

Abstract
Mycoplasma (MP), such as the species M. fermentans, possess remarkable immunoregulatory properties and can potentially establish chronic latent infections with little signs of disease. Atmospheric particulate matter (PM) is a complex and diverse component of air pollution associated with adverse health effects. We hypothesized that MP modulate the cellular responses induced by chemical stresses such as residual oil fly ash (ROFA), a type of PM rich in transition metals. We assessed the release of interleukin-6 (IL-6), a prototypic immune-modulating cytokine, in response to PM from different sources in human lung fibroblasts (HLF) deliberately infected with M. fermentans. We found that M. fermentans and ROFA together synergistically stimulated production of IL-6 compared to either stimuli alone. Compared to several other PM, ROFA appeared most able to potentiate IL-6 release. The potentiating effect of live MP infection could be mimicked by M. fermentans-derived macrophage-activating lipopeptide-2 (MALP-2), a known Toll-like receptor-2 agonist. The aqueous fraction of ROFA also contained potent IL-6 inducing activity in concert with MALP-2, and exposure to several defined metal salts indicated that Ni and, to a lesser extent V, (but not Cu) could synergistically act with MALP-2 to induce IL-6. These data indicate that microorganisms like MP can interact with environmental stimuli such as PM-derived metals to synergistically activate signaling pathways that control lung cell cytokine production and, thus, can potentially modulate adverse health effects of PM exposure.
Keywords: residual oil fly ash, mycoplasma, interleukin-6, particulate matter, MALP-2, human lung fibroblasts, nickel, transition metals, innate immunity, cytokines
Atmospheric particulate matter smaller than 10 μm (PM10) is listed as one of six “criteria pollutants” for which the EPA has set air quality standards. Epidemiological studies show that PM is associated with a variety of adverse health effects (Dockery et al., 1993; Pope and Kanner, 1993; Samet et al., 2000; Schwartz et al., 1993). PM arise through a variety human and natural activities and thus give rise to a myriad of PM types of diverse chemical composition and physical properties. While the biological effects of PM differ widely according to their size, composition, and quantity, certain entities including metals (Carter et al., 1997; Dye et al., 1999), organic aromatic hydrocarbons (Hiura et al., 1999), and microbial-derived biomolecules (Becker et al., 2002) have all been specifically implicated in the toxic effects. Numerous in vivo and in vitro studies reveal that PM exposure results in activation of inflammatory responses accompanied by elaboration of immune-modulating cytokines (Bayram et al., 1998; Becker et al., 1996; van Eeden et al., 2001). Residual oil fly ash (ROFA), a combustion product of fuel oil rich in transition metals, possesses potent biological effects (Ghio et al., 2002) and has been useful as a model particulate to study how metals mediate the biologic effects of air pollution particles.
Mycoplasmas (class Mollicutes) are a class of cell wall-free bacteria that represent the simplest self-replicating microorganisms known (Baseman and Tully, 1997). Due to their extremely small genome (0.58–2.20 Mb) and their limited metabolic options for replication and survival, these fastidious microorganisms have adopted a strict parasitic lifestyle in intimate relationship with a variety of animal and human hosts. With the exception of M. pneumoniae (Chanock et al., 1962), these microorganisms are not usually considered severely pathogenic; however, they may modulate host defense mechanisms (Rottem, 2002; Ruuth and Praz, 1989). Mycoplasma spp. are commonly found on various mucosal surfaces of healthy individuals, however, systemic dissemination and opportunistic growth of these microorganisms has been detected in patients with a variety of chronic inflammatory conditions including asthma (Cassell, 1998; Kraft et al., 1998). M. fermentans serves as a good example of these “stealth” pathogens (Ainsworth et al., 2000; Johnson et al., 2000; Saillard et al., 1990; Shibata et al., 1999; Vojdani et al., 1998).
We have previously observed that several early passage fibroblast cell lines derived from human lung were infected with M. fermentans (Fabisiak et al., 1993). Infection induced production of immune-modulating cytokines such as interleukin-6 (IL-6) and strongly potentiated the ability of known inducers such as TNF-β. Since exposure to PM, in general, and ROFA, in particular, can serve as potent stimuli for cytokine-dependent inflammatory responses in vitro and in vivo, we sought to test the hypothesis that M. fermentans exacerbates release of immune-modulatory factors induced by these chemical agents. We used our model of human lung fibroblasts (HLF) deliberately infected with M. fermentans to examine the ability of various components of PM to induce the induction of a prototypic immunomodulatory marker cytokine, IL-6. M. fermentans and ROFA synergistically interacted to increase the production of IL-6. These effects were mimicked with concurrent exposure to the Toll-like receptor-2 (TLR-2) agonist, M. fermentans-derived macrophage-activating lipopeptide-2 (MALP-2), and nickel. Thus, exposure to microbial-derived agents can strongly determine the cellular response to specific chemical stresses and warrants that the microbial ecology be taken in to consideration in the risks and mechanisms of toxicity posed by atmospheric pollutants such as particulate-derived metals.
MATERIALS AND METHODS
Materials
Cell culture medium, fetal bovine serum (FBS), and TRIzol™ were from Invitrogen (Gaithersburg, MD). Tissue culture plastic ware was from Falcon (Becton-Dickinson, Franklin Lakes, NJ). SP-4 mycoplasma growth media (Tully et al., 1977) was formulated with mycoplasma broth base, peptone, tryptone, Yeastolate, yeast extract (all from Becton-Dickinson Microbiological Systems, Sparks, MD), phenol red (Invitrogen), CMRL-1066 media, and Penicillin G (Sigma-Aldrich Chemical, St. Louis, MO). Calf thymus DNA, Hoechst 33258, and various metal salts were from Sigma (St. Louis, MO). Low endotoxin bovine-serum albumin (BSA) was from Intergen (Purchase, NY) and M. fermentans-derived macrophage-activating lipopeptide-2 (MALP-2) was from Alexis Biochemicals (San Diego, CA). Bradford protein assay reagent was from Bio-Rad (Hercules, CA). Murine leukemia reverse transcriptase, RNAse inhibitor, deoxynucleotides, oligo-dT, Taq DNA polymerase were from Promega (Madison, WI).
Preparations of PM were provided by Dr. Andrew Ghio, U.S.EPA. ROFA was collected by Southern Research Institute (Birmingham, AL) downstream from a cyclone scrubber at a power plant in FL burning a low sulfur #6 oil (Hatch et al., 1985). Other PM types included urban dust collected from Dusseldorf Germany (Dussel), volcanic ash from Mt. St. Helens (MSH), and an aqueous extract of PM collected from Provo Valley near Salt Lake City, Utah (SLC). The chemical and physical characteristics of these particles have been described in detail. (Becker et al., 1996; Frampton et al., 1999; Hatch et al., 1985; Prahalad et al., 1999). The ROFA particles were devoid of endotoxin activity as measured by limulus amoebocyte assay.
Cell culture
Human lung fibroblasts (HLF) were isolated as outgrowths from explanted surplus transbronchial biopsy tissues obtained during routine follow-up bronchoscopy of lung transplant recipients as previously described (Fabisiak et al., 1993) in accordance with a protocol approved by University of Pittsburgh Institutional Review Board. The individual cell lines used here were recovered from frozen stocks prepared at passage three and used for experiments over no more than eight additional subcultures. Greater than 95% purity of fibroblasts was determined by positive immunohistochemical staining for vimentin and negative staining for cytokeratin A3 or Factor VIII. Cells were maintained in Minimal Essential Medium (MEM)supplemented with FBS (10%, final concentration), glutamine (2 mM), penicillin (100 U/ml), streptomycin (100 μg/ml), and Fungizone (1.25 μg/ml) in a humidified incubator at 37°C with 5% CO2/95% air. All cultures were negative for mycoplasma as determined by fluorescent microscopy using Hoechst 33258 dye (Chen, 1977) prior to the deliberate introduction of M. fermentans.
