

Abstract
We investigate evolutionary dynamics of two-strategy matrix games with zealots in finite populations. Zealots are assumed to take either strategy regardless of the fitness. When the strategy selected by the zealots is the same, the fixation of the strategy selected by the zealots is a trivial outcome. We study fixation time in this scenario. We show that the fixation time is divided into three main regimes, in one of which the fixation time is short, and in the other two the fixation time is exponentially long in terms of the population size. Different from the case without zealots, there is a threshold selection intensity below which the fixation is fast for an arbitrary payoff matrix. We illustrate our results with examples of various social dilemma games.
Mathematics Subject Classification: 91A22, 60J70
Introduction
A standard assumption underlying evolutionary game dynamics, regardless of whether a player is social agent or gene, is that players tend to imitate successful others. In actual social evolutionary dynamics, however, there may be zealous players that stick to one option according to their idiosyncratic preferences regardless of the payoff that they or their peers earn. Collective social dynamics in the presence of zealots started to be examined for non-game situations such as the voter model representing competition between two equally strong opinions (i.e., neutral invasions) (Mobilia 2003; Galam and Jacobs 2007; Mobilia et al. 2007; Xie et al. 2011; Singh et al. 2012). Zealots seem to be also relevant in evolutionary game dynamics. For example, voluntary immunization behavior of individuals when epidemic spreading possibly occurs in a population can be examined by a public-goods dilemma game (Fu et al. 2011). In this situation, some individuals may behave as zealot such that they try to immunize themselves regardless of the cost of immunization (Liu et al. 2012).
In our previous work, we examined evolutionary dynamics of the prisoner’s dilemma and snowdrift games in infinite populations with zealots (Masuda 2012). Specifically, we assumed zealous cooperators and asked the degree to which the zealous cooperators facilitate cooperation in the entire population. We showed that cooperation prevails if the temptation of unilateral defection is weak or the selection strength is weak. For the prisoner’s dilemma, we analytically obtained the condition of cooperation.
In the present paper, we conduct a finite population analysis of evolutionary dynamics of a general two-person game with zealots. Evolutionary games in finite populations have been recognized as a powerful analytical tool for understanding properties of evolutionary games such as conditions of cooperation in social dilemma games. In addition, the outcome for finite populations is often different from that for infinite populations (Nowak et al. 2004; Taylor et al. 2004; Nowak 2006). We take advantage of this method to understand evolutionary dynamics of games with zealots for general matrix games.
It should be noted that the fixation probability, i.e., the probability that a given strategy eventually dominates the population as a result of stochastic evolutionary dynamics, is a primary quantity to be pursued in evolutionary dynamics in finite populations. Nevertheless, fixation trivially occurs in the presence of zealots if all zealots are assumed to take the same strategy; the zealots’ strategy always fixates. For example, if there is a single zealous cooperator in the population, cooperation always fixates even in the conventional prisoner’s dilemma game. However, in this adverse case, fixation of cooperation is expected to take long time; the relevant question here is the fixation time (Antal and Scheuring 2006; Traulsen et al. 2007; Altrock and Traulsen 2009; Altrock et al. 2010; Assaf and Mobilia 2010; Ewens 2010; Wu et al. 2010; Altrock et al. 2011; Assaf and Mobilia 2012; Kreindler and Young 2013). Here we examine the mean fixation time of the strategy selected by the zealots. This quantity serves as a probe to understand the extent to which zealots influence non-zealous players in the population. The fixation time would be affected by the payoff matrix, population size, number of zealous players, and strength of selection. We derive the asymptotic dependence of the mean fixation time on the population size when the fraction of zealots in the population is fixed. Mathematically, we extend the approach taken in Antal and Scheuring (2006) to the case with zealots.
Model
We assume a well-mixed population of players under evolutionary dynamics defined as follows. In each discrete time unit, each player selects either of the two strategies or . Each player plays a symmetric two-person game with all the other players in a unit time. The payoff matrix of the single game for the row player is given by
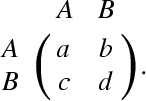 |
1 |
The fitness of a player on which the selection pressure operates is defined as the payoff summed over the opponents.
