

Abstract Abstract
Palaemon carteri (Gordon, 1935) and Palaemon ivonicus (Holthuis, 1950) are morphologically similar species of South American freshwater shrimps. Past studies have questioned the taxonomic status of both species, which are supposed to have partially sympatric geographic distributions in the Amazon basin. We analyzed a 550 bp fragment of the mitochondrial 16S rRNA gene from these Amazonian Palaemon species as well as from 11 palaemonids as the outgroup. Additionally, we checked diagnostic characters of the genus and family as well as other morphological characters that have been little explored before. Palaemon carteri and Palaemon ivonicus are allocated in two sister lineages, with wide genetic divergence and little morphological differentiation. The divergence time between these lineages was estimated as approximately 10 million years ago. Both molecular and morphological data support the taxonomic validity of both Palaemon carteri and Palaemon ivonicus, refuting the hypothesis of synonymy. In addition, a new species, Palaemon yuna sp. n., closely related to Palaemon ivonicus, is described. Our findings indicate that these species can be differentiated using the projection of the anterolateral margin and anterolateral spine of the first antennular segment, shape of the rostrum, and relative size of the appendix masculina.
Keywords: Amazon basin, divergence time, freshwater shrimp, Palaemon yuna, taxonomy
Introduction
The genus Palaemon Weber, 1795 comprises 84 marine, estuarine and freshwater species in tropical and subtropical regions, including Palaemonetes Heller, 1869, which was recently considered to be a junior synonym of Palaemon by De Grave and Ashelby (2013), and the new species described here. Four strictly freshwater species occur in South America, distributed in the Amazon, Orinoco and Paraguay/lower Paraná River basins: Palaemon carteri (Gordon, 1935), Palaemon ivonicus (Holthuis, 1950) Palaemon mercedae (Pereira, 1986) and a new species described herein.
Palaemon carteri and Palaemon ivonicus are freshwater species with abbreviated larval development (see Magalhães 1986 for larval description of Palaemon ivonicus) and are supposed to occur sympatrically in the Amazon basin as well as being very similar morphologically. They have been distinguished primarily based on rostral characters and on the position of the branchiostegal tooth. Holthuis (1952) stated that Palaemon ivonicus has the lower margin of the rostrum with two or three teeth, the rostrum rather high and straight, and the branchiostegal tooth removed a considerable distance from the anterior margin of the carapace, with its tip failing to reach beyond this margin. On the other hand, Palaemon carteri has the lower margin of the rostrum with four to seven teeth; the rostrum slender, often curved upwards; and the branchiostegal tooth removed a short distance from the anterior margin of the carapace. Nevertheless, some studies have shown that the main characters currently used (position of the branchiostegal tooth, number of rostral teeth, and rostral shape) are not enough to differentiate Palaemon ivonicus from Palaemon carteri consistently (Gomes-Corrêa 1977, Odinetz-Collart and Enriconi 1993, García-Dávila and Magalhães 2003). Therefore, the high interspecific morphological similarity and intraspecific variability found in these species raised doubts as to whether these two nominal species represent distinct biological entities (Gomes-Corrêa 1977, Odinetz-Collart and Enriconi 1993, García-Dávila and Magalhães 2003, García-Dávila et al. 2005).
A multivariate morphometric approach was applied in order to confirm whether the two species constitute separate biological entities in the Amazon basin, and a wide plasticity and overlap in these characters among populations was found (García-Dávila et al. 2005), enhancing the hypothesis of synonymy (Gomes-Corrêa 1977). However, populations from black- and clear-water river systems (Negro and Tapajós river basins, respectively) formed a group slightly distinct from those from white-water river systems (Solimões/Amazon river basin), based on morphometric patterns (García-Dávila et al. 2005; see Sioli 1984 for a characterization of the three Amazonian hydrological systems).
The occurrence of Palaemon ivonicus in the Amazon, Orinoco and Paraguay/lower Paraná basins, however, might indicate a Miocene origin for these lineages. In this period the Orinoco and Amazon basin were widely connected and different sequences of capture of headwater might have resulted in dispersal of species across boundaries of the Amazon and Paraguay basins during the Tertiary (see Lundberg et al. 1998 and Magalhães et al. 2005), which means that these populations could have had enough time to undergo speciation.
Regarding the morphological variability of this group, our study aimed to test the taxonomic status of Palaemon ivonicus and Palaemon carteri, as well as to verify the presence of a new species from the Negro River basin, on the basis of partial sequences of the large ribosomal subunit 16S and morphological analyses.
Methods
Abbreviations and symbols used
CCDB
CL
CNCR
INPA
OUMNH-ZC
MNRJ
MPEG
MV
MZUSP
NHM
USNM
MZUCR
♂: male, ♀: female, ♀ov: ovigerous female.
Sampling
Specimens from several localities were obtained from field collections as well as from visits to and loans from the above-mentioned carcinological collections (Fig. 1). Collected specimens were euthanized on ice, transferred to 96% ethanol, morphotyped under a Leica M205C stereomicroscope, and finally preserved in 80% ethanol for deposit in the CCDB. Specimens from donations and loans were also assigned to morphotypes, and each morphotype was deposited in the CCDB or in the original collection. The collections of species conducted in this study complied with current applicable state and federal laws of Brazil [FLC’s authorization from ICMBio (No. 25329); DIFAP/IBAMA/126/05; and permanent license to FLM for collection of Zoological Material No. 11777-1 MMA/IBAMA].
Figure 1.

Sample sites of Palaemon carteri, Palaemon ivonicus and Palaemon yuna sp. n. c1–Bragança, Pará; c2–Santa Maria do Pará, Pará; c3–National Forest of Amapá, Amapá; c4–Belém, Pará; i1–Solimões River, near Manaus, Amazonas; i2–Xingu River, Altamira, Pará; i3 and i4–Itacoatiara, Amazonas; AC-Acre; AM-Amazonas; AP-Amapá; MS-Mato Grosso do Sul; MT-Mato Grosso; PA-Pará and RO-Rondônia.
Molecular data
DNA extraction, amplification and sequencing protocols followed Schubart et al. (2000) with modifications as in Mantelatto et al. (2007, 2009) and Pileggi and Mantelatto (2010). Total genomic DNA was extracted from the muscle tissue of the abdomen. An approximately 550 bp region of the mitochondrial 16S rRNA gene was amplified from four specimens of Palaemon carteri, four of Palaemon ivonicus, one of Palaemon yuna sp. n. and ten of other palaemonids (Table 1). The amplification was performed by polymerase chain reaction (PCR) in an Applied Biosystems Veriti 96 Well Thermal Cycler® (thermal cycles: initial denaturing for 5 min at 95 °C; pairing for 40 cycles: 45 s at 95 °C, 45 s at 52 °C, 1 min at 72 °C; final extension 5 min at 72 °C) with universal 16S mtDNA primers 1472 (5’-AGATAGAAACCAACCTGG-3’) (Crandall and Fitzpatrick 1996) and 16S-L2 (5’-TGCCTGTTTATCAAAAACAT-3’) (Schubart et al. 2002). PCR products were purified using Sure Clean (Bioline) and sequenced with the ABI Big Dye® Terminator Mix (Applied Biosystems, Carlsbad, CA) in an ABI 3730 XL DNA Analyzer (Applied Biosystems, Foster City, CA) following Applied Biosystems protocols. All sequences were confirmed by sequencing both strands. A consensus sequence for the two strands was obtained using the BIOEDIT software (version 7.0.5) (Hall 2005).
Table 1.
Specimens of Palaemonidae used for the phylogenetic analyses. CCDB: Crustacean Collection of the Department of Biology of the Faculty of Philosophy, Sciences and Letters at Ribeirão Preto, University of São Paulo; CNCR: National Crustacean Collection of the Instituto de Biología, Universidad Nacional Autónoma de México; MPEG: Museu Paraense Emilio Goeldi; MV: Museum Victoria; MZUCR: Museo de Zoología, Universidad de Costa Rica.
| Taxon | Locality | Collection accession number | GenBank |
|---|---|---|---|
| Creaseria morleyi (Creaser, 1936) | Yucatan Peninsula, Mexico | --- | EU448997 |
| Palaemon carteri (Gordon, 1935) | (c1) Jequiri, Bragança, Pará, Brazil | MPEG 0787 | KF923721 |
| Palaemon carteri (Gordon, 1935) | (c2) Santa Maria do Pará, Pará, Brazil | CCDB 4339 | KF923720 |
| Palaemon carteri (Gordon, 1935) | (c3) Japim stream, National Forest of Amapá, Amapá, Brazil | MPEG 1108 | KF923727 |
| Palaemon carteri (Gordon, 1935) | (c4) Mocambo, Belém, Pará, Brazil | MPEG 0628 | KF923730 |
| Palaemon gracilis (Smith, 1871) | Pacific coast, Costa Rica | CCDB 3402 | KF923714 |
| Palaemon hancocki Holthuis, 1950 | Golfo Dulce, Puntarenas, Pacific coast, Costa Rica | MZUCR 2477-02 | KF923715 |
| Palaemon intermedius (Stimpson, 1860) | Victoria, Australia | MV J60843 | KF923725 |
| Palaemon ivonicus (Holthuis, 1950) | (i1) Solimões River, Manaus, Amazonas, Brazil | CCDB 1435 | KF923717 |
| Palaemon ivonicus (Holthuis, 1950) | (i2) Xingu River, Altamira, Pará, Brazil | MPEG 0715 | KF923726 |
| Palaemon ivonicus (Holthuis, 1950) | (i3) Poranga, Itacoatiara, Amazonas, Brazil | CCDB 4632 | KF923728 |
| Palaemon ivonicus (Holthuis, 1950) | (i4) Itacoatiara, Amazonas, Brazil | CCDB 4716 | KF923729 |
| Palaemon kadiakensis (Rathbun, 1902) | Convent, Louisiana, USA | CCDB 1600 | KF923718 |
| Palaemon longirostris H. Milne Edwards, 1837 | Guadiana River, Portugal | CCDB 2750 | KF923724 |
| Palaemon northropi (Rankin, 1898) | Mamanguape River, Paraíba, Brazil | CCDB 4332 | KF923722 |
| Palaemon pandaliformis (Stimpson, 1871) | Ilha Comprida, São Paulo, Brazil | CCDB 813 | KF923713 |
| Palaemon pugio (Holthuis, 1949) | River Delta, Gautier, Mississippi, USA | CCDB 3804 | KF923723 |
| Palaemon ritteri Holmes, 1895 | Bahía Wafer, Puntarenas, Pacific coast, Costa Rica | MZUCR 2396-04 | KF923719 |
| Palaemon suttkusi (Smalley, 1964) | Salado River, Zaragoza, Mexico | CNCR 25864 | KF923712 |
| Palaemon yuna sp. n. | Lago Tupé beach, Negro River, Manaus, Amazonas, Brazil | CCDB 4866 | KF923716 |
Phylogenetic analysis
Sequence alignments were conducted in MAFFT alignment software (version 7.058) with default settings (Katoh and Standley 2013). Estimates of uncorrected genetic divergence (p-distance) for sequence pairs were conducted in MEGA (version 5.2.2) (Tamura et al. 2011), and statistical selection of models of nucleotide substitution in jModelTest (version 2.1.4) (Darriba et al. 2012). The gene dataset was tested for nucleotide substitution saturation using the test by Xia et al. (2003) implemented in DAMBE v. 5.3.48, which revealed no significant saturation (P < 0.05) for symmetrical trees.
