in the general circulation coupled with its lipophilic nature renders retinol available to virtually all cells. However, not all tissues can oxidize retinol to RA; only those with a requirement for RA will do this. The ability of cells to synthesize RA and therefore to initiate retinoid signalling is determined by the expression of RA synthetic enzymes. Fig.1 illustrates cellular RA synthesis in two oxidative stages (Napoli 2012). Retinol is first converted to retinaldehyde, a reversible reaction catalysed by retinol dehydrogenases (RDHs) primarily RDH1 and RDH10 and dehydrogenase/reductase (SDR family) member 9 (DHRS9). Retinaldehyde is then converted to RA in an irreversible reaction catalysed by retinaldehyde dehydrogenases (RALDH1, 2 and 3, also called ALDH1A1, 2 and 3; McCaffery et al. 1992; Suzuki et al. 2000).
In both mouse and human, the three isoforms of RALDH are highly expressed by tissues of the developing embryo. RALDH1 is specifically expressed in the dorsal retina where RA signalling contributes to dorsoventral patterning (McCaffery et al. 1999), and is also present in the axons and terminals of a subset of tyrosine hydroxylase-expressing dopaminergic neurons of the mesostriatal and mesolimbic system. These neurons form a RA-generating projection from the substantia nigra to the corpus striatum and nucleus accumbens (McCaffery and Drager 1994; Jacobs et al. 2007). RALDH2 is widely expressed throughout the embryo during development; RALDH2-/- embryos fail to develop beyond E8.5, demonstrating the essential role of RALDH2 in development (Niederreither et al. 1999). RALDH3 expression is confined to the ventral retina (McCaffery et al. 1992) and the lateral ganglionic eminence of the telencephalon (Li et al. 2000). Here, RA signalling regulates cell proliferation, migration and fate determination (Molotkova et al. 2007; Crandall et al. 2011).
The RALDHs are key to the function of RA in the embryonic CNS and are strongly expressed in a variety of locations during embryogenesis (McCaffery et al. 1992, 1999; McCaffery and Drager 1994). However, in the adult rodent, there is a stark reduction in the number of tissues that express RALDHs, and by implication the number of tissues able to generate RA and participate in RA signalling. These limited regions include the hypothalamus in which the RALDHs are expressed in tanycytes to potentially control hypothalamic regulation of energy balance (Shearer et al. 2010) by mechanisms that include regulation of neurogenesis (Shearer et al. 2012b). In the hippocampus, RA is synthesized from retinaldehyde by the high levels of RALDH2 expressed in the meninges immediately adjacent to the dentate gyrus, which RA is presumed to enter by diffusion (Goodman et al. 2012). This is possible in the rodent hippocampus because of its relatively small size, but would be ineffective in supplying the much larger human brain. In the human hippocampus, high levels of all three RALDH enzymes are expressed by the neurons themselves in both the dentate gyrus and cornus ammonis (Fragoso et al. 2012). This potentially provides high levels of RA in the human hippocampus and the dynamics of RA supply in the human brain are likely different from that in the rodent. The discovery of widespread RA synthesis by neurons in the human hippocampus serves to emphasize that RA is crucial for hippocampal function in all mammalian species, regulating the neuroplasticity essential for learning and memory (Shearer et al. 2012a).
The actions of retinoic acid – regulation of transcription
Upon synthesis, the primary action of RA is to regulate the transcription of target genes, which is achieved through the activation of nuclear receptors. The retinoic acid receptors (RARα, β and γ) are ligand-induced transcription factors activated by the all-trans isomer of RA. Retinoic acid binding to RAR results in conformational changes within the ligand binding domain (LBD), promoting interaction with a number of co-regulator complexes to predominantly activate, but sometimes repress, transcription the latter also occurring in the absence of ligand (Rochette-Egly and Germain 2009). The RAR heterodimerizes with the retinoid X receptor (RXRα, β and γ). Although RXR can be bound by the 9-cis isomer of RA (Chambon 1996), this isomer is present in a very limited number of organs (Kane 2012) and given that ligand-bound RAR can effectively promote transcription with an RXR partner lacking bound ligand, it may commonly function in that way.
Retinoic acid regulation of transcription – new retinoic acid response elements
The RAR/RXR heterodimers regulate transcription by binding to DNA elements consisting of two direct repeats of an RGKTCA motif (R = A/G, K = G/T) traditionally viewed to have a spacing of 1, 2 or 5 bp (DR1, DR2 and DR5; reviewed by Bastien and Rochette-Egly 2004). In recent years, however, techniques such as ChIP-chip and ChIP-seq have allowed the large-scale study of DNA sequences bound by RARs. These studies suggest that the earlier described consensus is broadly correct, but also demonstrate the remarkable flexibility of RARs in binding a wide variety of sequences and suggest that the sites bound by RARs are highly cell type-specific. DR5 was identified as the most common RARE in MCF-7 cells (Hua et al. 2009) with a very low frequency of RAR-bound DR2 elements, while DR2 and DR5 were the most RAR-occupied sites in embryonic stem (ES) cells during early RA-induced differentiation (Mahony et al. 2011). However, other similar studies found that many sites occupied by RARs do not fit the canonical DR1, DR2 or DR5 consensus (Delacroix et al. 2010; Moutier et al. 2012). In another study using RA-treated mouse ES cells and F9 cells, DR0 elements were found to be the most common site bound by RARs (Moutier et al. 2012). DR8 was also found to be a common site of RAR binding, occurring with a similar frequency to DR5. Around 50% of DR8 RAREs incorporated a third motif within the other two, forming a compound RARE consisting of overlapping DR2 and DR0 sites. Analysis of these compound RAREs demonstrated that RAR-RXR heterodimers are able to bind the DR0, DR2 and DR8 elements and that DR8 sequences can act as RA-responsive elements, but also suggested that the DR2 and DR0 sites of the compound RAREs are preferentially occupied relative to the DR8. Although preferentially bound, a DR0-type RARE did not confer RA-responsiveness in a reporter assay. Therefore, despite being the most common site of RAR binding, DR0 RAREs do not appear to act as independent transcriptional regulators.
Prior to this analysis, several DR0-type RAREs had been described. In the context of the brain, one such DR0 RARE has been identified in the promoter of glial fibrillary acidic protein (GFAP), approximately 2.5 kb upstream of the transcription start site (Asano et al. 2009). RA acted synergistically with leukaemia inhibitory factor (LIF) to induce GFAP expression during differentiation of neural precursor cells to astrocytes, but the effect of RA in the absence of LIF was small, supporting the observation of Moutier et al. (2012) that DR0 RAREs do not act independently. RA increased histone acetylation in the GFAP promoter, facilitating the LIF-induced binding of pSTAT3, and the synergistic effect of RA on LIF could be mimicked using the histone deacetylase inhibitor valproic acid. Other potential DR0-type RAREs have been identified in the promoters of the rat genes encoding oxytocin (Adan et al. 1993) and vasopressin (AVP; Burbach et al. 1993). RAR-occupied DR0/8 elements, but not DR5s, were frequently located along with other conserved motifs within a larger regulatory element (Moutier et al. 2012). Therefore, RARs may act via DR0-type RAREs as enhancers of the action of other transcriptional regulators, perhaps through epigenetic modifications of their binding sites.
Additional actions of retinoic acid – regulation of protein translation and kinase activity
In addition to the canonical role of RARs as transcription factors, it is becoming increasingly apparent that RARs, in particular RARα, can have non-genomic functions. These mechanisms include regulation of protein translation. Cytoplasmic RARα regulates the translation of synaptic proteins, including the α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptor subunit glutamate receptor 1 (GLUR1) and RARα appears to directly bind to sequences in the 5′ UTR of the GLUR1 mRNA (Poon and Chen 2008; Chen et al. 2008; reviewed in Chen et al. 2012). RARα interacts with mRNAs via its LBD and the binding of RA disrupts the interaction and relieves repression of translation. Most recently, evidence has been published showing that, through RARα, RA may also non-genomically regulate inhibitory synaptic activity via the control of membrane trafficking of GABA receptors (Sarti et al. 2013).
