

Abstract
Dyslexia is one of the most common learning disabilities, yet its brain basis and core causes are not yet fully understood. Neuroimaging methods, including structural and functional magnetic resonance imaging, diffusion tensor imaging, and electrophysiology, have significantly contributed to knowledge about the neurobiology of dyslexia. Recent studies have discovered brain differences prior to formal instruction that likely encourage or discourage learning to read effectively, distinguished between brain differences that likely reflect the etiology of dyslexia versus brain differences that are the consequences of variation in reading experience, and identified distinct neural networks associated with specific psychological factors that are associated with dyslexia.
1. Introduction
Developmental dyslexia, an unexplained difficulty in word reading accuracy and/or fluency, affects 5–12% of children [1,2]. Dyslexia is associated with many undesirable outcomes, including reduced educational attainment and academic self-esteem [3]. Furthermore, children with dyslexia tend to read far less outside of school than their peers [4], resulting in a widening gap in reading skills. Over the past 15 years, neuroimaging has made visible and quantifiable the brain differences that are associated with dyslexia; here, we review progress in the past few years in understanding the biological basis of dyslexia at a neural systems level.
Reading is a complex and slowly learned skill requiring the integration of multiple visual, linguistic, cognitive, and attentional processes. Neuroimaging methods including functional magnetic resonance imaging (fMRI), electroencephalography (EEG, and event-related potentials or ERPs), and magnetoencephalography (MEG), have revealed the brain regions most consistently involved in single word reading. In typically reading adults, these regions are lateralized to the language-dominant left hemisphere, and include inferior frontal, superior and middle temporal, and temporo-parietal regions [5]. In addition, experienced readers recruit an area of the left fusiform gyrus, termed the visual word form area (VWFA), which becomes preferentially engaged for orthographic (print) processing with reading experience [6–8]. This reading network (Figure 1) develops over years as children gain both specific reading skills and other abilities relevant to reading (e.g., 9). White-matter pathways that connect the components of the reading network can be quantified in size and strength by diffusion tensor imaging (DTI). Major tracts involved in reading include the left arcuate/superior longitudinal fasciculus, which connects frontal and temporal language regions, the inferior longitudinal fasciculus, which connects occipital and temporal lobes, and the corona radiata, which connects cortex to subcortical structures [10].
Figure 1.
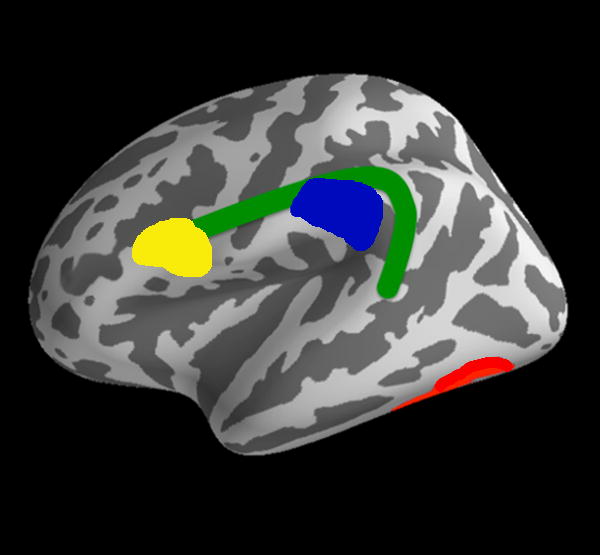
Schematic of the aspects of the reading brain in the left hemisphere. The inferior frontal gyrus (yellow) and the inferior parietal area (blue) are connected by the arcuate fasciculus (green). The fusiform gyrus, which includes the visual word form area, is in red. These regions are the most commonly found to be atypical in function or structure in dyslexia.
2. Psychological Bases of Dyslexia
Because reading involves multiple linguistic, visual, and attentional processes, it is likely that variable patterns of weakness may contribute to reading difficulty across children. Although it is unlikely that there is a single causal mechanism of dyslexia, some frequent likely causes have been identified (Table 1). The best understood cause for dyslexia is a weakness in phonological awareness (PA) for spoken (auditory) language that predicts and accompanies dyslexia [11]. Whereas learning a spoken language happens almost effortlessly, learning to read requires explicit knowledge and practice. Children must first become aware of the phonological structure of words, so that they can map those units of sound onto their corresponding printed letters.
Table 1.
