

Abstract
In an attempt to develop alternative medicine for the treatment of diabetes and related complications, the antidiabetic potential of the fruits of Crataegus pinnatifida was evaluated. The antidiabetic potential of the methanol (MeOH) extract as well as different solvent soluble fractions of the fruits of C. pinnatifida was evaluated via α-glucosidase, protein tyrosine phosphatase 1B (PTP1B), rat lens aldose reductase (RLAR), and advanced glycation end products (AGEs) formation inhibitory assays. The MeOH extract showed potent inhibitory activity against α-glucosidase, PTP1B, and AGEs formation with IC50 values of 122.11, 3.66 and 65.83 μg/ml respectively, while it showed moderate inhibitory activity against RLAR with the IC50 value of 160.54 μg/ml. Among different fractions, the ethyl acetate (EtOAc) and the dichloromethane (CH2Cl2) fractions were found as active fractions exhibiting potential α-glucosidase, PTP1B, RLAR inhibitory, and AGEs formation inhibitory activities. Seven compounds including hyperoside, chlorogenic acid, ursolic acid, oleanolic acid, 3-epicorosolic acid, β-sitosterol, β-sitosterol glucoside were isolated from these two fractions. 3-Epicorosolic acid showed both potent α-glucosidase and PTP1B inhibitory activities with IC50 values of 30.18 and 4.08 μg/ml respectively. Moreover, kinetic study revealed that 3-epicorosolic acid showed mixed type inhibition against PTP1B, while it showed uncompetitive inhibition against α-glucosidase. Therefore, these results suggest that the fruits of C. pinnatifida and its constituents have potential antidiabetic activity which might be used as a functional food for the treatment of diabetes and associated complications.
Keywords: Crataegus pinnatifida, Antidiabetic activity, 3-epicorosolic acid, Kinetics
INTRODUCTION
Diabetes mellitus is a progressive metabolic disorder of glucose metabolism in which the body either does not produce enough insulin or does not respond to the produced insulin, resulting in an increase of blood glucose levels and causing serious and irreparable damage to body systems, such as blood vessels and nerves (1). In Asia, the prevalence of diabetes is increasing at an alarming rate and expected to rise two or three folds by 2030 (2). All forms of diabetes are characterized by chronic hyperglycaemia and the development of diabetes-specific microvascular and macro-vascular complications which affect major organs such as eye, kidney, heart, brain, and lower extremities. Accumulated evidences suggested the four main hypotheses linked to hyperglycemia induced complications in diabetic patients: increased polyol pathway flux, increased advanced glycation end products, activation of protein kinase C, and increased hexosamine pathway flux (3). The control of hyperglycemia is therefore of prime importance to halt the progression of the disease. Hence, a number of viable pharmacologic options such as insulin, insulin secretagogues, and insulin sensitizers constitute a base line of therapy in order to maintain normoglycemia in established diabetes mellitus (4). The suppression of postprandial hyperglycemia subsequently delays the progression of micro and macro-vascular complications such as microangiopathy, cardio-vascular, and cerebrovascular diseases (5). However, pharmacologic maintenance of normoglycemia cannot prevent the eventual complications such as nephropathy, neuropathy and retinopathy (6). On the other hand, the modern medicines that are currently available for the management of diabetes mellitus exert serious side effects such as hepatotoxicity, abdominal pain, flatulence, diarrhea, and hypoglycemia (7,8). Therefore, researchers are continuously trying to find out new antidiabetic agents with high efficacy and low side effects for the management of diabetic mellitus.
Crataegus pinnatifida Bge. also known as Chinese hawthorn or ShanZha belonging to the family Rosaceae, is widely distributed throughout the northern temperate regions of the world with approximately 280 species, primarily in East Asia, Europe and North America (9). Traditionally, the fruits of C. pinnatifida have been used as food and medicine in the treatment of chronic heart failure, high blood pressure and various digestive ailments and as peptic agent. It has documented that the hot water extract exhibited free radical scavenging activity, inhibited low-density lipoprotein oxidation, decreased the serum total cholesterol, and triglycerides in hyperlipidemic humans (10,11). Recent studies demonstrated that the fruits exhibited potential antioxidant, anti-diabetic, anti-inflammatory, and anticancer activities (12,13,14,15). The recent studies showed that ShanZha has an ability to reduce the body weight and improve the dyslipidemia and suppress high cholesterol diet induced hypercholesterolemia in animal (16,17). The major components of C. pinnatifida include flavonoids, proanthocyanidin, triterpenes, organic acids, tannin, flavane and its polymers, among which flavonoids and triterpenes were reported as the main active hypolipidemic constituents (9,18). As a part of our continuous search for antidiabetic agents from natural sources, we investigated the methanol (MeOH) extract as well as its different solvent soluble fractions together with the isolated compounds from the active fractions of C. pinnatifida fruits via α-glucosidase, protein tyrosine phosphatase (PTP1B), rat lens aldose reductase (RLAR), and advanced glycation end products (AGEs) formation inhibitory assays.
MATERIALS AND METHODS
Chemicals and reagents
Yeast α-glucosidase, acarbose, p-nitrophenyl phosphate (pNPP), p-nitrophenyl α-D-glucopyranoside (pNPG), ethylenedi-aminetetraacetic acid (EDTA), nicotinamide adenine dinucleotide phosphate (NADPH), DL-glyceraldehyde dimer, quercetin, bovine serum albumin (BSA), D-(-)-fructose, D-(+)-glucose, aminoguanidine hydrochloride were purchased from Sigma-Aldrich (St. Louis, MO, USA). PTP1B (human recombinant) was purchased from Biomol® International LP (Plymouth Meeting, PA, USA), and dithiothreitol (DTT) was purchased from Bio-Rad Laboratories (Hercules, CA, USA). All other chemicals and solvents used were purchased from E. Merck, Fluka, and Sigma-Aldrich, unless otherwise stated.
Plant materials
The fruits of C. pinnatifida were purchased from Republic of Korea in the middle of 2010. A voucher specimen of the fruits registered (20100712) and deposited to the laboratory of Prof. J. H. Lee, Dongguk University, Gyeongju, Republic of Korea for future reference.
