

Abstract
Purpose.
Intramuscular innervation of horizontal rectus extraocular muscle (EOMs) is segregated into superior and inferior (transverse) compartments, whereas all EOMs are also divided into global (GL) and orbital (OL) layers with scleral and pulley insertions, respectively. Mechanical independence between both types of compartments has been demonstrated during passive tensile loading. We examined coupling between EOM compartments during active, ex vivo contraction.
Methods.
Fresh bovine EOMs were removed, and one compartment of each was coated with hydrophobic petrolatum. Contraction of the uncoated compartment was induced by immersion in a solution of 50 mM CaCl2 at 38°C labeled with sodium fluorescein dye, whereas tensions in both compartments were monitored by strain gauges. Control experiments omitted petrolatum so that the entire EOM contracted. After physiological experiments, EOMs were sectioned transversely to demonstrate specificity of CaCl2 permeation by yellow fluorescence dye excited by blue light.
Results.
In control experiments without petrolatum, both transverse and GL and OL compartments contracted similarly. Selective compartmental omission of petrolatum caused markedly independent compartmental contraction whether measured at the GL or the OL insertions or for transverse compartments at the scleral insertion. Although some CaCl2 spread occurred, mean (±SD) tension in the coated compartments averaged only 10.5 ± 3.3% and 6.0 ± 1.5% in GL/OL and transverse compartments, respectively relative to uncoated compartments. Fluorescein penetration confirmed selective CaCl2 permeation.
Conclusions.
These data confirm passive tensile findings of mechanical independence of EOM compartments and extend results to active contraction. EOMs behave actively as if composed of mechanically independent parallel fiber bundles having different insertional targets, consistent with the active pulley and transverse compartmental hypotheses.
Keywords: active pulley hypothesis, compartmental hypothesis, extraocular muscle
The orbital, global, and transverse compartments of individual bovine extraocular muscles generate tension independently of one another during ex vivo calcium activation. This finding, the result of parallel fibers in extraocular muscle, supports a postulate essential to the active pulley and compartmental hypotheses.
Introduction
In individual skeletal muscles, there may exist multiple neuromuscular compartments controlled by different sets of motor neurons.1–3 For example, the transversus abdominis,3 cricothyroid,4 and triceps brachii5 muscles each contain compartments innervated independently by separate motor nerve branches. Extraocular muscle (EOM) has numerous motor units (defined as sets of muscle fibers innervated by a single motor neuron), seemingly in excess of what would be necessary if all of the motor units within each individual EOM functioned similarly.6,7 Evidence has been accumulating, however, that motor units within individual EOMs may not all function in lock-step fashion. Peng et al.8 traced the intramuscular arborization of the abducens nerve within the lateral rectus (LR) muscles of monkey and human, and found that the nerve bifurcates to arborize within nonoverlapping superior and inferior zones throughout the EOM's length. Costa et al.9 confirmed the report by Peng et al.8 of compartmentalized LR innervation and extended the finding by tridimensional nerve reconstruction to the medial rectus (MR), and partially to the inferior rectus (IR) but not the superior rectus (SR) muscles of humans and monkeys. In both species, the LR and MR motor nerves bifurcate into dual, nonoverlapping divisions that innervate superior and inferior zones containing approximately equal numbers of EOM fibers. In contrast, motor nerve arborizations were partially overlapped in IR but highly mixed throughout the SR. It has been proposed that differential innervation in horizontal rectus zones can potentially mediate previously unrecognized vertical oculorotary actions. Although Costa et al.9 did not suggest that the dual compartments of the horizontal rectus EOMs are always differentially activated, the authors proposed that differential innervation might occur under some physiological conditions. Recently, functional evidence for differential compartmental activation of the human horizontal rectus EOMs was obtained from magnetic resonance imaging during ocular counter-rolling induced by head tilt,10 convergence,11 and vertical fusional vergence.12
Another compartmentalization of EOMs into global (GL) and orbital (OL) layers, orthogonal to the former transverse compartment, has also been described. The GL is the oculorotary muscle layer continuous from the annulus of Zinn to the scleral insertion. The active pulley hypothesis (APH) explains that the OL instead inserts on a pulley connective tissue ring posterior to the globe through which pass the GL fibers that, in turn, insert on the sclera to rotate the eye.13 Contraction of the OL causes the pulley to retract along the EOM to maintain a constant distance from the scleral insertion. Because this behavior is quantitatively similar for all four rectus EOMs, resulting shifts in pulley positions with gaze direction alter EOM pulling directions by half the change in ocular orientation to implement Listing's law of ocular torsion.13–17 The APH assumes at least some mechanical independence between GL and OL.
