

Abstract
AIM: To evaluate the influence of E2F-1 on the growth of human gastric cancer (GC) cells in vivo and the mechanism involved.
METHODS: E2F-1 recombinant lentiviral vectors were injected into xenograft tumors of MGC-803 cells in nude mice, and then tumor growth was investigated. Overexpression of transcription factor E2F-1 was assessed by reverse transcription-polymerase chain reaction (RT-PCR) and Western blotting analysis. Apoptosis rates were determined using a terminal deoxynucleotidyl transferase-mediated dUTP-biotin nick end labeling (TUNEL) assay. Expression levels of certain cell cycle regulators and apoptosis-related proteins, such as Bax, survivin, Bcl-2, cyclin D1, S-phase kinase-associated protein 2, and c-Myc were examined by Western blotting and RT-PCR.
RESULTS: Xenograft tumors of MGC-803 cells in nude mice injected with E2F-1 recombinant lentiviral vectors stably overexpressed the E2F-1 gene as measured by semi-quantitative RT-PCR (relative mRNA expression: 0.10 ± 0.02 vs 0.05 ± 0.02 for control vector and 0.06 ± 0.03 for no infection; both P < 0.01) and Western blotting (relative protein expression: 1.90 ± 0.05 vs 1.10 ± 0.03 in control vector infected and 1.11 ± 0.02 for no infection; both P < 0.01). The growth-curve of tumor volumes revealed that infection with E2F-1 recombinant lentiviral vectors significantly inhibited the growth of human GC xenografts (2.81 ± 1.02 vs 6.18 ± 1.15 in control vector infected and 5.87 ± 1.23 with no infection; both P < 0.05) at 15 d after treatment. TUNEL analysis demonstrated that E2F-1 overexpression promoted tumor cell apoptosis (18.6% ± 2.3% vs 6.7% ± 1.2% in control vector infected 6.3% ± 1.2% for no infection; both P < 0.05). Furthermore, lentiviral vector-mediated E2F-1 overexpression increased the expression of Bax and suppressed survivin, Bcl-2, cyclin D1, Skp2, and c-Myc expression in tumor tissue.
CONCLUSION: E2F-1 inhibits growth of GC cells via regulating multiple signaling pathways, and may play an important role in targeted therapy for GC.
Keywords: E2F-1, Gastric cancer, Lentiviral vector, Mouse model
Core tip: Transcription factor E2F-1 is the prototypical E2F and is often implicated in DNA synthesis and repair, cell proliferation, and apoptosis. Our preliminary study revealed that high expression of E2F-1 significantly suppressed gastric cancer (GC) cell line progression in vitro. However, the role of E2F-1 overexpression in GC in vivo remains unknown. Our results showed that overexpression of E2F-1 significantly inhibited tumor growth and promoted tumor cell apoptosis in vivo. Survivin, Bcl-2, cyclin D1, S-phase kinase-associated protein 2 and c-Myc were upregulated, and Bax was downregulated by E2F-1. E2F-1 inhibits growth of GC cells via regulating multiple signaling pathways.
INTRODUCTION
Although the incidence rate of gastric cancer (GC) has gradually decreased recently, it remains the second leading cause of cancer-related death worldwide[1]. Of all GC cases, > 70% occur in developing countries and half of the total cases worldwide occur in Eastern Asia (mainly China)[2]. Despite improvements in surgical techniques and the development of new chemotherapeutic regimens, patient outcome is often disappointing. Patients are mostly diagnosed at advanced stages, typically with a poor prognosis, with a five-year survival rate of < 30%[3]. Because of the patient’s own reasons, they lose the opportunity to receive surgery and chemotherapy[4]. Thus, it is necessary to find new therapies. Gene therapy as a novel strategy has been shown to have a therapeutic advantage for treating several types of tumors, including gastric carcinoma, and promises to be a new therapeutic approach to inhibit the proliferation of tumor cells, and avoids the side effects of drug therapy[5,6].
Since the E2F family factors have been reported, they have been considered as main regulators of cell growth and proliferation[7]. E2F-1 gene is one member of the E2F family, with the ability to induce apoptosis independently[8]. E2F-1 is also a key regulator for the G1/S phase transition[9]. On the one hand, a number of researchers have shown that high expression of E2F-1 is a risk factor for malignant tumors[10,11]. On the other hand, E2F-1 overexpression may play an important role in suppressing tumor growth in lung cancer, breast cancer and osteosarcoma[12-14]. These findings indicate that the E2F-1 gene has a dual effect in promoting cell proliferation and apoptosis. However, few studies have been reported concerning E2F-1 expression in GC. In particular, the functional mechanism of E2F-1 overexpression has not been determined. Our previous study indicated that E2F-1 overexpression had a significant influence on cell cycle progression and proliferation in an in vitro GC cell model[15,16], but the molecular mechanisms underlying inhibition of cell growth and increase of apoptosis by E2F-1 overexpression remain unclear.
It is widely known that undifferentiated cells and differentiated cells can be efficiently infected with lentivirus, and lentivirus-carrying genes are stably integrated into the host genome[17,18]. Therefore, transfer of recombinant lentiviral vectors is the best transgene method in various animals. Accordingly, we constructed E2F-1 recombinant lentiviral vectors and evaluated the influence of E2F-1 overexpression on the biologic behavior of MGC-803 cells in vivo using a xenograft tumor model. To explore the potential mechanism, we also examined the influence of E2F-1 overexpression on the expression of survivin, Bax, Bcl-2, cyclin D1, S-phase kinase-associated protein (Skp)2, and c-Myc in MGC-803 cells in vivo.
MATERIALS AND METHODS
Cell culture
The human GC MGC-803 cells and human kidney 293T cells were purchased from the Cell Bank of the Chinese Academy of Sciences (Shanghai, China). The cells were incubated in Dulbecco’s modified eagle medium (HyClone of Thermo Fisher Scientific Inc., Waltham, MA, United States), supplemented with 2 mmol/L glutamine, 0.05 g/L penicillin, 0.1 g/L streptomycin, and 10% fetal bovine serum at 37 °C in a humidified atmosphere containing 5% CO2. Culture media were replaced once every two days.
Antibodies
Specific rabbit anti-human antibodies to E2F-1, c-Myc, Skp2, Bax, Bcl-2, cyclin D1, survivin and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were obtained from Abcam (Cambridge, United Kingdom). Infrared-labeled secondary goat anti-rabbit antibodies to IRDye 800 were obtained from Li-Cor Biosciences (Lincoln, NE, United States).