Extraction and fractionation
The fruits of C. pinnatifida were dried and ground to powder. Then the powder of the fruits (5 kg) was refluxed in MeOH for 3 h (5 L × 3 times). The total filtrate was then concentrated to dryness in vacuo at 40°C in order to render the MeOH extract (1.31 kg). This extract was suspended in distilled water (H2O) and then successively partitioned with methylene chloride (CH2Cl2), ethyl acetate (EtOAc), and n-butanol (n-BuOH) to yield the CH2Cl2 (136.97 g), EtOAc (66.86 g), and n-BuOH (318.53 g) fractions, respectively, as well as a H2O residue (787.64 g).
α-Glucosidase inhibitory assay
The enzyme inhibition studies were carried out spectrophotometrically in a 96-well microplate reader using a procedure reported by Li and coworkers (19). A total 60 μl reaction mixture containing 20 μl of 100 mM phosphate buffer (pH 6.8), 20 μl of 2.5 mM pNPG in the buffer, and 20 μl of sample dissolved in 10% DMSO, were added to each well, followed by 20 μl of 10 mM phosphate buffer (pH 6.8) containing 0.2 U/ml α-glucosidase to the mixture of treatment terminated wells. The plate was incubated at 37°C for 15 min, then 80 μl of 0.2 mM sodium carbonate solution was added to stop the reaction. Right after that, absorbance was recorded at 405 nm using VERSA max (Molecular Devices, Sunnyvale, CA, USA) microplate reader. Control contained the same reaction mixture except the same volume of phosphate buffer was added instead of sample solution. Acarbose was dissolved in 10% DMSO and used as a positive control. The inhibition percentage (%) was calculated as: [(Ac – As)/Ac]×100 %, where, Ac is the absorbance of the control, and As, the absorbance of the sample.
Protein tyrosine phosphatase inhibitory assay
The inhibitory activity of the plant extracts against human recombinant PTP1B was evaluated using pNPP as substrate (20). To each 96 well (final volume 110 μl) were added 2 mM pNPP and PTP1B in a buffer containing 50 mM citrate (pH 6.0), 0.1 mM NaCl, 1 mM EDTA, and 1 mM DTT with or without sample. The plate was preincubated at 37°C for 10 min, and then 50 μl of pNPP in buffer was added. Following incubation at 37°C for 30 min, the reaction was terminated with the addition of 10 M NaOH. The amount of p-nitrophenyl produced after enzymetic dephosphorylation was estimated by measuring the absorbance at 405 nm using VERSA max (Molecular Devices, Sunnyvale, CA, USA) microplate reader. The nonenzymatic hydrolysis of 2 mM PNPP was corrected by measuring the increase in absorbance at 405 nm obtained in absence of PTP1B enzyme. The inhibition percentage (%) was calculated as: [(Ac – As)/Ac] × 100 %, where, Ac is the absorbance of the control, and As, the absorbance of the sample. Ursolic acid was used as positive control.
Rat lens aldose reductase inhibitory assay
In this experiment we followed the Guidelines for Care and Use of Laboratory Animals as approved by Pukyong National University. According to the modified method of Hayman and Kinoshita (1965), rat lens homogenate was prepared (21). Briefly, the lens were removed from the eyes of Sprague-Dawley rats (Samtako BioKorea, Inc.) weighing 250–280 g. The lens were homogenized in sodium phosphate buffer (pH 6.2), which was prepared from sodium phosphate dibasic (Na2HPO4.H2O, 0.66 g) and sodium phosphate monobasic (NaH2PO4.H2O, 1.27 g) in 100 ml of double distilled water. The supernatant was obtained by centrifugation of the homogenate at 10,000 rpm at 4°C for 20 min and was frozen until use. A crude RLAR homogenate, with a specific activity of 6.5 U/mg, was used in the evaluations for enzyme inhibition. Reaction solution consists of 620 μl of 100 mM sodium phosphate buffer (pH 6.2), 90 μl of RLAR homogenate, 90 μl of 1.6 mM NADPH, 9 μl of the samples (the extracts and fractions dissolved in 100% DMSO) or 9 μl of 100% DMSO, and 90 μl of 50 mM of DL-glyceraldehyde as the substrate. The RLAR activity was determined by measuring the decrease in NADPH absorption at 340 nm over a 4 min period using an Ultrospec®2100 pro UV/Visible spectrophotometer with SWIFT II Applications software (Amersham Biosciences, New Jersey, USA). Quercetin, a well-known aldose reductase inhibitor, was used as reference standard. The inhibition percentage (%) was calculated as [1 – (ΔA sample/min – ΔA blank/min)/(ΔA control/min – ΔA blank/min)] × 100%, where ΔA sample/min represents the reduction of absorbance for 4 min with the test sample and substrate, respectively, and ΔA control/min represents the same, but with 100 % DMSO instead of a sample. The IC50 values are expressed as means ± SEM of triplicate experiments.
Advanced glycation end products formation inhibitory assay
The inhibitory activity of AGEs formation was examined according to the modified method of Vinson and Howard (22). To prepare the AGE reaction solution, 10 mg/ml of bovine serum albumin in 50 mM sodium phosphate buffer (pH 7.4) was added to 0.2 mM fructose, 0.2 mM glucose, and 0.02% sodium azide to prevent bacterial growth. The reaction mixture (950 μl) was then mixed with various concentrations of the samples dissolved in 10 % DMSO. After incubating at 37°C for 7 days, the fluorescence intensity of the reaction products was determined on a spectrofluorometric detector (FLx800 microplate fluorescence reader, Bio-Tek Instrument, Inc., Winooski, USA) with excitation and emission wavelengths at 350 nm and 450 nm, respectively. The IC50 values are expressed as means ± SEM of triplicate experiments. Aminoguanidine hydrochloride, a nucleophilic hydrazine compound, was used as a reference in the AGEs formation inhibitory assay.
