

Abstract
Water chestnut (Trapa japonica Flerov.) is an annual aquatic plant. In the present study, we showed that the treatment of water chestnut extracted with boiling water resulted in a significant increase 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical-scavenging activity and decrease the intracellular H2O2-induced accumulation of reactive oxygen species. In addition, water chestnut extract (WCE) inhibited lipopolysaccharide (LPS)-induced nitric oxide production and suppressed mRNA and protein expression of the inducible nitric oxide synthase gene. The cytokine array results showed that WCE inhibited inflammatory cytokine secretion. Also, WCE reduced tumor necrosis factor-α-and interleukin-6-induced nuclear factor-αB activity. Furthermore, during sodium lauryl sulfate (SLS)-induced irritation of human skin, WCE reduced SLS-induced skin erythema and improved barrier regeneration. These results indicate that WCE may be a promising topical anti-inflammatory agent.
Keywords: Trapa japonica Flerov., Water chestnut, Anti-inflammatory effect, Cytokine, NF-αB
INTRODUCTION
The inflammation is a protective response that occurs in response to trauma, infection, tissue injury, or noxious stimuli and regulated by the activation of various immune cells (Zedler and Faist, 2006; Mariathasan and Monack, 2007; Kwon et al., 2008). Specifically, macrophages play a central role in mediating many different immune-pathological phenomena during inflammation, including the overexpression of pro-inflammatory cytokine and inflammatory mediators (Dokka et al., 2001) such as tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β), nitric oxide (NO), prostaglandin E2 (PGE2), and regulated on activation normal T cell expressed and secreted (RANTES) (Noel et al., 2004; Mosser and Edwards, 2008).
The expression of these inflammatory mediators can be regulated by activation of the transcription factor nuclear factor kappa-B (NF-αB), which plays a critical role in regulating the expression of various genes, including the cytokines, inducible NOS (iNOS) and cyclooxygenase-2 (COX-2) (Makarov, 2000). Since NF-αB plays a ubiquitous role in the pathogenesis of inflammatory gene expression, the cytokine is a current target for the treatment of various diseases (Yamamoto and Gaynor, 2001). In addition, NO production, which is critical in multiple cellular signaling pathways, is catalyzed by nitric oxide synthases (NOSs), which are classified as iNOS, endothelial NOS (eNOS), and neuronal NOS (nNOS) (Alderton et al., 2001). iNOS is highly expressed in macrophages, where its activation in several inflammatory and autoimmune diseases results in organ destruction. Another important inflammatory mediator is PGE2 that is produced from arachidonic acid metabolites by the catalysis of COX-2 (Wang et al., 2011). Therefore, a search for a new bioactive material is required to determine whether activator of anti-inflammation can repress these inflammatory mediators.
Lipopolysaccharide (LPS), a component of the cell wall of gram-negative bacteria, induces the activation of monocytes and macrophages, as well as the production of pro-inflammatory cytokines (Latorre et al., 2010). Also, LPS stimulation can induce the production of pro-inflammatory cytokines and inflammatory mediators in RAW 264.7 murine macrophage cells, therefore, this cell system provides an excellent model for drug screening and evaluating putative inhibitors of the inflammatory response (Yoon et al., 2009). In addition, sodium lauryl sulfate (SLS), an anionic detergent that is widely used as a synthetic cleansing agent in cosmetics and dentifrices (Babich and Babich, 1997), is the most widely utilized model for studying acute and cumulative skin irritation (Lee and Maibach, 1995). The detergent disrupts skin barrier function, resulting in increased transepidermal water loss (TEWL) and erythema (Froebe et al., 1990). Therefore, the development of materials that attenuate inflammation and skin irritation is needed in these model systems.
Water chestnut (Trapa Japonica Flerov.) is an aquatic plant that grows abundantly in shallow freshwater basins. The plant has been commercially cultivated for its edible fruits in aquatic areas in the lowlands, flatlands, or lakes in Korea, Japan, China, India, and North America (Hoque and Arima, 2002). Most studies on water chestnuts have focused on its nutritional and ecological value, but its biological activity has rarely been examined, with the exception of the antioxidant activity of Chinese water chestnuts (You et al., 2007). In this study, we examined the effect of water chestnut extracted with boiling water on intracellular ROS accumulation; LPS-induced production of pro-inflammatory cytokines and inflammatory mediators; and the inhibition of NF-αB. Furthermore, we demonstrated that WCE can reduce SLS-induced skin irritation in vivo.
MATERIALS AND METHODS
Preparation of plant extract
The epicarps of Trapa Japonica Flerov. were collected in October 2009 from the Weissan lake from Shandong, China. The plant extract was obtained from 1 kg of the epicarp of T. japonica Flerov. in 2.5 L of boiling water extraction for 1 h and then evaporated and filtered.