We assume that players may flip the strategy according to the Moran process (Moran 1958; Ewens 2010). We call these players the ordinary players. The other players are zealots that never change the strategy irrespectively of their fitness. Because our primary interest is in the possibility of cooperation in social dilemma games induced by zealous cooperators, we assume that all zealots take strategy ; is identified with cooperation in the case of a social dilemma game. We also assume that for the Moran process to be well-defined.
Because we have assumed a well-mixed population, the state of the evolutionary process is specified by the number of ordinary players selecting , which we denote by . In each time step, we select an ordinary player with the equal probability . The strategy of the selected player is updated. Then, we select a player, called the parent, whose strategy replaces that of the previously selected player. The parent is selected with the probability proportional to the fitness among the players including the zealots and the player whose strategy is to be replaced. The population size is constant over time. It should be noted that a player is updated once on average in time .
Because the zealots always select , the Moran process ends up with the unanimous population of players (we impose for this to be true). In other words, fixation of always occurs such that the issue of fixation probability is irrelevant to our model.
Results
We calculate the mean fixation time and its approximation in the case of a large population size by extending the framework developed in Antal and Scheuring (2006) (also see Van Kampen 2007; Redner 2001; Krapivsky et al. 2010; Ewens 2010).
Mean fixation time: exact solution
Consider the state of the population in which () ordinary players select strategy . A total of and players, including the zealots, select strategies and , respectively. The Moran process is equivalent to a random walk on the space in which is a reflecting boundary, and is the unique absorbing boundary.
The fitness of an and player is given by
| 2 |
and
| 3 |
respectively. In a single time step, increases by one, does not change, or decreases by one. We denote by and the probabilities that shifts to and , respectively. These probabilities are given by
| 4 |
and
| 5 |
We denote by the mean fixation time when there are initially ordinary players with strategy . As shown in Ewens (2010), pp. 86–91 (see Appendix A for a full derivation), we obtain
| 6 |
where
| 7 |
In Eq. (7), we interpret .
Deterministic approximation of the random walk
In this section we classify the deterministic dynamics driven by the expected bias of the random walk (i.e., ) into three cases, as is done in the analysis of populations without zealots (Taylor et al. 2004; Antal and Scheuring 2006). The obtained classification determines the dependence of the mean fixation time on , as we will show in Sect. 3.3.
We first identify the equilibrium points of the deterministic dynamics, i.e., satisfying . Equations (4) and (5) indicate that always yields , corresponding to the fact that is the unique absorbing state. Other equilibria are derived from
| 8 |
We set (), , and ignore terms in Eq. (8) to obtain
| 9 |
We define
| 10 |
| 11 |
| 12 |
and
| 13 |
We will use only when . In the continuous state limit, the deterministic dynamics driven by is classified into the following three cases, as summarized in Table 1. The derivation is shown in Appendix B.
-
Case (i): holds true for all () such that the dynamics starting from any initial condition tends to (Fig. 1a). In an infinite population, dominates . In a finite population, we expect that the fixation time is short. This case occurs when and one of the following conditions is satisfied:
- .
- and (i.e., ).
- , (i.e., and ), and .
- and (i.e., ).
-
Case(ii)
: has a unique solution () such that the dynamics starting from any initial condition converges to (Fig. 1b). In an infinite population, and coexist. In a finite population, we expect that the fixation time is long. This case occurs when .
-
Case (iii): has two solutions . Dynamics starting from converges to , and that starting from converges to (Fig. 1c). In an infinite population, a mixture of and and the pure configuration are bistable. In a finite population, we expect that the fixation time is long if the dynamics starts with and short if it starts with . This case occurs when
14 15
and16
are satisfied.17
Table 1.
Classification of the three cases of the mean fixation time when is large
| Case (i) | Case (i) or (iii) | |
| Case (ii) | Case (ii) |
Fig. 1.