Isolation and culture of M. fermentans
M. fermentans was isolated from previously described de novo infected HLF cell lines (Fabisiak et al., 1993) by inoculating 50 ml of SP-4 media (Tully et al., 1977) with 10 ml of spent tissue culture medium obtained from MP-infected HLF. Cultures were incubated in airtight flasks at 37°C until a red to yellow color change was observed indicative of microbial growth. Aliquots of MP cultures were cryo-preserved by addition of 0.8 ml of culture to 0.2 ml glycerol and freezing at −80°C. The total amount of MP was quantified by fluorometric determination of DNA content using a modification of Hoechst 33258 assay (Cesarone et al., 1979). The amount of viable organisms recovered from the frozen stocks was determined for each infection by determining the number of color changing units (ccu) measured with a limiting dilution assay (Rodwell and Whitcomb, 1983). The isolated strain of MP showed classical “fried-egg” morphology typical for the genus when grown on solid media and PCR positivity using M. fermentans sequence-specific primers (data not shown). In addition, the M. fermentans species was verified using a species-specific monoclonal antibody (personal communication, Dr. Shyh-Ching Lo, Armed Forces Institute of Pathology.). All experiments presented here utilized a single strain of M. fermentans derived from one infected cell line.
In vitro infection with M. fermentans
Uninfected HLF were seeded into T-75 flasks (6 × 105 cells/flask) and one P60 dish (2 × 105 cells/dish) and incubated for 24 h. At the time of infection, MP were rapidly thawed, centrifuged 12,000 × g for 15 min and washed twice with 0.25 M NaCl. Final pellet was resuspended in complete tissue culture, and each T-75 flask and P60 dish received 450 ng or 150 ng of mycoplasma DNA, respectively. Uninfected control cells were set up under identical conditions but did not receive mycoplasma and were maintained in a separate incubator. At 4 days post-infection cells were trypsinized, counted, and seeded into appropriate plates for experiments. Mycoplasma infection was verified by staining the P60 dish with Hoechst 33258 dye. Based on organism load expressed as ccu/ml and DNA content these conditions represent the introduction of ~100 viable organisms and ~300 total organisms per cell.
Cell viability
Cell viability was measured by quantifying the reduction of the fluorogenic dye, Alamar Blue (Biosource, Camarillo, CA). Briefly, MP-infected or uninfected cells were seeded into 24-well plates (4 × 104/ml) and allowed to attach for 24 h. Cells were then exposed to various concentrations of ROFA for 24 h. Media was then removed, cells washed once with PBS, and media replaced with 0.9 ml serum-free MEM. Alamar Blue (0.1 ml of a 10% solution made in serum-free MEM) was then added to each well, and cells returned to the incubator for 3 h. Fluorescence in each well was measured using a Cytofluor 2300 fluorescence plate reader using excitation 530 ± 25 nm and emission 590 ± 25 nm.
In vitro exposures and IL-6 measurement
M. fermentans-infected cells and uninfected cells were seeded into either 6-well plates (3–4.5 × 105 cells/3 ml/well) or 24-well plates (0.6–1 × 105 cells/1 ml/well) and cultured for 48 h. Media was removed and replaced with the same volume of serum-free media containing 0.1% BSA with or without various stimuli. Conditioned media was collected at indicated time points and stored at −80°C until further analysis. IL-6 content of conditioned media was measured using a specific enzyme-linked immunosorbent assay (ELISA) kit (R&D Systems, Minneapolis, MN) according to the manufacturer’s instructions. For experiments in 6-well plates IL-6 content was normalized to the producing cell number measured by nuclei counts obtained after lysis in 0.1% citric acid/0.1% crystal violet (Fabisiak et al., 1993). For experiments in 24-well plates IL-6 was normalized to the DNA content of the monolayer measured using Hoechst 33258 fluorescence (Cesarone et al., 1979).
Measurement of cytokine mRNA
Total cellular RNA was isolated using TRIzol™ reagent (Invitrogen, Gaithersburg, MD) according to the manufacturer’s instructions. cDNAs were generated from 0.5 μg total RNA by reverse transcription in a 30 μl reaction mixture containing Ambion 103 first strand buffer, 10 U RNase inhibitor, 0.33 mM each dNTP, 1.7 mM oligo dT, and 100 U M-MLV reverse transcriptase. Reactions were incubated at 44°C for 60 min in a MJ Research PTC-100 thermocycler. cDNA was stored at −20°C until further analysis. Specific primer pairs for IL-6 (forward 5′-GCCCAGCTATGAACTCCTTCTC; reverse 5′-GACTTGTCATGTCCTGCAGCC), IL-8 (forward 5′-ATGACTTCCAAGCTGGCCGTGGCT; reverse 5′-TCTCAGCCCTCTTCAAAAACTTCTC) and β-actin (forward 5′-GGGACCTGACCGACTACCTC; reverse 5′-GGGCGATGATCTTGATCTTC) were used to amplify the specific cDNAs. Specific cDNAs were amplified using 5 μl aliquots cDNA mixed with 1.25 μl of IL-6, IL-8, or β-actin forward and reverse primers, 2.5 U of Taq DNA polymerase, and 0.125 mM dNTP in Ambion complete PCR buffer in a final volume of 50 μl. PCR reactions were carried out for 20 s at 95°C, 30 s at 55°C, and 40 s at 72°C for 19 cycles for β-actin or 24 cycles IL-6 and IL-8. The number of cycles was demonstrated to be within the linear amplification range for each product. PCR products were either detected on 2% agarose gels stained with ethidium bromide or quantified in real-time fashion during the PCR amplification using the double-strand DNA fluorescent dye PicoGreen at 430 nm emission and 525 nm excitation. IL-8 and IL-6 mRNA expression were normalized to the housekeeping gene β-actin by determining the ratio of the IL-8 or IL-6 fluorescent signals to that for β-actin.
Statistical analysis
Data presented are expressed as mean ± SEM collected from at least three or four individual experiments for studies employing pooled HLF or cells derived from individual donors, respectively. Comparisons were made using a one-way ANOVA followed by appropriate group comparisons such as Dunnett’s multiple comparison to control or Bonferonni’s correction for multiple t-tests. Significance of response to a range of concentrations to single agents was determined by a test for linear trend across the applied concentrations. To control for variability between multiple individually derived cell lines repeated measures ANOVA and post hoc tests were applied with pairing of data collected within a specific cell line. When variability between cell lines precluded the assumption of a normal distribution, nonparametric Wilcoxon signed rank tests were used to compare groups. To compare observed dose-response relationships with combined stimuli to a theoretical additive model, concentrations were transformed to log scale, and a predicted additive response was derived as the algebraic sum to similar concentrations of each agent alone. Log–dose response curves were subjected to linear regression and the slopes compared by analysis of covariance. Statistical analyses were performed using GraphPad PRISM™, version 3.0 (GraphPad™ Software, San Diego, CA), and differences were considered significant at the p < 0.05.
RESULTS
M. fermentans and ROFA Synergistically Stimulate IL-6 Release from HLF
To explore possible interactions between ROFA and Mycoplasma fermentans (MP), we first exposed MP-infected and uninfected HLF to various concentrations of ROFA for 24 h. Production of the immune-modulating cytokine IL-6 during this period was then measured (Fig. 1). In the absence of ROFA, IL-6 release in uninfected cells was 54 ± 11 pg/105 cells and about 5-fold greater in cells infected with MP (445 ±127 pg/105 cells). Exposure of uninfected cells to ROFA alone produced a dose-dependent increase in IL-6 release that, at the highest dose tested (603 ± 141 pg/105 cells at 50 μg ROFA/ml), approximated that seen in the presence of MP alone. When MP-infected cells were exposed to ROFA, however, the response was dramatically enhanced even at the lowest concentration of ROFA used (2 μg/ml), with IL-6 production ranging from 21 to 50 times greater than the basal production seen in uninfected cells. In order to more clearly characterize the synergistic nature of this interaction, we compared the observed dose-response curve to ROFA in MP-infected cells to that predicted by an additive model for these two stimuli (IL-6 release from MP alone + IL-6 released by each concentration of ROFA applied to uninfected cells). Figure 1B shows the observed and predicted log–dose response curves. Linear regression of each line was performed and comparison revealed a highly significant difference between the slopes of the observed and predicted lines (p <0.02). Thus, the observed response to ROFA plus MP was always greater than that predicted in an additive model, and this difference grew in magnitude as the concentration of ROFA increased. This represents a synergistic interaction between these two stimuli.