Cell culture
The RAW 264.7 murine macrophage cell line was purchased from the Korea Cell Line Bank (Seoul, South Korea). The RAW 264.7 cells were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM; Gibco, NY, USA) supplemented with 10% fetal bovine serum (FBS; Gibco, NY, USA), 100 U/mL of penicillin, and 100 μg/mL of streptomycin in 5% CO2 at 37°C. HaCat cells (human keratinocyte cell line) and Hep3B cells were cultured in RPMI 1640 medium containing 10% (v/v) FBS and 1% penicillin-streptomycin (10,000 U and 10,000 μg/mL, respectively) in 5% CO2 at 37°C.
Measurements of the DPPH radical scavenging activity and intracellular ROS accumulation
The 2,2-diphenyl-1-picrylhydrazyl (DPPH) assay was used to determine the anti-oxidant capacity of WCE. The DPPH radical scavenging activity is generally quantified in terms of inhibition percentage of the pre-formed free radical by anti-oxidants, and the SC50 (concentration required to obtain a 50% scavenging effect) is a typically employed parameter to express the anti-oxidant capacity. Ascorbic acid was used as an anti-oxidant standard to define the SC50 parameters. HaCat cells were incubated at a density of 2.5×104 cells in clear bottom black polystyrene 96-well plates (Costar, Corning, NY, USA) overnight. Cells were gently washed with Dulbecco’s phosphate-buffered saline (DPBS) 2–3 times and treated with WCE for 1 h. The fluorescent probe, dichlorofluorescein diacetate (DCF-DA), was used to monitor the intracellular generation of H2O2-induced ROS. After 30 min of H2O2 (1 mM) treatment, DCF-DA (total 1 μM) was added to the cells, which were subsequently incubated for 30 min at 37°C. The intracellular ROS concentration was monitored using a Zeiss HBO 100 (Thornwood, NY, USA) inverted fluorescence microscope and fluorescence microplate reader Infinite 200 (Tecan, Männedorf, Switzerland) at excitation and emission wavelengths of 485 and 535 nm.
MTT assay
The MTT assay was used to evaluate the effects of WCE on cell viability. RAW 264.7 cells were plated in 96-well plates at a density of 1×104 cells/well for 24 h. The cells were treated with varying concentrations of WCE (10, 50, and 100 μg/mL) for 24 h at 37°C, followed by 50 μL of 2 mg/mL 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT; Sigma-Aldrich, St. Louis, MO, USA) solution to each well and further incubation for 4 h at 37°C. The media were discarded and 100 μL of dimethyl sulfoxide (DMSO) was added to each well to solubilize the formazan. Optical density was measured at 540 nm on a SpectraMax 190 microplate reader (Molecular Devices, Sunnyvale, CA, USA).
Assay for NO production
RAW 264.7 cells (2×105 cells/well) were seeded in a 96-well plate for 24 h at 37°C. Cells were treated with 1 μg/mL of LPS alone or in combination with different concentrations of WCE for 24 h. After 24 h, 100 μL of the medium was placed in a 96-well plate and an equal volume of Griess reagent (Sigma-Aldrich) was added. The cells in the plate were then incubated for an additional 15 min at room temperature, and the absorbance was measured at 540 nm with the microplate reader. The amount of NO generated was calculated using a sodium nitrite standard curve.
Assay for PGE2 production
RAW 264.7 cells (2×105 cells/well) were seeded in a 96-well plate for 24 h at 37°C. Cells were treated with 1 μg/mL of LPS alone or in combination with different concentrations of WCE for 24 h. The PGE2 concentration in the culture medium was quantified using a competitive enzyme immunoassay kit (R&D Systems, MN, USA) according to the manufacturer’s instructions.
Reverse-transcription polymerase chain reaction
RAW 264.7 cells (1.0×107 cells/100 π dish) were treated with 1 μg/mL of LPS alone or in combination with different concentrations of WCE for 24 h (for iNOS analyses), 4 h (for RANTES), or 6 h (for IL-6 analysis). Cells were washed with PBS, and total RNA was isolated using Trizol reagent according to the manufacturer’s instructions (Invitrogen, Carlsbad, CA, USA). Reverse transcription was carried out with 1 μg RNA samples by using reverse transcription master premix (ELPIS Biotech, Daejeon, South Korea) at 42°C for 60 min followed by inactivation of the enzyme at 94°C for 5 min. The PCR was carried out in a 20-μL reaction mixture containing 4 μL of rTaq Plus 5x PCR Premix (ELPIS Biotech), 13 μL of double-distilled DNase-free water, 1 μL of each primer (10 pmol), and 1 μL of reverse transcriptase (RT) product. The polymerase chain reaction (PCR) was performed on a GeneAmp PCR System 9700 system (Applied Biosystems, Carlsbad, CA, USA). The sequences of the specific primers for mouse iNOS, IL-6, RAN-TES, and β-actin are as follows: sense 5′-AGC CCA ACA ATA CAA ATG ACC CTA-3′ and antisense 5′-TTC CTG TTG TTT CTA TTT CCT TTG T-3′ for iNOS; sense 5′-CAT GTT CTC TGG GAA ATC GTG G-3′ and antisense 5′-AAC GCA CTA GGT TTG CCG AGT A-3′ for IL-6; sense 5′-CCT CAC CAT CAT CCT CAC TGC A-3′ and antisense 5′-CAT CCC CAA GCT GGC TAG GAC T-3′ for RANTES; and sense 5′-TGG AAT CCT GTG GCA TCC ATG AAA C-3′and antisense 5′-TAA AAC GCA GCT CAG TAA CAG TCC G-3′ for β-actin. Amplification was performed for 28-30 cycles. Each cycle consisted of denaturation for 10 sec at 95°C, annealing for 10 sec at the appropriate primer-specific temperature, and extension for 10 sec at 72°C. An additional incubation at 72°C for 10 min was executed after the last cycle. The PCR products were subject to electrophoresis on 1.8% agarose gels.