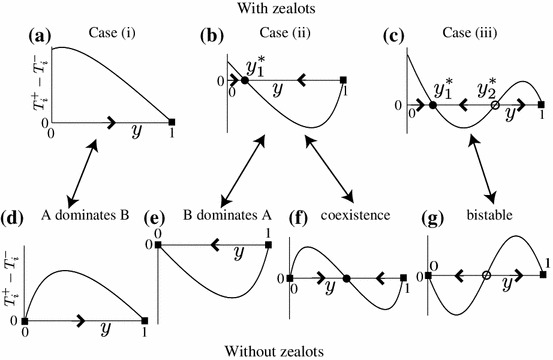
Schematic classification of the deterministic dynamics driven by . a–c Populations with zealots. d–g Populations without zealots. Filled and open circles represent stable and unstable equilibria, respectively. Filled squares represent the absorbing boundary condition. It should be noted that we identify
The condition given by Eq. (14) is related to the so-called cooperation facilitator assumed in a previous model (Mobilia 2012) as follows. Consider a hypothetical infinite population in which almost all players select , i.e., . Then, the payoff that a player with strategy gains by being matched with the other ordinary players and zealous players is equal to . The payoff that a player with strategy gains by being matched with the other ordinary players, but not zealous players, is equal to . Therefore, Eq. (14) represents the condition for the stability of the homogeneous population of strategy against invasion by when zealous players somehow contribute to the payoff of ordinary players and not to that of ordinary players. Such a zealous player is equivalent to the cooperation facilitator assumed in Mobilia (2012).
In the corresponding model without zealots, there are four scenarios: dominates (Fig. 1d), dominates (Fig. 1e), a mixture of and is stable (Fig. 1f), and and are bistable (Fig. 1g) (Antal and Scheuring 2006). The cases shown in Fig. 1d, f, and g are analogous to cases (i), (ii), and (iii), respectively, for the game with zealots. The case shown in Fig. 1e never occurs in the game with zealots because tends to increase in the absence of owing to the fact that unanimity of among the ordinary players is a reflecting boundary of our model. In fact, this case corresponds to case (ii) for the presence of zealots (Fig. 1b). If we set , we obtain case (i) when and , case (ii) when , and case (iii) when and . As is consistent with Antal and Scheuring (2006), the classification depends only on the and values. However, the scenario in which dominates (Fig. 1e) does not happen even with the vanishing density of zealots (i.e., ) because the unanimity of remains to be a reflecting boundary as long as there is at least one zealot.
Mean fixation time: large limit
In this section, we analyze the order of the mean fixation time in terms of when is large. We assume that the fraction of zealots in the population, i.e., , is fixed. Because the mean fixation time is by definition the largest for , i.e., the initial condition in which all ordinary players select , we focus on . To evaluate , we rewrite Eq. (6) for as
| 18 |
Case (i)
We obtain
| 19 |
where () is given by Eq. (9). In case (i), holds true. Therefore,
| 20 |
is satisfied for . By using Eq. (20), we obtain
| 21 |
Because the left-hand side of Eq. (21) is at least unity, we obtain
| 22 |
The substitution of and in Eq. (4) yields
| 23 |
In particular, we obtain
| 24 |
Equation (24) implies that
| 25 |
This result coincides with the previous result for the absence of zealots (Antal and Scheuring 2006).
Case (ii)
In Case (ii), for and for . Therefore, takes the minimum at . We denote the value of that satisfies and by . Such an exists if . If , we regard that . Using the relationship for a function () (Antal and Scheuring 2006) (also see Appendix C), we obtain
| 26 |
where
| 27 |
To derive the last line in Eq. (26), we used the steepest descent method (Antal and Scheuring 2006) (also see Appendix D).
Equations (23) and (24) imply that in Eq. (26) is safely ignored near the singularity at because it would contribute at most to the fixation time. Therefore, we obtain
| 28 |
where is a constant that depends on , and . The dependence of on is shown in Fig. 2 for sample payoff matrices for the prisoner’s dilemma game (solid line) and snowdrift game (dotted line). For both games, monotonically decreases with , implying that the fixation time decreases with . In particular, is equal to zero, which corresponds to , when is larger than a threshold value.
Fig. 2.