We performed four phylogenetic reconstructions: two by Bayesian inference, one by maximum likelihood and one by parsimony. All analyses were conducted using Creaseria morleyi (Creaser, 1936) as the outgroup. A consensus tree of the two Bayesian and maximum likelihood analyses was constructed by 50% majority-rule in the Mesquite package (2.75, build 566). Only posterior probabilities and bootstrap values above 50% are shown. All other software settings not mentioned below were maintained as default.
The Bayesian inferences were performed in the MrBayes software (version 3.2.2) (Ronquist et al. 2012). The Metropolis-coupled Markov chain Monte Carlo was used to empirically determine the posterior probability distribution of trees, branch lengths and substitution parameters. The nucleotide substitution model assumed was the 4by4 with general time-reversibility, gamma-distributed rate variation across sites and invariable sites (GTR+Γ+I). Five gamma rate categories were used. The prior probability distributions on the parameters of the model were maintained as default. Aiming to evaluate possible effects of overfitting, we also performed a second Bayesian inference using a simpler model (Hasegawa-Kishino-Yano with gamma-distributed rates: HKY+Γ) indicated by the Bayesian information criterion. In this second analysis, the parameter values given by jModelTest (nucleotide frequencies, transition/transversion ratio, shape of the gamma distribution and proportion of variable sites) were used to define the prior probability distributions on the parameters of the model. Both analyses were carried out with 107 generations in two independent runs, with one cold and four heated parallel chains. The parameter values were saved once every 1,000 rounds. The runs were stopped if they had reached stationarity (average standard deviation of the split frequencies below 0.01). The first quarter of parameters and trees was discarded (burn-in of 25%); see Ronquist et al. (2009) for further details.
The maximum likelihood (ML) analysis was conducted in the RAxML program (7.6.3) (Stamatakis 2006) implemented in CIPRES (“Cyberinfrastructure for Phylogenetic Research”; http://www.phylo.org). The consistency of topologies was measured by the bootstrap method. The number of bootstrap replicates (650) as well as the proportion of invariable sites was defined by RAxML.
The maximum parsimony analysis was performed using the branch-and-bound algorithm in PAUP (version 4.0 for Unix/Linux). We conducted a bootstrap analysis with 1,000 replicates. Gaps, missing and ambiguous characters were excluded in this analysis.
Molecular dating
The likelihood ratio test (LRT) was used to test the null hypothesis that the data evolved under a molecular clock. The likelihoods for the molecular trees, under the GTR+Γ+I model, with and without an imposed molecular clock were compared using the LRT calculator in jModelTest. The assumption of a molecular clock could not be rejected (P = 0.51). We used both a strict molecular clock and a relaxed molecular clock with independent gamma rates (IGR) in MrBayes, under the settings mentioned above, in order to estimate an approximate divergence time between the Palaemon carteri and Palaemon ivonicus / Palaemon yuna lineages. We assumed the rate of 0.0083 substitutions per site per million years to corrected divergence values for the 16S rRNA gene. This rate was estimated for neotropical palaemonid species of Palaemon, assuming that the isolation of the transisthmian estuarine sibling species Palaemon ritteri Holmes, 1895 and Palaemon paivai Fausto Filho, 1967 (CCDB 4334) occurred about three million years ago (Carvalho et al. unpublished data), as demonstrated for other estuarine carideans (Knowlton and Weigt 1998, Hurt et al. 2009). This rate is also compatible with rates estimated for other decapods (0.006–0.009) by Schubart et al. (2000) for the same gene.
Morphological analysis
Once the phylogenetic relationships based on molecular data were known, we analyzed adult morphological characters of South American species of Palaemon in order to verify the morphological support for the clades obtained, as well as provide new diagnoses for the Palaemon carteri / Palaemon ivonicus / Palaemon yuna group.
We checked out the diagnostic characters that were traditionally used to differentiate Palaemon carteri from Palaemon ivonicus: position of the branchiostegal tooth, number of rostral teeth, and rostral shape (Holthuis 1952, Melo 2003). We also analyzed other characters that were traditionally used in Palaemonidae, as well as others that have been little explored in the taxonomy of the group, such as the projection of the anterolateral margin of the first antennular segment, the anterolateral spine of the first antennular segment, and the relative size of the appendix masculina.
The search for morphological differences among species was conducted using the optimized comparison method described below. Initially, all pairwise differences found between the specimens from each clade in the tree were listed. Those differences with more than 80% constancy were chosen for further validation, using the remaining specimens from the same lots used in the molecular analyses. The consistency of each difference was evaluated step-by-step, analyzing blocks of 10 specimens from each clade. Characters with less than 80% constancy were discarded and no longer analyzed in the following blocks. Additional blocks of specimens from lots that were not included in the molecular analyses were used for a final check.
The diagnostic characters found were verified in the original descriptions (Gordon 1935, Holthuis 1950a, 1952) as well as in the type series of Palaemon carteri and Palaemon ivonicus in order to confirm the taxonomic entity of each clade.
Results
The two Bayesian analyses, maximum likelihood and parsimony analyses indicated the same topology. The mean standard deviation of the split frequencies after 107 generations was less than 0.003 for both GTR+Γ+I and HKY+Γ models. No important differences were found between the two Bayesian inferences using each model, since only slight differences were noted in the posterior probabilities of the clades and branch lengths, indicating that there is no perceptible effect of model overfitting. Similarly, only dissimilarities in the values of node support were observed between the maximum likelihood and the Bayesian inferences, with the GTR+Γ+I implemented in both analyses.
Of the 435 aligned positions (after exclusion of gaps, missing and ambiguous characters), 193 sites were variable, of which 54 (28%) were parsimony-informative. The parsimony reconstruction also showed more than 50% bootstrap values for all clades indicated by the previous analyses, with high support for most of them.
The monophyly of the Palaemon carteri / Palaemon ivonicus / Palaemon yuna group was indicated in all analyses, although this clade had weak bootstrap support in the parsimony and maximum-likelihood methods. All analyses supported the existence of two sister lineages for the specimens of this group (Fig. 2). The “carteri” lineage allocated specimens from eastern Amazonia, assigned to Palaemon carteri. The “ivonicus/yuna” lineage comprised specimens from central and western Amazonia, which were assigned to Palaemon ivonicus and Palaemon yuna sp. n. The population from the black water Negro River, assigned to Palaemon yuna sp. n. (Fig. 3), clearly fell outside the clade that comprised specimens of Palaemon ivonicus from other regions of the Amazon basins (with white and clear water).
Figure 2.

Bayesian (GTR+Γ+I and HKY+Γ models) and maximum likelihood 50% majority-rule consensus tree. Numbers in the nodes represent posterior probabilities (GTR+Γ+I and HKY+Γ, respectively), and bootstrap value for maximum likelihood and parsimony analyses, respectively. c1–Bragança, Pará; c2–Santa Maria do Pará, Pará; c3–National Forest of Amapá, Amapá; c4–Belém, Pará; i1–Solimões River, near Manaus, Amazonas; i2–Xingu River, Altamira, Pará; i3 and i4–Itacoatiara, Amazonas. MYBP–million years before present.
Figure 3.

Palaemon yuna sp. n. Holotype, male, CCDB 4865, habitus, lateral view.
The wide genetic dissimilarity between the “carteri” and “ivonicus/yuna” lineages (from 10.6% to 13.7%), compared among the representative members of the genus used here (from 3.5% to 23.0%; data not shown), also supports the hypothesis that Palaemon carteri and Palaemon ivonicus are valid species, as well as clearly genetically divergent (Fig. 4). The intralineage genetic variability was 0–3.6% in “carteri” and 0–4.3% in “ivonicus/yuna”. The interspecific dissimilarity between Palaemon ivonicus and Palaemon yuna was 4.1–4.3%.
Figure 4.
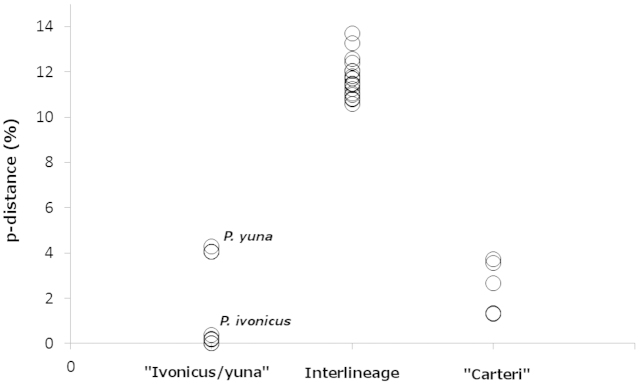
Intralineage and interlineage uncorrected genetic distance values for the “ivonicus/yuna” and “carteri” lineages.