Construction of E2F-1 recombinant lentiviral vectors
Lentiviral vectors with green fluorescent protein (GFP) were provided by Genechem (Shanghai, China). The lentiviral vector system was comprised of the vectors pGCL-GFP, pHelper 1.0 and pHelper 2.0. The full-length human E2F-1 gene (NM_0005225.2) was encoded by the pGCL-GFP-E2F-1 plasmid. The E2F-1 cDNA was accurately inserted into the plasmid, which was verified by DNA sequencing and PCR technology. The three plasmids (pHelper 1.0, pHelper 2.0, and pGCL-GFP or pGCL-GFP-E2F-1) were co-transfected into 293T cells using Lipofectamine 2000[19]. After a 12-h transfection, the medium was replaced with fresh medium supplemented with 10% fetal bovine serum. The lentivirus containing the E2F-1 gene was collected at 48 h after the transfection. The product was concentrated by ultracentrifugation (12000× g) and diluted in a series of gradients from 10-2 to 10-5. Various dilutions of lentivirus were used to transfect 293T cells. Then the cells were observed under a fluorescence microscope after 72 h. The cells with GFP expression were positive cells, and the number of positive cells was counted in each transfection. The lentiviral titer was calculated by the formula: lentiviral titer (TU/mL) = number of positive cells × dilution times/volume of lentivirus used.
Xenograft tumor model
Athymic nude male BALBC/c mice, aged 4-5 wk, were purchased from Guangxi Animal Center (Nanning, China). The mice were maintained in specific pathogen-free, temperature-controlled isolation conditions and fed with sterilized food and autoclaved water according to the experimental animal guidelines. All animal procedures were conducted to follow the provisions of the Ethics Committee of Guangxi Medical University in the research. Xenografted tumor models were prepared by subcutaneous injection of 3 × 106 MGC-803 cells suspended in phosphate-buffered saline (PBS) into male athymic nude mice. The xenografted tumor was referred to as a human GC xenograft.
Treatment of human GC xenografts in nude mice
When tumors reached a diameter of approximately 6 mm, mice were randomized into three groups (n = 8 each): LV-GFP-E2F-1, LV-GFP-NC, and PBS. The tumors of nude mice were injected with LV-GFP-E2F-1 or LV-GFP-NC at a concentration of 5 × 106 TU in 200 μL PBS, while the control group received an equal volume of PBS. The tumors of animals were injected in each group once every 2 d. Animals and tumors were observed daily. Tumor size was monitored every 2 d with a digital caliper and tumor volume (TV) was estimated by the equation: TV = a × b2/2, where a is the longest diameter and b the shortest diameter. The relative tumor volume (RTV) was calculated by the formula: RTV = Vt/V0 (V0 was the initial TV of the first day of treatment and Vt was the following TV measured)[20]. After a total of 15 d of treatment, nude mice were suffocated and tumors were assessed.
Semi-quantitative reverse transcriptase-PCR
Total RNA was extracted from MGC-803 tumor tissues and GC xenografts with TRI Reagent (Sigma-Aldrich, St Louis, MO, United States). cDNA was generated from DNase-1-treated RNA template with 0.2 μg random hexamer primers and 200 U RevertAid H-Minus M-MuLV reverse transcriptase enzyme (Roche, Switzerland). All the PCR primer sequences used in this study, including for c-Myc, Skp2, Bax, Bcl-2, cyclin D1, surviving and GAPDH are shown in Table 1. The PCR products were used for electrophoresis in 1.5% agarose gel stained with ethidium bromide (0.5 μg/mL). The mRNA expression levels of each gene were measured using GAPDH mRNA as a reference. Reverse transcriptase (RT)-PCR product bands were analyzed by densitometry using Quantity One 1-D gel image analysis software (BioRad, Hercules, CA, United States).
Table 1.
Sequences of primers used for semi-quantitative real-time polymerase chain reaction
| Gene | Primer | Base sequence | PCR product (bp) |
| E2F-1 | Forward | 5'-CCCAACTCCCTCTACCCT-3' | 217 |
| Reverse | 5'-CTCCCATCTCATATCCATCCTG-3' | ||
| Survivin | Forward | 5'-AAATGCACTCCAGCCTCTGT-3' | 311 |
| Reverse | 5'-TGTCGAGGAAGCTTTCAGGT-3' | ||
| Bax | Forward | 5'-CCAAGAAGCTGAGCGAGTGT-3' | 269 |
| Reverse | 5'-CCGGAGGAAGTCCAATGTC-3' | ||
| Bcl-2 | Forward | 5'-GACTTCGCCGAGATGTCCAG-3' | 259 |
| Reverse | 5'-CATCCCAGCCTCCGTTATCC-3' | ||
| Cyclin D1 | Forward | 5'-CCCTCGGTGTCCTACTTCAA-3' | 237 |
| Reverse | 5'-GGGGATGGTCTCCTTCATCT-3' | ||
| Skp2 | Forward | 5'-GCTGCTAAAGGTCTCTGGTGT-3' | 291 |
| Reverse | 5'-AGGCTTAGATTCTGCAACTTG-3' | ||
| c-Myc | Forward | 5'-TTCTCTCCGTCCTCGGATTC-3' | 282 |
| Reverse | 5'-GTAGTTGTGCTGATGTGTGG-3' | ||
| GAPDH | Forward | 5'-ACCACAGTCCATGCCATCAC-3' | 450 |
| Reverse | 5'-TCACCACCCTGTTGCTGTA-3' |
GAPDH: Glyceraldehyde-3-phosphate dehydrogenase; PCR: Polymerase chain reaction; Skp2: S-phase kinase-associated protein 2.
Western blot analysis
Protein was extracted in accordance with the kit instructions (OriGene, Rockville, MD, United States) in tumor tissue. Proteins were separated by 10% polyacrylamide-gel electrophoresis and then transferred onto polyvinylidene fluoride membranes (Roche) at 100 mA for 3 h, and later soaked for 2 h in a blocking solution (Tris-buffered saline containing 5% nonfat dry milk and 0.1% Tween-20), and immersed in a 1:1000 anti-E2F-1 antibody diluent, or anti-GAPDH monoclonal antibody used as an internal control, then incubated at 4 °C overnight. The membrane was washed with PBS with Tween 20 and immersed in a 1:10000 infrared-labeled secondary goat anti-rabbit antibody diluent for 30 min. The membranes were analyzed by densitometry using Odyssey version 3.0 (Li-Cor). Then the protein expression levels of each gene were measured using GAPDH protein as a reference.
Analyses of GC xenograft tissue apoptosis by terminal deoxynucleotidyl transferase-mediated dUTP-biotin nick end labeling
Paraffin sections of GC xenografts were processed with a terminal deoxynucleotidyl transferase-mediated dUTP-biotin nick end labeling (TUNEL) assay kit (Roche) treated according to the manufacturer’s instructions. Gastric tumor cell apoptosis was observed under microscopy. The primary antibody was replaced by phosphate buffer on the positive specimens in the negative controls. The brown particles in the nucleus were considered as apoptosis-positive cells. Eight fields were randomly selected in each slice under a high-power field (×400). The percentage of TUNEL-positive cells was calculated from a total of 200 cells in each field.
Statistical analysis
Data analyses were performed using SPSS version 16.0 (SPSS Inc., Chicago, IL, United States). Numerical data are reported as the mean ± SD. Group comparisons were evaluated by one-way analysis of variance. Two-sided tests were used to evaluate comparisons. Data were determined to have significant differences when P < 0.05.