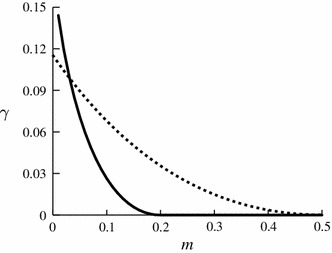
The exponent for the mean fixation time [Eq. (28)] plotted against the density of zealots for the prisoner’s dilemma game with , , , and (solid line) and the snowdrift game with , , , , with (dotted line). We calculated on the basis of Eqs. (26), (27), and (55)
Case (iii)
In this case, takes a local minimum at and a local maximum at . Therefore, behavior of the random walk in the range is qualitatively the same as that for case (ii), and that in the range is qualitatively the same as that for case (i). Because the former part makes the dominant contribution to the fixation time, the scaling of the mean fixation time is given by Eq. (28).
Case (iii) occurs when strategy is disadvantageous when it is rare and advantageous when it is frequent. The coordination game provides such an example (Sect. 5.4).
Summary and the borderline case
In summary, the mean fixation time in the limit of large is given by in case (i) and () in cases (ii) and (iii). For the parameter values on the boundary between the two scaling regimes, the same arguments as those for the model without zealots (Antal and Scheuring 2006) lead to .
Dependence of the mean fixation time on the selection strength
We examine the influence of the selection strength, denoted by , on the mean fixation time. To this end, we redefine the fitness to an and player by and , respectively, where and are given by Eqs. (2) and (3) (e.g., Nowak et al. 2004; Nowak 2006). Consequently, we replace the payoff matrix given by Eq. (1) by
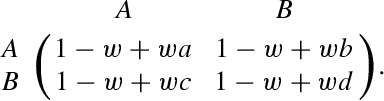 |
29 |
Equation (1) is reproduced with .
For sufficiently weak selection, we obtain , i.e., case (i), regardless of the payoff matrix. To prove this statement, we note that, by using the payoff matrix shown in Eq. (29), condition in the case of is generalized to
| 30 |
Therefore, if the original game in the case of belongs case (ii), i.e., , the game belongs to case (i) or (iii) (Table 1) if Eq. (30), or equivalently,
| 31 |
is satisfied. For a fixed payoff matrix, monotonically increases with , consistent with the intuition that existence of zealots would lessen the fixation time.
Next, the sign of is not affected by the selection strength. Therefore, we assume and prove that a condition for case (iii), i.e., Eq. (17), is violated with a sufficiently small . Because the value of given by Eq. (10) is also unaffected by , we start with assuming , which is a necessary condition for case (iii) [Eq. (16); see Appendix B]. The condition in the case of , where is defined by Eq. (11), is generalized to
| 32 |
Because the condition imposed on , which distinguishes cases (i) and (iii), is relevant only for (Table 1), Eq. (32) is satisfied for an arbitrary payoff matrix if
| 33 |
Therefore, case (iii) is excluded with a sufficiently small value.
The threshold value of below which , which we denote by , is given by
| 34 |
We can alternatively introduce the selection strength by replacing Eqs. (2) and (3) to redefine the fitness by
| 35 |
and
| 36 |
where is the selection strength (Traulsen et al. 2008). In Appendix E, we show that qualitatively the same result holds true in the sense that there is a threshold value of below which the fixation is fast irrespective of the , , , , and values. It should be noted that, with Eqs. (35) and (36), , , , and are allowed to take negative values.
Examples
We compare the mean fixation time for some games with that for the neutral game, i.e., . In the absence of zealots, the neural game yields (). The random walk is unbiased, and the so-called mean conditional fixation time is equal to (Antal and Scheuring 2006). The mean conditional fixation time is defined as the mean fixation time starting from state under the condition that the absorbing state at , not , is reached.
The neutral game in the presence of zealots yields and (). Therefore, the random walk is biased toward for all . More precisely, we obtain
| 37 |
As in Antal and Scheuring (2006), we say that fixation is fast (slow) if is smaller (larger) than the value given by Eq. (37). It should be noted that for the neutral game because it corresponds to .
Constant selection
As a first example, consider the case of frequency-independent selection such that and are equipped with fitness and 1 (under ), respectively. When and , the threshold selection strength below which , i.e., case (i), holds true is given by
| 38 |
If , case (ii) occurs. Even if is disadvantageous to , fixates fast with the help of zealots regardless of the selection strength if . This condition is more easily satisfied when is larger.