Western blot analysis
RAW 264.7 cells (1.0×107 cells/100 π dish) were treated with 1 μg/mL of LPS alone or in combination with different concentrations of WCE for 24 h. After 24 h, the cells were collected and washed twice with PBS. The cells were then lysed in RIPA buffer and kept on ice for 1 h. Cell lysates were centrifuged at 12,000×g at 4°C for 15 min, and the supernatants were stored at −70°C until required for analysis. Protein concentrations were measured using a protein assay kit (Bio-Rad, Hercules, CA, USA). Aliquots of the lysates (20 μg of protein) were applied to a 10% sodium dodecyl sulfate (SDS)-polyacrylamide gel and transferred onto a polyvinylidene fluoride (PVDF) membrane. After blocking nonspecific sites with 5% non-fat milk powder in Tris-buffered saline with 0.1% Tween-20 (TBST), the membranes were subsequently incubated with specific primary antibodies (goat anti-iNOS, and mouse anti-β-actin antibodies; Santa Cruz Biotechnology, Santa Cruz, CA, USA) for 3 h at room temperature. The membranes were subsequently incubated for 1 h with peroxide-conjugated secondary antibodies (Santa Cruz Biotechnology). The immunoreactive proteins were detected using an enhanced chemiluminescence (ECL) western blotting detection kit (Amersham Pharmacia Biotech, NY, USA).
Protein chip assay
The RayBio Mouse Cytokine Antibody Array I was purchased from RayBiotech (Norcross, GA, USA) and utilized according to manufacturer’s instructions. The array membranes were incubated with blocking buffer followed by undiluted culture supernatants for 1.5 h. Next, the membranes were washed and incubated with biotin-conjugated antibodies for 1.5 h, followed by HRP-conjugated streptavidin for 2 h. The membranes were then incubated in detection buffer and exposed to X-ray film.
Transfection and luciferase assay
Hep3B cells were grown overnight in 24-well plates and washed twice with serum-free medium. DNA-Lipofectamine complex containing 1.0 μg/mL NF-αB-luciferase vector, 0.5 μg/mL pSV β-galactosidase vector (Promega, Madison, WI, USA), and 5 μg/mL Lipofectamine (Invitrogen) were allowed to form at room temperature for 15 min. The cells were overlaid with the DNA-Lipofectamine complex and incubated at 37°C for 4–6 h. After washing the cells with serum-free medium, fresh growth medium containing 10% FBS was added. Transfected cells were treated with lysis buffer (Promega), and aliquots of the lysates were assayed for luciferase and β-galactosidase enzyme activity. β-Galactosidase enzyme activity was used to normalize for variability in transfection efficiency.
Skin irritation test
Ten healthy women (age, 20–50 years) were selected for the study according to the established inclusion and exclusion criteria commonly applied by the Ellead Skin Research Center (http://www.ellead.com) for clinical studies with protocol number (EL-P-4700). All the subjects signed informed consent guidelines pertaining to good clinical practice (GCP) for trials of pharmaceutical products, and the study was performed in accordance with the Declaration of Helsinki. No subject applied any moisturizer or skin care products on their arms during the 14-day testing period. Three different gels incorporating 500 and 1,250 μg/mL of WCE were prepared. The formulation of the solubilized gel consisted of the following: propanediol, hydroxypropyl cyclodextrin, betaine, ammonium acryloyldimethyltaurate/VP copolymer, dextrin, sodium benzotriazolyl butylphenol sulfonate, buteth-3, tributyl citrate, Tris (tetramethylhydroxypiperidinol) citrate, BHT, sodium bisulfite, disodium EDTA, preservatives, fragrance, and distilled water. WCE-containing gels and placebo were tested on the subjects’ ventral arms. The degree of skin irritation was measured with a Tewameter (Courage & Khazaka, Cologne, Germany), which measured the change in water evaporation. In this study, all volunteers were exposed to 1.5% sodium lauryl sulfate (SLS) for 24 h to induce skin irritation. Upon removal of the SLS patches (IQ chamber; Chemotechnique Diagnostics, Vellinge, Sweden), the skin was gently rinsed with water and allowed to dry. The gels were then applied to the SLS-treated areas once daily. The transepidermal water loss (TEWL) was measured at approximately the same time each day on days 0, 1, 2, and 14 d after administration.