Isolation of the active compounds from ethyl acetate and dichloromethane fractions
Since both EtOAc and CH2Cl2 soluble fractions of the MeOH extract of C. pinnatifida exhibited promising antidiabetic potential, these two fractions were selected for column chromatography to isolate active compounds. The EtOAc fraction was subjected to column chromatography on a silica gel column with a solvent mixture of hexane-EtOAc→EtOAc→EtOAc-MeOH (10:1→0:1→1:1) to get 22 subfractions (E1 to E22). Subfraction 9 was chromatographed repeatedly on a silica gel column with a solvent mixture of CH2Cl2-MeOH-H2O (7:1:0.1→0:1:0) to obtain 4 subfractions (E9-1 to E9-4). The subfraction E9-2 and E9-4 was chromatographed on a Sephadex LH-20 column with MeOH to get hyperoside (10 mg) and chlorogenic acid (11 mg). On the other hand, the CH2Cl2 fraction (136.97 g) was subjected to column chromatography on a silica gel column with a solvent mixture of CH2Cl2-MeOH (10:1→5:1→1: 1) to get 17 subfractions (C1 to C17). Among them, subfraction C9 was filtered to get precipitate as white powder as β-sitosterol glucoside 27 mg). From subfraction C8, β-sitosterol (36 mg) was found as white precipitate. Then subfraction C8 was chromatographed over silica gel column using a solvent mixture of hexane-acetone (10:1→1:1) to obtain 10 subfractions (C8-1 to C8-10). Repeated chromatography of subfractions C8-4 yielded oleanolic acid (41 mg). Subfractions C8-7 was chromatographed over a silica gel column with a solvent mixture of CH2Cl2:EtOAc (100:1→0:1) to get 3-epicorosolic acid (49 mg) and ursolic acid (2.01 g). Structures of the compounds isolated from two active fractions of C. pinnatifida are given in Fig. 1. All the compounds were identified based on their spectroscopic data including 1H and 13C-NMR as well as compared with the published data and TLC with standard.
Fig. 1.
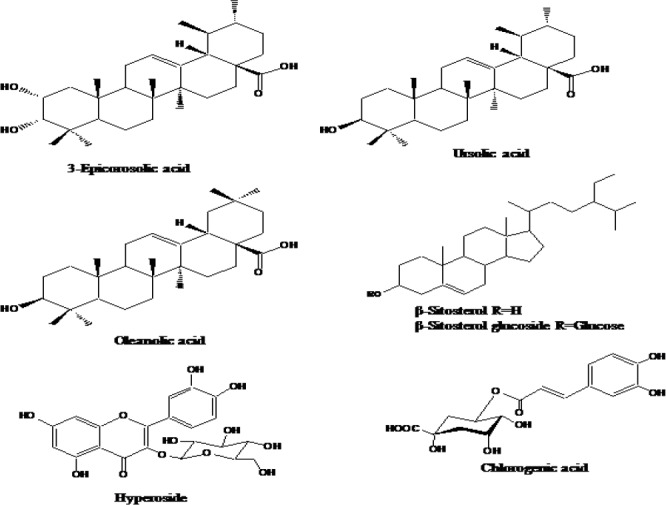
Structures of the compounds isolated from two active fractions of C. pinnatifida.
Kinetic analysis of 3-epicorosolic acid against α-glucosidase and protein tyrosine phosphatase enzymes
In order to determine the inhibition mechanism, each enzymatic inhibition at various concentrations of 3-epicorosolic acid was evaluated by monitoring the effect of different concentration of the substrate via Dixon plot (a single reciprocal plot).
Dixon plot for the inhibition of α-glucosidase by 3-epicorosolic acid obtained in the presence of different concentrations of pNPG substrate: 0.625 mM, 1.25 mM, and 2.5 mM. The test concentrations for the 3-epicorosolic acid in the α-glucosidase kinetic analysis were 33.05 and 16.52 μM. Dixon plot for PTP1B inhibition by the 3-epicorosolic acid was obtained in the presence of different concentrations of the pNPP substrate: 4 mM, 2 mM and 1 mM. The test concentrations of the 3-epicorosolic acid used in the PTP1B kinetic analysis was 21.19, 4.24 and 0.85 μM. Both enzymatic procedures applied the same aforementioned PTP1B and α-glucosidase assay methods. The inhibition constants (Ki) were determined by interpreting the Dixon plots, in which the value of the x-axis implies (Ki).
Statistical analysis
Statistical analyses were performed using Microsoft EXCEL. All results are expressed as the Mean ± Standard error of the Mean of triplicate experiments. Regression analysis was carried out by best fit methods to determine IC50 values.
RESULTS
α-Glucosidase inhibitory activity of the methanol extract and the solvent soluble fractions of Crataegus pinnatifida
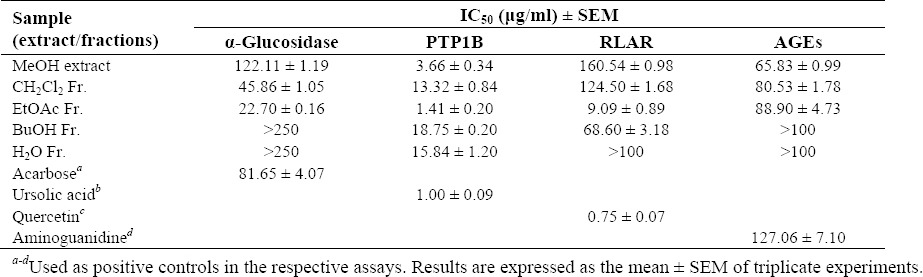
To evaluate the antidiabetic activity of the MeOH extract and the solvent soluble fractions of the fruits of C. pinnatifida, the α-glucosidase inhibitory activity was evaluated. The inhibitory activity is presented in Fig. 2. Among the tested fractions the EtOAc fraction has been found as the most active fraction with the IC50 value of 22.70 μg/ml compared to the positive control acarbose with the IC50 value of 81.65 μg/ml. The CH2Cl2 fraction also exhibited potent inhibitory activity with the IC50 value of 45.86 μg/ml, while the n-BuOH and H2O fractions did not show inhibitory activity within the tested concentrations (Table 1).
Fig. 2.