RESULTS
Evaluation of pGCL-GFP-E2F-1 lentiviral vector
Positive clones were screened by restriction enzyme analysis, and were verified by DNA sequencing (data not displayed). Positive clone sequencing was consistent with the target sequence of E2F-1 reported in Genebank indicating that the pGCL-GFP-E2F-1 had been constructed successfully.
Determination of lentiviral titers
As displayed in Figure 1, lentiviral vectors of LV-GFP-E2F-1 or LV-GFP-NC were mixed into 293T cell culture medium, and more than 90% of the cells were transfected. The concentration of the virus was greater than 6 × 108 TU/mL by GFP expression assay.
Figure 1.
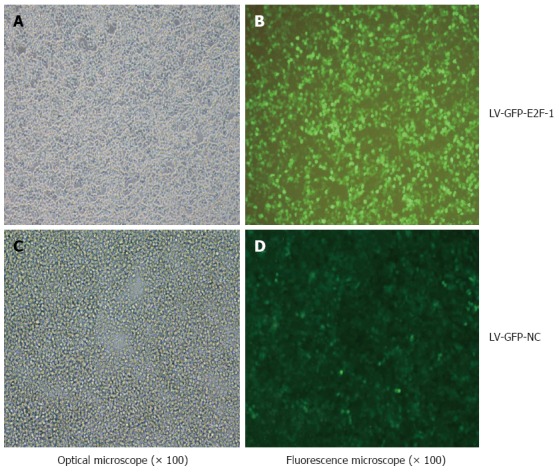
Determination of lentiviral titers. Recombinant lentivirus was transfected into 293T cells to determine viral titer by the method of end-point dilution, which involves counting the numbers of infected green cells under fluorescence microscopy (original magnification x 100). The viral dilution factor was 1:1000.
Overexpression of E2F-1 inhibited GC xenograft tumor growth
The tumors of the GC xenograft model were injected with E2F-1 recombinant lentiviral vectors. Compared with the LV-GFP-NC and PBS groups, the growth-curve of tumor volumes (Figure 2A) revealed that the growth of xenografts infected with E2F-1 recombinant lentiviral vectors had was significantly inhibited (P < 0.05). After a total of 15 d of treatment, GC xenograft tumor volume in the LV-GFP-E2F-1 group was significantly smaller than in the LV-GFP-NC and PBS groups (both P < 0.05), and there was no statistically significant difference between LV-GFP-NC and PBS groups (Figure 2B). The results indicated that overexpression of E2F-1 could effectively suppress gastric cancer xenograft tumor growth in vivo.
Figure 2.
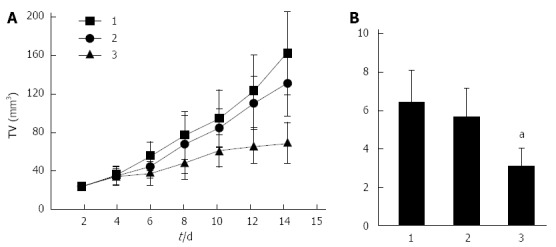
Overexpression of E2F-1 inhibits MGC-803 tumor growth. A: Tumor growth curve shows the growth tendency in the LV-GFP-E2F-1 group is suppressed in comparison to the control groups; B: Relative tumor volume (TV) in LV-GFP-E2F-1-treated mice was significantly smaller than in the control groups at 12 d after tumor injection (n = 8 animals for each condition). 1, phosphate-buffered saline (PBS) control group; 2, LV-GFP-NC group; 3, LV-GFP-E2F-1 group; comparisons made using analysis of variance and Student-Newman-Keuls analyses; aP < 0.05 vs control.
Protein and mRNA of E2F-1 gene were highly expressed in GC xenograft tissue
Compared with the LV-GFP-NC and PBS groups, mRNA and protein expressions of E2F-1 gene were significantly higher in the LV-E2F-1-GFP group by gray value analysis (P < 0.05), and there was no statistically significant difference between LV-GFP-NC and PBS groups (Figures 3 and 4). The results suggest that a nude mouse model with overexpression of E2F-1 was constructed successfully by injecting with E2F-1 recombinant lentiviral vectors.
Figure 3.
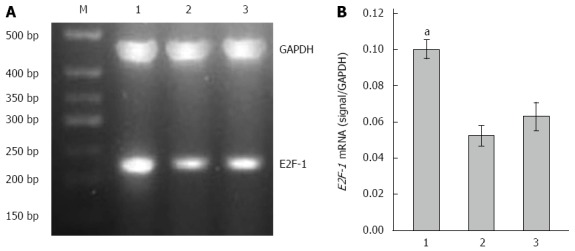
Overexpression of E2F-1 mRNA with LV-GFP-E2F-1. A: Agarose gel analysis of E2F-1 and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) real-time polymerase chain reaction products amplified from MGC-803 tumor tissues (M, 500 bp marker); B: Expression of E2F-1 mRNA was measured in the three groups and normalized to GAPDH (n = 8 animals for each condition). 1, LV-GFP-E2F-1 group; 2, LV-GFP-NC group; 3, phosphate buffered saline control group. aP < 0.05 using analysis of variance and Student-Newman-Keuls analyses.
Figure 4.
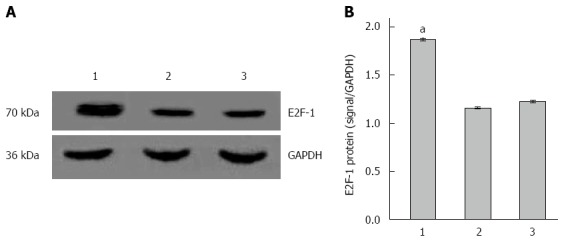
Overexpression of E2F-1 protein with LV-GFP-E2F-1. A: Western blot analysis of E2F-1 and glyceraldehyde-3-phosphate dehydrogenase (GAPDH; internal control) in MGC-803 tumor tissue; B: Expression of E2F-1 protein was measured in the three groups normalized to GAPDH (n = 8 animals for each condition). 1, LV-GFP-E2F-1 group; 2, LV-GFP-NC group; 3, PBS group. aP < 0.05 using analysis of variance and Student-Newman-Keuls analyses.
Overexpression of E2F-1 induced GC xenograft tissue cell apoptosis
Cell apoptosis was detected by the TUNEL staining in the GC xenograft tissue (Figure 5). The rate of cell apoptosis in the LV-GFP-NC, LV-GFP-NC and PBS groups were 18.6% ± 2.3%, 6.7% ± 1.2% and 6.3% ± 1.2%, respectively. Compared with LV-GFP-NC and PBS groups, the apoptosis rate of the LV-E2F-1-GFP group was significantly higher (P < 0.05). The results suggest that overexpression of E2F-1 effectively promotes GC xenograft tumor cell apoptosis in vivo.
Figure 5.