The divergence time between the “carteri” and “ivonicus/yuna” lineages, based on the 16S rRNA gene, was estimated as approximately 10 million years ago. This mean value varied between ~9 and ~11 Ma in our analysis, depending on the molecular-clock model used, as well as other parameter settings such as node constraints. The 95% credible intervals (highest posterior density–HDP) were 5.5–14.3 Ma using a relaxed clock, and 8.2–14.0 Ma using a strict clock.
Morphological characters supported these lineages. The morphological analysis (including the type series of all three species) of 122 specimens of Palaemon carteri, 333 specimens of Palaemon ivonicus and 125 specimens of Palaemon yuna sp. n. indicated that the projection of the anterolateral margin and the anterolateral spine of the first antennular segment were useful characters to differentiate these lineages, as long as adult specimens were considered. The “carteri” lineage had the projection of the anterolateral margin of the first antennular segment slightly shorter, not reaching the dorsal distal margin of the second segment. Additionally, the anterolateral spine of the first antennular segment usually reached the middle of the projection of the anterolateral margin (Fig. 5a). On the other hand, the “ivonicus/yuna” lineage had the projection of the anterolateral margin of the first antennular segment longer, reaching the dorsal distal margin of the second segment. Moreover, the anterolateral spine of the first antennular segment did not reach the middle of the projection of the anterolateral margin (Fig. 5b–c).
Figure 5.

Commonest shape of the antennular peduncle. Palaemon carteri (a MPEG 787), Palaemon ivonicus (b INPA 128) and Palaemon yuna sp. n. (c CCDB 4866).
The rostral characters (shape, relative size and number of ventral teeth of the rostrum) were helpful to differentiate Palaemon ivonicus from Palaemon yuna sp. n. (Fig. 6). Palaemon ivonicus had the rostrum high, straight or slightly curved upward, not overreaching the scaphocerite; the ventral margin had one to four teeth, usually three or fewer. On the other hand, Palaemon yuna sp. n. had the rostrum slightly curved upward, overreaching the scaphocerite; the ventral margin has two to five teeth, usually three or four. The relative length of the appendix masculina was also useful to distinguish adult individuals of the “ivonicus/yuna” lineage, since the appendix masculina reached up to 1.5 times the length of the appendix interna in Palaemon ivonicus and up to 1.1 times in Palaemon yuna sp. n.
Figure 6.

Commonest shape of the rostrum. Palaemon carteri (a MPEG 787), Palaemon ivonicus (b INPA 128) and Palaemon yuna sp. n. (c CCDB 4866).
Once the validity of Palaemon ivonicus and the new species for the Negro River basin were corroborated, we provide illustrations, diagnoses, identification key as well as a description of Palaemon yuna sp. n. in order to differentiate the three species from each other as well as from other South American species of Palaemon.
Palaemon carteri
(Gordon, 1935)
Palaemonetes carteri Gordon, 1935: 324, fig. 12;–Holthuis 1948: 113;–Holthuis 1950b: 32;–Holthuis 1966: 6 [part, spec. from Rio Tapajós];–Rodríguez 1980: 126;–Rodríguez 1981: 47 [in list];–Rodríguez 1982: 390;–Coelho and Ramos-Porto 1985: 408 [in table];–Holthuis 1993: 8;–Delgado et al. 1997: 16 [in list];–Ramos-Porto and Coelho 1998: 337 [in list];–Barros and Pimentel 2001: 20 [in list];–Vieira 2003: 61;–Magalhães and Pereira 2007: 9, 10, 12 [in list]; Mora-Day et al. 2009: 196 [in list];–Pereira et al. 2010a: 606 [in list];–Pereira et al. 2010b: 84 [in list];–Pileggi et al. 2013: 569 [part, material from Amapá and Bragança; ? material from rio Tapajós basin].
Palaemonetes (Palaemonetes) carteri –Holthuis 1950c: 10 [in list];–Holthuis 1952: 218, pl. 52, figs c-o, pl. 53, figs a–c;–Holthuis 1959: 81, text-fig. 9;–Kensley and Walker 1982: 11 [part, ? spec. from Rio Curua Una];–López and Pereira 1996: 54, fig. 8;–López and Pereira 1998: 77 [in list]; Melo 2003: 382 [part, not Amazonas].
Holotype.
Guyana, upper Cuyuni River, ♂, col. GS Carter (NHM 1935.5.20.19).
Paratypes.
Karow Creek, 2 m NE of Penal Settlement, Mazaruni, 1 ♂; River Cuyuni, 1♀ov; Forest Swamp, upper Cuyuni, 1♂; same data as holotype, 7♂ 3♀ 1♀ov 1 juvenile (NHM 1935.5.20.20-29).
Other material.
Suriname. Nickerie. Lower Naui Kreek, Southern Niew, 10♂ 10♀, col. DC Geijskes, 18 March 1971 (INPA 176). Brazil. Amapá. Floresta Nacional do Amapá, igarapé Japim, 5♂ 5♀, col. CRM Santos and JEM Nanzelor, 27 October 2009 (MPEG 1108); Porto Grande, Floresta Nacional do Amapá, tributary of rio Araguari, 1♂ 4♀, col. CRM Santos, 28 October 2009 (CCDB 2755); Macapá, stream in the home of Sr. Marcondes, 1♂ 4♀ov, col. J Cunha, 6 March 2005 (MZUSP 17676). Pará. Barcarena, Vila do Conde, 5♂ 2♀ 3♀ov, col. B Mascarenhas, 23 March 2002 (MPEG 739); Belém, Mocambo, 5♂ 5♀ov, col. FR Pimentel and R Maia, January 9 1998 (MPEG 528); Belém, Mocambo, Reserva Mocambo, 4♂ 6♀, col. FR Pimentel and J Dias, 18 June 1999 (MPEG 628); Bragança, Jequeri, Sítio Anacuã, 7♂ 9♀, 23 October 2002 (MPEG 787); Ilha do Marajó, cachoeira do Arari, 1♀, col. J Cunha and J Zuanon, 16 May 2008 (MZUSP 22753); Castanhal, 1♀ov, col. FL Carvalho et al., 14 December 2012 (CCDB 4338); Ilha de Marajó, cachoeira do Arari, rio Arari, igarapé Popudas, 1♂ 1♀, col. J Cunha and J Zuanon, 17 May 2006 (MZUSP 23224); Laranjal do Jari, igarapé Arapiranga, 2♀ 1♀ov, col. Moreira et al., 25 March 2008 (MZUSP 23225); Melgaço, Floresta Nacional de Caxiuanã, 5♂ 4♀, 8 November 1999 (MPEG 717); Santa Maria do Pará, 3 juveniles, col. FL Carvalho et al., 15 December 2012 (CCDB 4339); Tucuruí, rio Tocantins basin, igarapé Santos, 2♀, col. W Zuink and LCF Alvarenga, 16 September 1984 (MNRJ 23382).
Diagnosis.
Mandibular palp absent. Rostrum slender, curved upward, reaching or just overreaching the tip of the scaphocerite; dorsal margin with 6 to 10 teeth; ventral margin with 3 to 7 teeth, usually 4 or more. Projection of the anterolateral margin of the first antennular segment overreaching the middle of the second segment, but not reaching, sometimes almost reaching, the dorsal distal margin of the second segment; anterolateral spine of the first antennular segment usually reaches the middle of the projection of the anterolateral margin. Appendix masculina up to 1.1 the length of the appendix interna, measured from their junction. Telson carrying 2 plumose setae between the inner distal stout setae; inner distal stout setae overreaching the distal tip of the telson.
Geographic distribution.
Venezuela (Amazonas, Bolívar, Delta Amacuro, Monagas), Guyana, Suriname, French Guiana, Brazil (eastern Amazon: Amapá, Pará).
Ecological features.
Usually associated with riparian vegetation, leaf litter and similar microhabitats in lakes, streams and rivers, in areas with low flow. At least in the Amazon river basin, its occurrence is usually associated to clear water river systems.
Palaemon ivonicus
(Holthuis, 1950)
Palaemonetes ivonicus Holthuis, 1950a: 98;–Holthuis 1966: 4, fig. 1;–Rodríguez 1981: 47 [in list];–Coelho and Ramos-Porto 1985: 408 [in table];–Ramos-Porto and Coelho 1990: 99 [part, not Rio Negro, Catagalo];–Kochalka et al. 1996: 113 [in list];–Ramos-Porto and Coelho 1998: 337 [in list]; Magalhães 1999: 36, 85 [in list];–Magalhães 2000: 59 [in list];–Magalhães 2001: 70, 133 [in list];–Magalhães 2002: 1096, figs 5, 6;–García-Dávila and Magalhães 2003: 675, figs 21–27, 55;–Magalhães and Pereira 2007: 9, 10, 12 [in list];–Valencia and Campos 2010: 224, figs 3–4;–Pileggi et al. 2013: 570.
Palaemonetes carteri –Holthuis 1966: 6 [part, ? 2 spec. from Lago Redondo];–Magalhães 2005: 69, 71 [in list]; Pileggi et al. 2013: 569 [? part, material from rio Xingu and rio Trombetas].
Palaemonetes (Palaemonetes) ivonicus –Holthuis 1950c: 10 [in list];–Holthuis 1952: 222, pl. 53, figs d–h;–Melo 2003: 382.
Palaemonetes (Palaemonetes) carteri –Kensley and Walker 1982: 11 [part., ? spec. from Rio Madeira].
Holotype.
Bolivia, Beni, Ivon, Beni River, ♀ (CL 7.5 mm), col. WM Mann, February 1922 (USNM 85234).
Paratype.
same data as holotype, 1♀ (CL 6.6 mm) (USNM 85234).
Other material.