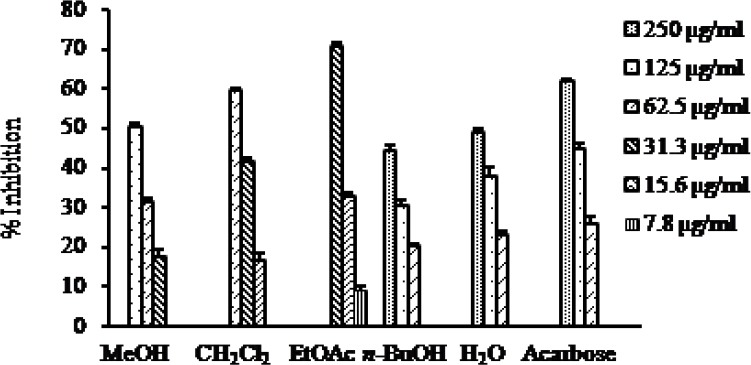
α-Glucosidase inhibitory activity of the MeOH extract as well as different solvent soluble fractions of C. pinnatifida.
Table 1.
Summary of the antidiabetic potential of the MeOH extract as well as different solvent soluble fractions of C. pinnatifida.
Protein tyrosine phosphatase inhibitory activity of the methanol extract and the solvent soluble fractions of Crataegus pinnatifida
The inhibitory activity of the MeOH extract and its different solvent soluble fractions of C. pinnatifida against PTP1B is presented in Fig. 3. The MeOH extract and all the fractions showed PTP1B inhibitory activity in a concentration dependent manner with IC50 values ranging from 1.41 to 18.75 μg/ml. Among the fractions, the EtOAc fraction showed the highest inhibitory activity with the IC50 value of 1.41 μg/ml followed by the CH2Cl2 fraction with the IC50 value of 13.32 μg/ml compared to the positive control ursolic acid with the IC50 value of 1.00 μg/ml (Table 1).
Fig. 3.
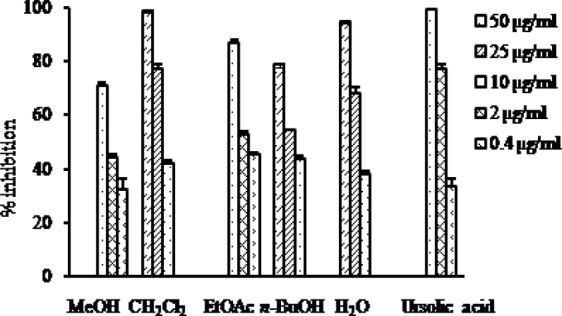
PTP1B inhibitory activity of the MeOH extract as well as different solvent soluble fractions of C. pinnatifida.
Rat lens aldose reductase inhibitory activity of the methanol extract and the solvent soluble fractions of Crataegus pinnatifida
The RLAR inhibitory activity of the MeOH extract and its solvent soluble fractions of the fruits of C. pinnatifida is presented in Fig. 4. It is evident from the Fig. 4 that the EtOAc fraction showed the highest RLAR inhibitory activity with the IC50 value of 9.09 μg/ml compared to the positive control quercetin with the IC50 value of 0.75 μg/ml. In addition, the CH2Cl2 fractions also exhibited remarkable RLAR inhibitory activities with the IC50 value of 124.50 μg/ml, while the n-BuOH fractions showed poor inhibitory activity with the IC50 value of 68.60 μg/ml. On the other hand, H2O fraction did not exhibit inhibitory activity against RLAR at the concentration tested (Table 1).
Fig. 4.
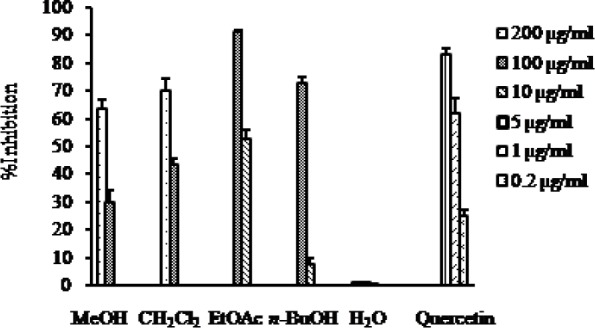
RLAR inhibitory activity of the MeOH extract as well as different solvent soluble fractions of C. pinnatifida.
Advanced glycation end products formation inhibitory activity of the methanol extract and the solvent soluble fractions of Crataegus pinnatifida
AGEs formation inhibitory activity of the MeOH extract and different solvent soluble fractions of C. pinnatifida is presented in the Fig. 5. As it is shown in Fig. 5, the MeOH extract exhibited strong AGEs formation inhibitory activity with the IC50 value of 65.83 μg/ml compared to the positive control aminoguanidine with the IC50 value of 127.06 μg/ml. Among the different fractions, the CH2Cl2 fraction and the EtOAc fraction showed similar type inhibition against AGEs formation with their corresponding IC50 values of 80.53 and 88.90 μg/ml, respectively which was much stronger (1.5 times) than the positive control aminoguanidine. Similar to the RLAR inhibitory assay, n-BuOH and H2O fractions were found inactive against AGEs formation at the concentration tested (Table 1).
Fig. 5.
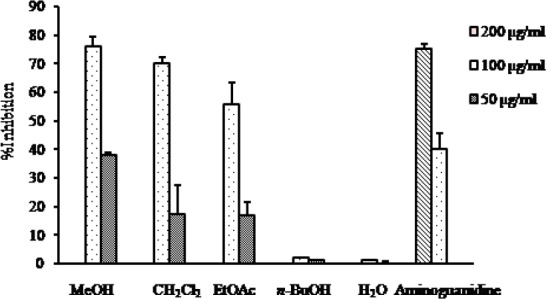
AGEs formation inhibitory activity of the MeOH extract as well as different solvent soluble fractions of C. pinnatifida.
α-Glucosidase and protein tyrosine phos-phatase inhibitory activities of 3-epicorosolic acid
In the present study we found that 3-epicorosolic acid showed 3 times potent α-glucosidase inhibitory activity with the IC50 value of 30.18 μg/ml compared to the positive control acarbose with the IC50 value of 92.80 μg/ml (Table 2). On the other hand, it also exhibited potent PTP1B inhibitory activity with the IC50 value of 4.08 μg/ml compared to the positive control ursolic acid with the IC50 value of 1.17 μg/ml (Table 2). Since it showed potent α-glucosidase and PTP1B inhibitory activities we further performed kinetic study in order to find out type of enzyme inhibition.