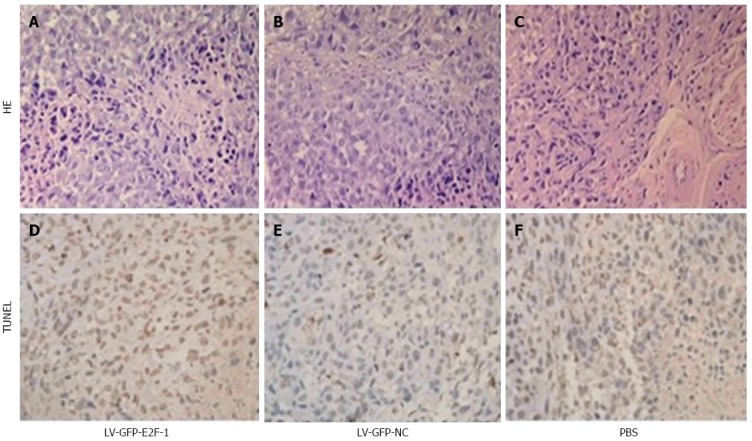
Overexpression of E2F-1 induces in situ MGC-803 tumor cell apoptosis. Apoptosis was assessed by terminal deoxynucleotidyl transferase-mediated dUTP-biotin nick end labeling (TUNEL) and hematoxylin and eosin staining. More apoptotic MGC-803 tumor cells were observed in the LV-GFP-E2F-1 group than in the LV-GFP-NC and phosphate buffered saline (PBS) groups (original magnification, × 400).
Overexpression of E2F-1 decreases expression of c-Myc, Skp2, Bcl-2, cyclin D1and survivin, and increases Bax expression
To probe the principles of E2F-1-induced cell apoptosis in GC xenografts, semiquantitative RT-PCR and Western blot were used to detect expression of apoptosis-related genes. Compared with the LV-GFP-NC and PBS groups, densitometry displayed that the mRNA levels of c-Myc, Skp2, Bcl-2, cyclin D1 and survivin were lower in the LV-GFP-E2F-1 group, whereas Bax levels were higher in the LV-GFP-E2F-1 group (all Ps < 0.05), and there was no statistically significant difference between LV-GFP-NC and PBS groups (Figure 6). Compared with LV-GFP-NC and PBS groups, densitometry showed that the protein levels of c-Myc, Skp2, Bcl-2, cyclin D1 and survivin were significantly lower, and the levels of Bax was significantly higher, in the LV-GFP-E2F-1 group (all P < 0.05), and there was no statistically significant difference between LV-GFP-NC and PBS groups (Figure 7). The results suggest that overexpression of E2F-1 decreases expression of c-Myc, Skp2, Bcl-2, cyclin D1 and survivin and increases Bax expression in MGC-803 tumors in vivo.
Figure 6.
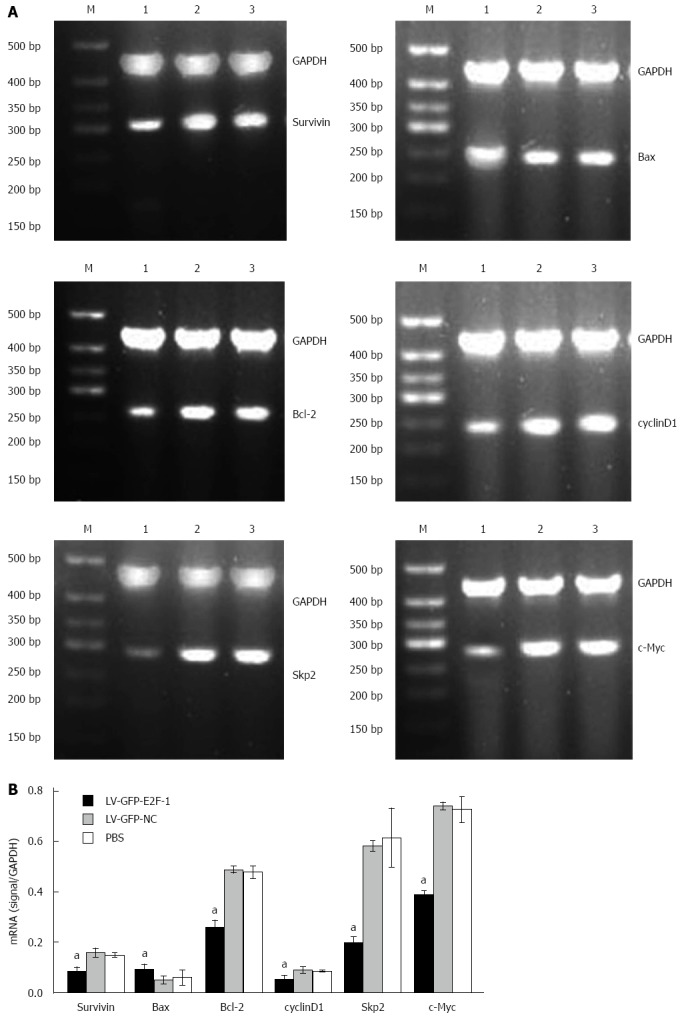
Overexpression of E2F-1 alters mRNA expression of apoptosis regulators. A: Agarose gel analysis of semiquantitative real-time polymerase chain reactions for c-Myc, S-phase kinase-associated protein 2 (Skp2), Bcl-2, cyclin D1, survivin, Bax and glyceraldehydes-3-phosphate dehydrogenase (GAPDH) in the MGC-803 tumor tissues. Lanes: 1, LV-GFP-E2F-1 group; 2, LV-GFP-NC group; 3, phosphate-buffered saline (PBS) group; M, 500 bp marker; B: Quantification showing downregulation of c-Myc, Skp2, Bcl-2, cyclin D1, survivin and upregulation of Bax mRNA levels (n = 8 animals for each condition). aP < 0.05 using analysis of variance and Student-Newman-Keuls analyses.
Figure 7.