Shin et al.18 used passive tensile elongation to demonstrate the independent mechanical behavior of ex vivo bovine EOM compartments. Using a dual-channel load cell, the experiment stretched one compartment of each EOM specimen, while the other compartment remained stationary, and forces in both channels were monitored. Compartments were operationally defined as arbitrary proportions of GL muscle or tendon fibers for transverse experiments or the OL versus GL at their respective insertions. Shin et al.18 showed that all rectus muscles and both oblique EOMs exhibited marked compartmental independence during tensile loading, consistent with separately controllable actions as required by the active pulley and transverse compartmental hypotheses. However, although passive tensile loading reflects the entire biomechanical repertoire of extraocular tendon, passive loading by the antagonist reflects only a portion of an EOM's biomechanical activity. It remains to be demonstrated whether compartments, defined by groupings of fibers within the same EOM, can generate differential tensions and transmit them to external loads.
Interaction of myosin and actin in the presence of ATP represents the essential mechanism of muscle contraction,19 and calcium ion is a critical regulatory and signaling molecule. Calcium ion content is 40-fold higher in EOM than in limb muscle.20 Because ionic calcium can induce EOM contraction, calcium was used in this study to induce ex vivo contraction.
This study used calcium-induced ex vivo contraction to investigate the potential for independent mechanical action of EOM compartments. If substantial coupling were demonstrated between EOM compartments during active contraction, the biomechanical basis of the APH and transverse compartmental hypotheses would be undermined. Conversely, substantial contractile mechanical independence between compartments would provide the prerequisite for the APH and transverse compartmental hypotheses.
Methods
Dual Channel Contraction Testing Apparatus
A dual-channel load cell was assembled, which consisted of two strain gauges (Grass Technology, Warwick, RI, USA) having 0.02 mN minimum resolution, vertically mounted using individual vernier height adjustments to measure isometric contraction force (Fig. 1). Each strain gauge was calibrated against a 10-g weight before every experiment. The lower, origin end of the EOM specimen was placed in a broad, spring steel clamp to avoid rotation artifacts, connected to a heavy anchor whose weight markedly exceeded all forces that might be generated by the EOM. The upper ends of each portion of the specimen were connected via two metal hooks to strain gauges using fine, stiff monofilament nylon with breaking strength exceeding 200 N. After conditioning by a bridge amplifier, the output of each strain gauge channel was low-pass filtered and digitally sampled at 10 Hz and 32-bit precision using LabVIEW (National Instruments Corp., Austin, TX, USA). Specimens were initially positioned in an empty glass beaker that was then filled with calcium solution preheated to 40°C to allow for cooling during the experiment.
Figure 1.

Dual-channel contraction testing apparatus. In each adjacent channel, two point sutures (2) connect the strain gauge (1) to the EOM compartment (3). The common ends of the EOM were clamped (4) to a heavy anchoring weight resting on the beaker's bottom (5). Contraction was initiated by filling the beaker with calcium chloride solution immersing the specimen. Finely adjustable screw jack stands permitted separate preloading of each channel.