Key Constructs in Reading and Potential Deficits in Dyslexia
| Construct | Definition | Example Tasks | Example Standardized Assessment |
|---|---|---|---|
| Phonological Awareness (PA) | Knowledge of, and ability to manipulate, the sound structure of words | - Say game without the /g/ - What word do these sounds make? /s/ - /i/ - /t/ - Name a word that rhymes with star |
- Comprehensive Test of Phonological Processing (CTOPP-2) [64] - Phonological Awareness Test (PAT-2) [65] |
| Rapid Automatized Naming (RAN) | Speed with which a series of familiar stimuli can be named aloud, reflecting efficient visual-verbal connections | Name, as quickly as possible, a 10×5 array of 5 randomly repeated objects, colors, letters, or numbers | - Rapid Automatized Naming–Rapid Alternating Stimulus Tests (RAN-RAS) [66] |
| Reading Fluency | Ability to read single words and connected text with sufficient accuracy and speed so as to support efficient comprehension | - Read aloud a list of common words or pseudowords as quickly and accurately as possible - Read aloud, quickly and accurately, paragraphs of increasing complexity |
- Test of Word Reading Efficiency (TOWRE-2) [67] - Gray Oral Reading Test (GORT-5) [68] |
A second psychological weakness associated with dyslexia relates to rapid automatized naming or RAN (Table 1). Slowness in naming may reflect difficulty in the integration of cognitive and linguistic processes involved in fluent reading [12]. Often, children who are especially poor readers have weaknesses in both PA and RAN [13], but some children exhibit only one of these weaknesses.
A third category of potential causal explanations for dyslexia relates to basic perceptual processes that may underlie the more proximal PA or RAN weaknesses, such as temporal sampling or processing [14–16], visual-spatial attention [17], or perceptual learning deficits [18]. These explanations are more mechanistic, but perhaps because they are more distal from reading per se, they are also more debated.
3. Functional and Structural Brain Differences in Dyslexia
Meta-analyses of primary research findings have identified broad patterns of functional and structural differences between typical and dyslexic readers. The most common functional brain differences, in children and adults, are reduced activations (hypoactivations) in left temporal, parietal, and fusiform (VWFA) regions [19–22]. In most cases, these hypoactivations arise from comparisons between two tasks or conditions, and thus reflect a lack of differential sensitivity to reading demands rather than a broader dysfunction of those brain regions. Increased activations in dyslexia are sometimes, but not consistently, observed in left inferior frontal and right-hemisphere regions. Variability across these findings may reflect differences in reading tasks, ages of participants, diversity among dyslexic groups, and other factors. Additionally, structural gray matter differences in dyslexia tend to co-localize with regions that show functional differences [23], but are also observed in the cerebellum, particularly in lobule VI [24, 25]. DTI studies often find reduced organization or volume in the left superior longitudinal fasciculus, including the arcuate fasciculus, and corona radiata fibers [26].
Because most neuroimaging studies of dyslexia have been conducted with children or adults who have had years of reading difficulty, it has been impossible to determine whether the brain differences are associated with the underlying neurobiological etiology of dyslexia, or are instead the consequence of years of altered and often vastly reduced reading experience (including compensatory alterations in reading networks). One approach to dissociating the cause and consequence of dyslexia in the brain has been to compare dyslexic children not only to age-matched typically reading children, but also to “ability-matched” children who are years younger than the dyslexic children but read at the same level. Ability-matched children are conceptualized as having approximately the same amount of reading experience as older dyslexic children. In one such study, dyslexic children exhibited reduced left parietal and occipito-temporal activations relative to both age- and ability-matched children, suggesting that these hypoactivations were related to the cause of dyslexia (in contrast, left prefrontal activations tracked ability level) [27].
A similar design challenged another idea about dyslexia, the magnocellular hypothesis of dyslexia. Previously, postmortem evidence from individuals with dyslexia revealed smaller magnocellular neurons in the lateral geniculate body [28], part of the visual pathway that is associated with motion perception. Accordingly, reduced activation for moving gratings in area MT, the cortical region most associated with motion perception, was found in adults with dyslexia [29]. When, however, children with dyslexia were examined, their MT activations were equivalent to ability-matched younger children, suggesting that the MT hypoactivation in dyslexia reflected reading experience [30]. This conclusion was further supported by evidence that remediation of the reading difficulty also enhanced MT activations in children with dyslexia [30]. These findings suggest that reduced MT activation for visual motion in dyslexia is a consequence, not a cause, of dyslexia. Similarly, many structural brain differences in dyslexia among age-matched groups were eliminated when a group with dyslexia was compared to ability-matched children [31].