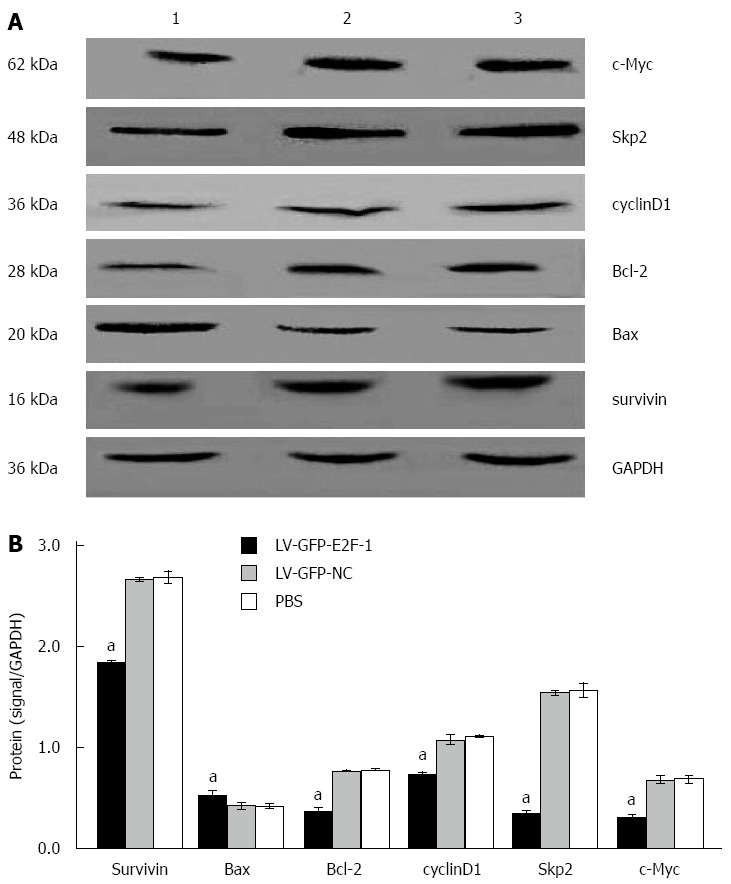
Overexpression of E2F-1 alters protein expression of apoptosis regulators. A: Western blot analysis of c-Myc, S-phase kinase-associated protein 2 (Skp2), Bcl-2, cyclin D1, survivin, Bax and glyceraldehyde-3-phosphate dehydrogenase (GAPDH; internal control) in the MGC-803 tumor tissues. Lanes: 1, LV-GFP-E2F-1 group; 2, LV-GFP-NC group; 3, phosphate-buffered saline (PBS) group; B: Quantification showing downregulation c-Myc, Skp2, Bcl-2, cyclin D1, survivin and upregulation of Bax protein levels (n = 8 animals for each condition). aP < 0.05 using analysis of variance and Student-Newman-Keuls analysis.
DISCUSSION
Cancer is a disease with dysregulation of the cell cycle and uncontrolled growth due to the combined effects of hereditary and environmental factors. The mechanism of cell cycle dysregulation is an important cause of the cell proliferation dropout that leads to cancer. Normally, each period of cell division and proliferation are strictly regulated by a variety of specific proteins.
The E2Fs are one of the most important cell cycle regulatory protein families[21]. Cell survival, mitosis, and apoptosis can be regulated by E2F family proteins[22]. E2F-1 is the most representative factor of this family, and it is generally considered to have the ability to induce apoptosis[23]. Many studies have shown that E2F-1 has anti-tumor effects in tumors such as colonic adenocarcinoma, glioma, and breast carcinoma[24-26]. However, E2F-1 also enhanced DNA replication and G1-to-S phase transition[27]. According to Conner et al[28], E2F-1 overexpression in the liver causes dysplasia and tumors. E2F-1 overexpression also enhances cell proliferation in epithelial ovarian cancer and may provide a useful prognostic indicator[29]. Expression of E2F-1 has been found in many human malignancies where it has two contrasting functions, including both inhibiting and promoting tumor growth. In addition, our previous studies have shown that downregulation of E2F-1 using RNAi successfully inhibits GC cell proliferation in vivo and in vitro[30,31]. It seems that E2F-1 is pleiotropic, and its functions are based on the dominant signaling pathway and cancer cell type.
In this study, MGC-803 cells were successfully inhibited by E2F-1 overexpression in vivo. This is consistent with our previous studies in vitro[15,16]. Therefore, lentiviral vector-mediated E2F-1 overexpression can be a new and effective treatment of GC. In this study, E2F-1 overexpression also decreased survivin, Bcl-2, cyclin D1, Skp2, and c-Myc expression, while Bax expression was increased. Thus, the E2F-1 gene might affect GC growth by modifying signaling pathways involving these genes.
The activation of mitochondrion-dependent processes is an effective approach for inducing apoptosis, which is regulated by the Bcl-2 family member. However, the proteins Bax and Bcl-2 have opposing roles in initiating mitochondrial apoptotic events and modulating apoptosis. Bcl-2 is associated with the outer mitochondrial membrane, where it plays a pivotal and important part in protecting mitochondrial structure and action, and prevents enzyme proteins from being released into the cytosol, thereby maintaining a stable intracellular environment[32]. In contrast, Bax, a dominant-negative inhibitor of Bcl-2, induces a mitochondrial permeability transition and promotes apoptosis. In this study, E2F-1 upregulated Bax and downregulated antiapoptotic Bcl-2, thereby elevating the ratio of Bax/Bcl-2 in MGC-803 cells. This suggests that cell apoptosis can be induced by a mitochondrion-dependent pathway, thereby activating caspase-9 and caspase-3[33]. Our current results are in good agreement with other recent studies, which demonstrated that E2F-1 induces apoptosis with Bax upregulation or Bcl-2 downregulation in melanoma cells[34] and human esophageal cancer cells[35].
Overexpression of E2F-1 downregulated the expression of c-Myc and Skp2, which are overexpressed in GC and promote tumor growth[36,37]. c-Myc is also involve in cell transcription and mitosis by management of cell cycle-related genes[38-41]. Zhang et al[42] confirmed that overexpression of c-Myc could enhance normal gastric cell growth and proliferation, as GC cell proliferation was inhibited by RNA interference targeting c-Myc gene silencing. Skp2 belongs to the family of F-box proteins. It has been known for a long time that Skp2 undergoes phosphorylation during cell cycle progression and growth factor stimulation[43,44]. Wei et al[45] reported that downregulation of Skp2 inhibited the growth and metastasis of GC cells in vitro and in vivo. Skp2 depletion also increased caspase-3 activity and impeded formation of filopodia and locomotion. Skp2 is associated with c-Myc transcription and degradation, and thus regulates Myc protein stability[46]. This establishes an unusual structured network that is the E2F-1/Skp2/c-Myc signaling pathway, by which E2F-1 suppresses MGC-803 cell growth in vivo.
Cyclin D1 is one of the most important proteins for cell cycle regulation, and is related to the development of many cancers and cancer cell growth. Cyclin D1 binds to and activates CDK4/6, which subsequently phosphorylates tumor-inhibiting protein Rb and promotes G1/S transition in the cell cycle[47,48]. So far, survivin is the strongest anti-apoptotic factor with the smallest molecular weight[49]. Survivin can also prevent tubulin from binding with spindle microtubules, and inhibit apoptosis in cell mitosis[50]. Furthermore, the proportion of cells in the G2/M phase were increased, while the number of cells in the G0/G1 phase decreased after transfecting survivin-specific siRNA into MGC-803 cells[51]. Thus, the proliferation of GC cells was inhibited, and apoptosis induced by survivin RNA interference. Dar et al[52] have shown that the proliferative capacity of melanoma cells is mediated by E2F-regulated Akt phosphorylation. Furthermore, the PTEN-Akt-p53-miR- 365-Cyclin D1/cdc25A axis serves as a new mechanism underlying gastric tumorigenesis[53]. Akt signaling translationally regulates survivin expression for metastatic progression of colorectal cancer and GC[54,55].