FIG. 1.

M. fermentans synergistically amplifies the ability of ROFA to induce IL-6 production in human lung fibroblasts (HLF). HLF deliberately infected with M. fermentans and uninfected control cells were seeded in 6-well plates (4 × 106 cells/well) and allowed to attach for 24 h. Cells were then washed with PBS and changed to serum-free MEM containing 0.1% BSA in the presence or absence of indicated concentrations of ROFA. Conditioned media was collected 24 h after stimulation and analyzed for IL-6 by ELISA. IL-6 content was normalized to the producing cell number by nuclei counts obtained after cell lysis in citric acid/crystal violet. Data represent mean ± SEM obtained from four independent cell lines derived from different donors. Panel A shows the IL-6 response of infected and uninfected cells to various concentrations of ROFA. *Denotes statistically significant difference compared to uninfected cells exposed to the equivalent concentration of ROFA by one-way repeated measures ANOVA and Bonferonni’s multiple t-tests. Panel B show a linear regression of the log–dose response curve to ROFA in infected cells (closed symbols) compared to that predicted by adding the individual responses to ROFA alone and infection alone (open symbols). Slope of the observed dose-response was significantly greater than the predicted by analysis of covariance (p = 0.018).
We also measured the viability of HLF after exposure using the vital reduction of Alamar Blue. The LD50 was approximately 100 μg/ml and did not appear to be altered by MP-infection, nor was it influenced by the presence or absence of 10% FBS during the ROFA exposure period (data not shown). The observed LD50 was considerably higher than concentrations sufficient to produce a synergistic effect of IL-6 production (Fig. 1). Thus, the synergistic interactions between ROFA and MP on induction of IL-6 production do not occur in the context of differential sensitivities to the overall toxic effects of ROFA.
Comparison of ROFA to PM from Other Sources
We next examined whether other types of PM might similarly augment IL-6 release from MP-infected cells. We measured IL-6 release during a 24-hour exposure of MP-infected and control uninfected HLF to equivalent concentrations (20 μg/ml) of urban dust collected from Dusseldorf, Germany (Dussel); aqueous extract of PM obtained from Provo Valley near Salt Lake City, Utah (SLC); Mt. St. Helens volcanic ash (MSH); as well as ROFA. Figure 2A shows that 20 μg/ml ROFA significantly increased IL-6 release about four-fold above that seen in untreated uninfected cells. In contrast, PM derived from other sources had little effect on IL-6 release at this same concentration. The slight increase observed with SLC did not reach statistical significance. When these same exposures were carried out on MP-infected cells, it was again observed that the combination of ROFA nearly tripled the release of IL-6 relative to untreated MP-infected cells alone (Fig. 2B). In keeping with the synergistic response, it is important to note that the amount of IL-6 release seen with the combination of ROFA plus MP is nearly 20-fold and 100-fold greater than that seen in uninfected cells in the presence and absence of ROFA, respectively. Figure 2B also shows that the other PM types at this concentration lacked sufficient ability to induce IL-6 in MP-infected cells similar to uninfected cells. Thus, the observed potency of ROFA to induce IL-6 release and interact with MP appears to arise from some unique property of this PM or relative enrichment of some chemical component compared to other PM.
FIG. 2.
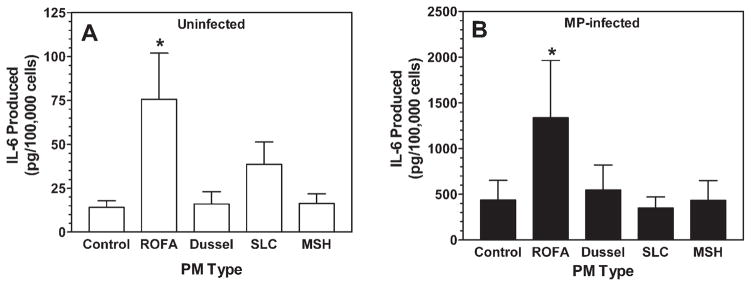
Comparison of various PM types on IL-6 production in control and MP-infected HLF. Uninfected control (panel A) and M. fermentans infected cells (panel B) were exposed to 20 μg/ml of each of the PM types in serum-free MEM for 24 h. Conditioned media were then collected and analyzed for IL-6 by ELISA. The various PM types tested included residual oil fly ash (ROFA), ambient urban dust collected from Dusseldorf, Germany (Dussel), an aqueous extract of ambient particles collected from Provo Valley near Salt Lake City, UT (SLC), and Mt. St. Helen’s volcanic ash (MSH). Data represent the means ± SEM obtained from six individual cell lines derived from different donors. *Denotes statistically significant difference compared to the respective unstimulated control for the uninfected and infected groups using a one-way ANOVA with Dunnett’s multiple comparisons to control (p < 0.05).
M. fermentans-Derived 2 kDa Lipopeptide (MALP-2) Mimics the Effect of M. fermentans to Synergistically Interact with ROFA and Stimulate IL-6 Release
We next sought to further define the signaling pathways that might be involved in IL-6 production during live infection with MP and hypothesized that activation of innate immune responses via activation of specific Toll-like receptor pathways could play a role. Activation of TLR-4 initiates the well-described induction of inflammatory cytokines by macrophages and other cells in response to lipopolysacharide (LPS) derived from gram-negative bacteria cell walls. Mycoplasma, however, lack a characteristic cell wall and hence are devoid of endotoxin. Several reports demonstrated that membrane lipoproteins derived from M. fermentans exhibit macrophage-stimulatory activities and induce the production of proinflammatory cytokines (Muhlradt et al., 1997; Rawadi and Roman-Roman, 1996). Macrophage-activating lipopeptide-2 (MALP-2), a 14 amino acid N-terminal fragment containing a unique diacylated cysteine, has been characterized and shown to induce the production of proinflammatory cytokines, as well as nitric oxide, via specific interaction with TLR-2 and TLR-6 (Nishiguchi et al., 2001; Takeuchi et al., 2001). We wondered if MALP-2 could recapitulate the effects of live infection and synergistically interact with ROFA to induce cytokine release. To test this possibility, we exposed HLFs to various concentrations of MALP-2 in the presence and absence of ROFA (40 μg/ml). Conditioned media was collected after both 24 h exposure (Fig. 3A) and 48 h exposure (Fig. 3B) and analyzed for IL-6 content. Exposure of HLF to MALP-2 alone induced a moderate concentration-dependent increase in IL-6. The effect of MALP-2 alone was first observed at 300 pg/ml and was further increased at 600 pg/ml. The amount of cytokine released relative to unstimulated cells after 48 h exposure to the highest concentration of MALP-2 alone (385 ± 277 with MALP-2 vs. 9 ±4 pg/μg DNA unstimulated) was approximately 2-fold higher than that observed after only 24 h (150 ± 105 vs. 4 ±1 pg/μg DNA), suggesting a fairly linear increase in IL-6 accumulation within the medium over this time. ROFA itself in the absence of MALP-2 had a very minimal effect on IL-6 release that reached 11 ± 4 and 45 ± 21 at the 24 (Fig. 3A) and 48 (Fig. 3B) h time points, respectively. The inclusion of MALP-2 in combination with ROFA, however, markedly stimulated the release of IL-6. The between cell line variability was sufficiently high to preclude a normal distribution of our data points. At each concentration of MALP-2, however, each of the six cell lines tested showed an enhanced response to combination of ROFA and MALP-2 (p <0.05, Wilcoxon’s signed rank test). At 24 and 48 h, the highest concentration of MALP-2 (600 pg/ml) in combination with ROFA released three- and six-fold more IL-6, respectively, compared to the same concentration of MALP-2 alone. We also compared the observed MALP-2 + ROFA response to values predicted by an additive model by summing the responses seen to ROFA alone and MALP-2 alone for each cell line (dotted lines in Figs. 3A and 3B). At both time points the response observed with both 300 and 600 pg/ml MALP-2 in combination with ROFA was significantly more than predicted from an additive model. To further demonstrate the synergistic nature of these interactions, we analyzed log–dose response relationships by linear regression using those concentrations of MALP-2 having a demonstrable effect (100–600 pg/ml) (Figs. 3A and 3B, insets). The slope of the observed response to MALP-2 plus ROFA was three-fold and seven-fold greater at the 24 and 48 h time points, respectively, compared to that predicted by an additive model; thus it is clear that the addition of ROFA dramatically synergized the effect of MALP-2 to induce IL-6.