Another strategy for identifying brain differences that underlie dyslexia has been the study of pre-reading children, typically in kindergarten, for whom brain differences cannot be the consequence of altered reading experience. Although pre-reading children cannot have a formal diagnosis of dyslexia, children can be identified as at-risk for dyslexia because of either a family history of dyslexia, which increases their risk of dyslexia by four times or more [32], or low performance on tests of pre-reading skills that tend to predict future reading difficulty (e.g., PA or RAN). Often, these children are followed longitudinally to determine which at-risk children actually progress to dyslexia.
Several neuroimaging studies have found brain differences preceding formal reading instruction in pre-reading children that resemble those observed in older children and adults. ERP studies of the mismatch negativity (MMN), an automatic response to an oddball auditory stimulus that is reduced in adults with dyslexia, have observed differences between infants with versus without a family history of dyslexia [33], and infants who do or do not develop dyslexia [34, 35]. Thus, the MMN may be a promising early endophenotype of dyslexia [36].
In MRI, pre-reading kindergartners with familial risk for dyslexia exhibited reduced bilateral occipitotemporal and left temporo-parietal activations for PA [37] and also bilaterally reduced grey matter volumes in similar posterior cortical regions [38]. Decreased grey-matter volumes in prefrontal and parieto-temporal regions were also found in 5- and 6-year-olds with maternal histories of reading difficulty [39]. In a heterogeneous sample of kindergartners, pre-reading children exhibited a positive correlation between measures of PA and both the size and microstructural white-matter organization of the left arcuate fasciculus [40]. Although it is not yet known which of these children will develop dyslexia, these studies support the idea that the most commonly observed functional and structural brain differences characterizing dyslexia are present before significant reading experience and therefore are more likely causes rather than consequences of dyslexia.
5. Advances in Understanding the Brain Basis of Aspects of Dyslexia
Brain Basis of Phonological Awareness (PA) Deficits
Impaired PA in dyslexia could reflect either a deficit in representing phonetic sounds and/or a deficit in access to and manipulation of those sounds (e.g., for mapping phonemes to print). Previously, a review of behavioral studies of dyslexia concluded that phonetic representations are intact, but access to those representations may be impaired [41]. Recently, a neuroimaging study with adults found that phonetic representations, as measured by multivoxel pattern analysis of activations in bilateral auditory cortices, were intact in dyslexia, but that functional and structural (DTI) connectivity between auditory cortices and left inferior frontal gyrus was reduced [42]. These findings favor the interpretation of dyslexia as being characterized by weakness in access to otherwise intact phonetic representations. Consistent with this conclusion is the finding that children with dyslexia exhibited reduced prefrontal activation when engaging in an auditory PA task, but no difference in temporal-lobe activation, as compared with both age- and ability-matched children [43].
Brain Basis of Rapid Automatized Naming (RAN) Deficits
RAN has been partially dissociated from PA as a skill essential for learning to read [12, 13], but now there is evidence for a neurobiological distinction between the two skills. A large structural MRI study of typical adult readers of Chinese found that phonological decoding ability was related to gray matter volume in left perisylvian cortex, whereas naming speed was related to volume in a more distributed network across all four lobes [44]. Further, functional activation to a PA task differed among groups of children with PA and RAN deficits, as predicted by the double deficit hypothesis. Activation in left inferior parietal lobule showed a gradient associated with PA ability, whereas activation in right cerebellar lobule VI showed a gradient with RAN ability [45].
Brain Basis of Reading Fluency Deficits
For older children with dyslexia who must read longer texts, slow reading is a major problem. Both the psychological and brain bases of reduced fluency for connected text, such as sentences and paragraphs, have been poorly understood relative to the many studies focusing on single-word reading. Two studies, however, examined reading fluency directly in dyslexia during fMRI by presenting sentences word-by-word at varying rates and testing comprehension, but the two studies reported disparate results [46, 47]. Both studies reported that more rapid reading resulted in greater activation of left fusiform cortex in the VWFA region. One study reported that children with dyslexia exhibited reduced activation related to fluency exclusively in left fusiform gyrus despite no significant differences in comprehension accuracy [46]. The other study reported that adults with dyslexia exhibited disproportionately worse comprehension accuracy and lesser activation in left prefrontal and superior temporal regions as a function of reading speed, but found no group difference in the VWFA region [47]. Although the populations and outcomes of the two studies differed, they have initiated the analysis of the brain basis of impaired reading fluency in dyslexia.