Table 2.
Summary of αglucosidase and PTP1B inhibitory activities of 3-epicorosolic acid.
Kinetic analysis of 3-epicorosolic acid
In an attempt to explain the mode of enzymatic inhibition, kinetic analyses were performed at different concentrations of the corresponding substrate (pNPG for α-glucosidase and pNPP for PTP1B) and various inhibitors concentrations. Dixon plots are the graphical method [a plot of 1/enzyme velocity (1/V) against concentration (I)] for determining the type of enzyme inhibition and dissociation or inhibition constant (Ki) for an enzyme inhibitor complex, and were easily determined. Fig. 6 and 7 demonstrate the enzymatic kinetic analysis of 3-epicorosolic acid against α-glucosidase and PTP1B. 3-epicorosolic acid showed uncompetitive inhibition against α-glucosidase, while it showed mixed type inhibition against PTP1B enzyme with Ki value of 3.08 μM. Since the Ki value represents the concentrations to form an enzyme-inhibitor complex, this value plays an important role in the development of preventive and therapeutic agent.
Fig. 6.

Dixon plot for inhibition of 3-epicorosolic acid on α-glucosidase in the presence of different concentrations of substrate: 0.625 mM (●); 1.25 mM (○); and 2.5mM (▼) showed uncompetitive inhibition.
Fig. 7.
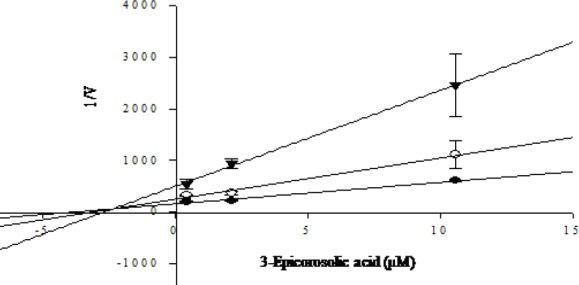
Dixon plot for inhibition of 3-epicorosolic acid on PTP1B in the presence of different concentrations of substrate: 4 mM (●); 2 mM (○); and 1 mM (▼) showed mixed type inhibition.
DISCUSSION
As a part of our continuous search for anti-diabetic agents from natural sources we investigated the MeOH extract as well as different solvent soluble fractions of C. pinnatifida fruits against α-glucosidase, PTP 1B, RLAR, and AGEs formation. Our data revealed that the MeOH extract as well as different solvent soluble fractions of C. pinnatifida fruits possess promising anti-diabetic and anti-diabetic complications potential which are attributed to its potent inhibitory activity against α-glucosidase, PTP1B, RLAR, and AGEs formation.
α-glucosidase is an enzyme which catalyzes the cleavage of glycosidic bonds in oligosaccharides or glycoconjugates to prevent their immediate breakdown into mono-saccharides, which then would be absorbed quickly into the bloodstream. Delaying the absorption of carbohydrates gives the β-cells in the pancreas more time to secrete adequate insulin to cover the meal (23). Thus in the present study, we investigated the α-glucosidase inhibitory activity of the MeOH extract and different solvent soluble fractions of the fruits of C. pinnatifida. Our study showed that the EtOAc fractions showed four times more portent α-glucosidase inhibitory activity, while CH2Cl2 fraction showed two times more potent inhibitory activity compared to acarbose, a commercially used α-glucosidase inhibitor. Although Chunmai and coworkers demonstrated poor α-glucosidase inhibitory activity of the MeOH extract of fruits of C. pinnatifida, we found moderate inhibitory activity of the MeOH extract (14). However, we found strong inhibitory activities of the EtOAc and CH2Cl2 fractions. These discrepancies between two results of the crude MeOH extract are probably due to difference in the time of collection and sources of the fruits or the method of extraction. On the other hand, strong inhibitory activities of the EtOAc and CH2Cl2 fractions are resulted from the differential distribution of the active compounds in these two fractions.
PTP1B, a key regulator of both insulin and leptin signaling pathway, is localized to the cytoplasmic face of the endoplasmic reticulum and is expressed ubiquitously, including in the classical insulin-targeted tissues such as liver, muscle and fat (24). PTP1B can associate with and dephosphorylate both activated insulin receptor or insulin receptor substrates. Clinical studies have found a correlation between insulin resistance states and levels of PTP1B expression in muscle and adipose tissues, suggesting that PTP1B has a major role in the insulin resistance associated with obesity and non-insulin dependent diabetes (25,26). Thus, inhibition of PTP1B has been demonstrated to be an effective therapeutic approach for insulin-sensitive drugs targeted for the treatment and prevention of metabolic disorders. To find out the potent PPTP1B inhibitors, the MeOH extract as well as different fractions of C. pinnatifida were evaluated against PTP1B. The MeOH extract and its different fractions showed potent PTP1B inhibitory activity. Among the fractions, the EtOAc fraction showed potent PTP1B inhibitory activity which is similar to the positive control ursolic acid. In addition, the CH2Cl2 fraction also showed moderate PTP1B inhibitory activity compared to the EtOAc fraction.
Diabetes mellitus associated complications can also arise from increased flux of glucose through the polyol pathway. Aldose reductaase, a cytosolic monomeric oxidoreductase, is the first enzyme in the polyol pathway (3). Under normoglycemic condition aldose reductase has low affinity for glucose and a small percentage of glucose is metabolized by this pathway whereas under hyperglycemic condition, increased intracellular glucose results in increased enzymatic conversion to sorbitol, with concomitant decreases in NADPH (3,27). The increased polyol pathway flux leads to accumulation of sorbitol in the lens fiber, which causes an influx of water, generation of osmotic stress, and cataract formation (28). Due to the poor penetration across membranes and inefficient metabolism, sorbitol and its metabolites accumulate in the nerves, retina, and kidneys, cause development of diabetic complications, including retinopathy, neuropathy, and nephropathy (2). Therefore, inhibition of aldose reductase has been emerged as an important therapeutic approach for preventing and reducing long term diabetic complications. In our investigation, the EtOAc fraction of C. pinnatifida showed potent RLAR inhibitory activity followed by n-BuOH fraction. In addition, CH2Cl2 fraction also exhibited moderate inhibitory activity against RLAR.