Calcium Depolarization of EOM
Preliminary experiments were performed to confirm that CaCl2 solution could induce ex vivo contraction of bovine EOM. To optimize conditions, separate EOM specimens were immersed in CaCl2 concentrations of 5, 50, and 100 mM as resulting length change was monitored for 1 hour. Every EOM immersed in at least 50 mM CaCl2 solution contracted more than 20% of initial length. Figure 2 shows an example of LR muscle that contracted 28% of initial length in 1 hour. Based upon this experiment, CaCl2 concentration for further experiments was set at 50 mM.
Figure 2.

Effect of CaCl2 solution-induced contraction on bovine LR muscle. The LR was immersed in 50 mM CaCl2. After 1 hour, LR length was reduced by 28% of initial length. The specimen was not fixed. Clamps are included in the photograph as distance reference only.
Compartment Isolation
In the present experiment, one EOM compartment could be exposed to CaCl2 solution to stimulate active contraction, whereas isolation of the other compartment was achieve with a water-repellent white petrolatum coating (Comforts, Kroger Corp., Cincinnati, OH, USA) applied before immersion. To verify specificity of compartmental isolation, aqueous-soluble fluorescein sodium dye (acid yellow [C20H10O5Na2]; Sigma Chemical Corp., St. Louis, MO, USA) was added at 0.4 mM to the CaCl2 solution. Fluorescein emits an apple-green fluorescence when excited by blue light. After immersion experiments, specimens, either whole or transected, were photographed under blue light-emitting diode illumination by a high-resolution camera with a 50-mm macro lens (EOS 5D Mark II model; Canon, Tokyo, Japan) equipped with a yellow filter. In both transverse and GL/OL compartment experiments, fluorescein penetration was detected mostly in the exposed compartment, and little fluorescein was evident in the isolated compartment, especially deep within the EOM. Figure 3 illustrates the arrangement for testing transverse and GL/OL compartmentalization and isolation of each compartment typically achieved.
Figure 3.

Isolation of transverse and GL/OL compartments for activation by 50 mM CaCl2 labeled with 0.4 mM sodium fluorescein dye. Inferior (A) and OL (B) compartments were coated with water-repellent white petrolatum. (C, D) Cross-sections of each compartment photographed under blue light using a yellow filter to demonstrate selective penetration of the fluorescein-labeled solution into the uncoated compartment. After immersion in fluorescein-labeled CaCl2 solution, superior ([C], left) and GL ([D], bottom) compartments were permeated with fluorescein marker, but inferior ([C], right) and OL ([D], top) compartments showed at most only minimal fluorescence.
Experimental Procedure
As chosen, 1 GL or OL or, alternatively, 1 transverse compartment of a freshly excised bovine EOM was coated with petrolatum to resist penetration by aqueous CaCl2 solution. Prior to clamping, both broad surfaces of each specimen end were fixed using cyanoacrylate glue between thin cardboard layers to form an anchor that was placed in the clamp. Then, one end of each compartment of the specimen was attached via monofilament line, light but stiff nylon (200-N strength), knotted to a stainless steel hook to one channel of a sensitive strain gauge, while the common end of the specimen was anchored to a heavy mass. The specimen and mass were placed in a glass beaker. Because contraction force cannot be measured by the strain gauge if initial slack exists, preloading of approximately 5-g force (gf) was applied by elongation of the specimen ends at each strain gauge. Fluorescein-labeled 50 mM CaCl2 solution prewarmed to 38°C was poured into the glass beaker, creating a slight reduction in tension due to specimen buoyancy. Thus, contraction of only one EOM compartment was activated for both the transverse and GL/OL compartment experiments, while the petrolatum-coated compartment was activated only minimally by what was assumed to be diffusion greatly hindered by the petrolatum coating. Force recording was performed continuously by the dual strain gauges.