Brain Basis of Basic Perceptual Processes
Neuroimaging findings have reported neural correlates of atypical basic perceptual processes in dyslexia. Successful parsing of the speech signal depends on the ability of left auditory cortex to selectively amplify phonemic information in the 30 Hz (low gamma) range [48]. MEG revealed reduced entrainment, or synchronization of neural firing, to the 30 Hz frequency range in dyslexia, as well as reduced left-hemisphere specialization for such oscillations [49, 50]. These differences may impede the efficient transfer of acoustic information into more abstract phonemic representations. Individuals with dyslexia also exhibited reduced neural entrainment in response to linguistic stimuli [51, 52], differences in EEG signals that reflect integration of auditory and visual stimuli [53], and greater variability of auditory brainstem responses to speech sounds [54].
An advantage of understanding dyslexia in terms of basic perceptual processes is that the neural mechanisms of those processes can be studied in animals. Animal research has linked dyslexia-associated genes such as KIAA0319 with atypical neural migration [55] and impaired speech sound discrimination [55, 56], suggesting that the mechanism by which cortical abnormalities result in behavioral deficits is through the disruption of synchronous firing in response to oral language [57]. In humans, variation in KIAA0319 and two other dyslexia susceptibility genes has been associated with variation in left-hemisphere white matter and reading skill [58]. Such research may integrate findings from the genetic, cellular, cognitive, and behavioral levels in understanding the core deficits in dyslexia.
6. Conclusion
Progress in understanding the cognitive neuroscience of dyslexia may be approaching translation from basic research to intervention for children who will struggle to read. Remediation is known to be most effective in beginning readers, so early and accurate identification may promote effective intervention for children before they experience prolonged reading failure. Neuroimaging has identified biomarkers that enhance or outperform current behavioral measures in predicting long-term reading outcomes [59–63]. With further progress in understanding specific components of dyslexia (e.g., PA, RAN, fluency) it may also become possible to develop personalized interventions that target the specific patterns of weaknesses that undermine learning to read in individual children.
Highlights.
Neuroimaging is identifying brain differences related to causes of dyslexia.
Brain bases of specific aspects of dyslexia have been better identified.
Genetics may bridge study of neural mechanisms to dyslexia in humans.
Acknowledgments
This work was supported by a grant from the National Institutes of Health/National Institute of Child Health and Human Development (Grant #R01 HD067312). We thank Rachel Romeo for assistance with manuscript preparation.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Lyon GR, Shaywitz SE, Shaywitz BA. A definition of dyslexia. Ann Dyslexia. 2003;53:1–14. [Google Scholar]
- 2.Peterson RL, Pennington BF. Developmental dyslexia. Lancet. 2012;379:1997–2007. doi: 10.1016/S0140-6736(12)60198-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Undheim AM. Dyslexia and psychosocial factors. A follow-up study of young Norwegian adults with a history of dyslexia in childhood. Nordic J Psychiat. 2010;57:221–226. doi: 10.1080/08039480310001391. [DOI] [PubMed] [Google Scholar]
- 4.Cunningham AE, Stanovich KE. What reading does for the mind. Am Educ. 1998;22:137–149. [Google Scholar]
- 5.Price C. A review and synthesis of the first 20 years of PET and fMRI studies of heard speech, spoken language and reading. Neuroimage. 2012;62:816–847. doi: 10.1016/j.neuroimage.2012.04.062. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Cantlon JF, Pinel P, Dehaene S, Pelphrey KA. Cortical representations of symbols, objects, and faces are pruned back during early childhood. Cereb Cortex. 2011;21:191–199. doi: 10.1093/cercor/bhq078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Dehaene S, Cohen L. The unique role of the visual word form area in reading. Trends Cogn Sci. 2011;15:254–262. doi: 10.1016/j.tics.2011.04.003. [DOI] [PubMed] [Google Scholar]