Brazil. Acre. Bujari, igarapé Mapinguari, 2♂ 18♀, col. LR Malabarba et al., 8 August 2001 (UFRGS 3179). Amazonas. Itacoatiara, canal Irandiba, 3♂, col. GY Hattori, April 2008 (CCDB 2753); Itacoatiara, igarapé Aeroporto, 2♂ 18♀ (CCDB 4725); Itacoatiara, Poranga, 7♂ 23♀ (CCDB 4632); Itacoatiara, 3♂ 15♀ (CCDB 4716); rio Madeira, Borba, 1♂ 9♀ (MNRJ 1078); rio Solimões, igarapé do Xiboreno, 1♂ 1♀, col. FL Carvalho and EA Souza-Carvalho, 28 January 2012 (CCDB 1435); rio Solimões, lago Janauacá, 1♀, col. J Donnath, 18 March 1978 (MZUSP 8183); rio Solimões, lago do Jacaré, 2♀, col. H Reichardt, 29 March 1967 (MZUSP 6405); Tefé, igarapé da Aeronáutica, 12♂ 8♀, col. JO Chaves, 21 March 1979 (INPA 128). Pará. Almeirim, rio Arraiolos, pesqueiro São Paulo, 7♂ 5♀, col. J Carvalho Júnior, 26 July 1999 (MPEG 689); Altamira, rio Xingu, 4♀, col. RM Sousa and Dionísio, 18 December 2000 (MPEG 715); Porto de Moz, rio Xingu, 1♂ 1♀ov, col. R Robles et al., 25 September 2013 (CCDB 4867); rio Tapajós, near the rio Cupari’s mouth, downstream Itaiatuba, 1♀, col. C Magalhães and LH Py-Daniel, 27 October 1991 (INPA 1176); Santarém, igarapé do Juá, 2♀, col. LM Sousa and JL Birindelli, 13 November 2006 (MZUSP 28358). Rondônia. Rio Guaporé, 1♂ 2♀, col. JC Malta, 25 September 1985 (INPA 326). Mato Grosso. Acorizal, 17♂ 6♀, col. Sebastiana, 28 August 1987 (MNRJ 1151); Acorizal, 9♂ 5♀ 4 juveniles, col. Sebastiana (MNRJ 1153); baía do Pio, Pantanal, 5♂ 15 juveniles, col. Sebastiana (MNRJ 1152); Poconé, baía do Pio, 14♂ 33♀ 19 juveniles (INPA 328). Mato Grosso do Sul. Rio Negro, córrego Anhumas, 9♂ 11♀, col. C Magalhães et al., 28 August 1998 (CCDB 4667). Peru. Loreto. Lago Urcococha, rio Amazonas, 10♂ 5♀, col. C García-Dávila, 10 January 1999 (INPA 883); Quistococha, río Itaya, 12♂ 7♀ov, col. C García-Dávila, 13 July 1998 (INPA 882).
Diagnosis.
Mandibular palp absent. Rostrum high, straight or slightly curved upward, not overreaching the scaphocerite; dorsal margin with 6 to 10 teeth; ventral margin with 1 to 4 teeth, usually 3 or fewer. Projection of the anterolateral margin of the first antennular segment reaching or overreaching the dorsal distal margin of the second segment; anterolateral spine of the first antennular segment almost reaching or overreaching the first third of the projection of the anterolateral margin. Appendix masculina up to 1.5 the length of the appendix interna, measured from their junction. Telson carrying 2 plumose setae between the inner distal stout setae; inner distal stout setae overreaching the distal tip of the telson.
Geographic distribution.
Venezuela? (Delta Amacuro, Monagas), Colombia? (Amazonas, Arauca, Casanare, Guainía, Meta, Vichada), Brazil (Acre, Amazonas, Mato Grosso, Mato Grosso do Sul, Pará), Bolívia (Beni, Cochabamba, Pando), Peru (Loreto, Madre de Díos), Paraguay.
Ecological features.
Usually associated with riparian vegetation, leaf litter and similar microhabitats in lakes, streams and rivers with white or clear water, in areas with low flow. In the western portion of the Amazon river basin it is commonly found in the floodplains of the white water river systems.
Palaemon yuna sp. n.
http://zoobank.org/1541C001-E4DD-4812-9D60-277706CE391D
Figure 7.

Palaemon yuna sp. n. Figure a holotype; figures b–n paratype (CCDB 4866, male, CL 5.5 mm). a anterior part of the carapace b right eye, dorsal view c left scaphocerite, ventral view d left mandible, ventral view e left maxillula, ventral view f left maxilla, ventral view g left second maxilliped, ventral view h left first maxilliped, ventral view i left first maxilliped, dorsal view j right third maxilliped, ventro-lateral view k right second pereiopod, ventro-lateral view l right first pereiopod, ventro-lateral view m right first chela, mesial view n right second chela, mesial view. Scale bar: a, c, k equal to 1 mm; others equal to 0.5 mm.
Figure 8.

Palaemon yuna sp. n. Figures i and m holotype; figures a–e, g, h, j–l paratype (CCDB 4866, male, CL 5.5 mm); figure f paratype (CCDB 4866, female, CL 5.5 mm). a left third pereiopod, lateral view b left fourth pereiopod, lateral view c distal portion of the left fifth pereiopod, lateral view d left fifth pereiopod, lateral view e left first pleopod, posterior view f left first pleopod, posterior view g left second pleopod, posterior view h left appendix masculina and appendix interna, posterior view i right posterior part of the abdomen, lateral view j pre-anal fig, ventral view k telson and uropods, dorsal view l distal part of the telson, dorsal view m left distal portion of the exopod of the uropod, dorsal view. Scale bar: a, b, d–g, i, k equal to 1 mm; c, j, m equal to 0.5 mm; h, l equal to 0.25 mm.
Palaemonetes (Palaemonetes) carteri –Kensley and Walker 1982: 11, figs 13–14 [part, at least spec. from Rio Negro and its basin]; Melo 2003: 382 [part, Amazonas].
Palaemonetes carteri –Ramos-Porto and Coelho 1990: 99 [Rio Negro, Cantagalo; igarapé afluente do Rio Mapiri, Santarém?];–Pileggi et al. 2013: 569 [? part, material from rio Tapajós basin, rio Xingu and rio Trombetas].
Palaemonetes ivonicus –Ramos-Porto and Coelho 1990: 99 [part, Rio Negro, Cantagalo].
Holotype.
Lago Tupé beach, lower Rio Negro tributary, Manaus, Amazonas, Brazil (003°02'42"S, 060°15'10"W), ♂, col. FL Carvalho and EA Souza-Carvalho, 27 January 2012 (CCDB 4865).
Paratypes.
same data as holotype, 28♂ 8♀ 17♀ov (CCDB 4866); same data as holotype, 10♂ 3♀ 3♀ov (INPA 2016); same data as holotype, 1♂ 1♀ 1♀ov (OUMNH-ZC 2013-08-001).
Other material.
Brazil. Amazonas. Parque Nacional de Anavilhanas, lake near the rio Apuaí’s mouth, 1♂ 1♀, col. J Zuanon, 20 August 2005 (INPA 1432); Manaus, Rio Negro basin, Igarapé do Camarão, 20♀ov, col. O Odinetz-Collart et al., 28 February 1989 (CCDB 4726); Rio Negro basin, igarapé Alagadiço, 20♂, col. O Odinetz-Collart et al., 17 January 1989 (CCDB 4727); São Gabriel da Cachoeira, igarapé Barixia, right bank of the Rio Negro, 4♀, col. J Cunha et al., 14 December 2005 (MZUSP 16907); Santa Izabel do Rio Negro, 2♀, 24 October 1972 (MZUSP 13645); rio Uatumã, near the igarapé do Miriti’s mouth, 1♂ 3♀, col. C Magalhães, 12 July 1985 (INPA 173).
Diagnosis.
Mandibular palp absent. Rostrum slender, slightly curved upward at the distal half, overreaching the scaphocerite; dorsal margin with 6 to 10 teeth; ventral margin with 2 to 5 teeth, usually 3 or 4. Projection of the anterolateral margin of the first antennular segment reaching the dorsal distal margin of the second segment; anterolateral spine of the first antennular segment not overreaching the first third of the projection of the anterolateral margin. Appendix masculina up to 1.1 the length of the appendix interna, measured from their junction. Telson carrying 2 plumose setae between the inner distal stout setae; inner distal stout setae overreaching the distal tip of the telson.
Description.
Carapace glabrous. Sub-orbital lobe and pterygostomial angle rounded. Branchiostegal suture located approximately with a half of the distance between the antennal and branchiostegal tooth. Branchiostegal tooth almost as strong as the antennal, placed behind the anterior margin of the carapace.
Rostrum slender, slightly curved upward at the distal half, overreaching the scaphocerite (Fig. 7a); dorsal margin with 6 to 10 teeth, 5 to 8 of them placed in the proximal 2/3, 1 or 2 dorsal teeth located behind the orbit, 1 or 2 subapical teeth; ventral margin with 2 to 5 teeth, usually 3 or 4, all of them placed in the distal half. Single row of setae present on the proximal ventral portion up to the second tooth; double continuous and uniformly spaced row of setae on the distal half.
Eye well developed with pigmented cornea (Fig. 7b); cornea slightly wider and smaller than the eyestalk; ocellus present on dorsal side.
Antennular peduncle not reaching the distal margin of the scaphocerite; first antenular segment with outer margin slightly convex and projection of the anterolateral margin rounded, reaching the dorsal distal margin of the second segment (Fig. 5c); anterolateral spine of the first antennular segment not overreaching the first third of the projection of the anterolateral margin; second segment as broad as and shorter than the third segment; inner ventromesial tooth present; upper antennular flagellum fused for about 1/5 of its length (4–6 segments fused, 14–16 free); free portion with two rows of two or three aesthetascs on each segment. Stylocerite short, not reaching the middle of the first antennular segment. Béc ocellaire with anterior margin concave, pronounced upwardly and bearing a pigmented spot dorsally.
Scaphocerite slender (Fig. 7c), laminar, 3.3 times as long as broad; outer margin slightly concave, terminating in a tooth, not overreaching the lamella; basal segment of antenna with strong lateral tooth. Flagellum of the antenna more than five times the length of the body.
Mandibular palp absent; incisor process with three teeth on both sides (Fig. 7d). Upper lacina of the maxillula just reaching the length of the inner lacina (Fig. 7e). First maxilliped with lobes of the epipod fused (Fig. 7h–i); anterior lobe elongated and almost triangular, with lateral border slightly convex; junction between the endites roundly curved. Epipod of the third maxilliped with anterior margin forming an angle of about 45° (Fig. 7j).