Our present study has shown that E2F-1 overexpression significantly inhibited the growth of transplanted tumors and promoted cell apoptosis. E2F-1 overexpression also inhibited cyclin D1 and survivin expression, either directly or indirectly via the Akt signaling pathway, and suppressed MGC-803 cell growth in vivo. Therefore, E2F-1 plays an important role in GC cell survival, apoptosis, and mitosis. This research provides a basis for treatment of GC via regulation of E2F-1 expression.
COMMENTS
Background
Gastric cancer (GC) remains the second leading cause of death from malignant disease worldwide. Despite improvements in surgical techniques and the development of new chemotherapeutic regimens, the results are often disappointing in patients with advanced tumors. Gene therapy is a very hopeful approach to inhibit the proliferation of tumor cells and avoid the side effects of drug therapy. E2F-1, a member of the E2F family of transcription factors, is crucial for the E2F-dependent apoptotic program. However, few studies have reported on E2F-1 in GC. In particular, the effect and functional mechanism of E2F-1 overexpression in gastric cancer in vivo remain unclear.
Research frontiers
E2F-1 plays an important part in manipulating cell cycle progression and other cell biologic behaviors. E2F-1 can show antitumor or tumor-promoting effects, which remain controversial. The research hotspot of E2F-1 is how it affects the progression of human cancer.
Innovations and breakthroughs
Previous studies have indicated that the growth and proliferation of GC cells were inhibited and G1 to S phase cell cycle transition was blocked by E2F-1 overexpression in vitro. Recent reports were focused on its roles on in vivo. The authors discovered that E2F-1 inhibited tumor growth and promoted tumor cell apoptosis in models of human gastric carcinoma in nude mice. In addition, E2F-1 upregulation increased the expression of Bax and decreased expression of survivin, Bcl-2, cyclin D1, Skp2, and c-Myc expression level.
Applications
In understanding the role and mechanism of E2F-1-mediated multiple signaling pathways in GC in vivo, this study is expected to suggest a way to improve clinical treatment.
Terminology
E2F-1 is a transcription factor involved with DNA replication, cell survival, mitosis, and apoptosis. The retinoblastoma 1 (pRb) factor binds to E2F-1, and E2F-1 activation is determined by the state of pRb phosphorylation; E2F-1 is inhibited by dephosphorylated pRb and activated by pRb phosphorylation.
Peer review
This study is a very enjoyable and valuable work. The results may represent a molecular mechanism of E2F-1 in GC.
Footnotes
Supported by National Natural Science Foundation of China, No. 30860273 and No. 81060201; Natural Science Foundation of Guangxi, No. 2011GXNSFA018273 and No. 2013GXNSFAA019163; and the Key Health Science Project of Guangxi, No. Key1298003-2-6.
Open-Access: This article is an open-access article which was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Peer-review started: March 19, 2014
First decision: April 21, 2014
Article in press: July 16, 2014
P- Reviewer: Tanyi M S- Editor: Gou SX L- Editor: AmEditor E- Editor: Ma S
References
- 1.Yasui W, Sentani K, Sakamoto N, Anami K, Naito Y, Oue N. Molecular pathology of gastric cancer: research and practice. Pathol Res Pract. 2011;207:608–612. doi: 10.1016/j.prp.2011.09.006. [DOI] [PubMed] [Google Scholar]
- 2.Ferlay J, Shin HR, Bray F, Forman D, Mathers C, Parkin DM. Estimates of worldwide burden of cancer in 2008: GLOBOCAN 2008. Int J Cancer. 2010;127:2893–2917. doi: 10.1002/ijc.25516. [DOI] [PubMed] [Google Scholar]
- 3.Mlkvý P. Multimodal therapy of gastric cancer. Dig Dis. 2010;28:615–618. doi: 10.1159/000320063. [DOI] [PubMed] [Google Scholar]
- 4.Shulman K, Haim N, Wollner M, Bernstein Z, Abdah-Bortnyak R, Bar-Sela G. Postoperative chemotherapy in gastric cancer, consisting of etoposide, doxorubicin and cisplatin, followed by radiotherapy with concomitant cisplatin: A feasibility study. Oncol Lett. 2012;3:1154–1158. doi: 10.3892/ol.2012.617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Kulig J, Kołodziejczyk P, Kulig P, Legutko J. Targeted therapy for gastric cancer--current status. J Oncol Pharm Pract. 2013;19:75–81. doi: 10.1177/1078155212449030. [DOI] [PubMed] [Google Scholar]
- 6.McNamara MJ, Adelstein DJ. Current developments in the management of locally advanced esophageal cancer. Curr Oncol Rep. 2012;14:342–349. doi: 10.1007/s11912-012-0239-7. [DOI] [PubMed] [Google Scholar]
- 7.DeGregori J, Johnson DG. Distinct and Overlapping Roles for E2F Family Members in Transcription, Proliferation and Apoptosis. Curr Mol Med. 2006;6:739–748. doi: 10.2174/1566524010606070739. [DOI] [PubMed] [Google Scholar]
- 8.Lazzerini Denchi E, Helin K. E2F1 is crucial for E2F-dependent apoptosis. EMBO Rep. 2005;6:661–668. doi: 10.1038/sj.embor.7400452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Hallstrom TC, Nevins JR. Balancing the decision of cell proliferation and cell fate. Cell Cycle. 2009;8:532–535. doi: 10.4161/cc.8.4.7609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Molina-Privado I, Rodríguez-Martínez M, Rebollo P, Martín-Pérez D, Artiga MJ, Menárguez J, Flemington EK, Piris MA, Campanero MR. E2F1 expression is deregulated and plays an oncogenic role in sporadic Burkitt’s lymphoma. Cancer Res. 2009;69:4052–4058. doi: 10.1158/0008-5472.CAN-08-4617. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Gala S, Marreiros A, Stewart GJ, Williamson P. Overexpression of E2F-1 leads to cytokine-independent proliferation and survival in the hematopoietic cell line BaF-B03. Blood. 2001;97:227–234. doi: 10.1182/blood.v97.1.227. [DOI] [PubMed] [Google Scholar]