EOM specimens displaced aqueous solution by their volumes. Due to resulting buoyancy, reduced tension was detected immediately after EOM immersion in both strain gauges, followed by increased tension due to EOM contraction. Buoyancy of each of the transverse compartments was expected to be roughly similar because transverse dimensions were set to be equal. However, because transverse cross-sections are not symmetrical, modest variation in buoyancy of transverse compartments was anticipated because magnetic resonance imaging has demonstrated 15% to 20% differences in the volumes of the similarly defined compartments of human horizontal rectus EOMs.10 Larger differences in buoyancy were anticipated between the GL and OL compartments because the OL is not only shorter than the GL but contains fewer fibers21 and so has considerably less volume. Data were collected during 5 minutes for each experiment, although analysis was ultimately confined to the first 30 seconds. Because permeation of ionic solution between compartments, or through gaps in the petrolatum, eventually caused undesired contraction in the petrolatum-coated compartment, force data in the early 30 seconds were used to calculate the degree of mechanical independence between compartments.
Results
Transverse Compartment Coupling
The degree of mechanical coupling of actively generated, internal force between the transverse compartments of 5 specimens was tested using 50 mM CaCl2. Representative specimen photographs and time series data are shown in Figure 4. Because the strain gauge and specimen were aligned vertically, positive values indicated contraction and negatives values indicated relative relaxation. Because of the eventual permeation of ionic solution between compartments, force data in early 30 seconds was analyzed. For calculating the degree of compartmental force coupling, the ratio of force increment in each compartment was calculated. Theoretically, 0% implies complete independence, although 100% implies complete coupling between compartments. All experimental data exhibited <10% force coupling, reflecting a high degree of compartmental independence during active contraction. For 5 specimens, the mean value (±SD) at 30 seconds was 6.03 ± 1.53%.
Figure 4.

Transverse compartment testing of bovine lateral rectus muscle. (A) En face view of specimen with inferior compartment isolated using water-repellent petrolatum coating. (B) Measured force is shown in each compartment after 5 minutes following CaCl2 immersion. Initial force reduction represents buoyancy effect of immersion in the aqueous solution; the difference in initial force reduction is attributable to differences in the volumes of the two compartments. (C) Time expanded time scale view. Force data have been shifted vertically to align the ordinates at 90 seconds for trend comparison.
GL and OL Compartment Coupling
Similar to transverse compartment experiments, GL and OL compartment experiments were performed to evaluate intercompartmental mechanical coupling. During previous experiments involving passive external loading, elongation of either the GL or OL symmetrically produced the same force coupling to the other compartment.18 Therefore, only the GL was activated by using CaCl2 in the present experiments, because the OL has fewer and shorter fibers than the GL and petrolatum coating readily isolated the superficially located OL, whereas the GL was not easily isolated. Figure 5 shows GL/OL specimen placement and resulting force time series data for the LR. Note that force in the contracting GL was rapidly increasing at the same time that force in the isolated OL was decreasing slightly. The dynamic trends in the two layers were opposite. Five EOM specimens were tested by GL activation during OL isolation, and in these tests, the coupling ratio averaged 10.50 ± 3.28%.
Figure 5.

Global/orbital compartment testing of LR muscle. (A) Transverse view of LR specimen with the orbital layer (OL) compartment isolated by petrolatum coating. (B) Force behavior of each compartment is shown after 5 minutes. Initial force reduction represents the buoyancy effect that was smaller for the OL because of its smaller volume. (C) Time expanded view beginning at 170 seconds.
Control Experiments Without Compartment Isolation
Control experiments were performed in order to validate the effectiveness of the petrolatum isolation. Six EOM specimens were tested from LR, SR, and IR muscles. For both transverse and GL/OL compartment experiments, whole EOM without petrolatum coating was immersed in 50 mM CaCl2 during dual-channel force recording for 5 minutes. Figure 6 shows resulting force in transverse and GL/OL compartment specimens. In the transverse compartment experiment (Fig. 6A), both inferior and superior compartments showed similar contraction force behavior, suggesting that the calcium solution evenly penetrated whole-EOM tissue and led to contraction in the roughly equal sized compartments. However, for the case of GL/OL compartment experiment (Fig. 6B), data showed different magnitudes and temporal patterns of force in the GL and OL compartments. The anatomically larger GL compartment contracted to a higher force level than the smaller OL compartment.