Thoracic sternal armature sexually similar. First thoracic sternite with an acute tooth and a conspicuous transverse ridge; second without tooth, bearing a triangular transverse ridge. Third to fifth without tooth and ridge incomplete.
First pereiopod slender (Fig. 7l), reaching the tip of scaphocerite with the fingers; ischium 1.8 times the length of basis; merus 1.7 times the length of ischium; carpus 1.3 times the length of merus; chela slightly less than 0.5 the length of carpus; fingers as long as palm (Fig. 7m).
Second pereiopod slender (Fig. 7k), overreaching the scaphocerite with about a half of the inflated distal part of the carpus; ischium about 4.5 times the length of basis; merus 0.8 the length of ischium; carpus 1.8 times the length of merus; chela about 0.6 times the length of carpus; fingers about 0.7 the length of palm (Fig. 7n).
Third pereiopod (Fig. 8a) slender, reaching the tip of the scaphocerite; ischium 1.8 times the length of basis; merus 1.9 times the length of ischium; carpus about 0.5 times the length of merus; propodus 1.6 times the length of carpus, ventral margin armed with 5–8 cuspidate setae; dactylus simple, about 0.3 times the length of the propodus.
Fourth pereiopod slender (Fig. 8b), overreaching the scaphocerite with all length of the dactylus; ischium 1.7 times the length of basis; merus 2.4 times the length of ischium; carpus about 0.5 length of merus; propodus 1.7 times the length of carpus, ventral margin armed with 7-13 cuspidate setae; dactylus simple, about 0.3 the length of propodus.
Fifth pereiopod slender (Fig. 8c–d), overreaching the scaphocerite with the end of the propodus; ischium 1.7 times the length of basis; merus 2.3 times the length of ischium; carpus about 0.6 length of merus; propodus 2.1 times the length of carpus; grooming brush comprises about 10 rows of setae on the distal third of the propodus; dactylus simple, about 0.2 the length of propodus.
First pleopod without appendix interna and sexually dimorphic in proportions; males with endopod 0.5 the length of exopod (Fig. 8e); females with endopod approximately 0.3 length of exopod (Fig. 8f). Second to fifth pleopods similar, with the endopod reaching about 0.8 the length of exopod and bearing an appendix interna (Fig. 8g). Appendix masculina up to 1.1 times the length of the appendix interna, measured from their junction (Fig. 8h).
Abdominal sternal armature sexually dimorphic; males with first and second sternites bearing median process; second more acute and bigger than the first process; females with median process less develop than males.
Abdominal pleura furnished with plumose setae on ventral margin; fifth pleuron elongated and disto-ventrally rounded (Fig. 8i), with dorsal posterior border concave; sixth segment 1.63 times the length of the fifth; posterolateral margin with small tooth and keel disto-ventrally. Anal fig unarmed (Fig. 8j).
Telson as long as sixth pleonite; dorsal surface with two pairs of cuspidate setae (Fig. 8k); proximal dorsal tuft of setae reduced to one or two simple setae; marginal setae absent; posterior margin ending abruptly in a triangular tip, not overreaching the inner stout setae (Fig. 8l); 1 pair of plumose setae and 2 pairs of stout setae, inner pair of stout setae about 2.3 times the length of the outer pair.
Uropods overreaching the telson by 0.3 of the length of exopod; exopod 1.25 times the length of endopod; mobile distolateral setae of exopod weak, reaching about the middle of the fixed tooth (Fig. 8m).
Geographic distribution.
Brazil (Amazonas, Pará?), Venezuela? (Apure).
Ecological features.
Usually associated with riparian vegetation, leaf litter and similar microhabitats in lakes, streams and rivers of black or clear water river systems, in areas with low flow.
Etymology.
The specific epithet is derived from the Tupi, the general language of the Brazilian indigenous people: y = water, river + úna = black, alluding to the environment where the species was first found (Fig. 9).
Figure 9.

Type locality of Palaemon yuna sp. n. Lago Tupé beach, lower Rio Negro tributary, Manaus, Amazonas, Brazil (003°02'42"S, 060°15'10"W).
Key for the species of the Palaemon carteri / Palaemon ivonicus / Palaemon yuna sp. n. group
| 1 | Projection of the anterolateral margin of the first antennular segment not overreaching the middle of the second segment | other South American Palaemoninae |
| – | Projection of the anterolateral margin of the first antennular segment overreaching the middle of the second segment | 2 |
| 2 | Mandibular palp present | other South American Palaemoninae |
| – | Mandibular palp absent | 3 |
| 3 | Projection of the anterolateral margin of the first antennular segment not reaching, rarely almost reaching, the dorsal distal margin of the second segment; anterolateral spine of the first antennular segment usually reaching the middle of the projection of the anterolateral margin (Fig. 5a); rostrum with 3 to 7 ventral teeth (usually 4 or more) | Palaemon carteri |
| – | Projection of the anterolateral margin of the first antennular segment reaching or overreaching the dorsal distal margin of the second segment; anterolateral spine of the first antennular segment not reaching the middle of the projection of the anterolateral margin (Fig. 5b–c); rostrum with 1 to 5 ventral teeth (usually 4 or fewer) | 4 |
| 4 | Rostrum high, straight or slightly curved upward, not overreaching the scaphocerite; rostrum with 1 to 4 ventral teeth (usually 3 or fewer) (Fig. 6b). Anterolateral spine of the first antennular segment generally reaching the first third of more of the projection of the anterolateral margin (Fig. 5b) | Palaemon ivonicus |
| – | Rostrum slender, slightly curved upward, overreaching the scaphocerite; rostrum with 2 to 5 ventral teeth (usually 3 or 4) (Fig. 6c). Anterolateral spine of the first antennular segment generally not reaching the first third of the projection of the anterolateral margin (Fig. 5c) | Palaemon yuna sp. n. |
Discussion
Both molecular and morphological data support the validity of Palaemon ivonicus, refuting the hypothesis that this species is a junior synonym of Palaemon carteri. The two species are allocated in two sisters Amazonian lineages, with great genetic divergence and morphological support. Additionally, a third species closely related to Palaemon ivonicus is described.
The projection of the anterolateral margin and anterolateral spine of the first antennular segment seems to be an important character in this group, although we have found some specimens with a state of character close to an intermediate form, making difficult a clear distinction between these lineages. Additionally, there is an ontogenetic variation, which needs to be considered in the analyses. Some large specimens of the “carteri” lineage have the antennular projection almost reaching the dorsal margin of the second segment. On the other hand, some small specimens of the “ivonicus/yuna” lineage also have the projection almost reaching the dorsal margin of the second segment. However, even considering those limitations, the antennular character was the most constant one to distinguish the “carteri” and “ivonicus/yuna” lineages. Therefore, we suggest that this character must be evaluated in further morphological analyses within the Palaemon genus.
The wide intraspecific morphological variability and interspecific similarity between Palaemon carteri and Palaemon ivonicus as well as the presumptive synonymy have been reported by previous studies since the 1970s (Gomes-Corrêa 1977, Odinetz-Collart and Enriconi 1993, García-Dávila and Magalhães 2003, García-Dávila et al. 2005). Despite of that similarity posteriorly reported, Holthuis (1950a, 1952) did not make a direct comparison of Palaemon carteri and Palaemon ivonicus in order to clearly differentiate them. Furthermore, the type series of Palaemon ivonicus is composed by only two specimens, making it impossible to evaluate the morphological variability spectrum within this species. Therefore, it is not surprising that misidentification may occur, which may have contributed to the hypothesis of synonymy.
The overlapping of several morphological characters demonstrated by Odinetz-Collart and Enriconi (1993) and García-Dávila et al. (2005) probably is due to the fact that all or most of the specimens examined by these authors were from the “ivonicus/yuna” lineage. These studies used the traditional rostral characters to identify some specimens from the central Amazon basin as Palaemon carteri. We were unable to find any sample of Palaemon carteri in the central and western Amazon basin, since specimens from these regions are more likely to be assigned to the “ivonicus/yuna” lineage. Therefore, the rostral similarity between Palaemon carteri and Palaemon yuna sp. n. probably has led many authors to use the name Palaemon carteri, which was the name available at the moment that better fitted the rostral characteristics of some populations from the central Amazon basin (see synonymic list of Palaemon yuna sp. n.).
The wide genetic dissimilarity between the “carteri” and “ivonicus/yuna” lineages shows that they have no recent divergence, as one could expect based only on their morphological similarity. The approximately 10 million years of divergence estimated for these lineages, based on the 16S rRNA gene, may be associated with marine incursion as well as colonization of different environments in western Amazonia during the Middle Miocene (~16 to 11.6 Ma) and Late Miocene (~11.6 to 5.3 Ma) (Fig. 10). From the Early Miocene until the early Late Miocene (~23 to 9 Ma), the western Amazonia region was mostly submerged and transformed into a continually shifting mosaic of lakes, wetlands and river belts (Wesselingh 2006), which are similar to the environments where specimens of the “ivonicus/yuna” lineage are currently found. This continually shifting mosaic, the Pebas system, had contact with the Caribbean Sea and underwent several marine incursions during the Middle and Late Miocene (Hoorn 1993, Wesselingh 2006). Particularly in the period between 11.8 and 10 Ma, close to our estimate for the time of divergence between the “carteri” and “ivonicus/yuna” lineages, there is evidence of an extensive marine transgression into the low-lying basins of South America (Lundberg et al. 1998). These marine incursions, which established brackish-water conditions in the late Middle Miocene and early Late Miocene (Hoorn 1993, Lundberg et al. 1998), might have isolated freshwater lineages of the Pebas system from others distributed in northern South America outside this wetland system, where the “carteri” lineage is currently found (Fig. 11). Similar biogeographical patterns and speciation events associated with the Pebas system and marine incursions in the Middle and Late Miocene have been reported for several groups in the Amazon region (Hoorn 1993, Lovejoy et al. 1998, 2006, Lundberg et al. 1998, Wesselingh 2006, Santos et al. 2009, Cooke et al. 2012a,b). Therefore, the proposed current parapatric distribution of the “carteri” and “ivonicus/yuna” lineages probably developed after the establishment of the modern west-to-east course of the Amazon River, which may have initiated approximately 8 Ma (Lundberg et al. 1998).