- 12.Peng B, Cao J, Yi S, Wang C, Zheng G, He Z. Inhibition of proliferation and induction of G1-phase cell-cycle arrest by dFMGEN, a novel genistein derivative, in lung carcinoma A549 cells. Drug Chem Toxicol. 2013;36:196–204. doi: 10.3109/01480545.2012.710620. [DOI] [PubMed] [Google Scholar]
- 13.Shen WH, Jackson ST, Broussard SR, McCusker RH, Strle K, Freund GG, Johnson RW, Dantzer R, Kelley KW. IL-1beta suppresses prolonged Akt activation and expression of E2F-1 and cyclin A in breast cancer cells. J Immunol. 2004;172:7272–7281. doi: 10.4049/jimmunol.172.12.7272. [DOI] [PubMed] [Google Scholar]
- 14.Liontos M, Niforou K, Velimezi G, Vougas K, Evangelou K, Apostolopoulou K, Vrtel R, Damalas A, Kontovazenitis P, Kotsinas A, et al. Modulation of the E2F1-driven cancer cell fate by the DNA damage response machinery and potential novel E2F1 targets in osteosarcomas. Am J Pathol. 2009;175:376–391. doi: 10.2353/ajpath.2009.081160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Xiao Q, Li L, Xie Y, Tan N, Wang C, Xu J, Xia K, Gardner K, Li QQ. Transcription factor E2F-1 is upregulated in human gastric cancer tissues and its overexpression suppresses gastric tumor cell proliferation. Cell Oncol. 2007;29:335–349. doi: 10.1155/2007/147615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Xie Y, Wang C, Li L, Ma Y, Yin Y, Xiao Q. Overexpression of E2F-1 inhibits progression of gastric cancer in vitro. Cell Biol Int. 2009;33:640–649. doi: 10.1016/j.cellbi.2009.02.015. [DOI] [PubMed] [Google Scholar]
- 17.Liu C, Wang L, Li W, Zhang X, Tian Y, Zhang N, He S, Chen T, Huang J, Liu M. Highly efficient generation of transgenic sheep by lentivirus accompanying the alteration of methylation status. PLoS One. 2013;8:e54614. doi: 10.1371/journal.pone.0054614. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Segura MM, Garnier A, Durocher Y, Coelho H, Kamen A. Production of lentiviral vectors by large-scale transient transfection of suspension cultures and affinity chromatography purification. Biotechnol Bioeng. 2007;98:789–799. doi: 10.1002/bit.21467. [DOI] [PubMed] [Google Scholar]
- 19.Hanawa H, Kelly PF, Nathwani AC, Persons DA, Vandergriff JA, Hargrove P, Vanin EF, Nienhuis AW. Comparison of various envelope proteins for their ability to pseudotype lentiviral vectors and transduce primitive hematopoietic cells from human blood. Mol Ther. 2002;5:242–251. doi: 10.1006/mthe.2002.0549. [DOI] [PubMed] [Google Scholar]
- 20.Wang XT, Xie YB, Xiao Q. siRNA targeting of Cdx2 inhibits growth of human gastric cancer MGC-803 cells. World J Gastroenterol. 2012;18:1903–1914. doi: 10.3748/wjg.v18.i16.1903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Tammali R, Saxena A, Srivastava SK, Ramana KV. Aldose reductase regulates vascular smooth muscle cell proliferation by modulating G1/S phase transition of cell cycle. Endocrinology. 2010;151:2140–2150. doi: 10.1210/en.2010-0160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Bao J, Li D, Wang L, Wu J, Hu Y, Wang Z, Chen Y, Cao X, Jiang C, Yan W, et al. MicroRNA-449 and microRNA-34b/c function redundantly in murine testes by targeting E2F transcription factor-retinoblastoma protein (E2F-pRb) pathway. J Biol Chem. 2012;287:21686–21698. doi: 10.1074/jbc.M111.328054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Wu Z, Zheng S, Yu Q. The E2F family and the role of E2F1 in apoptosis. Int J Biochem Cell Biol. 2009;41:2389–2397. doi: 10.1016/j.biocel.2009.06.004. [DOI] [PubMed] [Google Scholar]
- 24.Evangelou K, Kotsinas A, Mariolis-Sapsakos T, Giannopoulos A, Tsantoulis PK, Constantinides C, Troupis TG, Salmas M, Kyroudis A, Kittas C, et al. E2F-1 overexpression correlates with decreased proliferation and better prognosis in adenocarcinomas of Barrett oesophagus. J Clin Pathol. 2008;61:601–605. doi: 10.1136/jcp.2007.050963. [DOI] [PubMed] [Google Scholar]
- 25.Mitlianga PG, Gomez-Manzano C, Kyritsis AP, Fueyo J. Overexpression of E2F-1 leads to bax-independent cell death in human glioma cells. Int J Oncol. 2002;21:1015–1020. [PubMed] [Google Scholar]
- 26.Ho GH, Calvano JE, Bisogna M, Van Zee KJ. Expression of E2F-1 and E2F-4 is reduced in primary and metastatic breast carcinomas. Breast Cancer Res Treat. 2001;69:115–122. doi: 10.1023/a:1012230115173. [DOI] [PubMed] [Google Scholar]
- 27.van den Heuvel S, Dyson NJ. Conserved functions of the pRB and E2F families. Nat Rev Mol Cell Biol. 2008;9:713–724. doi: 10.1038/nrm2469. [DOI] [PubMed] [Google Scholar]
- 28.Conner EA, Lemmer ER, Omori M, Wirth PJ, Factor VM, Thorgeirsson SS. Dual functions of E2F-1 in a transgenic mouse model of liver carcinogenesis. Oncogene. 2000;19:5054–5062. doi: 10.1038/sj.onc.1203885. [DOI] [PubMed] [Google Scholar]
- 29.Suh DS, Yoon MS, Choi KU, Kim JY. Significance of E2F-1 overexpression in epithelial ovarian cancer. Int J Gynecol Cancer. 2008;18:492–498. doi: 10.1111/j.1525-1438.2007.01044.x. [DOI] [PubMed] [Google Scholar]
- 30.Xie Y, Yin Y, Li L, Ma Y, Xiao Q. Short interfering RNA directed against the E2F-1 gene suppressing gastric cancer progression in vitro. Oncol Rep. 2009;21:1345–1353. doi: 10.3892/or_00000360. [DOI] [PubMed] [Google Scholar]
- 31.Wang XT, Xie YB, Xiao Q. Lentivirus-mediated RNA interference targeting E2F-1 inhibits human gastric cancer MGC-803 cell growth in vivo. Exp Mol Med. 2011;43:638–645. doi: 10.3858/emm.2011.43.11.072. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Yang J, Liu X, Bhalla K, Kim CN, Ibrado AM, Cai J, Peng TI, Jones DP, Wang X. Prevention of apoptosis by Bcl-2: release of cytochrome c from mitochondria blocked. Science. 1997;275:1129–1132. doi: 10.1126/science.275.5303.1129. [DOI] [PubMed] [Google Scholar]
- 33.Jiang CG, Liu FR, Yu M, Li JB, Xu HM. Cimetidine induces apoptosis in gastric cancer cells in vitro and inhibits tumor growth in vivo. Oncol Rep. 2010;23:693–700. doi: 10.3892/or_00000686. [DOI] [PubMed] [Google Scholar]