Figure 6.
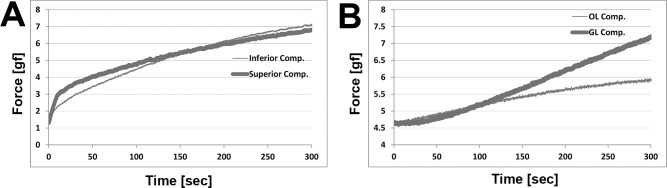
Control experiment with simultaneous calcium activation of transverse (A) and GL/OL (B) compartments. (A) Transverse compartment forces are shown for superior rectus. (B) Global/orbital compartment forces are shown for inferior rectus.
Dynamics of Compartmental Behavior
The ratio of force increments in each compartment was used as an indicator of compartmental independence during active contraction, and no values greatly exceeded 10%. This implies considerable independence between transverse and GL/OL compartments. However, the dynamics of active contraction provide further evidence of compartment independence. Figure 7 shows the contractile behaviors of each compartment in transverse and GL/OL compartment experiments. In each figure, the arrows indicate the time interval between onset of contraction in each compartment following force reduction due to buoyancy. During this period, tension in the two compartments exhibited opposite trends: increase in superior and GL compartments but decrease in inferior and OL compartments, representing totally opposite, independent compartmental mechanical behavior.
Figure 7.
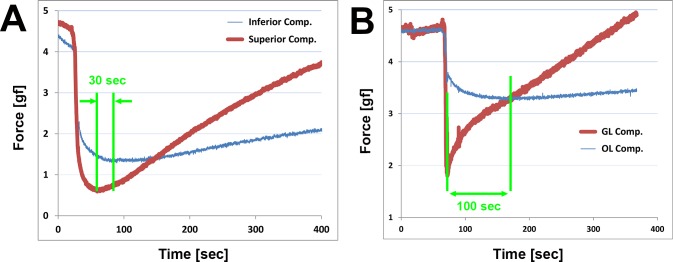
Active contraction in superior (A) and GL (B) compartments. Initiation of a positive slope in the force curves indicates contraction exceeding buoyancy. In each figure, the arrow indicates the time interval between asynchronous onset of contractions in each compartment. (A) Transverse compartment force shows an approximately 30-second interval between onset of force increase in the superior compartment first and inferior compartment later. (B) Time interval of approximately 100 seconds between increases in force first in the GL, followed by onset of smaller force increases in the OL. During the interval, tension in the two compartments exhibited opposite trends.
Discussion
Although induction of individual muscle fiber contraction by CaCl2 has been reported in several studies,20,22,23 macro-scale contraction shortening of whole EOM has yet to be reported. A 50 mM CaCl2 concentration is effective for active compartmental contraction experiments. Fresh EOM was required to obtain useful experimental data. In this study, elapsed time for every experiment was kept within 2 hours from animal slaughter, including transport and preparation time, in order to minimize rigor mortis, which is muscle stiffening beginning after 3 to 4 hours, reaching maximum at 12 hours and gradually dissipating from approximately 24 hours after death.24 Although the EOM contraction reported here induced by CaCl2 was not physiological, it nevertheless represents active force generation.
Multiple lines of biomechanical data here indicate that regions of EOMs and their tendons have a substantial degree of mechanical independence during active contraction. Not only was the force in the ionically isolated transverse compartment only approximately 6% of that in the chemically activated fellow compartment, the time course of force changes in the two layers differed dynamically. Calcium-activated contractile force in the GL was only approximately 10% transmitted to the OL, again with different dynamic changes in the two layers. Moreover, the foregoing values of intercompartmental force coupling are likely to represent upper boundaries for this phenomenon, because any imperfections in chemical isolation of “noncontracting” compartments would have resulted in some non-zero level of contraction. During physiological EOM activation in vivo, contraction would presumably be exquisitely specific to the motor units activated individually. These results are consistent with mechanical independence among orthotropic EOM and tendon compartments demonstrated during passive tensile elongation, yet uncharacteristic of isotropic material such as latex rubber.18 Structure of EOM and tendon underlies compartmental mechanical independence.