HPLC analysis
HPLC analysis was performed on a Waters HPLC system consisting of a 2960 separations module, a 996 Photodiode Array Detector (Waters, Milford, MA, USA), and WATERS Millennium 32 software for peak identification and integration. The separation was carried out on a Phenomenex Prodigy ODS-2 column (150×3.2 mm, 5 μm). Acetonitrile (solvent A) and 0.03% (v/v) phosphoric acid in water (solvent B) were used. The samples were eluted according to the following gradient: a linear increase from 15% to 95% of solvent A in 30 min, started with 85% of solvent B. The flow rate was 0.8 mL/min and the detection wavelength was 280 nm.
Statistical analysis
Data are presented as means ± standard error of the mean (SEM) from more than 3 separate experiments performed in triplicate. They were analyzed using the two-sided Student’s t test. Statistical significance was defined as p<0.05, p<0.01, and p<0.001.
RESULTS
Anti-oxidant effects of WCE
To determine the anti-oxidant capacity of WCE, its antioxidant activity was determined by measuring the DPPH radicalscavenging reaction. Compared to treatment of the cells with the vitamin C positive control, treatment of HaCat cells with WCE markedly increased DPPH radical-scavenging activity according to concentration (Fig. 1A). WCE exhibited highly radical-scavenging activity (SC50=12.8 μg/mL; vitamin C, SC50=2.1 μg/mL). The intracellular ROS-scavenging activity of WCE was monitored by the dichlorofluorescein (DCF) fluorescence intensity of H2O2 induction. The WCE reduced intracellular ROS in a dose dependently manner (Fig. 1B). These results indicate that WCE has the anti-oxidant capacity.
Fig. 1.

Effect of WCE on DPPH radical-scavenging activity and intracellular ROS accumulation induced by H2O2. (A) HaCat cells were treated with WCE or vitamin C (Vit. C). and then DPPH radical-scavenging activities were estimated, black bar WCE treated group, white bar vitamin C treated group. *** p<0.001 compared to the untreated control group. §p<0.05 compared to the vitamin C treated group. (B) Ha-Cat cells were treated with DCF-DA in the presence or absence of WCE for 2 h, followed by H2O2 treatment, black bar WCE treated group, white bar H2O2 treated group. **p< 0.01 compared to H2O2 treated group. Values are presented as means ± standard error of the mean (SEM).
Effects of WCE on NO and PGE2 production
Cell viability was not affected by 100 μg/mL WCE (data not shown). Therefore, all subsequent experiments on inflammatory mediator production were conducted with 100 μg/mL WCE. To investigate the anti-inflammatory effects of WCE, we examined its effect on NO production in LPS-induced macrophages. NO production greatly increased after 24-h LPS treatment; subsequent addition of WCE resulted in dose-dependent reduction of LPS-stimulated NO production (Fig. 2A). We subsequently examined the effects of WCE on PGE2 production in LPS-stimulated RAW 264.7 cells. After RAW 264.7 cells were stimulated with LPS for 24 h, PGE2 levels increased in the culture medium were suppressed by subsequent addition of WCE (Fig. 2B).
Fig. 2.

Effects of WCE on LPS-induced NO and PGE2 production, and iNOS expression. (A) RAW 264.7 cells were treated with 1 μg/mL of LPS alone or in combination with varying concentrations of WCE for a nitrite assay. The extracellular medium containing nitrate was analyzed by a Griess regent system. (B) The PGE2 concentration in the supernatants was determined by ELISA *p<0.05, **p<0.01 compared to the LPS treated group. Values are presented as means ± standard error of the mean (SEM). (C) Western blot analysis of iNOS. β-actin was used as an internal standard. (D) RT-PCR analysis of iNOS. β-actin was used as an internal standard. The PCR products were resolved on a 2% agarose gel.
Effects of WCE on LPS-induced iNOS expression
To elucidate the mechanism by which WCE inhibits LPS-induced NO production, we investigated its effects on iNOS mRNA and protein expression. The levels of iNOS protein, which were minimal in RAW 264.7 cells in their resting state, markedly increased upon LPS exposure. Subsequent addition of WCE dose-dependently inhibited LPS-induced iNOS mRNA and protein expression (Fig. 2C, D). This observation suggests that the inhibitory effects of WCE on LPS-induced NO production is attributable to LPS-induced iNOS gene expression.