- 34.Dong YB, Yang HL, Elliott MJ, Liu TJ, Stilwell A, Atienza C, McMasters KM. Adenovirus-mediated E2F-1 gene transfer efficiently induces apoptosis in melanoma cells. Cancer. 1999;86:2021–2033. doi: 10.1002/(sici)1097-0142(19991115)86:10<2021::aid-cncr20>3.0.co;2-1. [DOI] [PubMed] [Google Scholar]
- 35.Yang HL, Dong YB, Elliott MJ, Liu TJ, McMasters KM. Caspase activation and changes in Bcl-2 family member protein expression associated with E2F-1-mediated apoptosis in human esophageal cancer cells. Clin Cancer Res. 2000;6:1579–1589. [PubMed] [Google Scholar]
- 36.Han JC, Zhang KL, Chen XY, Jiang HF, Kong QY, Sun Y, Wu ML, Huang L, Li H, Liu J. Expression of seven gastric cancer-associated genes and its relevance for Wnt, NF-kappaB and Stat3 signaling. APMIS. 2007;115:1331–1343. doi: 10.1111/j.1600-0643.2007.00695.x. [DOI] [PubMed] [Google Scholar]
- 37.Honjo S, Kase S, Osaki M, Ardyanto TD, Kaibara N, Ito H. COX-2 correlates with F-box protein, Skp2 expression and prognosis in human gastric carcinoma. Int J Oncol. 2005;26:353–360. [PubMed] [Google Scholar]
- 38.Xiangming C, Natsugoe S, Takao S, Hokita S, Ishigami S, Tanabe G, Baba M, Kuroshima K, Aikou T. Preserved Smad4 expression in the transforming growth factor beta signaling pathway is a favorable prognostic factor in patients with advanced gastric cancer. Clin Cancer Res. 2001;7:277–282. [PubMed] [Google Scholar]
- 39.Fernandez PC, Frank SR, Wang L, Schroeder M, Liu S, Greene J, Cocito A, Amati B. Genomic targets of the human c-Myc protein. Genes Dev. 2003;17:1115–1129. doi: 10.1101/gad.1067003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Hu J, Liu ZS, Tang SL, He YM. Effect of hydroxyapatite nanoparticles on the growth and p53/c-Myc protein expression of implanted hepatic VX2 tumor in rabbits by intravenous injection. World J Gastroenterol. 2007;13:2798–2802. doi: 10.3748/wjg.v13.i20.2798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Liu GY, Luo Q, Xiong B, Pan C, Yin P, Liao HF, Zhuang WC, Gao HZ. Tissue array for Tp53, C-myc, CCND1 gene over-expression in different tumors. World J Gastroenterol. 2008;14:7199–7207. doi: 10.3748/wjg.14.7199. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Zhang L, Hou Y, Ashktorab H, Gao L, Xu Y, Wu K, Zhai J, Zhang L. The impact of C-MYC gene expression on gastric cancer cell. Mol Cell Biochem. 2010;344:125–135. doi: 10.1007/s11010-010-0536-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Zhang H, Kobayashi R, Galaktionov K, Beach D. p19Skp1 and p45Skp2 are essential elements of the cyclin A-CDK2 S phase kinase. Cell. 1995;82:915–925. doi: 10.1016/0092-8674(95)90271-6. [DOI] [PubMed] [Google Scholar]
- 44.Ju Y, Yu A, Sun X, Wu D, Zhang H. Glucosamine, a naturally occurring amino monosaccharide, inhibits A549 and H446 cell proliferation by blocking G1/S transition. Mol Med Rep. 2013;8:794–798. doi: 10.3892/mmr.2013.1584. [DOI] [PubMed] [Google Scholar]
- 45.Wei Z, Jiang X, Liu F, Qiao H, Zhou B, Zhai B, Zhang L, Zhang X, Han L, Jiang H, et al. Downregulation of Skp2 inhibits the growth and metastasis of gastric cancer cells in vitro and in vivo. Tumour Biol. 2013;34:181–192. doi: 10.1007/s13277-012-0527-8. [DOI] [PubMed] [Google Scholar]
- 46.Kim SY, Herbst A, Tworkowski KA, Salghetti SE, Tansey WP. Skp2 regulates Myc protein stability and activity. Mol Cell. 2003;11:1177–1188. doi: 10.1016/s1097-2765(03)00173-4. [DOI] [PubMed] [Google Scholar]
- 47.Fu M, Wang C, Li Z, Sakamaki T, Pestell RG. Minireview: Cyclin D1: normal and abnormal functions. Endocrinology. 2004;145:5439–5447. doi: 10.1210/en.2004-0959. [DOI] [PubMed] [Google Scholar]
- 48.Xia W, Li J, Chen L, Huang B, Li S, Yang G, Ding H, Wang F, Liu N, Zhao Q, et al. MicroRNA-200b regulates cyclin D1 expression and promotes S-phase entry by targeting RND3 in HeLa cells. Mol Cell Biochem. 2010;344:261–266. doi: 10.1007/s11010-010-0550-2. [DOI] [PubMed] [Google Scholar]
- 49.Oliveras-Ferraros C, Vazquez-Martin A, Cufí S, Torres-Garcia VZ, Sauri-Nadal T, Barco SD, Lopez-Bonet E, Brunet J, Martin-Castillo B, Menendez JA. Inhibitor of Apoptosis (IAP) survivin is indispensable for survival of HER2 gene-amplified breast cancer cells with primary resistance to HER1/2-targeted therapies. Biochem Biophys Res Commun. 2011;407:412–419. doi: 10.1016/j.bbrc.2011.03.039. [DOI] [PubMed] [Google Scholar]
- 50.Conway EM, Pollefeyt S, Cornelissen J, DeBaere I, Steiner-Mosonyi M, Ong K, Baens M, Collen D, Schuh AC. Three differentially expressed survivin cDNA variants encode proteins with distinct antiapoptotic functions. Blood. 2000;95:1435–1442. [PubMed] [Google Scholar]
- 51.Wenying Z, Zhaoning J, Zhimin Y, Dongyun C, Lili S. Survivin siRNA inhibits gastric cancer in nude mice. Cell Biochem Biophys. 2012;62:337–341. doi: 10.1007/s12013-011-9315-0. [DOI] [PubMed] [Google Scholar]
- 52.Dar AA, Majid S, de Semir D, Nosrati M, Bezrookove V, Kashani-Sabet M. miRNA-205 suppresses melanoma cell proliferation and induces senescence via regulation of E2F1 protein. J Biol Chem. 2011;286:16606–16614. doi: 10.1074/jbc.M111.227611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Guo SL, Ye H, Teng Y, Wang YL, Yang G, Li XB, Zhang C, Yang X, Yang ZZ, Yang X. Akt-p53-miR-365-cyclin D1/cdc25A axis contributes to gastric tumorigenesis induced by PTEN deficiency. Nat Commun. 2013;4:2544. doi: 10.1038/ncomms3544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Ye Q, Cai W, Zheng Y, Evers BM, She QB. ERK and AKT signaling cooperate to translationally regulate survivin expression for metastatic progression of colorectal cancer. Oncogene. 2014;33:1828–1839. doi: 10.1038/onc.2013.122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Cao W, Yang W, Fan R, Li H, Jiang J, Geng M, Jin Y, Wu Y. miR-34a regulates cisplatin-induce gastric cancer cell death by modulating PI3K/AKT/survivin pathway. Tumour Biol. 2014;35:1287–1295. doi: 10.1007/s13277-013-1171-7. [DOI] [PubMed] [Google Scholar]