In addition to the increased polyol pathway flux, prolonged hyperglycemia also accelerates the formation of AGEs in body tissues. AGEs are a heterogeneous group of substances, involves a series of reactions between the carbonyl group on reducing sugars and the amino group on proteins, nucleic acids, or phospholipids, forming a Schiff base followed by an Amadori rearrangement and further non-oxidative and/or oxidative modifications, which lead to the formation of AGEs (30,31). Enhanced formation and accumulation of AGEs have been implicated as a major pathogenic process leading to diabetic complications such as peripheral neuropathy, vascular damage, arterial wall stiffening, decreased myocardial compliance, normal aging, atherosclerosis, and Alzheimer's disease (32,33,34). In our present study the CH2Cl2 farction showed the highest AGEs formation inhibitory activity followed by the EtOAc fraction. Moreover, the CH2Cl2 and EtOAc fraction showed similar type inhibition against AGEs formation which is much stronger (1.5 times) than the positive control aminoguanidine.
Based on results obtained from all assays, both EtOAc and CH2Cl2 fractions of the MeOH extract of C. pinnatifida have been shown to exhibit promising antidiabetic potential. Therefore, these two fractions were selected for column chromatography to isolate active compounds. Repeated column chromatography of the EtOAc fraction yielded hyperoside and chlorogenic acid, while the CH2Cl2 fraction yielded three triterpenic acid including ursolic acid, oleanolic acid and 3-epicorosolic acid along with β-sitosterol and β-sitosterol glucoside. Although 3-epicorosolic acid was isolated from other species, it is the first report of 3-epicorosolic acid isolated from the fruits of C. pinnatifda. Two compounds hyperoside and chlorogenic acid isolated from the EtOAc fraction have been already reported to possess potent α-glucosidase, PTP1B, RLAR, and AGEs formation inhibitory activities (35,40). On the other hand, two triterpenic acids, ursolic acid and oleanolic acid have been already reported as potent inhibitors of α-glucosidase and PTP1B enzymes (41,42). In addition, ursolic acid showed noncompetitive inhibition against α-glucosidase while competitive inhibition against PTP1B enzyme (41,43). Yin and Chan reported that ursolic acid and oleanolic acid inhibited the nonenzymatic formation of glycative products such as pentosidine and carboxymethyllysine (44). In addition, oleanolic acid and ursolic acid markedly suppressed renal aldose reductase activity and enhanced glyoxalase I activity, which contributed to decrease renal AGEs formation and improve renal function (45). Therefore, it is anticipated that the presence of these compounds largely contributed to the antidiabetic effect of the fruits of C. pinnatifida. Since antidiabetic potential of all the compounds except 3-epicorosolic acid were extensively investigated, only 3-epicorosolic acid was selected for further investigation. 3-Epicorosolic acid showed 3 fold strong inhibitory potential than acarbose against α-glucosidase while slightly lower than ursolic acid against PTP1B enzyme. Since it showed strong inhibitory activity against both α-glucosidase and PTP1B enzymes, we performed kinetic analysis in order to explain the mode of enzyme inhibition. Kinetic analyses revealed that 3-epicorosolic acid showed uncompetitive inhibition against α-glucosidase, while it showed mixed type inhibition against PTP1B enzyme.
CONCLUSION
Considering the importance and severity of diabetic complications, mainly cardiovascular diseases, nephropathies, retinopathies and neuropathies, any new therapeutic innovation is of interest to prevent deleterious effects of hyperglycemia. In the present study, the MeOH extract as well as different solvent soluble fractions of C. pinnatifida showed promising inhibitory potential against α-glucosidase, PTP1B, RLAR, and AGEs formation which might be attributed to the presence of triterpinic acid (ursolic acid, oleanolic acid, 3-epicorosolic acid), hyperoside and chlorogenic acid. Taken together all these results demonstrate the promising antidiabetic potential of the fruits of C. pinnatifida which could be used as functional food in the treatment of diabetes and diabetes related complications. Moreover, 3-epicorosolic acid isolated first time from C. pinnatifida fruits exhibited strong inhibitory activities against α-glucosidase and PTP1B which could be further explored in order to develop therapeutic agents for the treatment of diabetes and related disease.
ACKNOWLEDGMENT
This work was financially supported by the Blue-Bio Industry Regional Innovation Center (RIC) at Dong-Eui University as a RIC program of Korea Institute of Industrial Technology Evaluation and Planning (ITEP) under Ministry of Knowledge Economy (MKE) and Busan City, and the Food and Drug Administration, Republic of Korea (KFDA).