It is evident from such control of translation that RARs can function in the cytoplasm, in addition to their regulatory role to control transcription in the nucleus. A series of reports have described an additional extranuclear function for RARs in the control of kinase activity that can lead to rapid cellular changes, often within minutes. During neuronal differentiation, RARγ can act in the cytoplasm to regulate neurite outgrowth via a direct interaction with c-SRC (Dey et al. 2007). The RARs can also function in the plasma membrane. RARα directly interacts with phosphatidylinositol-3-kinase in the plasma membrane to activate extracellular signal-regulated kinase 1/2 (ERK1/2) MAPK signalling pathways (Masiá et al. 2007). Alternatively RARα can associate with G protein αq within lipid rafts to activate the p38MAPK/MSK1 pathway (Piskunov and Rochette-Egly 2012). ERK1/2 MAPK pathways are activated in neuronal cells and embryonic stem cells by RA (Bost et al. 2002; Masiá et al. 2007), while in other cells types p38MAPK is activated by RA (Alsayed et al. 2001; Piskunov and Rochette-Egly 2012). This latter pathway can be upstream of activation of MSK1 (Bruck et al. 2009) and provides one mechanism by which a rapid action by RA to promote protein phosphorylation then feeds back to RARs more traditional role of control of transcription. These non-genomic and genomic mechanisms intersect as a result of MSK1′s phosphorylation of RARα, promoting its capacity to induce gene transcription. Overall, these non-genomic pathways likely provide an important route by which RA has rapid control of cellular events such as neurite outgrowth (Dey et al. 2007), growth cone turning (Farrar et al. 2009) and control of neuronal differentiation (Cañón et al. 2004).
Retinoic acid catabolism
The concentration of RA available to activate the RAR/RXR complex and therefore to contribute to retinoid signalling within tissues is determined by the relative expression of both the synthetic and degradative enzymes. Endogenous degradation of RA is primarily via oxidation to a number of polar metabolites including 4-oxo-RA and 4-hydroxy-RA. A number of cytochrome P450 enzymes are able to catalyse this reaction, including CYP2C8, CYP3A4 and CYP2C9 (Marill et al. 2000). However, the high KM values of these reactions mean that their relative contributions to RA metabolism in vivo are likely to be negligible (Gomaa et al. 2011).
The principal CYP450 family responsible for RA degradation in vivo is CYP26. There are three known isoforms encoded by separate genes: CYP26A1, CYP26B1 and CYP26C1. The CYP26s sequentially oxidize RA to more polar metabolites such as 4-hydroxy-RA and 4-oxo-RA using oxygen and nicotinamide adenine dinucleotide phosphate reduced (NADPH) co-factors (McCaffery and Simons 2007; Topletz et al. 2012). In the embryo, the CYP26 enzymes play a role in determining the cellular exposure to RA by inactivating it in cells that do not participate in retinoid signalling, or where RA-activated gene transcription would be detrimental to the cell, for example through the perturbation of the developmental programme (Thatcher and Isoherranen 2009). All three enzymes are expressed during mammalian development in a highly regulated spatio-temporal pattern (Kudoh et al. 2002). During development, CYP26A1 and CYP26B1 are essential for generating an uneven distribution of RA necessary for the development of the hindbrain in the mouse embryo (Abu-Abed et al. 2001). Furthermore, CYP26A1 expression in the prospective head region of the gastrulating mouse embryo is vital for the formation of anterior structures, as RA receptors in this region must be in an unliganded state to allow the proper development of the head (Ribes et al. 2006).
In the adult, CYP26A1 expression is much sparser with only low expression found in the brain (Wang et al. 2002). In contrast, CYP26B1 is considered a brain-specific RA-degrading enzyme with a high level of expression found in the adult cerebellum (Trofimova-Griffin and Juchau 2002) and hippocampus (Abu-Abed et al. 2002). In situ hybridization analysis found at the Allen Brain Atlas database shows an even more widespread distribution including the cortex and amygdala (http://www.brain-map.org/).
Taken together, net RA production is determined by the relative expression of RA synthetic and degradative enzymes in a given tissue. A further element of control is demonstrated by the ability of RA to induce the CYP26 enzymes and therefore limit its own production via a negative feedback pathway (Reijntjes et al. 2005). CYP26A1 is regulated directly by RA, its promoter containing a functional RARE (Loudig et al. 2000, 2005; Hu et al. 2008). This is a key regulatory mechanism necessary to achieve appropriate vitamin A homeostasis.
Retinoic acid in the regulation of circadian and seasonal rhythms
Animals are able to anticipate change in the environment over both the 24-h day and through the changing seasons by the use of a variety of rhythmic systems. The 24-h circadian clock is based on a central rhythm generator in the suprachiasmatic nucleus (SCN), acting as a master clock for the body, although most tissues also contain their own peripheral clock (Dibner et al. 2010). These clocks rely on an internal oscillation network within the cell reliant on a set of feedback loops (Fig.4), but entrained by daylight to keep the rhythm synchronized with environmental day and night. For seasonal rhythms the synchronizing cue is photoperiod, the annual cycle of changing day length. Photoperiodic information is then conveyed to the whole organism through the secretion of melatonin from the pineal gland. Secretion of melatonin at night is a continuous signal, the duration of which reflects the length of the subjective night (Morgan and Hazlerigg 2008).
Fig 4.
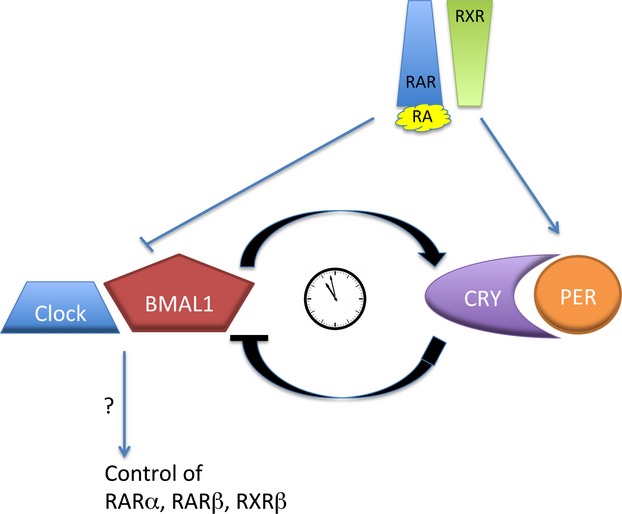
The influence of retinoic acid (RA) on the circadian oscillator. The core set of proteins that create the cellular circadian pacemaker achieve this by forming a positive and a negative feedback loop (Ko and Takahashi 2006). The positive loop involves a heterodimer of CLOCK:BMAL1. These activate transcription of target genes that contain an enhancer box (E-box) sequence, such as PER and CRY. The opposing half of the circuit is the PER and CRY heterodimer which exerts a repressive action on CLOCK:BMAL transcriptional activity. In addition, not shown in the diagram, are multiple interlocked feedback loops that likely both stabilize and provide robustness to the rhythm. These additional loops include pathways that require retinoic acid receptor-related orphan receptors (RORs). RA is reported to influence the central components of the circadian feedback loop by inhibiting the expression or activity of CLOCK/BMAL (McNamara et al. 2001; Shirai et al. 2006) and inducing expression of PER (Shirai et al. 2006). In turn, the genes that encode retinoic acid receptors (RAR)α, β and retinoid X receptor (RXR)β may be regulated by CLOCK/BMAL via their E-box sites (Navigatore-Fonzo et al. 2013).