Effects of WCE on LPS-induced cytokine expression
A mouse cytokine antibody array (Fig. 3A) was applied to broadly observe the effects of WCE on LPS-induced cytokine expression. After the cells were incubated with WCE for 24 h, the cytokine expression pattern in the treated cells were differentially compared to that in the control cells (Fig. 3B). After LPS treatment, the protein expression levels of GCSF, GMCSF, IL-6, IL-9, IL-12 p40/p70, MCP-1, RANTES, and TNF-α all increased. Each LPS-induced dot in the array was quantified, and the results indicated that WCE inhibited LPS-induced secretion of IL-6, RANTES, IL-9, IL-12 p40/p70, and GM-CSF proteins in RAW 264.7 cells (Fig. 3C). Using RT-PCR, we further analyzed whether WCE inhibited LPS-induced mRNA expression levels of IL-6 and RANTES. LPS-induced IL-6 and RANTES mRNA expression peaked after 6 h (data not shown). After incubation 6 h, WCE treatment induced the inhibition of IL-6 and RANTES mRNA expression (Fig. 3D). In contrast, WCE did not affect expression of the housekeeping gene β-actin.
Fig. 3.

Cytokine profile of the effect of WCE on LPS-activated RAW 264.7 cells. (A) Each cytokine is represented by duplicate spots in the location shown. (B) RAW 264.7 cells (1.0×107cells/100 π dish) were treated with 1 μg/ml of LPS only or with different concentrations of water chestnut extract for 24 h. At the end of the incubation period, RAW 264.7 supernatants were collected and assayed for cytokine production using the Mouse Cytokine Array I. (C) The ratio of average intensity of each cytokine spot to each positive spot on the same chip is shown. Values are presented as mean ± S.E.M. *p<0.05 compared to the LPS group. White bars, no treatment control group; hatched bars, LPS group; gray bars, WCE group treated at 50 μg/mL after LPS treatment; black bars, WCE group treated at 100 μg/mL after LPS treatment. (D) RAW 264.7 cells were treated with 1 μg/mL of LPS alone or in combination with different concentrations of WCE for 6 h. Total mRNA was isolated, and mRNA levels of IL-6, RANTES, and β-actin were measured by RT-PCR. β-actin was used as an internal standard.
Inhibitory effects of WCE on NF-αB induced by TNF-α and IL-6
To further elucidate the effects of LPS and cytokine treatment on the expression of NF-αB-regulated genes, we performed transfection studies using an NF-αB-luciferase construct in human hepatocellular carcinoma Hep3B cells. In this study, the NF-αB activity induced by TNF-α and IL-6 treatment dose-dependently reduced on WCE addition (Fig. 4). These results indicate that WCE downregulates iNOS protein and mRNA, as well as cytokine (i.e., TNF-α and IL-6) expression, by suppressing NF-αB activity.
Fig. 4.

Inhibitory effect of WCE on TNF-α and IL-6 induced NF-αB activity. Hep3B cells were treated with 10 ng/mL TNF-α or IL-6 by using the serum-free media supplemented with 0.1% bovine serum albumin and cultured for 24 h. At the end of the incubation period, luciferase and β-galactosidase activity were determined. Values are presented as mean ± S.E.M. *p<0.05, **p<0.01, and ***p<0.001 compared to the TNF-α and IL-6 alone-treated group respectively.
Anti-skin irritation effect of WCE
We further investigated the mechanism by which gel emulsion containing the WCE can reduce SLS-induced irritation. We repeatedly applied skin care products over a 2-week period onto the skin of subjects and induced acute irritant contact dermatitis by 24-h occlusion with 1.5% (w/v) SLS. The response to SLS was monitored by measuring TEWL. The increase in TEWL values caused by SLS was statistically reduced by WCE in comparison with the placebo at 1, 2 and 7 day (Fig. 5).
Fig. 5.
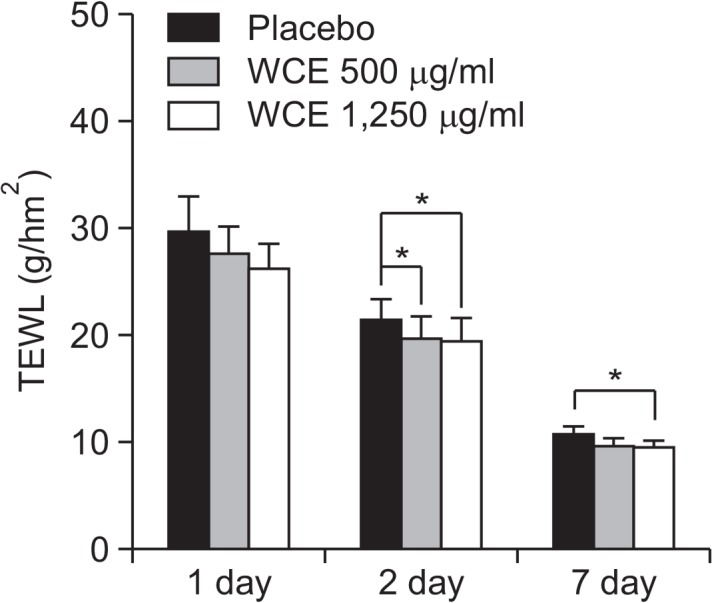
Anti-irritation effect of WCE on SLS-induced irritation. All 10 volunteers were exposed to 1.5% SLS for 24 h to induce skin irritation. The gel emulsions were subsequently applied to the SLStreated areas once daily. The skin TEWL was measured at approximately the same time each day on days 0, 1, 2, and 7. Values are presented as mean ± S.E.M. *p< 0.05 compared to the placebotreated group.