FIG. 3.

M. fermentans -derived macrophage activating peptide (MALP-2) and ROFA synergistically stimulate IL-6 release from HLF. HLF were seeded into 24-well plates and allowed to attach overnight. Cells were then exposed to ROFA (40 μg/ml) in the presence or absence of various concentrations of MALP-2 (30–600 pg/ml) in serum-free MEM with 0.1% BSA. Control cells received MALP-2 alone in the absence of ROFA. IL-6 content in the media was measured at both 24 (panel A) and 48 h (panel B) after exposure. Cytokine release is expressed per μg cellular DNA in the monolayer. Data represent the mean ± SEM obtained from six individual cell lines derived from different donors. The large variability observed in both basal and induced responses observed in this experiment did not permit the assumption of a normally distributed population, and therefore nonparametric statistics were applied. Wilcoxon signed rank tests were used to compared the observed response to MALP-2 plus ROFA to the response observed to that same concentration of MALP-2 alone (* = p < 0.05) and the predicted additive response (dotted line) calculated as the algebraic sum of the ROFA alone and MALP-2 alone responses (# = p < 0.05). Insets show the linear regression of the mean log–dose response curves observed with effective MALP-2 concentrations (100–600 pg/ml) in combination with ROFA and those predicted by an additive model. Individual slopes were calculated using the data collected from each cell line and compared by Wilcoxon signed rank test. The slope of the observed line was significantly different from that predicted by the additive model at both time points (p = 0.0313).
The Effect of ROFA Is Mimicked by Its Water-Soluble Fraction and Specific Metal Constituents
Airborne products derived from the combustion of fossil fuels are complex mixtures containing an elemental carbon core with adsorbed organic hydrocarbons and various metal salts and oxides depending on the origin of the fuel. ROFA is extremely rich in its metal content, most notably vanadium (V), iron (Fe), and nickel (Ni), among others. Many studies have implied that ROFA’s soluble metallic components are responsible for its bioactivity. To first evaluate the effect of soluble components in ROFA, we prepared a standard solution of ROFA particles in serum-free tissue culture media as used in the experiments described above. Half of this mixture was filtered through a 0.22 μm filter to essentially obtain an aqueous extract of ROFA. We exposed HLF to either untreated ROFA (20 and 40 μg/ml) or the soluble fraction corresponding to the same amounts of whole ROFA. Table 1 shows the IL-6 response to soluble ROFA (filtrate) and total ROFA in the presence and absence of MALP-2 (600 pg/ml) after treatment for 24 and 48 h. As expected, exposure of the cells to ROFA or MALP-2 alone has very minimal effects of IL-6 release at either time point. When cells were exposed to the lowest concentration of total or soluble ROFA in combination with MALP-2 there was no enhancement at the 24 h time point, but by 48 h cells exposed to either total ROFA or soluble ROFA in the presence of MALP-2 released approximately ten-fold more IL-6 than cells exposed to any stimulus alone. With this small number of observations the response observed with 20 μg/ml ROFA filtrate plus MALP-2 failed to reach statistical significance, however; no significant difference was detected compared to the response seen with the same concentration of total ROFA plus MALP-2. When a higher concentration of ROFA (40 μg/ml) was used, significant synergy was observed between MALP-2 and total ROFA at both time points. The response seen with 40 μg/ml ROFA filtrate plus MALP-2 was about 50% and 75% of that seen with total ROFA plus MALP-2 at the 24 and 48 h time points, respectively. Thus, these results show that soluble components of ROFA can synergistically interact with MALP-2 in producing IL-6 in a manner similar to that seen with exposure of HLF to the entire particle. Although the synergy seen with ROFA filtrate is slightly weaker than ROFA as a whole, there is no statistically significant difference between intact ROFA and its soluble components.
TABLE 1.
Water-Soluble Components of ROFA Mimic the Intact Particle to Induce IL-6 in HLF in the Presence of MALP-2
| Treatment | IL-6 Produced (pg/μg DNA)
|
|||
|---|---|---|---|---|
| 24 h | 48 h | |||
| Untreated | 3 ± 0.4 | 21 ± 13 | ||
| MALP-2 alone (600 pg/ml) | 33 ± 14 | 64 ± 22 | ||
| 20 μg/ml ROFA | ||||
| Total alone | 11 ± 4 | 57 ± 25 | ||
| Total + MALP-2 | 38 ± 15 |
|
773 ± 249a |
|
| Filtrate alone | 11 ± 7b | 39 ± 29 | ||
| Filtrate + MALP-2 | 36 ± 10 | 544 ± 281 | ||
| 40 μg/ml ROFA | ||||
| Total alone | 25 ± 15 | 91 ± 39 | ||
| Total + MALP-2 | 499 ± 235a |
|
1097 ± 338a |
|
| Filtrate alone | 39 ± 30 | 42 ± 13 | ||
| Filtrate + MALP-2 | 196 ± 71 | 836 ± 185a | ||
Note. Data represent mean ±SEM obtained from four individual experiments using a single pooled HLF cell line.
Statistically significant difference (p < 0.05) compared to the predicted additive response obtained by summing the individual responses observed with MALP-2 alone and the ROFA fraction alone in each experiment by one-way ANOVA and Bonferroni’s Multiple Comparisons.
ns denotes no significant difference between total ROFA and soluble ROFA fraction in the presence of MALP-2.
Since ROFA’s soluble metallic components may be responsible for this synergy, we next sought to define which metal elements found in ROFA might alone synergize with MALP-2 to induce IL-6 release from HLFs. The ROFA preparation used in the previous experiments contains high levels of vanadium, nickel, iron, and copper. Therefore, we first treated HLFs with three concentrations of various metallic salts in the presence or absence of 600 pg/ml MALP-2 (Fig. 4). The most dramatic interaction for stimulation of IL-6 production was seen with NiSO4 and MALP-2. Exposure of HLF to 200 μM NiSO4 by itself produced about a ten-fold increase in IL-6 production and was about the same magnitude as that seen with MALP-2 alone. The combination of MALP-2 along with 200 μM Ni, however, stimulated IL-6 release to a level over 5 times that seen with either stimulus alone and over 50 times greater than that observed in unstimulated cells. Exposure of cells to lower concentrations of Ni alone (2 and 20 μM) was ineffective and did not modulate the response to MALP-2. Exposures of HLF to similar concentrations of CuSO4 had no stimulatory effect by themselves and, in fact, decreased the response to MALP-2 alone. The two vanadium compounds, vanadyl (V4+) sulfate (VOSO4) and sodium vanadate (V5+) (Na3VO4), induced small increases in cytokine production when applied at 200 μM in the absence of MALP-2 that were similar to that seen with the same concentration of Ni alone. While the addition of MALP-2 to 200 μM Na3VO4 further increased IL-6 release, the observed response appeared was much less than that observed with Ni + MALP-2. The response to 200 μM VOSO4 in combination with MALP-2 appeared only slightly greater than the additive effects of each stimulus alone and also was substantially less than that seen with nickel.
FIG. 4.

Effect of several soluble metal salts on IL-6 release in the presence and absence of MALP-2. A composite cell line representing early passage-cells pooled from six different donors was seeded into 24-well plates and allowed to attach for 48 h and then exposed to varying concentrations of the specific metal salts in the presence or absence of MALP-2 (600 pg/ml) in serum-free MEM with 0.1% BSA for 48 h. IL-6 was measured in the conditioned media by ELISA and normalized to DNA content of the monolayer at the end of the incubation. Data represent mean ± SEM of four different experiments. *Denotes statistically significant difference compared to control cells in the absence of any stimuli, and **denotes statistically significant difference compared to cells treated with MALP-2 alone by one-way ANOVA and Dunnett’s multiple comparison to control.