Both systems are dependent on light: the circadian system is entrained by daylight while photoperiodic information relies on day length. Of course the eye is essential for both of these and, in turn, vitamin A is the key to both eye development and visual function. RA patterns retinal development (McCaffery et al. 1999) while in the photoreceptors retinaldehyde is the chromophore for visual pigment essential for visual transduction via photoreceptors, but is also the chromophore for melanopsin in the intrinsically photosensitive retinal ganglion cells that innervate and signal to the SCN (Palczewski 2012). The evolution of retinoid function as part of this system may have overlapped with the involvement of retinoids in other rhythmic systems in the body and brain. The roles of RA in both circadian and seasonal rhythms are outlined below.
Circadian rhythms in the suprachiasmatic nucleus and hippocampus
RA may influence the body's central clock as high levels of RARα expression are found in the vasopressin-expressing neurons of the human SCN (Meng et al. 2011) and our own preliminary studies show RARα present in processes and cell bodies in the SCN while mRNA transcript levels cycle with a circadian rhythmicity (Fig.3a, b). The function of vasopressin neurons in the SCN was not clear until a recent study demonstrated that mice lacking the V1a/V1b vasopressin receptors are able to adjust rapidly to phase-shifts in the light-dark cycle, making them extremely resistant to jet lag (Yamaguchi et al. 2013) and this was replicated by pharmacological blockade of V1a/b. This indicates that vasopressin signalling in the SCN may stabilize the central circadian clock. In a screen of nuclear receptor regulators of circadian genes RA was found to induce expression of AVP (the vasopressin gene), as well as the clock genes PER1 and PER2, via an enhancer box (E-box)-dependent mechanism (Shirai et al. 2006). PER proteins are central to the cellular circadian rhythm through reciprocal positive and negative feedback loops acting via dimers of CLOCK:BMAL1 and CRY:PER (Fig.4). The results of Shirai et al. also suggested that RA could inhibit expression of CLOCK:BMAL1, although only when RARα was present. This supports an earlier study on blood vessels in which RA could inhibit CLOCK:BMAL1 dimer transcriptional activity (as well as that of MOP4:BMAL1), acting via a RARα:RXRα heterodimer (McNamara et al. 2001). Thus, from these studies, RA may promote CRY:PER and inhibit CLOCK:BMAL1 activity (Fig.4).
Fig 3.
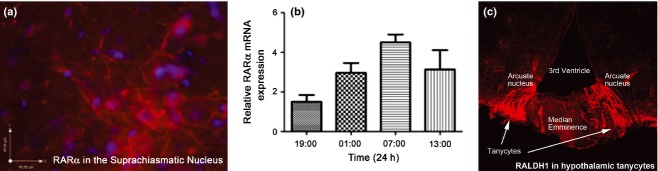
Rhythmic retinoic acid signalling proteins in the hypothalamus. (a) Immunohistochemistry shows that retinoic acid receptor (RAR)α protein is present in both cell bodies and processes in the adult rat suprachiasmatic nucleus. (b) mRNA expression of the encoding gene, RARA, appears to oscillate through the 24-h day in the rat suprachiasmatic nucleus. (c) Retinaldehyde dehydrogenase (RALDH)1, the retinoic acid (RA)-synthesizing enzyme which shows seasonal changes in rat hypothalamic tanycytes, is also strongly expressed in ventral tanycytes in the mouse hypothalamus.
RA may also play a role in the regulation of circadian genes in the hippocampus, given that vitamin A deficiency can phase shift the expression patterns of the clock gene BMAL1 and completely abolishes the circadian expression of PER1 (Golini et al. 2012; Navigatore-Fonzo et al. 2013). Phase-shifting effects of RA are also evident in the capacity of RA to alter rhythmic expression of vascular PER2 both in vitro and in vivo (McNamara et al. 2001). Further implying the involvement of RA in circadian control in the hippocampus is the finding that many of the components of the retinoid signalling pathway show a significant diurnal oscillation (Navigatore-Fonzo et al. 2013). Of the potential downstream elements that RA may control in the hippocampus, the circadian rhythm of brain-derived neurotrophic factor (BDNF) is disrupted in response to vitamin A deficiency (Golini et al. 2012).
The E-box is one of the key enhancers used to control gene expression of several of the circadian genes and is the target of CLOCK:BMAL1. Of the genes required to transduce the RA signal, perfect E-box or E-box-like sites are found in RARA, RARB and RXRB making these genes potential outputs of the circadian cycle (Navigatore-Fonzo et al. 2013). Conversely, RAREs were found in the promoter regions of BMAL1 and PER1 (Golini et al. 2012), suggesting that there may be direct transcriptional regulation of these genes by RA (Navigatore-Fonzo et al. 2013).
Circadian cycles controlled by the pineal gland
The pineal gland receives information about day and night from the retina via the suprachiasmatic nucleus (SCN), the central clock that synchronizes the functions of the body. There is some evidence favouring a role for RA signalling in the generation of circadian rhythms by the mammalian body clock. Nocturnal activity of arylalkylamine N-acetyltransferase, the rate-limiting enzyme in the synthesis of melatonin from serotonin, is greatly reduced in the pineal gland of vitamin A-deficient rats (Herbert and Reiter 1985), although the circadian timing of the nocturnal increase in melatonin appeared unaffected. Expression of RORB, which encodes a retinoid orphan receptor that binds RA with high affinity, is increased at night in the pineal gland (Bailey et al. 2009); genes encoding the retinoid-binding proteins RBP3 and transthyretin (TTR) are also expressed. RA is known to regulate the MAP kinases and the circadian activation of ERK1/2 is lost in the pineal of vitamin A-deficient (VAD) rats (Guillaumond et al. 2005). This function of vitamin A was found to be SCN-dependent, but independent of the superior cervical ganglion (SCG) neurons, raising the possibility that blood-borne nutrients are able to alter the function of the pineal gland.
Seasonal rhythms
Certain species display marked physiological responses to seasonal change in body weight, food intake and reproductive status. These changes are driven by the hypothalamus and the duration of nocturnal melatonin synthesis is the means by which the mammalian hypothalamus determines the season. When the night is short, the reduction in melatonin disinhibits the release of thyroid-stimulating hormone from the pars tuberalis of the pituitary (Ono et al. 2008), up-regulating thyroid hormone signalling in the hypothalamus. Hypothalamic thyroid hormone signalling downstream of melatonin appears to be crucial for seasonal responses in birds (Yoshimura et al. 2003) and mammals (Barrett et al. 2007; Ross et al. 2009).
There is evidence that vitamin A and RA are mediators of seasonal changes in feeding and energy balance though their actions in the hypothalamus. Evidence for this comes from earlier studies showing that vitamin A deficiency in rats causes a decrease in food intake accompanied by a rapid drop in bodyweight which persists even after force-feeding (Anzano et al. 1979). Also, mice lacking the RA synthesizing enzyme RALDH1 are extremely resistant to diet-induced obesity (Ziouzenkova et al. 2007). These two observations suggest a pivotal role for RA in the control of energy balance and more recently, the hypothalamus, the region of the brain controlling growth and feeding, has been recognized as a site of RA action (Ross et al. 2004; Shearer et al. 2010, 2012b; Helfer et al. 2012).
The RA synthetic enzymes RALDH1 and RALDH2 are found in rat tanycytes (RALDH1 only in mice; Shearer et al. 2010 and Fig.3c), cells which line the third ventricle adjacent to the hypothalamus and are thought to be important for the transduction of photoperiodic signals into the hypothalamic nuclei. The tanycytes project RALDH1-containing processes into the arcuate nucleus and the dorsomedial hypothalamic nucleus (Shearer et al. 2012b). RA synthesis by RALDH1/2 in the tanycytes is sensitive to photoperiod with significantly higher levels of RA in the hypothalamus of F344 rats maintained on summer-like long day photoperiods relative to short days (winter; Helfer et al. 2012). Such a seasonal change in RA production has recently been summarized by Shearer et al. (Shearer et al. 2012a). This is in contrast to hypothalamic T3, which shows little change with photoperiod (Ross et al. 2011).