Prisoner’s dilemma game
Consider the prisoner’s dilemma game with a standard payoff matrix given by , , , and , where . Strategies and represent cooperation and defection, respectively. It should be noted that . With a general selection strength, the conditions derived in Sect. 3.2 imply that , i.e., case (i), if , and with case (ii) if . This condition coincides with that for the dominance of cooperators in the case of the infinite population (Masuda 2012).
The mean fixation time with and obtained by direct calculations of Eq. (18) is shown in Fig. 3a. In this and the following figures, the values are those normalized by that for the neutral game [Eq. (37)]. The behavior of is qualitatively different according to whether is larger or smaller than . If , the ratio of for the prisoner’s dilemma game to for the neutral game seems to approach a constant as . This is consistent with case (i). In contrast, if , grows rapidly, which is consistent with case (ii). To be more quantitative, divided by the value for the neutral game is shown by the dashed line in Fig. 3a. It should be noted that 400 is a constant for fitting and that value is theoretically determined as described in Sect. 3.3.2. The theory (dashed line) agrees well with the exact numerical results (thinnest solid line). We remark that the normalized behaves non-monotonically in ; it takes a minimum at an intermediate value of .
Fig. 3.
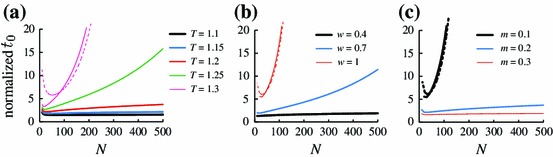
The normalized mean fixation time for the prisoner’s dilemma game as a function of . We set , , , and . In a, we set and . In b, we set and . In c, we set and . The dashed lines represent divided by the value for the neutral game
Next, to examine the effect of the selection strength, we set and . The mean fixation time as a function of and is shown in Fig. 3b. Equation (34) implies that when . Consistent with this result, grows fast as a function of when is large (i.e., and 1). In particular, for , normalized by the value for the neutral game (dashed line in Fig. 3b) agrees well with the exact results (thin solid line). For small (i.e., ), seems to scale with (thick solid line).
Figure 3c shows the dependence of on for different densities of zealots (i.e., ). It should be noted that the baseline value derived from the neutral game depends on the value of . Because we set and in Fig. 3c, the threshold value of is equal to 0.2. In fact, the normalized diverges according to when (dashed line and thick solid line), whereas it seems to converge to a constant value when (thin solid line).
Figure 3 indicates that for the prisoner’s dilemma game is always larger than that for the neutral game (i.e., the normalized is larger than unity). This is consistent with the intuition that cooperation is difficult to attain in the prisoner’s dilemma game as compared to the neutral game.
Finally, consider the symmetrized donation game, which is another standard form of the prisoner’s dilemma game, given by , , , and , where is the benefit, and is the cost. For the Moran process to be well-defined, we require , i.e., . For this payoff matrix, we obtain
| 39 |
Fixation occurs fast for a large benefit-to-cost ratio, large , or small selection strength.
Snowdrift game
In this section, we examine the snowdrift game (Maynard Smith 1982; Sugden 1986; Hauert and Doebeli 2004) defined by , , , and , where . Strategies an are identified as cooperation and defection, respectively. Each player is tempted to defect if the other player cooperates, as in the prisoner’s dilemma game. However, different from the prisoner’s dilemma game, a player is better off by cooperating if the partner defects; mutual defection is the worst outcome. In the infinite well-mixed population without zealots, the game has the unique mixed Nash equilibrium in which the fraction of cooperation is equal to .
Numerical evidence for the replicator dynamics, corresponding to an infinite population, suggests that cooperation is dominant if is large or is small (Masuda 2012). For the finite population, we obtain
| 40 |
If , we obtain , i.e., case (i). If , we obtain with case (ii). A large value of or makes the fixation time smaller. This result makes sense because a large generally favors cooperation.
Coordination game
The coordination game given by and has two pure Nash equilibria in the infinite well-mixed population without zealots. For a finite population in the presence of zealots, Eq. (34) yields
| 41 |
If , we obtain , i.e., case (i). It should be noted that any strength of selection yields if there are sufficiently many zealots, similar to the game with constant selection, prisoner’s dilemma game, and snowdrift game. If , we obtain with case (iii).