Both passive stretching and active contraction occur in EOMs during agonist and antagonist action. Passive stretching was replicated by tensile elongation in a previous study,18 whereas in the current study, active contraction was induced by CaCl2 depolarization. Highly independent contractile force in the transverse and GL/OL compartments was similar to that during passive tensile loading.
Because both passive and active compartmental independence has now been demonstrated in EOM, it is plausible that selective compartmental intramuscular innervation could induce separate mechanical behaviors. Horizontal rectus EOMs of humans, monkeys, and other mammals exhibit two compartments,8,9 so the groups of EOM fibers separately innervated by the superior and inferior motor nerve trunks can be separately controlled. The present demonstration of biomechanical independence implies that groups of horizontal rectus EOM and tendon fibers may constitute functionally independent superior and inferior muscle actuators whose tensions are delivered to separate insertions on the ocular sclera. In vivo magnetic resonance imaging has provided further evidence for independent compartmental behavior. During ocular counter-rolling, the superior versus inferior compartments of the human LR contract differently.10,25 Selective compartmental function in MR and to a lesser extent LR may also contribute to vertical duction in humans.26 Moreover, the superior MR compartment exhibits greater contractility than other horizontal rectus compartments during convergence.11
Independent compartmental behavior is also supported by significantly different electromyographic activities in the GL and OL of the human MR.27 The current demonstration of independent compartmental behavior easily satisfies the requirement of the APH, so that the OL can control pulley connective tissues to adjust EOM pulling direction, while the GL can generate sometimes different oculorotary tension to rotate the globe. Structural,13 electrophysiological,14 hemodynamic,15 metabolic,16 and genetic expression17 differences between GL and OL are also consistent with compartmental independence.
The anisotropy of EOM comes from the roughly parallel structure of EOM fibers, which is probably also the reason for mechanical compartmental independence. Fibers of EOM are arranged into long, roughly parallel fascicles. Anatomical studies using serial EOM sections stained by Mason trichrome in human and monkey have demonstrated long EOM fibers with relatively few fiber-to-fiber junctions,28,29 although other anatomical studies have reported more myomyous junctions in species such as cat,30 rabbit,31 and squirrel monkey.32 Notwithstanding some anatomical couplings, these structural observations cannot refute repeated biomechanical studies demonstrating independent mechanical compartmental behavior.
The recent hypothesis of transverse neuromuscular compartmental innervation of horizontal rectus EOMs is tenable in light of the current demonstration of compartmental biomechanical independence. However, because biomechanical independence is only a necessary but not a sufficient condition for the independent neuromuscular compartmental behavior in EOMs, the degree of selective innervational control of EOM compartments must be established under additional relevant physiological conditions. In other words, the anatomical and physiological capacity for independent compartmental control is analogous to a string quartet: the capacity of each instrument to play a different score does not preclude playing in unison if the composer desires it. During many but not all physiologic eye movement behaviors, compartments of the same EOM probably act in unison. Conversely, just as a defective quartet might suffer the absence of a disabled viola player, one compartment of an EOM could experience compartmental denervation and thus lose function in a single compartment. Such differential compartmental pathology is suggested by the frequent finding of selective atrophy of the superior portion of the LR in abducens palsy.33 The recent demonstration of selective intramuscular innervation in the superior oblique muscle apparently selective for compartmental advantage to vertical versus torsional action may extend the compartmental principle to the traditional cyclovertical EOMs.34
Acknowledgments
We thank Howard Ying, MD, PhD, for helpful suggestions about calcium activation of muscle contraction. We also thank Manning Beef, LLC (Pico Rivera, CA, USA) for its generous contribution of bovine specimens; Jose Martinez, Claudia Tamayo, and Ramiro Carlos for assistance with specimen preparation; and Alan Le for assistance with photography and preparation of specimens for experiments.