A further indicator of the photoperiodic role of RA in the hypothalamus has been demonstrated in Siberian hamsters and F344 rats in which a long day photoperiod results in large up-regulation of many genes involved in retinoid signalling, including those encoding extracellular and intracellular retinoid-binding proteins (STRA6, TTR, CRABP1, CRABP2, CRBP1), retinoic acid receptors (RAR) and RA-degrading enzymes (CYP26B1; Ross et al. 2004, 2005; Shearer et al. 2010; Helfer et al. 2012). Intriguingly, RPE65, previously thought to be restricted to the retinal pigment epithelium of the eye, is also expressed and photoperiodically regulated in the cells lining the third ventricle in the rat (Helfer et al. 2012). In the retinal pigment epithelium, RPE65 catalyses the isomerization of all-trans retinol to 11-cis retinaldehyde and its expression may be a vestige of photoreceptor-like cells in the mammalian hypothalamus. Deep-brain photoreceptors are present in many non-mammalian species (Vigh et al. 2002) and the periventricular region of the hypothalamus of birds is known to detect light directly (Thayananuphat et al. 2007). These changes suggest a large seasonal up-regulation of RA signalling in the hypothalamus during periods of increased somatic growth and feeding. Many of these changes in gene expression precede physiological changes and, together with the effects of vitamin A deficiency on energy balance (Anzano et al. 1979), this implies a causative role for RA (Ross et al. 2005).
The targets of RA in the hypothalamus are currently not well understood. Some potential targets of RA have been previously discussed (Shearer et al. 2012a). Recently, it has been found that components of the Wnt signalling pathway are photoperiodically regulated in the hypothalamus of F344 rats, with expression of two inhibitors of Wnt signalling, DKK3 and SFRP2, increased in long day photoperiods when hypothalamic RA levels are high (Helfer et al. 2012). Wnt signalling and RA signalling are both involved in adult neurogenesis in the hippocampus (Maden 2007; Inestrosa and Arenas 2010), and there is some evidence that Wnt signalling components are regulated by RA during neurogenesis and neuronal differentiation (Katoh 2002; Jacobs et al. 2006). In F9 teratocarcinoma cells, SFRP2 is specifically up-regulated by RARγ in response to RA (Su and Gudas 2008). In the medial arcuate nucleus, the number of cells expressing nuclear RARγ, suggestive of an increase in transcriptional activity, was increased in rats in long day photoperiods compared to short day (Shearer et al. 2010). Such evidence hints at the Wnt signalling pathway as one mediator of RA's action in the hypothalamus (Helfer et al. 2012).
Vitamin A signalling in sleep regulation
A further biological rhythm in which vitamin A plays a key role is the daily cycle of sleep. Humans spend approximately one-third of their lives sleeping, but the purpose of this vital process remains obscure. Sleep is characterized by measurable differences in brain activity of which slow-wave sleep (SWS, or non-rapid eye movement, NREM, sleep) featuring an electroencephalogram (EEG) delta oscillation of < 2 Hz is most associated with CNS maintenance and repair (Sei 2008). The power of the delta oscillation is a reliable predictor of the homeostatic requirement for sleep after an extended period of wakefulness, and is closely related to the recovery function of sleep (Maret et al. 2005). Indeed, Tononi and Cirelli (2006) have proposed a synaptic homeostasis hypothesis for the purpose of the delta wave in sleep. Briefly, wakefulness and daily activity are both linked to synaptic potentiation in cortical circuits owing to our interaction with the environment leading to increased cortical activation. This synaptic potentiation is associated with the homeostatic regulation of sleep in which increasing length of wakefulness greatly increases the power of the delta oscillation during sleep. When sleeping, we are not interacting with the environment around us and this results in a net downscaling of synaptic strength. Synaptic downscaling could have many benefits to brain function such as reducing the energy expenditure of neural circuits (Atwell and Laughlin 2001), reducing the space required for potentiated synapses (Eyre et al. 2003), and improving the memory function of cortical neurons during further wakeful periods (Huber et al. 2004). There is the intriguing possibility that this synaptic downscaling may be related to the role of RA in the process of homeostatic synaptic plasticity, which provides a homeostatic mechanism of neural circuit adaptation (Chen et al. 2012).
A definitive role for vitamin A signalling however is evident in the regulation of delta oscillations. This was first proposed by Maret (Maret et al. 2005), who observed that the relative contribution of the delta wave to SWS is determined by the RA receptor RARβ1. Moreover, vitamin A deficiency is known to significantly reduce the power of the delta oscillation in mice (Kitaoka et al. 2007), an effect that is reversed by sleep deprivation. Intriguingly, this vitamin A deficiency-induced reduction in delta power is accompanied by decreased dopamine metabolism in the striatum, an observation suggestive of a role for vitamin A in dopaminergic neurotransmission (Kitaoka et al. 2007). Indeed, the RA antagonist LE540 induced a significant reduction in wakefulness and delta oscillation power in mice treated over a period of 4 weeks. This was associated with a reduction in protein expression of the dopamine receptor D1DR in the striatum, and also tyrosine hydroxylase in the hippocampus (Kitaoka et al. 2011). It is possible therefore that retinoids may reduce wakefulness indirectly via RAR interaction with dopamine neurotransmission.
Further clues as to the involvement of vitamin A in the regulation of delta oscillations come from the study of sleep in mouse models of ageing. The SAMP8 senescence-accelerated prone mouse shows age-related deterioration in the sleep/wake cycle in comparison with the related senescence-accelerated resistant mouse (SAMR1; Butterfield and Poon 2005). EEG recordings from SAMP8 mice show a significantly reduced delta power in comparison with that of the SAMR1 mouse, accompanied by increased wakefulness, reduction in NREM sleep and reduction in cortical acetylcholine content (Kitaoka et al. 2010). The expression of several components of the retinoid cascade is reduced in the hippocampus and brainstem of the SAMP8 mouse including a highly significant reduction in the expression of RARα in the hippocampus, suggesting that a reduction in vitamin A signalling may be responsible in part for the decline in sleep observed in this model. Surprisingly, however, the RAR agonist AM80 does not improve delta oscillation power, and no improvement in rapid eye movement (REM) sleep is observed. It is likely therefore that the SAMP8 phenotype is only partially explained by deficiencies in the RAR activation (Kitaoka et al. 2010).
It is thus clear that vitamin A deficiency disrupts SWS through interruption of the delta oscillation in mice. However, given that specific RAR activation is unable to recover sleep abnormalities associated with age, it may be the case that vitamin A regulates sleep indirectly via interaction with dopamine and acetylcholine neurotransmission. Furthermore, vitamin A signalling and the molecular clock show significant cross-regulation in the hippocampus. Given the importance of this brain region to learning and memory, vitamin A may be an important factor in maintaining optimal molecular performance throughout the solar day.
Conclusions
The observations described above make it clear that vitamin A status has an influence on the mammalian biological clock, impacting on the body's response to both the subjective day and night, and also on seasonal regulation of physiology. A key function of circannual rhythms is to anticipate favourable times of year for seasonal behaviours such as reproduction and hibernation, and nutrients are known to inform the clock about the quality and availability of food in the environment. It is clear that vitamin A is a key nutrient in this function.
Acknowledgments
Thanks to Biological Sciences Research Council Grant BB/G014272/1 and BB/K001043/1 for financial support for part of this study.