The mean first-passage time from state 0 (i.e., all ordinary players select ) to state , i.e., , is shown in Fig. 4. It should be noted that is equal to this first-passage time to exit . We set , , , , and . Equation (41) implies for these parameter values. Because , we obtain case (iii).
Fig. 4.
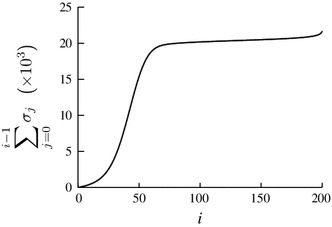
Mean first-passage time for the coordination game. We set , , , , and
The first-passage time increases slowly as increases when is small. It rapidly increases with for intermediate values of , Once the random walker passes the critical value, it feels a positive bias such that the first-passage time only gradually increases with for large . The values of that separate the three regimes are roughly consistent with the analytical estimates and [Eqs. (12), (13)]. It should be noted that the first-passage time shows representative behavior of case (iii) although is only slightly larger than .
Discussion
We extended the results for the fixation time under the Moran process (Antal and Scheuring 2006) to the case of a population with zealous players. Similar to the case without zealots (Antal and Scheuring 2006), we identified three regimes in terms of the payoff matrix, number of zealots, and selection strengths. In one regime, the fixation time is small (i.e., ). In the other two regimes, it is large (i.e., with ). We illustrated our results with representative games including the prisoner’s dilemma game, snowdrift game, and coordination game.
Zealots have several impacts on evolutionary dynamics in finite populations. First, fixation of one strategy always occurs with zealots because we assumed that all zealots permanently take . Second, there is a case in which fixation is fast if the fraction of players is sufficiently large, whereas fixation is slow if the fraction of is small. This scenario occurs for the coordination game. In the absence of zealots, the same game shows bistability such that the fixation to the unanimity of or that of occurs fast (Antal and Scheuring 2006). Third, for a selection strength smaller than a threshold value, the fixation is fast for any payoff matrix. In the absence of zealots, the dependence of the mean fixation time on for large values is completely determined by the signs of and (Antal and Scheuring 2006). Therefore, the scaling of the mean fixation time on is independent of the selection strength because manipulating the selection strength does not change the sign of the effective or value. If the payoff matrix is given in the slow fixation regime, the fixation is exponentially slow even for a small selection strength. In contrast, in the presence of zealots, slow fixation can be accelerated if we lessen the selection strength.
Mobilia examined the prisoner’s dilemma game with cooperation facilitators (Mobilia 2012). A cooperation facilitator was assumed to cooperate with cooperators and not to play with defectors. The cooperation facilitator and zealous cooperator in the present study are common in that they never change the strategy. However, they are different. First, zealous cooperators are embedded in a well-mixed population such that they myopically cooperate with defectors as well as cooperators. Second, the ordinary players may imitate the zealous cooperator’s strategy (i.e., cooperation). In contrast, players do not imitate the cooperation facilitator’s strategy (i.e., cooperation) in Mobilia’s model. As a consequence, cooperation does not always fixate in his model.
Examination of the case of imperfect zealots, in which zealots change the strategy with a small probability (Masuda 2012), warrants future work.
Acknowledgments
We thank Bin Wu for carefully reading the manuscript. NM acknowledges the support provided through Grants-in-Aid for Scientific Research (No. 23681033) from MEXT, Japan, the Nakajima Foundation, CREST JST, and the Aihara Innovative Mathematical Modelling Project, the Japan Society for the Promotion of Science (JSPS) through the “Funding Program for World-Leading Innovative R&D on Science and Technology (FIRST Program),” initiated by the Council for Science and Technology Policy (CSTP).
Appendix A: Derivation of Eq. (6)
Denote by the probability that the random walker starting from state at time 0 is absorbed to state at time . The normalization is given by . It should be noted that and (). The mean fixation time when ordinary players initially select strategy is given by
| 42 |
It should be noted that .
satisfies the recursion relation given by
| 43 |
By multiplying both sides of Eq. (43) by and taking the summation over , we obtain
| 44 |
In terms of , Eq. (44) can be rewritten as
| 45 |
The solution of Eq. (45) is given by
| 46 |
where and is given by Eq. (7).