To explore the dose-response relationships of Ni in more detail and further demonstrate the synergistic relationship between NiSO4 and MALP-2, we treated HLFs with various concentrations of NiSO4 between 20 and 200 μM in the presence or absence of 600 pg/ml MALP-2 (Fig. 5). The effects of nickel alone were extremely small, with IL-6 release remaining below 100 pg/μg DNA at the highest concentration for both the 24 h (Fig. 5A) and 48 h (Fig. 5B) time points. NiSO4, however, dramatically increased IL-6 release in a dose-dependent manner in the presence of MALP-2 to a level that was ten times that seen with MALP-2 or the highest concentration of Ni alone. At the 48 h time point in the presence of MALP-2, even the lowest concentration of Ni provided enhancement over 20 μM Ni alone (101 ± 19 vs. 6.1 ±0.7 pg IL-6/μg DNA) and was nearly twice that predicted from an additive model (62.3 ± 11.7 pg IL-6/μg DNA). The synergistic interaction between Ni and MALP-2 was apparent when the slope of the observed dose-response to Ni plus MALP-2 was compared to that predicted by adding the observed responses to each agent alone, where the slope of the observed response was ten-fold greater than that predicted by an additive model. Thus, soluble nickel ions may be one of the primary components in ROFA capable of synergistically interacting with MALP-2 to stimulate IL-6 production from HLF.
FIG. 5.

NiSO4 and MALP-2 synergistically stimulate IL-6 release from HLF. HLF from the pooled composite cell line were seeded into 24-well plates and allowed to attach overnight. Cells were then exposed to MALP-2 (600 pg/ml) in the presence or absence of various concentrations of NiSO4 (20–200 μM) in serum-free MEM with 0.1% BSA (closed symbols). Control cells received NiSO4 alone in the absence of MALP-2 (open symbols). IL-6 content in the media was measured at both 24 (panel A) and 48 h (panel B) after exposure. Cytokine release is expressed per μg cellular DNA in the monolayer. Data represent the mean ± SEM obtained from 6 observations. *Denotes statistically significant difference between with and without MALP-2 for any given concentration of Ni by one-way ANOVA and Bonferroni’s multiples comparisons (p < 0.05). A theoretical additive response was calculated as the algebraic sum of the responses observed at each Ni concentration in the absence of MALP-2 and that observed with MALP-2 alone (dotted line). #Denotes significant difference between the response observed with each concentration of Ni plus MALP-2 compared to that predicted by the additive model (p <0.05). Insets show the linear regression of the mean log–dose response curves observed with effective NiSO4 concentrations (50–200 μM) in combination with MALP-2 and those predicted by an additive model. Slope of the observed log dose-response was significantly greater than that predicted by the additive model by analysis of covariance (p < 0.0001).
The effects of ROFA and Ni were further investigated at the level of cytokine mRNA and compared for their ability to modulate gene expression of IL-6 and another cytokine, interleukin-8 (IL-8). IL-8 is a potent inflammatory cytokine with neutrophil chemotactic activity whose regulation shares many similarities to that of IL-6. The steady-state levels of both IL-6 and IL-8 mRNA in HLF were measured after 24 h stimulation with Ni (200 μM) or ROFA (40 μg/ml) in the presence or absence of MALP-2 using RT-PCR, and β-actin gene expression was assessed as a control. Figure 6A shows a representative experiment and the specific ethidium bromide-stained PCR products obtained after agarose gel electrophoresis. Note that both IL-6 and IL-8 transcripts were barely detectable in unstimulated cells. Increased expression of IL-6 and IL-8 was observed after 24 h treatment with either NiSO4 or ROFA alone and was similar in magnitude to that seen after MALP-2 alone. The combinations of MALP-2/ROFA or MALP-2/NiSO4, however, produced a dramatic accumulation of mRNA for both cytokines that was much greater than that seen with any single agent alone. The level of each cytokine mRNA species was measured using quantitative real-time RT-PCR, normalized to the amount of β-actin transcript, and then compared between each treatment (Figs. 6B and 6C). The steady-state level of IL-6 mRNA was induced about ten-fold above unstimulated control when cells were exposed to any of the stimuli as single agents; in contrast, the abundance of IL-6 transcripts were approximately 50-fold greater with the combinations of MALP-2 + ROFA and MALP-2 + NiSO4. A similar pattern of induction was also seen for IL-8, although the difference between MALP-2 + NiSO4 and NiSO4 alone just failed to reach statistical significance, in part, because of the unequal variances between the combination and single-stimuli exposure groups.
FIG. 6.

Effects of MALP-2 plus ROFA or NiSO4 on steady-state level of IL-6 and IL-8 mRNA. Cells were seeded into 6-well plates, incubated overnight, and then treated with MALP-2 (600 pg/ml), NiSO4 (200 μM), and/or ROFA (40 μg/ml) in serum-free medium containing 0.1% BSA. Twenty-four h after treatment total RNA was harvested and IL-6, IL-8, and β-actin mRNA levels were measured by RT-PCR. Panel A shows the representative PCR products observed from a typical experiment visualized by ethidium bromide staining, a and b denote duplicate samples. Panels B and C show the results of real-time quantitative PCR of cytokine mRNA normalized relative to β-actin. Data represent mean ± SEM from 6 RNA samples and are expressed as fold-induction relative to untreated control cells. *Denotes statistically significant difference (p <0.05) relative to the same condition in the absence of MALP-2 by one-way ANOVA and Bonferroni’s multiple comparisons.
DISCUSSION
Our study is the first to observe a potent and significant interaction between a particular PM, ROFA, and exposure to the microorganism, M. fermentans. Using IL-6, a prototypic immune-modulating cytokine, as a biomarker for cellular activation of inflammatory responses, we demonstrate that levels of chemical stress and microbial stimulation that produce minimal response alone produce profound cell-activating effects when applied together. These interactions are clearly synergistic since, (1) the observed response to simultaneous chemical and microbial exposure is greater than that predicted by the algebraic sum of each stimulus alone, and (2) the shape and slope of the dose-response curves to each agent can be reciprocally modulated by minimally-effective concentrations of the other.
Dose-response relationships determined in specific pathogen-free animals and sterile cell culture models are often used to set risk assessment guidelines for various environmental agents. This rarely, however, mimics relevant “real-life” exposures, where individuals may be exposed in the presence of microbial infection/colonization. Such infection can range from clinically evident disease (pneumonia, tracheobronchitis), to subclinical latent chronic infection, to colonization with various “harmless” commensals. Moreover, ambient PM itself is a complex mixture that contains particulates from many sources, including those of biological origin. In fact, bacterial endotoxin and other microbial products have been implicated in some of the biological activity of ambient PMs (Becker et al., 2002). Thus, the synergistic interactions between different components of the PM mixture and concurrent presence of microorganisms need to be considered when evaluating their overall toxicity.
M. fermentans, serves as an example of a mycoplasma with potential to establish chronic subclinical infection. Originally isolated from the genitourinary (Ruiter and Wentholt, 1950), M. fermentans has also been documented in joint fluid of patients with rheumatoid arthritis but not those with other arthrides (Horowitz et al., 2000; Johnson et al., 2000), leukemic bone marrow (Murphy et al., 1970), and a disproportionately high percentage of blood samples from those with chronic fatigue syndrome (Vojdani et al., 1998). Recently, Mycoplasma spp. were detected in the airways of humans in absence of symptoms of acute infection, and the incidence was greater in asthmatics (Kraft et al., 1998). Using sensitive PCR-based detection methods, high incidences of M. fermentans positivity have been noted in saliva (Shibata et al., 1999), blood (Ainsworth et al., 2000), and urine (Kovacic et al., 1996) from apparently normal healthy subjects. Data regarding the presence of M. fermentans within the human lung or its ability to establish chronic “symptomless” pulmonary infection, however, are severely limited.
It remains unresolved as to what particular components of PM contribute to its adverse health effects. ROFA is frequently used as a surrogate to study the adverse effects of PM, particularly in the context of particulate-derived metal (Ghio et al., 2002); however, it represents a very small component of total air PM and does not accurately reflect the complex composition of ambient particulate mixtures. Several studies have pointed to the importance of the “fine” (<2.5 μm) and “ultrafine” (<0.1 μm) fractions in mediating ambient PM toxicity (Laden et al., 2000; Peters et al., 1997). Although these size fractions represent only <50% and <10%, respectively, of the total mass fraction of PM10, these fractions are enriched in their metal content. Multiple sources of PM-derived metals exist in addition to ROFA. Whether ambient PMs are capable of interacting with microbial stimuli in a manner analogous to ROFA will undoubtedly depend on their sources, chemical composition, size distribution, and physical properties.