Glossary
- ARAT
acyl-CoA:retinol acyltransferase
- AVP
arginine vasopressin
- BDNF
Brain-derived neurotrophic factor
- BMAL
brain and muscle ARNT (arylhydrocarbon receptor nuclear translocator)-like protein
- ChIP
chromatin immunoprecipitation
- CLOCK
circadian locomotor output cycles kaput
- CNS
central nervous system
- CRABP
cellular retinoic acid-binding protein
- CRBP
cellular retinol binding protein
- CRY
cryptochrome
- CYP
cytochrome P450
- DHRS
dehydrogenase/reductase (SDR family)
- DKK3
Dickkopf 3
- DR
direct repeat
- EEG
electroencephalogram
- ERK
extracellular signal-regulated kinase
- ES
embryonic stem
- GABA
gamma-aminobutyric acid
- GFAP
glial fibrillary acidic protein
- GLUR1
glutamate receptor 1
- LBD
ligand binding domain
- LIF
leukaemia inhibitory factor
- LRAT
lecithin:retinol acyltransferase
- MAPK
mitogen-activated protein kinase
- MOP
member of PAS protein
- MSK1
mitogen- and stress-activated protein kinase-1
- NADPH
nicotinamide adenine dinucleotide phosphate reduced
- NREM
non-rapid eye movement
- PER
period circadian clock
- pSTAT
phosphorylated Signal transducer and activator of transcription 3
- RA
retinoic acid
- RALDH
retinaldehyde dehydrogenases
- RAR
retinoic acid receptors
- RARE
retinoic acid response element
- RBP
retinol binding protein
- RDHs
retinol dehydrogenases
- REM
rapid eye movement
- ROR
retinoid-related orphan receptor
- RPE65
retinal pigment epithelium-specific protein 65kDa
- RXR
retinoid X receptor
- SCG
superior cervical ganglion
- SCN
suprachaiasmatic nucleus
- SFRP2
Secreted frizzled-related protein 2
- STRA6
stimulated by retinoic acid 6
- SWS
slow-wave sleep
- TTR
transthyretin
- VAD
vitamin A-deficient
References
- Abu-Abed S, Dolle P, Metzger D, Beckett B, Chambon P, Petkovich M. The retinoic acid metabolising enzyme, CYP26A1, is essential for normal hindbrain patterning, vertebral identity, and development of posterior structures. Genes Dev. 2001;15:226–240. doi: 10.1101/gad.855001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Abu-Abed S, MacLean G, Fraulob V, Chambon P, Petkovich M, Dolle P. Differential expression of the retinoic acid metabolising enzymes CYP28A1 and CYP26B1 during murine organogenesis. Mech. Dev. 2002;110:173–177. doi: 10.1016/s0925-4773(01)00572-x. [DOI] [PubMed] [Google Scholar]
- Adan RA, Cox JJ, Beischlag TV, Burbach JP. A composite hormone response element mediates the transactivation of the rat oxytocin gene by different classes of nuclear hormone receptors. Mol. Endocrinol. 1993;7:47–57. doi: 10.1210/mend.7.1.8383287. [DOI] [PubMed] [Google Scholar]
- Alsayed Y, Uddin S, Mahmud N, Lekmine F, Kalvakolanu DV, Minucci S, Bokoch G, Platanias LC. Activation of Rac1 and the p38 mitogen-activated protein kinase pathway in response to all-trans-retinoic acid. J. Biol. Chem. 2001;276:4012–4019. doi: 10.1074/jbc.M007431200. [DOI] [PubMed] [Google Scholar]
- Anzano MA, Lamb AJ, Olson JA. Growth, appetite, sequence of pathological signs and survival following the induction of rapid, synchronous vitamin A deficiency in the rat. J. Nutr. 1979;109:1419–1431. doi: 10.1093/jn/109.8.1419. [DOI] [PubMed] [Google Scholar]
- Asano H, Aonuma M, Sanosaka T, Kohyama J, Namihira M, Nakashima K. Astrocyte differentiation of neural precursor cells is enhanced by retinoic acid through a change in epigenetic modification. Stem Cells. 2009;27:2744–2752. doi: 10.1002/stem.176. [DOI] [PubMed] [Google Scholar]
- Atwell D, Laughlin SB. An energy budget for signalling in the grey matter of the brain. J. Cereb. Blood Flow Metab. 2001;12:1133–1145. doi: 10.1097/00004647-200110000-00001. [DOI] [PubMed] [Google Scholar]
- Bailey MJ, Coon SL, Carter DA, et al. Night/day changes in pineal expression of >600 genes: central role of adrenergic/cAMP signalling. J. Biol. Chem. 2009;284:7606–7622. doi: 10.1074/jbc.M808394200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barrett P, Ebling FJ, Schuhler S, et al. Hypothalamic thyroid hormone catabolism acts as a gatekeeper for the seasonal control of body weight and reproduction. Endocrinology. 2007;148:3608–3617. doi: 10.1210/en.2007-0316. [DOI] [PubMed] [Google Scholar]
- Bastien J, Rochette-Egly C. Nuclear retinoid receptors and the transcription of retinoid target genes. Gene. 2004;328:1–16. doi: 10.1016/j.gene.2003.12.005. [DOI] [PubMed] [Google Scholar]
- Blomhoff R, Green MH, Berg T, Norum KR. Transport and storage of vitamin A. Science. 1990;250:399–404. doi: 10.1126/science.2218545. [DOI] [PubMed] [Google Scholar]
- Bost F, Caron L, Marchetti I, Dani C, Le Marchand-Brustel Y, Binétruy B. Retinoic acid activation of the ERK pathway is required for embryonic stem cell commitment into the adipocyte lineage. Biochem. J. 2002;361:621–627. doi: 10.1042/0264-6021:3610621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bruck N, Vitoux D, Ferry C, Duong V, Bauer A, de Thé H, Rochette-Egly C. A coordinated phosphorylation cascade initiated by p38MAPK/MSK1 directs RARalpha to target promoters. EMBO J. 2009;28:34–47. doi: 10.1038/emboj.2008.256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burbach JPH, Adan RAH, Cox JJ, da Silva SL. Transactivation of the rat oxytocin and vasopressin promoters by nuclear hormone receptors. Regul. Pept. 1993;45:31–35. doi: 10.1016/0167-0115(93)90178-b. [DOI] [PubMed] [Google Scholar]
- Butterfield DA, Poon HF. The senescence-accelerated prone mouse (SAMP8): a model of age related cognitive decline with relevance to alterations of the gene expression and protein abnormalities in Alzheimer's disease. Exp. Gerontol. 2005;40:774–783. doi: 10.1016/j.exger.2005.05.007. [DOI] [PubMed] [Google Scholar]
- Cañón E, Cosgaya JM, Scsucova S, Aranda A. Rapid effects of retinoic acid on CREB and ERK phosphorylation in neuronal cells. Mol. Cell. Biol. 2004;15:5583–5592. doi: 10.1091/mbc.E04-05-0439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chambon P. A decade of molecular biology of retinoic acid receptors. FASEB J. 1996;10:940–954. [PubMed] [Google Scholar]