We set in Eq. (44) and use to obtain
| 47 |
Therefore, we obtain
| 48 |
Using Eq. (48), we reduce Eq. (46) to
| 49 |
The mean fixation time is given by
| 50 |
Appendix B: Classification of the deterministic dynamics induced by the biased random walk
B.1 When
We obtain for . Because
| 51 |
| 52 |
where we used the assumption in Eq. (51), we distinguish the following two cases. If , is satisfied for , yielding case (i) in the main text. If , a certain exists such that for , and for . Therefore, case (ii) occurs.
B.2 When
We obtain for . In this situation, Eq. (51) holds true.
If , i.e., , a certain () exists such that for , and for . Therefore, case (ii) occurs.
Suppose that , i.e., . To analyze this case, let us write
| 53 |
where
| 54 |
-
(i)
If , i.e., , we obtain for . Therefore, case (i) occurs.
-
(ii)
If , i.e., , then , yielding case (i).
-
(iii)If , we have the following two subcases:
- If , has two solutions . In the deterministic dynamics driven by the bias , and are stable and unstable, respectively. Therefore, case (iii) occurs.
- If , we obtain for all , where the equality holds true only when and . Therefore, case (i) occurs.
B.3 When
The quadratic term in disappears when . The classification of the dynamics in this case coincides with that for .
Appendix C: Derivation of
To derive the relationship , we write
| 55 |
where and . Because the integral on the right-hand side of Eq. (55) is independent of , we obtain . It should be noted that is consistent with .
Appendix D: Steepest descent method
As done in Antal and Scheuring (2006), we use the steepest descent method to evaluate in Eq. (26) as follows:
| 56 |
where . We approximate the integral by a Gaussian integral to obtain
| 57 |
with and such that
| 58 |
Appendix E: Weak selection introduced via an exponential function leads to fast fixation
Assume that the fitness of an A and B player is given by Eqs. (35) and (36), respectively. Then, we obtain
| 59 |
If , i.e.,
| 60 |
holds true for any () and , the fixation occurs fast (i.e., ). By substituting and in Eq. (60) and ignoring terms, we obtain
| 61 |
Because the right-hand side of Eq. (61) is positive, there exists such that when . It should be noted that, in contrast to the assumption throughout the present article, , , , and are allowed to be negative in the present analysis because and given by Eqs. (35) and (36) are positive irrespective of the , , , and values.
References
- Altrock PM, Traulsen A. Fixation times in evolutionary games under weak selection. New J Phys. 2009;11:013012. doi: 10.1088/1367-2630/11/1/013012. [DOI] [Google Scholar]
- Altrock PM, Gokhale CS, Traulsen A. Stochastic slowdown in evolutionary processes. Phys Rev E. 2010;82:011925. doi: 10.1103/PhysRevE.82.011925. [DOI] [PubMed] [Google Scholar]
- Altrock PM, Traulsen A, Reed FA. Stability properties of underdominance in finite subdivided populations. PLOS Comput Biol. 2011;7:e1002260. doi: 10.1371/journal.pcbi.1002260. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Antal T, Scheuring I. Fixation of strategies for an evolutionary game in finite populations. Bull Math Biol. 2006;68:1923–1944. doi: 10.1007/s11538-006-9061-4. [DOI] [PubMed] [Google Scholar]
- Assaf M, Mobilia M. Large fluctuations and fixation in evolutionary games. J Stat Mech. 2010;2010:P09009. [Google Scholar]
- Assaf M, Mobilia M. Metastability and anomalous fixation in evolutionary games on scale-free networks. Phys Rev Lett. 2012;109:188701. doi: 10.1103/PhysRevLett.109.188701. [DOI] [PubMed] [Google Scholar]
- Ewens WJ. Mathematical population genetics I. Theoretical introduction. New York: Springer; 2010. [Google Scholar]