REFERENCES
- 1.Matsui T, Tanaka T, Tamura S, Toshima A, Tamaya K, Miyata Y, et al. Alpha-glucosidase inhibitory profile of catechins and theaflavins. J Agric Food Chem. 2007;55:99–105. doi: 10.1021/jf0627672. [DOI] [PubMed] [Google Scholar]
- 2.Shaw JE, Sicree RA, Zimmet PZ. Global estimates of the prevalence of diabetes for 2010 and 2030. Diabetes Res Clin Pract. 2010;87:4–14. doi: 10.1016/j.diabres.2009.10.007. [DOI] [PubMed] [Google Scholar]
- 3.Brownlee M. Biochemistry and molecular cell biology of diabetic complications. Nature. 2001;414:813–820. doi: 10.1038/414813a. [DOI] [PubMed] [Google Scholar]
- 4.Ghadyale V, Takalikar S, Haldavnekar V, Arvindekar A. Effective control of postprandial glucose level through inhibition of intestinal alpha glucosidase by Cymbopogon martinii (roxb.) Evid Based Complement Alternat Med 2012. 2012:1–6. doi: 10.1155/2012/372909. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Chaisson JL. Acarbose for the prevention of diabetes, hypertension, and cardiovascular disease in subjects with impaired glucose tolerance: the study to prevent non-insulin-dependent diabetes mellitus (STOP-NIDDM) Endocr Pract. 2006;12:25–30. doi: 10.4158/EP.12.S1.25. [DOI] [PubMed] [Google Scholar]
- 6.Giannoukakis N. Ranirestat as a therapeutic aldose reductase inhibitor for diabetic complications. Expert Opin Investig Drugs. 2008;17:575–581. doi: 10.1517/13543784.17.4.575. [DOI] [PubMed] [Google Scholar]
- 7.Fujisawa T, Ikegami H, Inoue K, Kawabata Y, Ogihara T. Effect of two α-glucosidase inhibitors, voglibose and acarbose, on postprandial hyperglycemia correlates with subjective abdominal symptoms. Metabolism. 2005;54:387–390. doi: 10.1016/j.metabol.2004.10.004. [DOI] [PubMed] [Google Scholar]
- 8.Singh SK, Rai PK, Jaiswal D, Watal G. Evidence based critical evaluation of glycemic potential of Cynodon dactylon. Evid Based Complement Alternat Med. 2008;5:415–420. doi: 10.1093/ecam/nem044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Zhang ZS, Ho WKK, Huang Y, Chen ZY. Hypocholesterolemic activity of hawthorn fruit is mediated by regulation of cholesterol-7α-hydroxylase and acyl CoA: cholesterol acyltransferase. Food Res Int. 2002;35:885–891. [Google Scholar]
- 10.Chu CY, Lee MJ, Liao CL, Lin WL, Yin YF, Tseng TH. Inhibitory effect of hot-water extract from dried fruit of Crataegus pinnatifida on low-density lipoprotein (LDL) oxidation in cell and cell-free systems. J Agric Food Chem. 2003;51:7583–7588. doi: 10.1021/jf030407y. [DOI] [PubMed] [Google Scholar]
- 11.Chen JD, Wu YZ, Tao ZL, Chen ZM, Liu XP. Hawthorn (shan zha) drink and its lowering effect on blood lipid levels in humans and rats. World Rev Nutr Diet. 1995;77:147–154. doi: 10.1159/000424470. [DOI] [PubMed] [Google Scholar]
- 12.Kao ES, Wang CJ, Lin WL, Yin YF, Wang CP, Tseng TH. Anti-inflammatory potential of flavonoid content from dried fruits of Crataegus pinnatifida in vitro and in vivo. J Agric Food Chem. 2005;53:430–436. doi: 10.1021/jf040231f. [DOI] [PubMed] [Google Scholar]
- 13.Kao ES, Wang CJ, Lin WL, Chu CY, Tseng TH. Effects of polyphenols derived from fruits of Crataegus pinnatifida on cell transformation, dermal edema and skin tumor formation by phorbol ester application. Food Chem Toxicol. 2007;45:795–804. doi: 10.1016/j.fct.2007.03.016. [DOI] [PubMed] [Google Scholar]
- 14.Li C, Son HJ, Huang C, Lee SK, Lohakare J, Wang MH. Comparison of Crataegus pinnatifida Bunge var typica Schneider and C pinnatifida Bunge fruits for antioxidant, anti-α-glucosidase, and anti-inflammatory activities. Food Sci Biotechnol. 2010;19:769–775. [Google Scholar]
- 15.Li H, Song F, Xing J, Rong T, Liu Z, Liu S. Screening and structural characterization of α-glucosidase inhibitors from Hawthorn leaf flavonoids extract by ultrafiltration LC-DAD-MSn and SORI-CID FTICR MS. J Am Soc Mass Spectrom. 2009;20:1496–1503. doi: 10.1016/j.jasms.2009.04.003. [DOI] [PubMed] [Google Scholar]
- 16.Kuoa DH, Yehb CH, Shieha PC, Chengc KC, Chena FA, Cheng JT. Effect of ShanZha, a Chinese herbal product, on obesity and dyslipidemia in hamsters receiving high-fat diet. J Ethnopharmacol. 2009;124:544–550. doi: 10.1016/j.jep.2009.05.005. [DOI] [PubMed] [Google Scholar]
- 17.Kwok CY, Wong CNY, Yau MYC, Yua PHF, Au ALS, Poon CCW, et al. Consumption of dried fruit of Crataegus pinnatifida (hawthorn) suppresses high-cholesterol diet-induced hypercholesterolemia in rats. J Functional Foods. 2010;2:179–186. [Google Scholar]
- 18.Lin Y, Vermeer MA, Trautwein EA. Triterpenic acids present in hawthorn by inhibiting intestinal ACAT activity in hamsters. Evid Based Complement Alternat Med 2009. 2011:1–9. doi: 10.1093/ecam/nep007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Li T, Zhang XD, Song YW, Liu JW. A microplate-based screening method for α-glucosidase inhibitors. Chin J Clin Pharmacol Ther. 2005;10:1128–1134. [Google Scholar]
- 20.Cui L, Na M, Oh H, Bae EY, Jeong DG, Ryu SE, et al. Protein tyrosine phosphatase 1B inhibitors from Morus root bark. Bioorg Med Chem Lett. 2006;16:1426–1429. doi: 10.1016/j.bmcl.2005.11.071. [DOI] [PubMed] [Google Scholar]
- 21.Hayman S, Kinoshita JH. Isolation and properties of lens aldose reductase. J Biol Chem. 1965;240:877–882. [PubMed] [Google Scholar]
- 22.Vinson JA, Howard TB., III Inhibition of protein glycation and advanced glycation endproducts by ascorbic acid and other vitamins and nutrients. J Nutr Biochem. 1996;7:659–663. [Google Scholar]