HPLC analysis
Several peaks were monitored in the HPLC profile of WCE (Fig. 6). Two peaks were qualitatively identified based on retention times of 30 HPLC reference compounds used and the other were not identified owing to lack of authentic references. Among the identified compounds, there were major compounds, that is, gallic acid and ellagic acid. Gallic acid was the most abundant.
Fig. 6.

HPLC analysis of WCE. Each arrow indicates gallic acid and ellagic acid. The other peaks were not identified owing to lack of authentic compounds.
DISCUSSION
Inflammation is characterized by swelling, redness, and warmth at the site, often accompanied by pain and a response by the immune system to tissue damage. Macrophages participate in inflammatory responses by releasing pro-inflammatory cytokines such as TNF-α and inflammatory factors (Baumann and Gauldie, 1994; Dokka et al., 2001). In addition, it has been reported that cytokines were abundantly expressed in atopic skin disease (Gutowska-Owsiak and Ogg, 2013), and inflammatory cytokine attenuated ceramide production in the whole epidermis, which are predominantly responsible for the skin barrier function (Sawada et al., 2012). Therefore, the inhibition of these inflammatory factors has been considered as a novel candidate for a skin therapeutic agent. In addition, the potential inhibitors of iNOS have been considered as potential anti-inflammatory agents (Murakami and Ohigashi, 2007). In present study, we found that WCE inhibited iNOS mRNA and protein expression, resulting in reduced NO in LPS-stimulated cells.
WCE inhibits LPS-induced protein secretion of IL-6, RANTES, IL-9, IL-12 p40/p70, and GM-CSF. Since protein secretion of IL-6 and RANTES was more markedly reduced than that of other cytokines. Interleukin 6 is a pivotal pro-inflammatory cytokine generated by monocytes, fibroblasts, T-cells, and B-cells and has a putative role in mycobacterial infection through its involvement in adaptive cell activation and acutephase protein and immunoglobulin production (Van Snick, 1990). Similarly, RANTES is a member of a large family of chemotactic cytokines, known as chemokines, which can recruit and activate a wide variety of proinflammatory cells (Oppenheim et al., 1991; Schall, 1991). Moreover, RANTES promotes cell accumulation and activation in chronic inflammatory diseases (Schall, 1991). Our study suggests that WCE inhibits the production of NO, IL-6, and RANTES via reduction in transcriptional down-regulation of the iNOS, IL-6, and RANTES genes, respectively.
NF-αB controls the expression of genes encoding several pro-inflammatory cytokines (e.g., IL-1, IL-2, IL-6, and TNF-α), chemokines (e.g., IL-8, MIP-1α, MCP1, RANTES, and eotaxin), adhesion molecules (e.g., ICAM, VCAM, and E-selectin), inducible enzymes (iNOS), growth factors, acute phase proteins, and immune receptors, all of which play a critical role in modulating inflammatory processes. Many reports suggest that plant-derived compounds have antioxidant properties and act by preventing NF-αB activation, followed by inhibiting iNOS and cytokine gene expression (Froebe et al., 1990; Kang et al., 2008). Our study demonstrated that WCE treatment reduced the extent of IL-6-and TNF-α-induced increases in NF-αB-dependent luciferase activity. The results indicate that WCE may exert anti-inflammatory effects via reduction of the pro-inflammatory mediators through suppression of the NF-αB-mediated signaling pathways.
Reactive oxygen species are associated with the inflammatory response and frequently contribute to the tissue-damaging effects of inflammatory reactions (Johar et al., 2004). We elucidated that WCE increased DPPH radical scavenging and reduced intracellular H2O2-induced ROS accumulation. Anti-irritants are used in cosmetic products to prevent or treat skin irritations that arise during daily life. Our study demonstrated a strong anti-inflammatory effect of WCE on human skin, as measured by TEWL and skin sensitivity to SLS-induced irritation. Therefore, these agents may also be effective in preventing acute irritant contact dermatitis.