- Chen N, Onisko B, Napoli JL. The nuclear transcription factor RARalpha associates with neuronal RNA granules and suppresses translation. J Biol. Chem. 2008;283:20841–20847. doi: 10.1074/jbc.M802314200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen L, Lou AG, Sorti F. Synaptic retinoic acid signalling and homeostatic synaptic plasticity. J. Neuropharmacol. 2012 doi: 10.1016/j.neuropharm.2012.12.004. doi: 10.1016/j.neuropharm.2012.12.004 ]Epub ahead of print[ [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crandall JE, Goodman T, McCarthy DM, Duester G, Bhide PG, Dräger UC, McCaffery P. Retinoic acid influences neuronal migration from the ganglionic eminence to the cerebral cortex. J. Neurochem. 2011;119:723–735. doi: 10.1111/j.1471-4159.2011.07471.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delacroix L, Moutier E, Altobelli G, Legras S, Poch O, Choukrallah MA, Bertin I, Jost B, Davidson I. Cell-specific interaction of retinoic acid receptors with target genes in mouse embryonic fibroblasts and embryonic stem cells. Mol. Cell. Biol. 2010;30:231–244. doi: 10.1128/MCB.00756-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dey N, De PK, Wang M, Zhang H, Dobrota EA, Robertson KA, Durden DL. CSK controls retinoic acid receptor (RAR) signalling: a RAR-c-SRC signalling axis is required for neuritogenic differentiation. Mol. Cell. Biol. 2007;27:4179–4197. doi: 10.1128/MCB.01352-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dibner C, Schibler U, Albrecht U. The mammalian circadian timing system: organization and coordination of central andperipheral clocks. Annu. Rev. Physiol. 2010;72:517–549. doi: 10.1146/annurev-physiol-021909-135821. [DOI] [PubMed] [Google Scholar]
- Eyre MD, Richter-Levin G, Avital A, Stuart MG. Morphological changes in hippocampal dentate gyrus synapses following spatial learning in rats are transient. Eur. J. Neurosci. 2003;17:1973–1980. doi: 10.1046/j.1460-9568.2003.02624.x. [DOI] [PubMed] [Google Scholar]
- Farrar NR, Dmetrichuk JM, Carlone RL, Spencer GE. A novel, nongenomic mechanism underlies retinoic acid-induced growth cone turning. J. Neurosci. 2009;29:14136–14142. doi: 10.1523/JNEUROSCI.2921-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fragoso YD, Shearer KD, Sementilli A, de Carvalho LV, McCaffery PJ. High expression of retinoic acid receptors and synthetic enzymes in the human hippocampus. Brain Struct. Funct. 2012;217:473–483. doi: 10.1007/s00429-011-0359-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fu Z, Kato H, Kotera N, Sugahara K, Kubo T. Regulation of the expression of N-acetyltransferase gene in Japanese quail (Coturnix Japonica): II effect of vitamin A deficiency. J. Pineal Res. 1999;27:34–41. doi: 10.1111/j.1600-079x.1999.tb00594.x. [DOI] [PubMed] [Google Scholar]
- Golini RS, Delgado SM, Navigatore Fonzo LS, Ponce IT, Lacoste MG, Anzulovich AC. Daily patterns of clock and cognition-related factors are modified in the hippocampus of vitamin A-deficient rats. Hippocampus. 2012;22:1720–1732. doi: 10.1002/hipo.22007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gomaa S, Bridgens CE, Aboraia AS, Veal GJ, Redfern CPF, Brancale A, Armstrong JL, Simons C. Small molecule inhibitors of retinoic acid 4-hydroxylase (CYP26): synthesis and biological evaluation of imidiazole methyl 3-(4-(aryl-2-ylamino)phenyl) propanoates. Med. Chem. 2011;54:2778–2791. doi: 10.1021/jm101583w. [DOI] [PubMed] [Google Scholar]
- Goodman T, Crandall JE, Nanescu SE, Quadro L, Shearer K, Ross A, McCaffery P. Patterning of retinoic acid signalling and cell proliferation in the hippocampus. Hippocampus. 2012;22:2171–2183. doi: 10.1002/hipo.22037. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guillaumond F, Giraudet F, Becquet D, Sage D, Laforge-Anglade G, Bosler O, François-Bellan AM. Vitamin A is a necessary factor for sympathetic-independent rhythmic activation of mitogen-activated protein kinase in the rat pineal gland. Eur. J. Neurosci. 2005;21:798–802. doi: 10.1111/j.1460-9568.2005.03901.x. [DOI] [PubMed] [Google Scholar]
- Helfer G, Ross AW, Russell L, Thomson LM, Shearer KD, Goodman TH, McCaffery PJ, Morgan PJ. Photoperiod regulates vitamin A and Wnt/β-catenin signalling in F344 rats. Endocrinology. 2012;153:815–824. doi: 10.1210/en.2011-1792. [DOI] [PubMed] [Google Scholar]
- Herbert DC, Reiter RJ. Changes in pineal indoleamine metabolism in vitamin A deficient rats. Life Sci. 1985;37:2515–2522. doi: 10.1016/0024-3205(85)90609-5. [DOI] [PubMed] [Google Scholar]
- Herr FM, Ong DE. Differential interaction of lecithin-retinol acyltransferase with cellular retinol binding proteins. Biochemistry. 1992;31:6748–6755. doi: 10.1021/bi00144a014. [DOI] [PubMed] [Google Scholar]
- Hu P, Tian M, Bao J, Xing G, Gu X, Gao X, Linney E, Zhao Q. Retinoid regulation of the zebrafish cyp26a1 promoter. Dev. Dyn. 2008;237:3798–3808. doi: 10.1002/dvdy.21801. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hua S, Kittler R, White KP. Genomic antagonism between retinoic acid and estrogen signalling in breast cancer. Cell. 2009;137:1259–1271. doi: 10.1016/j.cell.2009.04.043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huber R, Ghilardi MF, Massimini M, Tahran G. Local sleep and learning. Nature. 2004;430:78–81. doi: 10.1038/nature02663. [DOI] [PubMed] [Google Scholar]
- Inestrosa NC, Arenas E. Emerging roles of Wnts in the adult nervous system. Nat. Rev. Neurosci. 2010;11:77–86. doi: 10.1038/nrn2755. [DOI] [PubMed] [Google Scholar]
- Jacobs S, Lie DC, DeCicco KL, Shi Y, DeLuca LM, Gage FH, Evans RM. Retinoic acid is required early during adult neurogenesis in the dentate gyrus. Proc. Natl Acad. Sci. USA. 2006;103:3902–3907. doi: 10.1073/pnas.0511294103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jacobs FM, Smits SM, Noorlander CW, von Oerthel L, van der Linden AJ, Burbach JP, Smidt MP. Retinoic acid counteracts developmental defects in the substantia nigra caused by Pitx3 deficiency. Development. 2007;134:2673–2684. doi: 10.1242/dev.02865. [DOI] [PubMed] [Google Scholar]
- Kane MA. Analysis, occurrence, and function of 9-cis-retinoic acid. Biochim. Biophys. Acta. 2012;1821:10–20. doi: 10.1016/j.bbalip.2011.09.012. [DOI] [PubMed] [Google Scholar]
- Katoh M. Regulation of WNT signalling molecules by retinoic acid during neuronal differentiation in NT2 cells: threshold model of WNT action. Int. J. Mol. Med. 2002;10:683–687. [PubMed] [Google Scholar]
- Kitaoka K, Hattori A, Chikahisa S, Miyamoto K, Nakaya Y, Sei H. Vitamin A deficiency induces a decrease in EEG delta power during sleep in mice. Brain Res. 2007;1150:121–130. doi: 10.1016/j.brainres.2007.02.077. [DOI] [PubMed] [Google Scholar]
- Kitaoka K, Sano A, Chikahisa S, Yoshizaki K, Séi H. Disturbance of rapid eye movement sleep in senescence-accelerated mouse prone/8 mice is improved by retinoic acid receptor agonist Am 80 (Tamibarotene) Neuroscience. 2010;167:573–582. doi: 10.1016/j.neuroscience.2010.01.059. [DOI] [PubMed] [Google Scholar]
- Kitaoka K, Shimizu M, Shimizu N, Chikahisa S, Nakagomi M, Shudo K, Yoshizaki K, Séi H. Retinoic acid receptor antagonist LE540 attenuated wakefulness via the dopamine D1 receptor in mice. Brain Res. 2011;1423:10–16. doi: 10.1016/j.brainres.2011.09.023. [DOI] [PubMed] [Google Scholar]
- Ko CH, Takahashi JS. Molecular components of the mammalian circadian clock. Hum. Mol. Genet. 2006;15:R271–R277. doi: 10.1093/hmg/ddl207. [DOI] [PubMed] [Google Scholar]
- Kudoh T, Wilson SW, David IB. Distinct roles for FGF, Wnt, and retinoic acid in posteriorising the neural ectoderm. Development. 2002;129:4335–4346. doi: 10.1242/dev.129.18.4335. [DOI] [PubMed] [Google Scholar]