Figure 10.
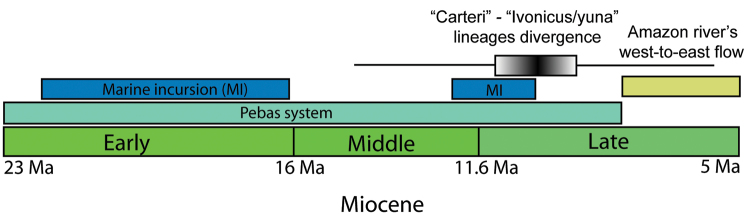
Historical context for the proposed divergence time between the “carteri” and “ivonicus/yuna” lineages.
Figure 11.

Putative current distribution of the “carteri” and “ivonicus/yuna” lineages. [Pebas system during the Late Miocene (~11.8 to 10 Ma) according to Lundberg et al. 1998.]
The specimens from the Negro River (Palaemon yuna sp. n.) have considerable genetic divergence from the specimens collected in the Solimões-Amazon and Xingu rivers, being allocated outside the Palaemon ivonicus sensu stricto clade. The specimens from the Negro River also show some differences in the rostrum, in the antennular characters and in the appendix masculina compared to specimens from the type locality of Palaemon ivonicus as well as other basins (Figs 5–6). Additionally, a morphometric study with specimens from the Negro River revealed that they form a morphometrically distinct group from populations that inhabit white water, although some overlapping was found (García-Dávila et al. 2005). Studies with fishes reported a similar pattern, providing evidence for the effect of divergent natural selection associated with the difference in water colour between the Negro River and Solimões-Amazon River (Cooke et al. 2012a,b). Genetic divergence between hydrologically different, but interconnected, environments has been reported for other palaemonid species without a conspicuous and consistent morphological differentiation (Carvalho et al. 2013). However, the characters described above allow the morphological distinction between Palaemon ivonicus and Palaemon yuna sp. n. Therefore, regarding the genetic, morphological and ecological differentiation, there is sufficient evidence to justify the proposal of a new species for the populations from the Negro River basin. The actual distributional range of Palaemon yuna sp. n. may be wider than indicated by the material available for this study. Specimens from the lower Tapajós River basin might be co-specific with the specimens from the Negro River basin, as suggested by the results of a molecular study conducted by García-Dávila (2002) in an unpublished academic thesis, using the mitochondrial cytochrome c oxidase subunit I gene (COI).
Specimens from the upper Orinoco River basin have antennular characters similar to those of Palaemon yuna sp. n. Nevertheless, some specimens fail to have a curved upward rostrum overreaching the scaphocerite. The connection between the Negro and Orinoco river basins through the Casiquiare River opens the possibility of the existence of a conspecific group occurring in the Negro and Orinoco rivers. A similar biogeographical pattern has been reported for fish species, using molecular data (Willis et al. 2010). However, despite logistic difficulties, a more thorough genetic and morphological sampling along the Negro River and the upper Orinoco River basins should be carried out in order to verify the phylogenetic relationships among the populations from the upper Orinoco and other populations of the “ivonicus/yuna” lineage as well as its taxonomic status. Similarly, the records of Palaemon ivonicus from the Orinoco River basin (López and Pereira 1996, 1998, Pereira et al. 2010a, b) should be verified, since morphologically (judged by the illustration provided by López and Pereira 1996: 55, fig. 9) and zoogeographically these specimens probably do not belong to this taxon.
The occurrence of Palaemon ivonicus in the Paraguay/lower Paraná River basin is an issue that needs further analyses in order to verify the phylogenetic relationships of these populations. As discussed by Magalhães et al. (2005), the decapod fauna of the Amazon and the Paraguay-Paraná river lowlands has several common elements whose current distributions may be result of dispersal across the paleobasins of these systems during Tertiary and Quaternary as some geological events changed their boundaries promoting different sequences of capture of headwater (Lundberg et al. 1998). Even in recent times, sporadic or seasonal contact between the Amazon and the Paraguay-Paraná fluvial nets can occur, as some landscape features along their boundary favor transfluences, headwater captures, floods and spillouts to one or other side (Iriondo and Paira 2007).
An additional record of Palaemon ivonicus from the São Francisco River basin, state of Minas Gerais, Brazil (Melo 2003), cannot be confirmed as it was not documented by voucher material, and we were unable to find any samples from this basin in the collections visited by us. In addition, we could not find any individual of this species in field collections made in that state. Moreover, the geological history of the São Francisco River basin does not show any evidence of a connection with the Amazonian basin during the Neogene or Quaternary period, which makes unlikely a natural occurrence of Palaemon ivonicus in this basin. The report of Palaemon carteri for Mexico (Felder et al. 2009) seems to be an error, since the notes associated to the species do not match with the distribution and habitat of the species, and no other studies have reported this species for the Gulf of Mexico.
The fact that our target group seems to be closer related to three neotropical species of Palaemon (Palaemon gracilis, Palaemon hancocki and Palaemon pandaliformis) than to other species of the genus require further studies. Ashelby et al. (2012) recovered Palaemon gracilis and Palaemon pandaliformis outside the clade that comprised the majority of the species of Palaemon. Therefore, Palaemon carteri, Palaemon ivonicus and Palaemon yuna might also not have a close relationship with Palaemon sensu stricto. However, as this was not the goal of the present study, a broader taxonomic sampling of the subfamily using both mitochondrial and nuclear genes is needed to address properly these generic-level questions.
Our study is part of a project aiming to investigate the American species of Palaemon, and this is the first one which uses a multidisciplinary approach aiming to clarify this taxonomic issue. Our data clearly show that there are at least two morphologically and genetically distinct lineages, which might have diverged ~10 Ma. A multilocus approach is needed to provide more molecular support for this estimated divergence time. The possibility of hybridization cannot be rejected and must be deeply investigated in further studies. Moreover, the morphological variability found in some populations still assigned to Palaemon ivonicus as well as the molecular variability found within the “carteri” lineage need to be further investigated to verify whether there are other morphologically similar species not yet described.
Supplementary Material
Acknowledgements
The present study is part of a long-term project to evaluate the taxonomy of freshwater decapods in Brazil, and was supported by scientific grants provided to FLM by the São Paulo Research Foundation–FAPESP (2002/08178-9, 2009/54931-0 and 2010/50188-8) and the Conselho Nacional de Desenvolvimento Científico e Técnológico–CNPq (472746/2004-9, 491490/2004-6, 473050/2007-2, 471011/2011-8, 504322/2012-5; Research Scholarships PQ 301261/2004-0, 301359/2007-5, 302748/2010-5), and to CM by CNPq (Research Scholarships PQ 304468/2009-6, 303837/2012-8). FLC is supported by PhD fellowship from CNPq (140199/2011-0) and “Sandwich” PhD fellowship from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior–CAPES (7711-13-1). We are grateful to the Department of Biology and Postgraduate Program in Comparative Biology of the FFCLRP/USP and Instituto Nacional de Pesquisa da Amazônia–INPA for partial financial and logistic support, to the Xingu Project funded by the U.S. National Science Foundation (DEB-1257813) and CNPq (Proc. N° 1300.001303/2011-2) for providing some specimens from the Xingu River, and to many colleagues and friends (Cléverson Santos, Darryl Felder, Edinaldo Nelson dos Santos-Silva, Edvanda Souza-Carvalho, Emmanoela Ferreira, Fernando Alvarez, Gary Poore, Ingo Wehrtmann, Irene Cardoso, José Luis Villalobos, José Montoya, Joana Palma, Marcos Tavares, Mariana Terossi, Natalia Rossi, Rafael Robles, Ray Bauer, Tatiana Magalhães) for helping in collections, making available some essential specimens and information, lending material from collections used in our research, and for critical discussion during the preparation of this manuscript. Special thanks to all members of LBSC for their assistance during the development of this study, to Dr. Janet Reid (JWR Associates) for providing the English review service, to anonymous reviewers and Ingo Wehrtmann for all comments and suggestions.