Our study showed that phenolic compounds such as gallic acid were detected in WCE. It has been reported that gallic acid suppressed melanogenesis in melanoma cells by increasing of extracellular signal-regulated kinase (ERK)/AKT phosphorylation and significantly inhibited melanin production followed by the down-regulation of microphthalmia-associated transcription factor (MITF), tyrosinase, tyrosinase-related protein-1 (TRP1), and dopachrome tautomerase (Dct) (Kumar et al., 2013; Su et al., 2013). In addition, gallic acid has been shown to have various biological properties, including antioxidant, anti-inflammatory, anti-viral, and anti-cancer activities (Kroes et al., 1992; Chuang et al., 2010). Therefore, our study suggests that gallic acid acts as a main anti-inflammatory activity in WCE. These findings provide evidence that WCE may be a putative therapeutic agent against skin inflammatory disorders.
REFERENCES
- Alderton WK, Cooper CE, Knowles RG. Nitric oxide synthases: structure, function and inhibition. Biochem J. 2001;357:593–615. doi: 10.1042/0264-6021:3570593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Babich H, Babich JP. Sodium lauryl sulfate and triclosan: in vitro cytotoxicity studies with gingival cells. Toxicol Lett. 1997;91:189–196. doi: 10.1016/S0378-4274(97)00022-2. [DOI] [PubMed] [Google Scholar]
- Baumann H, Gauldie J. The acute phase response. Immunol. Today. 1994;15:74–80. doi: 10.1016/0167-5699(94)90137-6. [DOI] [PubMed] [Google Scholar]
- Chuang CY, Liu HC, Wu LC, Chen CY, Chang JT, Hsu SL. Gallic acid induces apoptosis of lung fibroblasts via a reactive oxygen species-dependent ataxia telangiectasia mutatedp53 activation pathway. J Agric Food Chem. 2010;58:2943–2951. doi: 10.1021/jf9043265. [DOI] [PubMed] [Google Scholar]
- Dokka S, Shi X, Leonard S, Wang L, Castranova V, Rojanasakul Y. Interleukin-10-mediated inhibition of free radical generation in macrophages. Am J Physiol Lung Cell Mol Physiol. 2001;280:L1196–1202. doi: 10.1152/ajplung.2001.280.6.L1196. [DOI] [PubMed] [Google Scholar]
- Froebe CL, Simion FA, Rhein LD, Cagan RH, Kligman A. Stratum corneum lipid removal by surfactants: relation to in vivo irritation. Dermatologica. 1990;181:277–283. doi: 10.1159/000247822. [DOI] [PubMed] [Google Scholar]
- Gutowska-Owsiak D, Ogg GS. Cytokine regulation of the epidermal barrier. Clin. Exp. Allergy. 2013;43:586–598. doi: 10.1111/cea.12023. [DOI] [PubMed] [Google Scholar]
- Hoque A, Arima S. Overcoming phenolic accumulation during callus induction and in vitro organogenesis in water chestnut (Trapa Japonica Flerov) In Vitro Cell. Dev. Biol. Plant. 2002;38:342–346. doi: 10.1079/IVP2002305. [DOI] [Google Scholar]
- Johar D, Roth JC, Bay GH, Walker JN, Kroczak TJ, Los M. Inflammatory response, reactive oxygen species, programmed (necrotic-like and apoptotic) cell death and cancer. Rocz. Akad Med Bialymst. 2004;49:31–39. [PubMed] [Google Scholar]
- Kang OH, Chae HS, Choi JG, Oh YC, Lee YS, Kim JH, Seung MJ, Jang HJ, Bae KH, Lee JH, Shin DW, Kwon DY. Ent-pimara-8(14), 15-dien-19-oic acid isolated from the roots of Aralia cordata inhibits induction of inflammatory mediators by blocking NF-kappaB activation and mitogen-activated protein kinase pathways. Eur J Pharmacol. 2008;601:179–185. doi: 10.1016/j.ejphar.2008.10.012. [DOI] [PubMed] [Google Scholar]
- Kroes BH, van den Berg AJ, van Ufford HCQ, van Dijk H, Labadie RP. Anti-inflammatory activity of gallic acid. Planta Med. 1992;58:499–504. doi: 10.1055/s-2006-961535. [DOI] [PubMed] [Google Scholar]
- Kumar KJ, Vani MG, Wang SY, Liao JW, Hsu LS, Yang HL, Hseu YC. In vitro and in vivo studies disclosed the depigmenting effects of gallic acid: a novel skin lightening agent for hyperpigmentary skin diseases. Biofactors. 2013;39:259–270. doi: 10.1002/biof.1064. [DOI] [PubMed] [Google Scholar]
- Kwon HS, Park JH, Kim DH, Kim YH, Shin HK, Kim JK. Licochalcone A isolated from licorice suppresses lipopolysaccharide-stimulated inflammatory reactions in RAW264.7 cells and endotoxin shock in mice. J Mol Med. 2008;86:1287–1295. doi: 10.1007/s00109-008-0395-2. [DOI] [PubMed] [Google Scholar]