The ability of ROFA to produce cell and tissue injury and stimulate inflammatory cytokines has been linked to its high metal content (Carter et al., 1997; Dreher et al., 1995; Dye et al., 1999; Pritchard et al., 1996; Samet et al., 1997). In much the same way, our data support a role for particulate-derived metal in mediating the synergistic effects with MP. Vanadium and iron have been implicated as the primary transition metals in ROFA capable of inducing oxidative stress and activating inflammatory responses such as cytokine release. In contrast, our data point to an important role for Ni in mediating the effects observed here. The preparation of ROFA used here contains 37.5 mg Ni/gm, which if completely soluble would produce from 6 to 25 μM Ni with the ROFA concentrations used in Figure 1 (10–40 μg/ml). It should be pointed out, however, that the threshold for the effect of NiSO4 was between 20 and 50 μM; therefore, it is likely that other metals or alternate forms of Ni also participate in the overall effects of ROFA. Ni is most often considered as an occupational hazard in exposed workers (Morgan and Usher,1994), although considerable exposure does occur in the general population, often in the form of ultrafine metal-rich combustion-derived PM. Nearly 1 million and 200 million people living in the vicinity of Ni-emitting sources are exposed to median concentrations of 200 and 50 ng Ni/m3 respectively (Leikauf, 2002). Assuming an ambient level of 100 ng/m3, 50% deposition, negligible elimination, and normal respiration, we estimate a daily dose of 864 ng Ni that if uniformly distributed within the noncellular volume of the lung (22 ml) (Weibel, 1985) would produce ≈1 μM Ni. While this value is below that necessary to produce synergy, a small change in breathing parameters, potential for uneven distribution, and possible accumulation over time could produce Ni levels closer to those used in our experiments. Using data from the Six Cities studies, Laden et al. (2000) demonstrated that Ni was positively associated with daily deaths. In animal studies, Ni synergistically interacts with V to initiate untoward cardio-vascular effects following inhalational exposure (Campen et al., 2001).
Our data do not speak directly to the cellular and molecular mechanisms that account for the synergistic interactions between ROFA and MP. Since ROFA contains a variety of transition metals (Fe, V, Ni, among others) it is possible that enhanced formation of reactive oxygen species (ROS) during metal and microbial exposure can subsequently modulate signal transduction pathways culminating in IL-6 release. The SLC PM, however, was inhibitory towards MALP-2-induced IL-6 despite the fact that this PM is particularly rich in redox-active Cu (Kennedy et al., 1998). Although Ni leads to formation of ROS (Andrew et al., 2001; Huang et al., 1993; Misra et al., 1990) it is not considered as redox-active as other transition metals. It is possible, however, that Ni coordinated to specific ligands enhances its redox activity (Misra et al., 1993; Shi et al., 1992) and/or imparts an intracellular regio-specificity that is critical for these effects.
Numerous transcription factors such as NF-κB and AP-1 play well-established roles in the regulation of cytokine gene expression following diverse microbial and chemical stressors. Activation of these pathways by ROFA (Quay et al., 1998; Samet et al., 2002) and MALP-2 (Rawadi et al., 1999) can occur through activation of protein kinase-dependent and oxidant-sensitive mechanisms. Other mechanisms of Ni-dependent regulation gene expression include stabilization of hypoxia-inducible factor (HIF-1α) (Andrew et al., 2001), as well as other novel, as yet defined, transcription factors (Barchowsky et al., 2002). Other possibilities for mechanisms of regulation-include metal-dependent inactivation of specific signal-transducing protein tyrosine phosphatases (Samet et al., 1997, 1998) and modulation of mRNA stability (Winzen et al., 1999). Thus, it is possible that synergy arises via amplified activation of one or more of these specific transcription factors or signaling pathways or, alternatively, that each stimulus produces a unique profile of signaling events that converge to govern gene expression in an interactive way.
HLF are useful as in vitro model for these studies since they (1) support live MP infection, (2) represent a relatively normal untransformed human lung cell type without artifacts of prolonged tissue culture, and (3) play an active role in the cytokine and fibrotic response following tissue injury. Other cells such as macrophages and epithelial cells, however, may arguably be more relevant since they are amongst the first to encounter PM and infectious stimuli. Since regulation of cytokine production in diverse cell types likely follows common mechanisms, we anticipate that microbial stimuli and particulate-derived metals will similarly interact to activate other cell types within the lung.
In summary, our studies demonstrate the potential for profound synergistic interactions between microbial products and PM in the ability of HLF to produce immune-modulating/inflammatory cytokines. These effects occur in absence of cytotoxicity and with concentrations of stimuli that produce minimal effects by themselves. Since it appears that mycoplasma are signaling through TLR-dependent mechanisms, the phenomenon can likely be extended to a range of diverse microorganisms. Our studies provide an experimental model to further examine the cellular and molecular mechanisms by which these microbial and chemical stimuli interact to modulate the expression of immune-modulating cytokines and other gene products important in the response to various environmental stresses.
Acknowledgments
The authors are deeply indebted to Dr. Andrew Ghio (U.S. Environmental Protection Agency) for providing the various PM types and information regarding their chemical and physical characteristics. We would also like to thank Dr. Shyh-Ching Lo, Armed Forces Institute of Pathology, for his help in characterizing the M. fermentans isolates, as well as Robyn Thomson and Linda Klei for invaluable technical assistance. This work was supported by the U.S. EPA (STAR Grant R827151) and NIH (ES 10638).
References
- Ainsworth JG, Hourshid S, Webster AD, Gilroy CB, Taylor-Robinson D. Detection of Mycoplasma fermentans in healthy students and patients with congenital immunodeficiency. J Infect. 2000;40:138–140. doi: 10.1016/s0163-4453(00)80005-5. [DOI] [PubMed] [Google Scholar]
- Andrew AS, Klei LR, Barchowsky A. Nickel requires hypoxia-inducible factor-1α, not redox signaling, to induce plasminogen activator inhibitor-1. Am J Physiol Lung Cell Mol Physiol. 2001;281:L607–L615. doi: 10.1152/ajplung.2001.281.3.L607. [DOI] [PubMed] [Google Scholar]
- Barchowsky A, Soucy NV, O’Hara KA, Hwa J, Noreault TL, Andrew AS. A novel pathway for nickel-induced interleukin-8 expression. J Biol Chem. 2002;2002:24225–24231. doi: 10.1074/jbc.M202941200. [DOI] [PubMed] [Google Scholar]