Supported by the US Public Health Service, National Eye Institute Grants EY08313 and EY0331, and an Unrestricted Grant from Research to Prevent Blindness. JLD is a Leonard Apt Professor of Ophthalmology.
Disclosure: A. Shin, None; L. Yoo, None; J.L. Demer, None
References
- 1. English AW, Wolf SL, Segal RL. Compartmentalization of muscles and their motor nuclei: the partitioning hypothesis. Phys Ther. 1993; 73: 857–867. [DOI] [PubMed] [Google Scholar]
- 2. Holtermann A, Roeleveld K, Mork PJ, et al. Selective activation of neuromuscular compartments within the human trapezius muscle. J Electromyogr Kinesiol. 2009; 19: 896–902. [DOI] [PubMed] [Google Scholar]
- 3. Urquhart DM, Hodges PW. Differential activity of regions of transversus abdominis during trunk rotation. Eur Spine. 2005; 14: 393–400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4. Mu L, Sanders I. The human cricothyroid muscle: three muscle bellies and their innervation patterns. J Voice. 2009; 23: 21–28. [DOI] [PubMed] [Google Scholar]
- 5. Lucas-Osma AM, Collazos-Castro JE. Compartmentalization in the triceps brachii motoneuron nucleus and its relation to muscle architecture. J Comp Neurol. 2009; 516: 226–239. [DOI] [PubMed] [Google Scholar]
- 6. Goldberg SJ, Wilson KE, Shall MS. Summation of extraocular motor unit tensions in the lateral rectus muscle of the cat. Muscle Nerve. 1997; 20: 1229–1235. [DOI] [PubMed] [Google Scholar]
- 7. Goldberg SJ, Meredith MA, Shall MS. Extraocular motor unit and whole-muscle responses in the lateral rectus muscle of the squirrel monkey. J Neurosci. 1998; 18: 10629–10639. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8. Peng M, Poukens V, da Silva Costa RM, Yoo L, Tychsen L, Demer JL. Compartmentalized innervation of primate lateral rectus muscle. Invest Ophthalmol Vis Sci. 2010; 51: 4612–4617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9. da Silva Costa RM, Kung J, Poukens V, Yoo L, Tychsen L, Demer JL. Intramuscular innervation of primate extraocular muscles: unique compartmentalization in horizontal recti. Invest Ophthalmol Vis Sci. 2011; 52: 2830–2836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10. Clark RA, Demer JL. Differential lateral rectus compartmental contraction during ocular counter-rolling. Invest Ophthalmol Vis Sci. 2012; 53: 2887–2896. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Demer JL, Clark RA. Magnetic resonance imaging of differential compartmental function of horizontal rectus extraocular muscles during conjugate and converged ocular adduction. J Neurophysiol. 2014; 112: 845–855. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12. Demer JL. Compartmentalization of extraocular muscle function [published online ahead of print October 24, 2014]. Eye (Lond). doi:10.1038/eye.2014.246. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13. Demer JL, Oh SY, Poukens V. Evidence for active control of rectus extraocular muscle pulleys. Invest Ophthalmol Vis Sci. 2000; 41: 1280–1290. [PubMed] [Google Scholar]
- 14. Demer JL. The orbital pulley system: a revolution in concepts of orbital anatomy. Ann N Y Acad Sci. 2002; 956: 17–32. [DOI] [PubMed] [Google Scholar]
- 15. Demer JL. Pivotal role of orbital connective tissues in binocular alignment and strabismus. The Friedenwald lecture. Invest Ophthalmol Vis Sci. 2004; 45: 729–738. [DOI] [PubMed] [Google Scholar]