- Li H, Wagner E, McCaffery P, Smith D, Andreadis A, Dräger UC. A retinoic acid synthesizing enzyme in ventral retina and telencephalon of the embryonic mouse. Mech. Dev. 2000;95:283–289. doi: 10.1016/s0925-4773(00)00352-x. [DOI] [PubMed] [Google Scholar]
- Loudig O, Babichuk C, White J, Abu-Abed S, Mueller C, Petkovich M. Cytochrome P450RAI(CYP26) promoter: a distinct composite retinoic acid response element underlies the complex regulation of retinoic acid metabolism. Mol. Endocrinol. 2000;14:1483–1497. doi: 10.1210/mend.14.9.0518. [DOI] [PubMed] [Google Scholar]
- Loudig O, Maclean GA, Dore NL, Luu L, Petkovich M. Transcriptional co-operativity between distant retinoic acid response elements in regulation of Cyp26A1 inducibility. Biochem. J. 2005;392:241–248. doi: 10.1042/BJ20050874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maden M. Retinoid signalling in the development of the central nervous system. Nat. Rev. Neurosci. 2002;3:843–853. doi: 10.1038/nrn963. [DOI] [PubMed] [Google Scholar]
- Maden M. Retinoic acid in the development, regeneration and maintenance of the nervous system. Nat. Rev. Neurosci. 2007;8:755–765. doi: 10.1038/nrn2212. [DOI] [PubMed] [Google Scholar]
- Mahony S, Mazzoni EO, McCuine S, Young RA, Wichterle H, Gifford DK. Ligand-dependent dynamics of retinoic acid receptor binding during early neurogenesis. Genome Biol. 2011;12:R2. doi: 10.1186/gb-2011-12-1-r2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maret S, Franken P, Dauvilliers Y, Ghyselinck NB, Chambon P, Tafti M. Retinoic acid signalling affects cortical synchrony during sleep. Science. 2005;310:111–113. doi: 10.1126/science.1117623. [DOI] [PubMed] [Google Scholar]
- Marill J, Cresteil T, Lanotte M, Chabott GG. Identification of human cytochrome P450s involved in the formation of all-trans-retinoic acid principal metabolites. Mol. Pharmacol. 2000;58:1341–1348. doi: 10.1124/mol.58.6.1341. [DOI] [PubMed] [Google Scholar]
- Masiá S, Alvarez S, de Lera AR, Barettino D. Rapid, nongenomic actions of retinoic acid on phosphatidylinositol-3-kinase signalling pathway mediated by the retinoic acid receptor. Mol. Endocrinol. 2007;21:2391–2402. doi: 10.1210/me.2007-0062. [DOI] [PubMed] [Google Scholar]
- McCaffery P, Drager U. High levels of a retinoic acid generating dehydrogenase in the meso-telencephalic dopamine system. Proc. Natl Acad. Sci. USA. 1994;91:7772–7776. doi: 10.1073/pnas.91.16.7772. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCaffery P, Simons C. Prospective Teratology of Retinoic Acid Metabolic Blocking Agents (RAMBAs) and Loss of CYP26 Activity. Curr. Pharm. Des. 2007;13:3020–3037. doi: 10.2174/138161207782110534. [DOI] [PubMed] [Google Scholar]
- McCaffery P, Lee MO, Wagner MA, Sladek NE, Drager UC. Asymmetrical retinoic acid synthesis in the dorsoventral axis of the retina. Development. 1992;115:371–382. doi: 10.1242/dev.115.2.371. [DOI] [PubMed] [Google Scholar]
- McCaffery P, Wagner E, O'Neil J, Petkovich M, Drager UC. Dorsal and ventral retinoic acid territories defined by retinoic acid synthesis, break-down and nuclear receptor expression. Mech. Dev. 1999;85:203–214. doi: 10.1016/s0925-4773(99)00132-x. [DOI] [PubMed] [Google Scholar]
- McCaffery PJ, Zhang J, Crandall JE. Retinoic acid signalling and function in the adult hippocampus. J. Neurobiol. 2006;66:780–791. doi: 10.1002/neu.20237. [DOI] [PubMed] [Google Scholar]
- McNamara P, Seo SB, Rudic RD, Sehgal A, Chakravarti D, FitzGerald GA. Regulation of CLOCK and MOP4 by nuclear hormone receptors in the vasculature: a humoral mechanism to reset a peripheral clock. Cell. 2001;105:877–889. doi: 10.1016/s0092-8674(01)00401-9. [DOI] [PubMed] [Google Scholar]
- Meng QY, Chen XN, Zhao J, Swaab DF, Zhou JN. Distribution of retinoic acid receptor-α immunoreactivity in the human hypothalamus. Neuroscience. 2011;174:132–142. doi: 10.1016/j.neuroscience.2010.11.058. [DOI] [PubMed] [Google Scholar]
- Molotkova N, Molotkov A, Duester G. Role of retinoic acid during forebrain development begins late when RALDH3 generates retinoic acid in the ventral subventricular zone. Dev. Biol. 2007;303:601–610. doi: 10.1016/j.ydbio.2006.11.035. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morgan PJ, Hazlerigg DG. Photoperiodic signalling through the melatonin receptor turns full circle. J. Neuroendocrinol. 2008;20:820–826. doi: 10.1111/j.1365-2826.2008.01724.x. [DOI] [PubMed] [Google Scholar]
- Moutier E, Ye T, Choukrallah M-A, et al. Retinoic acid receptors recognize the mouse genome through binding elements with diverse spacing and topology. J. Biol. Chem. 2012;287:26328–26341. doi: 10.1074/jbc.M112.361790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Napoli JL. Physiological insights into all-trans-retinoic acid biosynthesis. Biochim. Biophys. Acta. 2012;1821:152–167. doi: 10.1016/j.bbalip.2011.05.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Navigatore-Fonzo LS, Golini RL, Ponce IT, Delgado SM, Plateo-Pignatari MG, Gimenez MS, Anzulovich AC. Retinoic acid receptors move in time with the clock in the hippocampus. effect of a vitamin-A-deficient diet. J. Nutr. Biochem. 2013;24:859–867. doi: 10.1016/j.jnutbio.2012.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Newcomer ME, Ong DE. Plasma retinol binding protein: Structure and function of the prototypic lipocalin. Biochim. Biophys. Acta. 2000;1482:57–64. doi: 10.1016/s0167-4838(00)00150-3. [DOI] [PubMed] [Google Scholar]
- Niederreither K, Subbarayan V, Dolle P, Chambon P. Embryonic retinoic acid synthesis is essential for early mouse post-implantation development. Nat. Genet. 1999;21:444–448. doi: 10.1038/7788. [DOI] [PubMed] [Google Scholar]
- Noy N. Retinoid-binding proteins: mediators of retinoid action. Biochem. J. 2000;348:481–495. [PMC free article] [PubMed] [Google Scholar]
- Ono H, Hoshino Y, Yasuo S, Watanabe M, Nakane Y, Murai A, Ebihara S, Korf HW, Yoshimura T. Involvement of thyrotropin in photoperiodic signal transduction in mice. Proc. Natl Acad. Sci. USA. 2008;105:18238–18242. doi: 10.1073/pnas.0808952105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palczewski K. Chemistry and biology of vision. J. Biol. Chem. 2012;287:1612–1619. doi: 10.1074/jbc.R111.301150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Piskunov A, Rochette-Egly C. A retinoic acid receptor RARα pool present in membrane lipid rafts forms complexes with G protein αQ to activate p38MAPK. Oncogene. 2012;31:3333–3345. doi: 10.1038/onc.2011.499. [DOI] [PubMed] [Google Scholar]