- Baseman JB, Tully JG. Mycoplasmas: Sophisticated, reemerging, and burdened by their notoriety. Emerg Infect Dis. 1997;3:21–32. doi: 10.3201/eid0301.970103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bayram H, Devalia JL, Sapsford RJ, Ohtoshi T, Miyabara Y, Sagai M, Davis RJ. The effect of diesel exhaust particles of cell function and release of inflammatory mediators from human bronchial epithelial cells in vitro. Am J Respir Cell Mol Biol. 1998;18:441–448. doi: 10.1165/ajrcmb.18.3.2882. [DOI] [PubMed] [Google Scholar]
- Becker S, Fenton MJ, Soukup JM. Involvement of microbial components and toll-like receptors 2 and 4 in cytokine responses to air pollution particles. Am J Respir Cell Mol Biol. 2002;27:611–618. doi: 10.1165/rcmb.4868. [DOI] [PubMed] [Google Scholar]
- Becker S, Soukoup JM, Gilmour I, Devlin RB. Stimulation of human and rat alveolar macrophages by urban air particulates: Effects on oxidant radical generation and cytokine production. Toxicol Appl Pharmacol. 1996;141:637–648. doi: 10.1006/taap.1996.0330. [DOI] [PubMed] [Google Scholar]
- Campen MJ, Nolan JP, Schladweiler MC, Kodavanti UP, Evansky PA, Costa DL, Watkinson WP. Cardiovascular and thermoregulatory effects of inhaled PM-associated transition metals: A potential interaction between PM-associated transition nickel and vanadium sulfate. Toxicol Sci. 2001;64:243–252. doi: 10.1093/toxsci/64.2.243. [DOI] [PubMed] [Google Scholar]
- Carter JD, Ghio AJ, Samet JM, Devlin RB. Cytokine production by human airway epithelial cells after exposure to an air pollution particle is metal-dependent. Toxicol Appl Pharmacol. 1997;146:180–188. doi: 10.1006/taap.1997.8254. [DOI] [PubMed] [Google Scholar]
- Cassell GH. Infectious causes of chronic inflammatory diseases and cancer. Emerg Infect Dis. 1998;4:475–487. doi: 10.3201/eid0403.980339. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cesarone CF, Bolognesi L, Santi L. Improved microfluorometric DNA determination in biological material using 33258 Hoechst. Anal Biochem. 1979;100:188–197. doi: 10.1016/0003-2697(79)90131-3. [DOI] [PubMed] [Google Scholar]
- Chanock RM, Hayflick L, Barile MF. Growth on artificial medium of an agent associated with a typical pneumonia and its identification as a PPLO. Proc Natl Acad Sci USA. 1962;48:41–49. doi: 10.1073/pnas.48.1.41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen TR. In situ detection of mycoplasma contamination in cell cultures by fluorescent Hoechst 33258 stain. Exp Cell Res. 1977;140:255–262. doi: 10.1016/0014-4827(77)90089-1. [DOI] [PubMed] [Google Scholar]
- Dockery DW, Pope CA, III, Xu X, Spengler JD, Ware JH, Fay ME, Ferris BG, Jr, Speizer PE. An association between air pollution and mortality in six US cities. New Engl J Med. 1993;329:1753–1759. doi: 10.1056/NEJM199312093292401. [DOI] [PubMed] [Google Scholar]
- Dreher KL, Jaskot RH, Lehmann JR, Richards JH, McGee JK, Ghio AJ, Costa DL. Soluble transition metals mediate oil fly ash induced acute lung injury. J Toxicol Environ Health. 1995;50:285–305. [PubMed] [Google Scholar]
- Dye JA, Adler KB, Richards JH, Dreher KL. Role of soluble metals in oil fly ash-induced airway epithelial injury and cytokine gene expression. Am J Physiol Lung Cell Mol Physiol. 1999;277:L498–L510. doi: 10.1152/ajplung.1999.277.3.L498. [DOI] [PubMed] [Google Scholar]
- Fabisiak JP, Weiss RD, Powell GA, Dauber JH. Enhanced secretion of immune-modulating cytokines by human lung fibroblasts during in vitro infection with Mycoplasma fermentans. Am J Respir Cell Mol Biol. 1993;8:358–364. doi: 10.1165/ajrcmb/8.4.358. [DOI] [PubMed] [Google Scholar]
- Frampton MW, Ghio AJ, Samet JM, Carson JL, Carter JD, Devlin RB. Effects of aqueous extracts of PM10 filters from Utah Valley on human airway epithelial cells. Am J Physiol Lung Cell Mol Physiol. 1999;277:L960–L967. doi: 10.1152/ajplung.1999.277.5.L960. [DOI] [PubMed] [Google Scholar]
- Ghio AJ, Silbajoris R, Carson JL, Samet JM. Biologic effects of oil fly ash. Environ Heath Prespect. 2002;11(Suppl 1):89–94. doi: 10.1289/ehp.02110s1189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hatch GE, Boykin E, Graham JE, Lewtas J, Pott F, Loud K, Mumford JL. Inhalable particles and pulmonary host defense: In vivo and in vitro effects of ambient and combustion particles. Environ Res. 1985;36:67–80. doi: 10.1016/0013-9351(85)90008-8. [DOI] [PubMed] [Google Scholar]
- Hiura TS, Kaszubowski MP, Li N, Nel AE. Chemicals in diesel exhaust particles generate reactive oxygen radicals and induce apoptosis in macrophages. J Immunol. 1999;163:5582–5591. [PubMed] [Google Scholar]
- Horowitz S, Evinson B, Borer A, Horowitz J. Mycoplasma fermentans in rheumatoid arthritis and other inflammatory arthritides. J Rheumatol. 2000;27:2747–2753. [PubMed] [Google Scholar]
- Huang X, Frenkel K, Klein CB, Costa M. Nickel induces increased oxidants in intact cultured mammalian cells as detected by dichlorofluorescein fluorescence. Toxicol Appl Pharmacol. 1993;120:29–36. doi: 10.1006/taap.1993.1083. [DOI] [PubMed] [Google Scholar]
- Johnson S, Sidebottom D, Bruckner F, Collins D. Identification of Mycoplasma fermentans in synovial fluid samples from arthritis patients with inflammatory disease. J Clin Microbiol. 2000;38:90–93. doi: 10.1128/jcm.38.1.90-93.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kennedy T, Ghio AJ, Reed W, Samet J, Zagorski J, Quay J, Carter J, Dailey L, Hoidal JR, Devlin RB. Copper-dependent inflammation and Nuclear Factor-κB activation by particulate air pollution. Am J Respir Cell Mol Biol. 1998;19:366–378. doi: 10.1165/ajrcmb.19.3.3042. [DOI] [PubMed] [Google Scholar]
- Kovacic R, Launay V, Tuppin P, Lafeuillade A, Feuillie V, Montagnier L, Grau O. Search for the presence of six Mycoplasma species in peripheral blood mononuclear cells of subjects seropositive and seronegative for human immunodeficiency virus. J Clin Microbiol. 1996;34:1808–1810. doi: 10.1128/jcm.34.7.1808-1810.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kraft M, Casell GH, Hensen JE, Watson H, Williamson J, Marimion BP, Gaydos CA, Martin RJ. Detection of Mycoplasma pneumoniae in the airways of adults with chronic asthma. Am J Respir Crit Care Med. 1998;158:998–1001. doi: 10.1164/ajrccm.158.3.9711092. [DOI] [PubMed] [Google Scholar]
- Laden F, Neas LM, Dockery DW, Schwartz J. Association of fine particulate matter from different sources with daily mortality in six U.S. cities. Environ Health Perspect. 2000;108:941–947. doi: 10.1289/ehp.00108941. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leikauf GD. Hazardous air pollutants and asthma. Environ Health Prespect. 2002;110:505–526. doi: 10.1289/ehp.02110s4505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Misra M, Olinski R, Dizaroglu M, Kasprzak KS. Enhancement by L-histidine of nickel(II)-induced DNA–protein cross-linking and oxidative DNA base damage in the rat kidney. Chem Res Toxicol. 1993;6:33–37. doi: 10.1021/tx00031a005. [DOI] [PubMed] [Google Scholar]
- Misra M, Rodriguez RE, Kasprzak KS. Nickel-induced lipid peroxidation in the rat: Correlation with nickel effect on antioxidant defense systems. Toxicology. 1990;64:1–17. doi: 10.1016/0300-483x(90)90095-x. [DOI] [PubMed] [Google Scholar]
- Morgan LG, Usher V. Health problems associated with nickel refining and use. Ann Occup Hyg. 1994;38:189–198. doi: 10.1093/annhyg/38.2.189. [DOI] [PubMed] [Google Scholar]