- Fu F, Rosenbloom DI, Wang L, Nowak MA. Imitation dynamics of vaccination behaviour on social networks. Proc R Soc B. 2011;278:42–49. doi: 10.1098/rspb.2010.1107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galam S, Jacobs F. The role of inflexible minorities in the breaking of democratic opinion dynamics. Physica A. 2007;381:366–376. doi: 10.1016/j.physa.2007.03.034. [DOI] [Google Scholar]
- Hauert C, Doebeli M. Spatial structure often inhibits the evolution of cooperation in the snowdrift game. Nature. 2004;428:643–646. doi: 10.1038/nature02360. [DOI] [PubMed] [Google Scholar]
- Van Kampen NG. Stochastic processes in physics and chemistry. 3. Netherlands: Elsevier; 2007. [Google Scholar]
- Krapivsky PL, Redner S, Ben-Naim E. A kinetic view of statistical physics. Cambridge: Cambridge University Press; 2010. [Google Scholar]
- Kreindler GE, Young HP. Fast convergence in evolutionary equilibrium selection. Games Econ Behav. 2013;80:39–67. doi: 10.1016/j.geb.2013.02.004. [DOI] [Google Scholar]
- Liu XT, Wu ZX, Zhang L. Impact of committed individuals on vaccination behavior. Phys Rev E. 2012;86:051132. doi: 10.1103/PhysRevE.86.051132. [DOI] [PubMed] [Google Scholar]
- Masuda N. Evolution of cooperation driven by zealots. Sci Rep. 2012;2:646. doi: 10.1038/srep00646. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maynard Smith J. Evolution and the theory of games. Cambridge: Cambridge University Press; 1982. [Google Scholar]
- Mobilia M. Does a single zealot affect an infinite group of voters? Phys Rev Lett. 2003;91:028701. doi: 10.1103/PhysRevLett.91.028701. [DOI] [PubMed] [Google Scholar]
- Mobilia M. Stochastic dynamics of the prisoner’s dilemma with cooperation facilitators. Phys Rev E. 2012;86:011134. doi: 10.1103/PhysRevE.86.011134. [DOI] [PubMed] [Google Scholar]
- Mobilia M, Petersen A, Redner S (2007) On the role of zealotry in the voter model. J Stat Mech: P08029
- Moran PAP. Random processes in genetics. Proc Cambridge Philos Soc. 1958;54:60–71. doi: 10.1017/S0305004100033193. [DOI] [Google Scholar]
- Nowak MA, Sasaki A, Taylor C, Fudenberg D. Emergence of cooperation and evolutionary stability in finite populations. Nature. 2004;428:646–650. doi: 10.1038/nature02414. [DOI] [PubMed] [Google Scholar]
- Nowak MA. Evolutionary dynamics. MA: The Belknap Press of Harvard University Press; 2006. [Google Scholar]
- Redner S. A guide to first-passage processes. Cambridge: Cambridge University Press; 2001. [Google Scholar]
- Singh P, Sreenivasan S, Szymanski BK, Korniss G. Accelerating consensus on coevolving networks: the effect of committed individuals. Phys Rev E. 2012;85:046104. doi: 10.1103/PhysRevE.85.046104. [DOI] [PubMed] [Google Scholar]
- Sugden R. The economics of rights, co-operation and welfare. New York: Blackwell; 1986. [Google Scholar]
- Taylor C, Fudenberg D, Sasaki A, Nowak MA. Evolutionary game dynamics in finite populations. Bull Math Biol. 2004;66:1621–1644. doi: 10.1016/j.bulm.2004.03.004. [DOI] [PubMed] [Google Scholar]
- Traulsen A, Pacheco JM, Nowak MA. Pairwise comparison and selection temperature in evolutionary game dynamics. J Theor Biol. 2007;246:522–529. doi: 10.1016/j.jtbi.2007.01.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Traulsen A, Shoresh N, Nowak MA. Analytical results for individual and group selection of any intensity. Bull Math Biol. 2008;70:1410–1424. doi: 10.1007/s11538-008-9305-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu B, Altrock PM, Wang L, Traulsen A. Universality of weak selection. Phys Rev E. 2010;82:046106. doi: 10.1103/PhysRevE.82.046106. [DOI] [PubMed] [Google Scholar]
- Xie J, Sreenivasan S, Korniss G, Zhang W, Lim C, Szymanski BK. Social consensus through the influence of committed minorities. Phys Rev E. 2011;84:011130. doi: 10.1103/PhysRevE.84.011130. [DOI] [PubMed] [Google Scholar]