- Poon MM, Chen L. Retinoic acid-gated sequence-specific translational control by RARalpha. Proc. Natl Acad. Sci. USA. 2008;105:20303–20308. doi: 10.1073/pnas.0807740105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reijntjes S, Blentic A, Gale E, Maden M. The control of morphogen signalling: regulation of the synthesis and catabolism of retinoic acid in the developing embryo. Dev. Biol. 2005;285:224–237. doi: 10.1016/j.ydbio.2005.06.019. [DOI] [PubMed] [Google Scholar]
- Ribes V, Otto DM, Dickmann L, Schmidt K, Schuhbaur B, Henderson C, Blomhoff R, Wolf CR, Tickle C, Dolle P. Rescue of cytochrome P450 oxidoreductase (por) mutants reveals functions in vasculogenesis, brain, and limb patterning linked to retinoic acid homeostasis. Dev. Biol. 2006;303:66–81. doi: 10.1016/j.ydbio.2006.10.032. [DOI] [PubMed] [Google Scholar]
- Rochette-Egly C, Germain P. Dynamic and combinatorial control of gene expression by nuclear retinoic acid receptors (RARs) Nucl. Recept. Signal. 2009;7:e005. doi: 10.1621/nrs.07005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ronne H, Ocklind C, Wilman K, Rask L, Obrink B, Peterson PA. Ligand-dependent regulation of intracellular protein transport: effect of vitamin A on the secretion of the retinol binding protein. J. Cell Biol. 1983;96:907–910. doi: 10.1083/jcb.96.3.907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ross SA, McCaffery PJ, Drager UC, De Luca LM. Retinoids in embryonal development. Physiol. Rev. 2000;80:1021–1054. doi: 10.1152/physrev.2000.80.3.1021. [DOI] [PubMed] [Google Scholar]
- Ross AW, Webster CA, Mercer JG, Moar KM, Ebling FJ, Schuhler S, Barrett P, Morgan PJ. Photoperiodic regulation of hypothalamic retinoid signalling: association of retinoid X receptor gamma with body weight. Endocrinology. 2004;145:13–20. doi: 10.1210/en.2003-0838. [DOI] [PubMed] [Google Scholar]
- Ross AW, Bell LM, Littlewood PA, Mercer JG, Barrett P, Morgan PJ. Temporal changes in gene expression in the arcuate nucleus precede seasonal responses in adiposity and reproduction. Endocrinology. 2005;146:1940–1947. doi: 10.1210/en.2004-1538. [DOI] [PubMed] [Google Scholar]
- Ross AW, Johnson CE, Bell LM, Reilly L, Duncan JS, Barrett P, Heidemann PD, Morgan PJ. Divergent regulation of hypothalamic neuropeptide Y and agouti-related protein by photoperiod in F344 rats with differential food intake and growth. J. Neuroendocrinol. 2009;21:610–619. doi: 10.1111/j.1365-2826.2009.01878.x. [DOI] [PubMed] [Google Scholar]
- Ross AW, Helfer G, Russell L, Darras VM, Morgan PJ. Thyroid hormone signalling genes are regulated by photoperiod in the hypothalamus of F344 rats. PLoS ONE. 2011;6:e21351. doi: 10.1371/journal.pone.0021351. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sarti F, Zhang Z, Schroeder J, Chen L. Rapid suppression of inhibitory synaptic transmission by retinoic acid. J. Neurosci. 2013;33:11440–11450. doi: 10.1523/JNEUROSCI.1710-13.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sei H. Vitamin A and sleep regulation. J. Med. Invest. 2008;55:1–8. doi: 10.2152/jmi.55.1. [DOI] [PubMed] [Google Scholar]
- Shearer KD, Goodman TH, Ross AW, Reilly L, Morgan PJ, McCaffery PJ. Photoperiodic regulation of retinoic acid signalling in the hypothalamus. J. Neurochem. 2010;112:246–257. doi: 10.1111/j.1471-4159.2009.06455.x. [DOI] [PubMed] [Google Scholar]
- Shearer KD, Stoney PN, Morgan PJ, McCaffery P. A vitamin for the brain. Trends Neurosci. 2012a;35:733–741. doi: 10.1016/j.tins.2012.08.005. [DOI] [PubMed] [Google Scholar]
- Shearer KD, Stoney PN, Nanescu SE, Helfer G, Barrett P, Ross AW, Morgan PJ, McCaffery PJ. Photoperiodic expression of two RALDH enzymes and the regulation of cell proliferation by retinoic acid in the rat hypothalamus. J. Neurochem. 2012b;122:789–799. doi: 10.1111/j.1471-4159.2012.07824.x. [DOI] [PubMed] [Google Scholar]
- Shirai H, Oishi K, Ishida N. Bidirectional CLOCK/BMAL1-dependent circadian gene regulation by retinoic acid in vitro. Biochem. Biophys. Res. Commun. 2006;351:387–391. doi: 10.1016/j.bbrc.2006.10.031. [DOI] [PubMed] [Google Scholar]
- Su D, Gudas LJ. Gene expression profiling elucidates a specific role for RARgamma in the retinoic acid-induced differentiation of F9 teratocarcinoma stem cells. Biochem. Pharmacol. 2008;75:1129–1160. doi: 10.1016/j.bcp.2007.11.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Suzuki R, Shintani T, Sakuta H, Kato A, Ohkanara T, Osumi N, Noda M. Identification of RALDH3, a novel retinaldehyde dehydrogenase expressed in the ventral region of the retina. Mech. Dev. 2000;98:37–50. doi: 10.1016/s0925-4773(00)00450-0. [DOI] [PubMed] [Google Scholar]
- Thatcher JE, Isoherranen N. The role of CYP26 enzymes in retinoic acid clearance. Expert Opin. Drug Metab. Toxicol. 2009;5:875–886. doi: 10.1517/17425250903032681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Thayananuphat A, Kang SW, Bakken T, Millam JR, El Halawani ME. Rhythm-dependent light induction of the c-fos gene in the turkey hypothalamus. J. Neuroendocrinol. 2007;19:407–417. doi: 10.1111/j.1365-2826.2007.01544.x. [DOI] [PubMed] [Google Scholar]
- Tononi G, Cirelli C. Sleep function and synaptic homeostasis. Sleep Med. Rev. 2006;10:49–62. doi: 10.1016/j.smrv.2005.05.002. [DOI] [PubMed] [Google Scholar]
- Topletz AR, Thatcher JE, Zelter A, Lutz JD, Tay S, Nelson WL, Isoherranen N. Comparison of the function and expression of CYP26A1 and CYP26B1, the two retinoic acid hydroxylases. Biochem. Pharmacol. 2012;83:149–163. doi: 10.1016/j.bcp.2011.10.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trofimova-Griffin ME, Juchau MR. Developmental expression of cytochrome CYP26B1 (P450RAI-2) in human cephalic tissues. Brain Res. Dev. Brain Res. 2002;136:175–178. doi: 10.1016/s0165-3806(02)00305-x. [DOI] [PubMed] [Google Scholar]
- Vigh B, Manzano MJ, Zadori A, Frank CL, Lukats A, Rohlich P, Szel A, David C. Nonvisual photoreceptors of the deep brain, pineal organs and retina. Histol. Histopathol. 2002;17:555–590. doi: 10.14670/HH-17.555. [DOI] [PubMed] [Google Scholar]
- Wang Y, Zolfaghari R, Ross AC. Cloning of rat cytochrome P450RAI (CYP26) cDNA and regulation of its gene expression by all-trans-retinoic acid in vivo. Arch. Biochem. Biophys. 2002;401:235–243. doi: 10.1016/S0003-9861(02)00043-7. [DOI] [PubMed] [Google Scholar]
- Yamaguchi Y, Suzuki T, Mizoro Y, et al. Mice genetically deficient in vasopressin V1a and V1b receptors are resistant to jet lag. Science. 2013;342:85–90. doi: 10.1126/science.1238599. [DOI] [PubMed] [Google Scholar]
- Yoshimura T, Yasuo S, Watanabe M, Iigo M, Yamamura T, Hirunagi K, Ebihara S. Light-induced hormone conversion of T4 to T3 regulates photoperiodic response of gonads in birds. Nature. 2003;426:178–181. doi: 10.1038/nature02117. [DOI] [PubMed] [Google Scholar]
- Ziouzenkova O, Orasanu G, Sharlach M, et al. Retinaldehyde represses adipogenesis and diet-induced obesity. Nat. Med. 2007;13:695–702. doi: 10.1038/nm1587. [DOI] [PMC free article] [PubMed] [Google Scholar]