- Latorre D, Puddu P, Valenti P, Gessani S. Reciprocal interactions between lactoferrin and bacterial endotoxins and their role in the regulation of the immune response. Toxins. 2010;2:54–68. doi: 10.3390/toxins2010054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee CH, Maibach HI. The sodium lauryl sulfate model: an overview. Contact Dermatitis. 1995;33:1–7. doi: 10.1111/j.1600-0536.1995.tb00438.x. [DOI] [PubMed] [Google Scholar]
- Makarov SS. NF-kappaB as a therapeutic target in chronic inflammation: recent advances. Mol. Med. Today. 2000;6:441–448. doi: 10.1016/S1357-4310(00)01814-1. [DOI] [PubMed] [Google Scholar]
- Mariathasan S, Monack DM. Inflammasome adaptors and sensors: intracellular regulators of infection and inflammation. Nat Rev Immunol. 2007;7:31–40. doi: 10.1038/nri1997. [DOI] [PubMed] [Google Scholar]
- Murakami A, Ohigashi H. Targeting NOX, iNOS and COX-2 in inflammatory cells: chemoprevention using food phytochemicals. Int. J. Cancer. 2007;121:2357–2363. doi: 10.1002/ijc.23161. [DOI] [PubMed] [Google Scholar]
- Mosser DM, Edwards JP. Exploring the full spectrum of macrophage activation. Nat Rev Immunol. 2008;8:958–969. doi: 10.1038/nri2448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Noel W, Raes G, Hassanzadeh Ghassabeh G, De Baetselier P, Beschin A. Alternatively activated macrophages during parasite infections. Trends Parasitol. 2004;20:126–133. doi: 10.1016/j.pt.2004.01.004. [DOI] [PubMed] [Google Scholar]
- Oppenheim JJ, Zachariae CO, Mukaida N, Matsushima K. Properties of the novel proinflammatory supergene “intercrine” cytokine family. Annu Rev Immunol. 1991;9:617–648. doi: 10.1146/annurev.iy.09.040191.003153. [DOI] [PubMed] [Google Scholar]
- Sawada E, Yoshida N, Sugiura A, Imokawa G. Th1 cytokines accentuate but Th2 cytokines attenuate ceramide production in the stratum corneum of human epidermal equivalents: an implication for the disrupted barrier mechanism in atopic dermatitis. J Dermatol Sci. 2012;68:25–35. doi: 10.1016/j.jdermsci.2012.07.004. [DOI] [PubMed] [Google Scholar]
- Schall TJ. Biology of the RANTES/SIS cytokine family. Cytokine. 1991;3:165–183. doi: 10.1016/1043-4666(91)90013-4. [DOI] [PubMed] [Google Scholar]
- Su TR, Lin JJ, Tsai CC, Huang TK, Yang ZY, Wu MO, Zheng YQ, Su CC, Wu YJ. Inhibition of melanogenesis by gallic acid: possible involvment of the PI3K/Akt, MEK/ERK and Wnt/β-Catenin signaling pathways in B16F10 cells. Int J Mol Sci. 2013;14:20443–20458. doi: 10.3390/ijms141020443. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Snick J. Interleukin-6: an overview. Annu Rev Immunol. 1990;8:253–278. doi: 10.1146/annurev.iy.08.040190.001345. [DOI] [PubMed] [Google Scholar]
- Wang CC, Choy CS, Liu YH, Cheah KP, Li JS, Wang JT, Yu WY, Lin CW, Cheng HW, Hu CM. Protective effect of dried safflower petal aqueous extract and its main constituent, carthamus yellow, against lipopolysaccharide-induced inflammation in RAW264.7 macrophages. J Sci Food Agric. 2011;91:218–225. doi: 10.1002/jsfa.4172. [DOI] [PubMed] [Google Scholar]
- Yamamoto Y, Gaynor RB. Role of the NF-kappaB pathway in the pathogenesis of human disease states. Curr Mol Med. 2001;1:287–296. doi: 10.2174/1566524013363816. [DOI] [PubMed] [Google Scholar]
- Yoon WJ, Ham YM, Kim SS, Yoo BB, Moon JY, Baik JS, Lee NH, Hyun CG. Suppression of pro-inflammatory cytokines, iNOS, and COX-2 expression by brown algae Sargassum micracanthum in RAW 264.7 macrophages. EurAsia J Biosci. 2009;3:130–143. doi: 10.5053/ejobios.2009.3.0.17. [DOI] [Google Scholar]
- You Y, Duan X, Wei X, Su X, Zhao M, Sun J, Ruenroengklin N, Jiang Y. Identification of major phenolic compounds of Chinese water chestnut and their antioxidant activity. Molecules. 2007;12:842–852. doi: 10.3390/12040842. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zedler S, Faist E. The impact of endogenous triggers on trauma-associated inflammation. Curr. Opin. Crit. Care. 2006;12:595–601. doi: 10.1097/MCC.0b013e3280106806. [DOI] [PubMed] [Google Scholar]