- 23.Bilous R, Donelly R. 4th ed. Oxford: Wiley-Blackwell; 2010. Handbook of diabetes; p. 238. [Google Scholar]
- 24.Tonks NK. PTP1B: from the sidelines to the front lines! FEBS Lett. 2003;546:140–148. doi: 10.1016/s0014-5793(03)00603-3. [DOI] [PubMed] [Google Scholar]
- 25.Kusari J, Kenner KA, Shu KI, Hill DE, Henry RR. Skeletal muscle protein tyrosine phosphatase activity and tyrosine phosphatase 1B protein content are associated with insulin action and resistance. J Clin Invest. 1994;93:1156–1162. doi: 10.1172/JCI117068. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ahmad F, Azevedo JJ, Cortright R, Dohm G, Goldstein B. Alteration in skeletal muscle protein-tyrosine phosphatase activity and expression in insulin-resistant human obesity and diabetes. J Clin Invest. 1997;100:449–458. doi: 10.1172/JCI119552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kinoshita JH, Varma SD, Fukui HN. Aldose reductase in diabetes. Jpn J Ophthalmol. 1976;20:399–410. [Google Scholar]
- 28.Kubo E, Miyoshi N, Fukuda M, Akagi Y. Cataract formation through the polyol pathway is associated with free radical production. Exp Eye Res. 1999;68:457–464. doi: 10.1006/exer.1998.0624. [DOI] [PubMed] [Google Scholar]
- 29.Kador PF, Kinoshita JH, Tung WH, Chylack LT. Differences in the susceptibility of various aldose reductases to inhibition. II. Invest Ophthalmol Vis Sci. 1980;19:980–982. [PubMed] [Google Scholar]
- 30.Reddy VP, Beyaz A. Inhibitors of the Maillard reaction and AGE breakers as therapeutics for multiple diseases. Drug Discov Today. 2006;11:646–654. doi: 10.1016/j.drudis.2006.05.016. [DOI] [PubMed] [Google Scholar]
- 31.Thorpe SR, Baynes JW. Maillard reaction products in tissue proteins: new products and new perspectives. Amino Acids. 2003;25:275–281. doi: 10.1007/s00726-003-0017-9. [DOI] [PubMed] [Google Scholar]
- 32.Ahmed N. Advanced glycation endproducts-role in pathology of diabetic complications. Diabetes Res Clin Pract. 2005;67:3–21. doi: 10.1016/j.diabres.2004.09.004. [DOI] [PubMed] [Google Scholar]
- 33.Thomas MC, Baynes JW, Thorpe SR, Cooper ME. The role of AGEs and AGE inhibitors in diabetic cardiovascular disease. Curr Drug Targets. 2005;6:453–474. doi: 10.2174/1389450054021873. [DOI] [PubMed] [Google Scholar]
- 34.Wada R, Yagihashi S. Role of advanced glycation end products and their receptors in development of diabetic neuropathy. Ann N Y Acad Sci. 2005;1043:598–604. doi: 10.1196/annals.1338.067. [DOI] [PubMed] [Google Scholar]
- 35.Muthusamy VS, Saravanababu C, Ramanathan M, Raja RB, Sudhagar S, Anand S, et al. Inhibition of protein tyrosine phosphatase 1B and regulation of insulin signalling markers by caffeoyl derivatives of chicory (Cichorium intybus) salad leaves. Br J Nutr. 2010;104:813–823. doi: 10.1017/S0007114510001480. [DOI] [PubMed] [Google Scholar]
- 36.Kim J, Jeong IH, Kim CS, Lee YM, Kim JM, Kim JS. Chlorogenic acid inhibits the formation of advanced glycation end products and associated protein cross-linking. Arch Pharm Res. 2011;34:495–500. doi: 10.1007/s12272-011-0319-5. [DOI] [PubMed] [Google Scholar]
- 37.Kim CS, Kim J, Lee YM, Sohn E, Jo K, Kim JS. Inhibitory effects of chlorogenic acid on aldose reductase activity in vitro and cataractogenesis in galactose-fed rats. Arch Pharm Res. 2011;34:847–852. doi: 10.1007/s12272-011-0519-z. [DOI] [PubMed] [Google Scholar]
- 38.Lee J, Jang DS, Yoo NH, Lee YM, Kim JH, Kim JS. Single-step separation of bioactive flavonol glucosides from Osteomeles schwerinae by high-speed counter-current chromatography. J Sep Sci. 2010;33:582–586. doi: 10.1002/jssc.200900693. [DOI] [PubMed] [Google Scholar]
- 39.Beaulieu LP, Harris CS, Saleem A, Cuerrier A, Haddad PS, Martineau LC, et al. Inhibitory effect of the Cree traditional medicine wiishichimanaanh (Vaccinium vitis-idaea) on advanced glycation endproduct formation: identification of active principles. Phytother Res. 2010;24:741–747. doi: 10.1002/ptr.3025. [DOI] [PubMed] [Google Scholar]
- 40.Fan P, Terrier L, Hay AE, Marston A, Hostettmann K. Antioxidant and enzyme inhibition activities and chemical profiles of Polygonum sachalinensis F. Schmidt ex Maxim (Polygonaceae) Fitoterapia. 2010;81:124–131. doi: 10.1016/j.fitote.2009.08.019. [DOI] [PubMed] [Google Scholar]
- 41.Kang W, Zhang L, Song Y. α-Glucosidase inhibitors from Luculia pinciana. Zhongguo Zhong Yao Za Zhi. 2009;34:406–409. [PubMed] [Google Scholar]
- 42.Thuong PT, Lee CH, Dao TT, Nguyen PH, Kim WG, Lee SJ, et al. Triterpenoids from the leaves of Diospyros kaki (Persimmon) and their inhibitory effects on protein tyrosine phosphatase 1B. J Nat Prod. 2008;71:1775–1778. doi: 10.1021/np800298w. [DOI] [PubMed] [Google Scholar]
- 43.Na M, Yang S, He L, Oh H, Kim BS, Oh WK, et al. Inhibition of protein tyrosine phosphatase 1B by ursane-type triterpenes isolated from Symplocos paniculata. Planta Med. 2006;72:261–263. doi: 10.1055/s-2005-873194. [DOI] [PubMed] [Google Scholar]
- 44.Yin MC, Chan KC. Non-enzymatic antioxidative and antiglycative effects of oleanolic acid and ursolic acid. J Agric Food Chem. 2007;55:7177–7181. doi: 10.1021/jf071242m. [DOI] [PubMed] [Google Scholar]
- 45.Wang ZH, Hsu C, Huang CC, Yin MC. Anti-glycative effects of oleanolic acid and ursolic acid in kidney of diabetic mice. Eur J Pharmacol. 2010;628:255–260. doi: 10.1016/j.ejphar.2009.11.019. [DOI] [PubMed] [Google Scholar]