Citation
Carvalho FL, Magalhães C, Mantelatto FL (2014) Molecular and morphological differentiation between two Miocene-divergent lineages of Amazonian shrimps, with the description of a new species (Decapoda, Palaemonidae, Palaemon). In: Wehrtmann IS, Bauer RT (Eds) Proceedings of the Summer Meeting of the Crustacean Society and the Latin American Association of Carcinology, Costa Rica, July 2013. ZooKeys 457: 79–108. doi: 10.3897/zookeys.457.6771
References
- Ashelby CW, Page TJ, De Grave S, Hughes JM, Johnson ML. (2012) Regional scale speciation reveals multiple invasions of freshwater in Palaemoninae (Decapoda). Zoologica Scripta 41(3): 293–306. doi: 10.1111/j.1463-6409.2012.00535.x [Google Scholar]
- Barros MP, Pimentel FR. (2001) Decapoda (Crustácea) do estado do Para, Brasil: lista preliminar das espécies. Boletim do Museu Paraense Emílio Goeldi Boletim do Museu Paraense Emílio Goeldi, série Zoologia17: 15–41. [Google Scholar]
- Carvalho FL, Pileggi LG, Mantelatto FL. (2013) Molecular data raise the possibility of cryptic species in the Brazilian endemic prawn Macrobrachium potiuna (Decapoda, Palaemonidae). Latin American Journal of Aquatic Research 41(4): 707–717. doi: 10.3856/vol41-issue4-fulltext-7 [Google Scholar]
- Coelho PA, Ramos-Porto M. (1985) Camarões de água doce do Brasil: distribuição geográfica. Revista Brasileira de Zoologia 2(6): 405–410. doi: 10.1590/S0101-81751984000200014 [Google Scholar]
- Cooke GM, Chao NL, Beheregaray LB. (2012a) Natural selection in the water: freshwater invasion and adaptation by water colour in the Amazonian pufferfish. Journal of Evolutionary Biology 25: 1305–1320. doi: 10.1111/j.1420-9101.2012.02514.x [DOI] [PubMed] [Google Scholar]
- Cooke GM, Chao NL, Beheregaray LB. (2012b) Five Cryptic Species in the Amazonian catfish Centromochlus existimatus identified based on biogeographic predictions and genetic data. PLoS ONE 7(11): . doi: 10.1371/journal.pone.0048800 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crandall KA, Fitzpatrick JF. (1996) Crayfish molecular systematics: using a combination of procedures to estimate phylogeny. Systematic Biology 45: 1–26. [Google Scholar]
- Darriba D, Taboada GL, Doallo R, Posada D. (2012) jModelTest 2: more models, new heuristics and parallel computing. Nature Methods 9(8): . doi: 10.1038/nmeth.2109 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delgado JG, Severeyn HJ, Godoy AR, Reverol YM, Ewald JJ. (1997) Camarones dulceacuícolas y estuarinos de Venezuela (Atyidae, Palaemonidae): nuevos registros para los estados Zulia y Falcón. Boletín del Centro de Investigaciones Biológicas 31(1): 11–32. [Google Scholar]
- De Grave S, Ashelby CW. (2013) A re-appraisal of the systematic status of selected genera in Palaemoninae (Crustacea: Decapoda: Palaemonidae). Zootaxa 3734(3): 331–344. doi: 10.11646/zootaxa.3734.3.3 [DOI] [PubMed] [Google Scholar]
- Felder DL, Alvarez F, Goy JW, Lemaitre R. (2009) Decapoda (Crustacea) of the Gulf of Mexico, with comments on the Amphionidacea. In: Felder DL, Camp DK. (Eds) Gulf of Mexico Origin, Waters and Biota Vol. 1, Biodiversity Texas A&M University Press, College Station, 1019–1104. [Google Scholar]
- García-Dávila CR. (2002) Estudos morfométricos e moleculares comparativos das populações de Palaemonetes carteri (Gordon, 1935) e Palaemonetes ivonicus Holthuis, 1950 (Decapoda: Palaemonidae) na Amazônia. PhD thesis, Instituto Nacional de Pesquisas da Amazônia/Universidade Federal do Amazonas, Manaus, Brazil: [Unpublished] [Google Scholar]
- García-Dávila CR, Magalhães C. (2003) Revisão taxonômica de camarões de água doce (Crustacea: Decapoda: Palaemonidae e Sergestidae) da Amazônia peruana. Acta Amazonica 33(4): 663–686. [Google Scholar]
- García-Dávila CR, Magalhães C, Guerrero JCH. (2005) Morphometric variability in populations of Palaemonetes spp. (Crustacea, Decapoda, Palaemonidae) from the Peruvian and Brazilian Amazon Basin. Iheringia, Série Zoologia 95(3): 327–334. doi: 10.1590/S0073-47212005000300013 [Google Scholar]
- Gomes-Corrêa MM. (1977) Palaemonídeos do Brasil (Crustacea–Decapoda–Natantia). MSc thesis, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil: [Unpublished] [Google Scholar]
- Gordon I. (1935) On new or imperfectly known species of Crustacea Macrura. Journal of the Linnean Society of London, Zoology 39(266): 307–351. [Google Scholar]
- Hall T. (2005) BioEdit v.7.0.5. Biological Sequences Alignment Editor for Windows. Ibis Therapeutics, a Division of Isis Pharmaceuticals; http://www.mbio.nesu.edu/bioefit.html [Google Scholar]
- Holthuis LB. (1948) Notes on some Crustacea Decapoda Natantia from Surinam. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen 51: 1104–1113, figs 1–27. [Google Scholar]
- Holthuis LB. (1950a) Preliminary descriptions of twelve new species of palaemonid prawns from American waters (Crustacea Decapoda). Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen 53: 93–99. [Google Scholar]
- Holthuis LB. (1950b) Scientific results of the Surinam Expedition 1948–1949. Part II. Zoology No. 1 Crustacea Decapoda Macrura. Zoologische Mededelingen 31(3): 25–37. [Google Scholar]
- Holthuis LB. (1950c) The Decapoda of the Siboga Expedition. Part X. The Palaemonidae collected by the Siboga and Snellius Expeditions with remarks on other species. I. Subfamily Palaemoninae. Siboga Expéditie 39(A9): 1–268. [Google Scholar]
- Holthuis LB. (1952) A general revision of the Palaemonidae (Crustacea: Decapoda: Natantia) of the Americas, II: The subfamily Palaemoninae. Allan Hancock Foundation Publications 12: 1–396. [Google Scholar]
- Holthuis LB. (1959) The Crustacea Decapoda of Suriname. Zoologische Verhandelingen 44: 1–296, pls. 1–15. [Google Scholar]
- Holthuis LB. (1966) A collection of freshwater prawns (Crustacea Decapoda, Palaemonidae) from Amazonia, Brazil, collected by Dr. G. Marlier. Bulletin de l'Institut Royal des Sciences Naturelles de Belgique 42: 1–11. [Google Scholar]
- Holthuis LB. (1993) The decapod Crustacea of Brokopondo Lake, Suriname. Nauplius 1: 1–12. [Google Scholar]
- Hoorn C. (1993) Marine incursions and the influence of Andean tectonics on the Miocene depositional history of northwestern Amazonia: results of a palynostratigraphic study. Palaeogeography, Palaeoclimatology, Palaeoecology 105: 267–309. doi: 10.1016/0031-0182(93)90087-Y [Google Scholar]
- Hurt C, Anker A, Knowlton N. (2009) A multilocus test of simultaneous divergence across the isthmus of Panama using snapping shrimp in the genus Alpheus. Evolution 63: 514–530. doi: 10.1111/j.1558-5646.2008.00566.x [DOI] [PubMed] [Google Scholar]
- Iriondo MH, Paira AR. (2007) Physical Geography of the Basin. In: Iriondo MH, Paggi JC, Parma MJ. (Eds) The Middle Paraná River: Limnology of a Subtropical Wetland. Springer-Verlag, Heidelberg, 7–31. doi: 10.1007/978-3-540-70624-3_1 [Google Scholar]
- Katoh K, Standley DM. (2013) MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Molecular Biology and Evolution 30(4): 772–780. doi: 10.1093/molbev/mst010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kensley B, Walker I. (1982) Palaemonid shrimps from the Amazon basin, Brazil (Crustacea: Decapoda: Natantia). Smithsonian Contributions to Zoology 362: 1–28. [Google Scholar]
- Knowlton N, Weigt LA. (1998) New dates and new rates for divergence across the Isthmus of Panama. Proceedings of the Royal Society of London B 265: 2257–2263. doi: 10.1098/rspb.1998.0568 [Google Scholar]
- Kochalka JA, Torres D, Garcete BR, Aguilar C, et al. (1996) Lista de invertebrados de Paraguay pertenecientes a las colecciones del Museo Nacional de Historia Natural del Paraguay. In: Colecciones de Flora y Fauna del Museo Nacional de Historia Natural del Paraguay. DPNVS, MAG, Asunción, 69–283. [Google Scholar]
- López B, Pereira G. (1996) Inventario de los crustáceos decápodos de las zonas alta y media del delta del Río Orinoco, Venezuela. Acta Biologica Venezolana 16(3): 45–64. [Google Scholar]
- López B, Pereira G. (1998) Actualización del inventario de crustáceos decápodos del Delta del Orinoco. In: López Sánchez JL, Saavedra Cuadra II, Dubois Martínez M. (Eds) El Río Orinoco. Aprovechamiento Sustentable. Universidad Central de Venezuela, Caracas, 76–85 [Memorias de las Primeras Jornadas Venezolanas de Investigación sobre el Rio Orinoco.] [Google Scholar]
- Lovejoy NR, Albert JS, Crampton WGR. (2006) Miocene marine incursions and marine/freshwater transitions: Evidence from neotropical fishes. Journal of South American Earth Sciences, 21: 5–13. doi: 10.1016/j.jsames.2005.07.009 [Google Scholar]
- Lovejoy NR, Bermingham E, Martin AP. (1998) Marine incursion into South America. Nature 396: 421–422. doi: 10.1038/24757 [Google Scholar]
- Lundberg JG, Marshall JG, Guerrero J, Horton B, Malabarba MCSL, Wesslingh F. (1998) The stage for neotropical fish diversification: a history of tropical South American rivers. In: Malabarba LR, Reis RE, Vari RP, Lucena ZMS, Lucena CAS. (Eds) Phylogeny and Classification of Neotropical Fishes. EDIPUCRS, Porto Alegre, 13–48. [Google Scholar]
- Magalhães C. (1986) The larval development of palaemonid shrimps from the Amazon region reared in the laboratory IV. Abbreviated development of Palaemonetes ivonicus Holthuis 1950 Crustacea Decapoda. Amazoniana 10: 63–78. [Google Scholar]