- Muhlradt PF, Keiss M, Meyer H, Sussmuth R, Jung G. Isolation, structural elucidation, and synthesis of a macrophage stimulatory lipopeptide from Mycoplasma fermentans acting at picomolar concentrations. J Exp Med. 1997;185:1951–1958. doi: 10.1084/jem.185.11.1951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murphy WH, Bullis C, Dabich L, Heyn R, Zarafonetis JD. Isolation of mycoplasma from leukemic and nonleukemic patients. J Natl Cancer Inst. 1970;45:243–251. [PubMed] [Google Scholar]
- Nishiguchi M, Matsumoto M, Takao T, Hoshino M, Shimonishi Y, Tsuji S, Begum NA, Takeuchi O, Akira S, Toyoshima K, et al. Mycoplasma fermentans lipoprotein M161Ag-induced cell activation is mediated by Toll-like receptor 2: Role of N-terminal hydrophobic portion in its multiple functions. J Immunol. 2001;166:2610–2616. doi: 10.4049/jimmunol.166.4.2610. [DOI] [PubMed] [Google Scholar]
- Peters A, Wichmann HE, Tuch T, Heinrich J, Heyder J. Rerspiratory effects are associated with the number of ultrafine particles. Am J Respir Crit Care Med. 1997;155:1376–1383. doi: 10.1164/ajrccm.155.4.9105082. [DOI] [PubMed] [Google Scholar]
- Pope CA, Kanner RE. Acute effects of PM10 pollution on pulmonary function of smokers with mild to moderate chronic obstructive pulmonary disease. Am Rev Respir Dis. 1993;147:1336–1340. doi: 10.1164/ajrccm/147.6_Pt_1.1336. [DOI] [PubMed] [Google Scholar]
- Prahalad AK, Soukup JM, Inmon J, Willis R, Ghio AJ, Becker S, Gallagher JE. Ambient air particles: Effects on cellular oxidant radical generation in relation to particulate elemental chemistry. Toxicol Appl Pharmacol. 1999;158:81–91. doi: 10.1006/taap.1999.8701. [DOI] [PubMed] [Google Scholar]
- Pritchard R, Ghio AJ, Lehmann JR, Winsett DW, Tepper JS, Park P, Gilmour MI, Dreher KL, Costa DL. Oxidant generation and lung injury after particulate air pollution exposure increase with the concentration of associated metals. Inhal Toxicol. 1996;8:457–477. [Google Scholar]
- Quay JL, Reed W, Samet J, Devlin RB. Air pollution particles induce IL-6 gene expression in human epithelial cells via NF-κB activation. Am J Respir Cell Mol Biol. 1998;19:98–106. doi: 10.1165/ajrcmb.19.1.3132. [DOI] [PubMed] [Google Scholar]
- Rawadi G, Garcia J, Lemercier B, Roman-Roman S. Signal transduction pathways involved in the activation of NF-κB, AP-1, and c-fos by Mycoplasma fermentans membrane lipoproteins in macrophages. J Immunol. 1999;162:2193–2203. [PubMed] [Google Scholar]
- Rawadi G, Roman-Roman S. Mycoplasma membrane lipoproteins induce proinflammatory cytokines by a mechanism distinct from that of lipopolysaccharide. Infect Immun. 1996;64:637–643. doi: 10.1128/iai.64.2.637-643.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rodwell AW, Whitcomb RH. Methods for direct and indirect measurement of mycoplasma growth. In: Tully JG, Razin S, editors. Methods in Mycoplasmology. Academic Press; New York: 1983. pp. 185–196. [Google Scholar]
- Rottem S. Interaction of mycoplasmas with host cells. Physiol Rev. 2002;83:417–432. doi: 10.1152/physrev.00030.2002. [DOI] [PubMed] [Google Scholar]
- Ruiter M, Wentholt HMM. A pleuropneumonia-like organism in primary fusopirochetal gangrene of the penis. J Invest Dermatol. 1950;15:301–304. doi: 10.1038/jid.1950.104. [DOI] [PubMed] [Google Scholar]
- Ruuth E, Praz F. Interactions between mycoplasmas and the immune system. Immunol Rev. 1989;112:133–160. doi: 10.1111/j.1600-065x.1989.tb00556.x. [DOI] [PubMed] [Google Scholar]
- Saillard C, Carle P, Bove JM, Bebear C, Lo SC, Shih JWK, Wang RYH, Rose DL, Tully JG. Genetic and serologic relatedness between M. fermentans and a mycoplasma recently identified in tissues of AIDS and non-AIDS patients. Res Virol. 1990;141:441–448. doi: 10.1016/0923-2516(90)90010-g. [DOI] [PubMed] [Google Scholar]
- Samet JM, Dominici F, Curriero FC, Coursac I, Zeger SL. Fine particulate air pollution and mortality in 20 U.S. cities. N Engl J Med. 2000;343:1742–1749. doi: 10.1056/NEJM200012143432401. [DOI] [PubMed] [Google Scholar]
- Samet JM, Graves LM, Quay J, Dailey LA, Devlin RB, Ghio AJ, Wu W, Bromberg PA, Reed W. Activation of MAPKs in bronchial epithelial cells exposed to metals. Am J Physiol Lung Cell Mol Physiol. 1998;275:L551–L558. doi: 10.1152/ajplung.1998.275.3.L551. [DOI] [PubMed] [Google Scholar]
- Samet JM, Silbajoris R, Huang T, Jaspers I. Transcription factor activation following exposure of an intact lung preparation to metallic particulate matter. Environ Health Prespect. 2002;110:985–990. doi: 10.1289/ehp.02110985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Samet JM, Stonehuerner J, Reed W, Devlin RB, Dailey LA, Kennedy TP, Bromberg PA, Ghio AJ. Disruption of protein tyrosine phosphate homeostasis in bronchial epithelial cell exposed to oil fly ash. Am J Physiol Lung Cell Mol Physiol. 1997;272:L426–L432. doi: 10.1152/ajplung.1997.272.3.L426. [DOI] [PubMed] [Google Scholar]
- Schwartz J, Slater D, Larson TV, Pierson WE, Koenig JQ. Particulate air pollution and hospital emergency room visits for asthma in Seattle. Am Rev Respir Dis. 1993;147:826–831. doi: 10.1164/ajrccm/147.4.826. [DOI] [PubMed] [Google Scholar]
- Shi X, Dalal NS, Kasprzak KS. Generation of free radicals from lipid hydroperoxides by Ni2+ in the presence of oligopeptides. Arch Biochem Biophys. 1992;299:154–162. doi: 10.1016/0003-9861(92)90257-w. [DOI] [PubMed] [Google Scholar]
- Shibata K, Kaga M, Kudo M, Dong L, Hasebe A, Domon H, Sato Y, Oguchi H, Watanabe T. Detection of Mycoplasma fermentans in saliva sampled from infants, preschool, and school children, adolescents and adults by a polymerase chain reaction-based assay. Microb Immunol. 1999;43:521–525. doi: 10.1111/j.1348-0421.1999.tb02437.x. [DOI] [PubMed] [Google Scholar]
- Takeuchi O, Kawai T, Muhlradt PF, Morr M, Radolf JD, Zychlinsky A, Takeda K, Akira S. Discrimination of bacterial lipoproteins by toll-like receptor 6. Intl Immunology. 2001;13(7):933–940. doi: 10.1093/intimm/13.7.933. [DOI] [PubMed] [Google Scholar]
- Tully JG, Whitcomb RF, Clark HF, Williamson DL. Pathogenic mycoplasmas: Cultivation and vertebrate pathogenicity of a new spiroplasma. Science. 1977;195:892–894. doi: 10.1126/science.841314. [DOI] [PubMed] [Google Scholar]
- van Eeden SF, Tan WC, Suwa T, Mukae H, Terashima T, TF, Qui D, Vincent R, Hogg JC. Cytokines involved in the systemic inflammatory response induced by exposure to particulate matter air pollutants (PM10) Am J Respir Crit Care Med. 2001;164:826–830. doi: 10.1164/ajrccm.164.5.2010160. [DOI] [PubMed] [Google Scholar]
- Vojdani A, Choppa PC, Tagle C, Andrin R, Samimi B, Lapp CW. Detection of Mycoplasma genus and Mycoplasma fermentans by PCR in patients with chronic fatigue syndrome. FEMS Immunol Med Microbiol. 1998;22:355–365. doi: 10.1111/j.1574-695X.1998.tb01226.x. [DOI] [PubMed] [Google Scholar]
- Weibel ER. Lung cell biology. In: Fishman AP, editor. Handbook of Physiology: The Respiratory System I. Circulation and nonrespiratory functions. American Physiological Society; Betheseda, MD: 1985. pp. 47–91. [Google Scholar]
- Winzen R, Kracht M, Ritter B, Wilhelm A, Chen AYA, Shyum AB, Muller M, Gaestel M, Resch K, Holtmann H. The p38 MAP kinase pathway signals for cytokine-induced mRNA stabilization via MAP-kinase-activated protein kinase 2 and an AU-rich region-targeted mechanism. EMBO J. 1999;18(18):4969–4980. doi: 10.1093/emboj/18.18.4969. [DOI] [PMC free article] [PubMed] [Google Scholar]