- 16. Demer JL. Current concepts of mechanical and neural factors in ocular motility. Curr Opin Neurol. 2006; 19: 4–13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Demer JL. Mechanics of the orbita. Dev Ophthalmol. 2007; 40: 132–157. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18. Shin A, Yoo L, Chaudhuri Z, Demer JL. Independent passive mechanical behavior of bovine extraocular muscle compartments. Invest Ophthalmol Vis Sci. 2012; 53: 8414–8423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19. Szent-Gyorgyi A. Chemistry of Muscular Contraction. 2nd ed. New York: Academic Press; 1951. [Google Scholar]
- 20. Porter JD, Karathanasis P. The development of extraocular muscle calcium homeostasis parallels visuomotor system maturation. Biochem Biophys Res Commun. 1999; 257: 678–683. [DOI] [PubMed] [Google Scholar]
- 21. Oh SY, Poukens V, Demer JL. Quantitative analysis of rectus extraocular muscle layers in monkey and humans. Invest Ophthalmol Vis Sci. 2001; 42: 10–16. [PubMed] [Google Scholar]
- 22. Berchtold MW, Brinkmeier H, Müntener M. Calcium ion in skeletal muscle: its crucial role for muscle function, plasticity, and disease. Physiol Rev. 2000; 1215–1265. [DOI] [PubMed] [Google Scholar]
- 23. Feng C-Y, Hennig GW, Corrigan RD, Smith TK, von Bartheld CS. Analysis of spontaneous and nerve-evoked calcium transients in intact extraocular muscles in vitro. Exp Eye Res. 2012; 100: 73–85. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24. Saladin KS. Anatomy and Physiology. 6th ed. New York: McGraw-Hill; 2010. [Google Scholar]
- 25. Demer JL, Clark RA, da Silva Costa RM, Kung J, Yoo L. Expanding repertoire in the oculomotor periphery: selective compartmental function in rectus extraocular muscles. Ann N Y Acad Sci. 2011; 1233: 8–16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26. Clark RA, Demer JL. Functional morphometry of horizontal rectus extraocular muscles during horizontal ocular duction. Invest Ophthalmol Vis Sci. 2012; 53: 7375–7379. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27. Collins CC. The human oculomotor control system. In: Lennerstrand G, Bach-y-Rita P. eds Basic Mechanisms of Ocular Motility and Their Clinical Implications. New York: Pergamon; ; 1975: 145–180. [Google Scholar]
- 28. Lim KH, Poukens V, Demer JL. Fascicular specialization in human and monkey rectus muscles: evidence for anatomic independence of global and orbital layers. Invest Ophthalmol Vis Sci. 2007; 48: 3089–3097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29. Demer JL, Poukens V, Ying H, Shan X, Tian J, Zee DS. Effects of intracranial trochlear neurectomy on the structure of the primate superior oblique muscle. Invest Ophthalmol Vis Sci. 2010; 51: 3485–3493. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30. Mayr R, Gottschall J, Gruber H, Neuhuber W. Internal structure of cat extraocular muscle. Anat Embryol. 1975; 148: 25–34. [DOI] [PubMed] [Google Scholar]
- 31. McLoon LK, Rios L, Wirtschafter JD. Complex three-dimensional patterns of myosin isoform expression: differences between and within specific extraocular muscles. J Muscle Res Cell Motil. 1999; 20: 771–783. [DOI] [PubMed] [Google Scholar]
- 32. Shall MS, Dimitrova DM, Goldberg SJ. Extraocular motor unit and whole-muscle contractile properties in the squirrel monkey. Exp Brain Res. 2003; 151: 338–345. [DOI] [PubMed] [Google Scholar]
- 33. Clark RA, Demer JL. Lateral rectus superior compartment palsy. Am J Ophthalmol. 2014; 157: 479–487. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34. Le A, Poukens V, Demer JL. Compartmentalized innervation of the superior oblique (SO) muscle: double nerves for double duty? J AAPOS. 2014; 18: e18–e19. [Google Scholar]