- Magalhães C. (1999) Diversity and abundance of decapod crustaceans in the rio Tahuamanu and rio Manuripi basins. In: Chernoff B, Willink PW. (Eds) A Biological Assessment of the Aquatic Ecosystems of the Upper Río Orthon basin, Pando, Bolivia. Appendix 5. Bulletin of Biological Assessment 15 Conservation International, Washington, D.C., 35–38. [Google Scholar]
- Magalhães C. (2000) Diversity and abundance of decapod crustaceans in the Rio Negro basin, Pantanal, Mato Grosso do Sul, Brazil. In: Chernoff B, Alonso LE, Montambault JR, Lourival R. (Eds) A biological Assessment of the Aquatic Eecosystems of the Pantanal, Mato Grosso do Sul, Brazil. Appendix 5. Bulletin of Biological Assessment 18 Conservation International, Washington, D.C., 56–62. [Google Scholar]
- Magalhães C. (2001) Diversity, distribution, and habitats of the macro-invertebrate fauna of the Río Paraguay and Río Apa, Paraguay, with emphasis on decapod crustaceans. (Appendices 10-11). In: Chernoff B, Willink PW, Montambault JR. (Eds) A Biological Assessment of the Aquatic Ecosystems of the Río Paraguay Basin, Alto Paraguay, Paraguay. Conservation International, Washington, D.C., 68–72. [Google Scholar]
- Magalhães C. (2002) A rapid assessment of the decapod fauna in the Rio Tahuamanu and Rio Manuripi Basins, with new records of shrimps and crabs for Bolivia (Crustacea, Decapoda, Palaemonidae, Sergestidae, Trichodactylidae). Revista Brasileira de Zoologia 19(4): 1091–1103. [Google Scholar]
- Magalhães C. (2005) Macrocrustacean survey of selected localities in the Río Pastaza basin in Ecuador and Perú: diversity, habitat, zoogeographical aspects, and conservation implications. In: Willink PW, Chernoff B, McCullough J. (Eds) A Rapid Biological Assessment of the Aquatic Ecosystems of the Pastaza River Basin, Ecuador and Perú. RAP Bulletin of Biological Assessment, Washington, D.C.33: 67–74. [Google Scholar]
- Magalhães C, Pereira G. (2007) Assessment of the decapod crustacean diversity in the Guayana Shield region aiming at conservation decisions. Biota Neotropica 7: 111–124. doi: 10.1590/S1676-06032007000200013 [Google Scholar]
- Magalhães C, Bueno SLS, Bond-Buckup G, Valenti WC, Silva HLM, Kiyohara F, Mossolin EC, Rocha SS. (2005) Exotic species of freshwater decapod crustaceans in the state of São Paulo, Brazil: records and possible causes of their introduction. Biodiversity and Conservation 14(8): 1929–1945. doi: 10.1007/s10531-004-2123-8 [Google Scholar]
- Mantelatto FL, Robles R, Felder DL. (2007) Molecular phylogeny of the Western Atlantic species of the genus Portunus (Crustacea, Brachyura, Portunidae). Zoological Journal of the Linnean Society of London 150: 211–220. doi: 10.1111/j.1096-3642.2007.00298.x [Google Scholar]
- Mantelatto FL, Robles R, Schubart CD, Felder DL. (2009) Molecular phylogeny of the genus Cronius Stimpson, 1860, with reassignment of C. tumidulus and several American species of Portunus to the genus Achelous De Haan, 1833 (Brachyura: Portunidae). In: Martin JW, Crandall KA, Felder DL. (Eds) Crustacean Issues, 18: Decapod Crustacean Phylogenetics. Taylor & Francis/CRC Press, Boca Raton, 567–579. [Google Scholar]
- Melo GAS. (2003) Famílias Atyidae, Palaemonidae e Sergestidae. In: Melo GAS. (Ed.) Manual de Identificação dos Crustáceos Decápodos de Água Doce Brasileiros. Editora Loyola, São Paulo, 289–415. [Google Scholar]
- Mora-Day J, Magalhães C, El Souki M, Blanco-Belmonte L. (2009) Apêndice 6. Lista sistemática de los macroinvertebrados colectados durante el RAP Alto Cuyuni 2008, Estado Bolívar, Venezuela. In: Lasso CA, Señaris JC, Rial A, Flores AL. (Eds) Evaluación Rápida de la Biodiversidad de los Ecosistemas Acuáticos de la Cuenca Alta del Río Cuyuní, Guayana Venezolana. Conservation International, Arlington, 196–197. [Google Scholar]
- Odinetz-Collart O, Enriconi A. (1993) Estratégia reprodutiva e alguns aspectos demográficos do camarão Palaemonetes carteri Gordon, 1935 na Amazônia Central, Rio Negro. Acta Amazonica 23(2-3): 227–243. [Google Scholar]
- Palumbi SR, Benzie J. (1991) Large mitochondrial DNA differences between morphologically similar Penaeid shrimp. Molecular Marine Biology and Biotechnology 1(1): 27–34. [PubMed] [Google Scholar]
- Pereira G, Lasso CA, Mora-Day J, Magalhães C. (2010a) Crustáceos decápodos de la Orinoquia Venezolana: biodiversidad, consideraciones biogeográficas y conservación. In: Lasso CA, Usma JS, Trujillo F, Rial A. (Eds) Biodiversidad de la Cuenca del Orinoco: Bases Científicas para la Identificación de Areas Prioritarias para la Conservación y Uso Sostenible de la Biodiversidad. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, WWF Colombia, Fundación Omacha, Fundación La Salle and Instituto de Estudios de la Orinoquia (Universidad Nacional de Colombia), Bogotá, 357–365. [Google Scholar]
- Pereira G, Lasso C, Mora-Day J, Magalhães C, Morales-Betancourt M, Campos M. (2010b) Lista de los crustáceos decápodos de la cuenca del río Orinoco (Colombia-Venezuela). Biota Colombiana 10(1-2): 75–87 [2009]. [Google Scholar]
- Pileggi LG, Mantelatto FL. (2010) Molecular phylogeny of the freshwater prawn genus Macrobrachium (Decapoda, Palaemonidae), with emphasis on the relationships among selected American species. Invertebrate Systematics 24: 194–208. doi: 10.1071/IS09043 [Google Scholar]
- Pileggi LG, Magalhães C, Bond-Buckup G, Mantelatto FL. (2013) New records and extension of the known distribution of some freshwater shrimps in Brazil. Revista Mexicana de Biodiversidad 84: 563–574. doi: 10.7550/rmb.30504 [Google Scholar]
- Ramos-Porto M, Coelho PA. (1990) Sinopse dos crustáceos decápodos brasileiros (Família Palaemonidae). Anais da Sociedade Nordestina de Zoologia 3: 93–111. [Google Scholar]
- Ramos-Porto M, Coelho PA. (1998) Malacostraca. Eucarida. Caridea (Alpheoidea excluded). In: Young PS. (Ed.) Catalogue of Crustacea of Brazil. Museu Nacional, Rio de Janeiro, 325–350. [Google Scholar]
- Rodríguez G. (1980) Crustaceos Decapodos de Venezuela. IVIC, Caracas, 494 pp. [Google Scholar]
- Rodríguez G. (1981) Decapoda. In: Hurlbert SH, Rodríguez G, Santos ND. (Eds) Aquatic Biota of Tropical South America, Part 1: Arthropoda. San Diego State University, San Diego, 41–51. [Google Scholar]
- Rodríguez G. (1982) Fresh-water shrimps (Crustacea, Decapoda, Natantia) of the Orinoco basin and the Venezuelan Guayana. Journal of Crustacean Biology 2(3): 378–391. doi: 10.2307/1548054 [Google Scholar]
- Ronquist F, van der Mark P, Huelsenbeck JP. (2009) Bayesian phylogenetic analysis using MRBAYES. In: Lemey P, Salemi M, Vandamme A-M. (Eds) The Phylogenetic Handbook. Cambridge University Press, Cambridge, 210–266. doi: 10.1017/CBO9780511819049.009 [Google Scholar]
- Ronquist F, Teslenko M, van der Mark P, Ayres D, Darling A, Höhna S, Larget B, Liu L, Suchard MA, Huelsenbeck JP. (2012) MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Systematic Biology 61: 539–542. doi: 10.1093/sysbio/sys029 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santos JC, Coloma LA, Summers K, Caldwell JP, Ree R, Cannatella DC. (2009) Amazonian amphibian diversity is primarily derived from Late Miocene Andean lineages. PLoS Biology 7: 448–461. doi: 10.1371/journal.pbio.1000056 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schubart CD, Cuesta JA, Felder DL. (2002) Glyptograpsidae, a new brachyuran family from Central America: larval and adult morphology, and a molecular phylogeny of the Grapsoidea. Journal of Crustacean Biology 22(1): 28–44. doi: 10.1163/20021975-99990206 [Google Scholar]
- Schubart CD, Neigel JE, Felder DL. (2000) Use of the mitochondrial 16S rRNA gene for phylogenetic and population studies of Crustacea. Crustacean Issues 12: 817–830. [Google Scholar]
- Sioli H. (1984) The Amazon and its main effluents: hydrography, morphology of the river courses, and river types. In: Sioli H. (Ed) The Amazon, Limnology and Landscape Ecology of a Mighty Tropical River and its Basin. Dr. W. Junk Publishers, Dordrecht, 127–166. [Google Scholar]
- Stamatakis A. (2006) RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 22(21): 2688–2690. doi: 10.1093/bioinformatics/btl446 [DOI] [PubMed] [Google Scholar]
- Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. (2011) MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Molecular Biology and Evolution 28(10): 2731–2739. doi: 10.1093/molbev/msr121 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Valencia DM, Campos MR. (2010) Freshwater shrimps of the Colombian tributaries of the Amazon and Orinoco rivers (Palaemonidae, Euryrhynchidae, Sergestidae). Caldasia 32(1): 221–234. [Google Scholar]
- Vieira IM. (2003) Diversidade de crustáceos das ressacas da Lagoa dos Índios, Tacacá e APA do Curiaú. In: Takiyama LR, Silva AQ. (orgs) Diagnóstico das Ressacas do Estado do Amapá: Bacias do Igarapé da Fortaleza e Rio Curiaú. CPAQ/IEPA, DGEO/SEMA, Macapá, 53–62. [Google Scholar]
- Wesselingh FP. (2006) Miocene long-lived lake Pebas as a stage of mollusc radiations, with implications for landscape evolution in western Amazonia. Scripta Geologica 133: 1–17. [Google Scholar]
- Willis SC, Nunes M, Montaña CG, Farias IP, Ortí G, Lovejoy NR. (2010) The Casiquiare river acts as a corridor between the Amazonas and Orinoco river basins: biogeographic analysis of the genus Cichla. Molecular Ecology 19: 1014–1030. doi: 10.1111/j.1365-294X.2010.04540.x [DOI] [PubMed] [Google Scholar]
- Xia X, Xie Z, Salemi M, Chen L, Wang Y. (2003) An index of substitution saturation and its application. Molecular Phylogenetics and Evolution 26: 1–7. doi: 10.1016/S1055-7903(02)00326-3 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.