- 8*.Vogel AC, Petersen SE, Schlaggar BL. The VWFA: it’s not just for words anymore. Front Hum Neurosci. 2014;8 doi: 10.3389/fnhum.2014.00088. This review summarized recent task-based and resting-state functional connectivity data from MRI to address the debate of whether the purported “visual word form area” (VWFA) is specific to reading or general to other visual processes. Understanding the nature of the VWFA could clarify how this region differs in dyslexia. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Brem S, Bach S, Kucian K, Guttorm TK, Martin E, Lyytinen H, Brandeis D, Richardson U. Brain sensitivity to print emerges when children learn letter-speech sound correspondences. Proc Natl Acad Sci U S A. 2010;107:7939–7944. doi: 10.1073/pnas.0904402107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Vandermosten M, Boets B, Wouters J, Ghesquière P. A qualitative and quantitative review of diffusion tensor imaging studies in reading and dyslexia. Neurosci Biobehav Rev. 2012;36:1532–1552. doi: 10.1016/j.neubiorev.2012.04.002. [DOI] [PubMed] [Google Scholar]
- 11.Melby-Lervåg M, Lyster S-AH, Hulme C. Phonological skills and their role in learning to read: a meta-analytic review. Psychol Bull. 2012;138:322–352. doi: 10.1037/a0026744. [DOI] [PubMed] [Google Scholar]
- 12.Norton ES, Wolf M. Rapid automatized naming (RAN) and reading fluency: Implications for understanding and treatment of reading disabilities. Annu Rev Psychol. 2012;63:427–452. doi: 10.1146/annurev-psych-120710-100431. [DOI] [PubMed] [Google Scholar]
- 13.Wolf M, Bowers PG. The double-deficit hypothesis for the developmental dyslexias. J Ed Psych. 1999;91:415. [Google Scholar]
- 14.Farmer ME, Klein RK. The evidence for a temporal processing deficit linked to dyslexia: A review. Psychon Bull Rev. 1995;2:460–493. doi: 10.3758/BF03210983. [DOI] [PubMed] [Google Scholar]
- 15.Tallal P, Miller S, Fitch RH. Neurobiological basis of speech: A case for the preeminence of temporal processing. Ann N Y Acad Sci. 1993;682:27–47. doi: 10.1111/j.1749-6632.1993.tb22957.x. [DOI] [PubMed] [Google Scholar]
- 16.Pammer K. Temporal sampling in vision and the implications for dyslexia. Front Hum Neurosci. 2014;7:933. doi: 10.3389/fnhum.2013.00933. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Franceschini S, Gori S, Ruffino M, Pedrolli K, Facoetti A. A causal link between visual spatial attention and reading acquisition. Curr Biol. 2012;22:814–819. doi: 10.1016/j.cub.2012.03.013. [DOI] [PubMed] [Google Scholar]
- 18.Gori S, Facoetti A. Perceptual learning as a possible new approach for remediation and prevention of developmental dyslexia. Vision Res. 2014;99:78–87. doi: 10.1016/j.visres.2013.11.011. [DOI] [PubMed] [Google Scholar]
- 19.Maisog JM, Einbinder ER, Flowers DL, Turkeltaub PE, Eden GF. A meta-analysis of functional neuroimaging studies of dyslexia. Ann N Y Acad Sci. 2008;1145:237–259. doi: 10.1196/annals.1416.024. [DOI] [PubMed] [Google Scholar]
- 20.Richlan F, Kronbichler M, Wimmer H. Functional abnormalities in the dyslexic brain: a quantitative meta-analysis of neuroimaging studies. Hum Brain Mapp. 2009;30:3299–3308. doi: 10.1002/hbm.20752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Richlan F, Kronbichler M, Wimmer H. Meta-analyzing brain dysfunctions in dyslexic children and adults. Neuroimage. 2011;56:1735–1742. doi: 10.1016/j.neuroimage.2011.02.040. [DOI] [PubMed] [Google Scholar]
- 22.Richlan F. Developmental dyslexia: dysfunction of a left hemisphere reading network. Front Hum Neurosci. 2012;6:120. doi: 10.3389/fnhum.2012.00120. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Linkersdörfer J, Lonnemann J, Lindberg S, Hasselhorn M, Fiebach CJ. Grey matter alterations co-localize with functional abnormalities in developmental dyslexia: an ALE meta-analysis. PLoS One. 2012;7:e43122. doi: 10.1371/journal.pone.0043122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Stoodley CJ, Stein JF. Cerebellar function in developmental dyslexia. Cerebellum. 2013;12:267–276. doi: 10.1007/s12311-012-0407-1. [DOI] [PubMed] [Google Scholar]
- 25.Stoodley CJ. Distinct regions of the cerebellum show gray matter decreases in autism, ADHD, and developmental dyslexia. Front Syst Neurosci. 2014;8:92. doi: 10.3389/fnsys.2014.00092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Vandermosten M, Boets B, Poelmans H, Sunaert S, Wouters J, Ghesquière P. A tractography study in dyslexia: neuroanatomic correlates of orthographic, phonological and speech processing. Brain. 2012;135:935–948. doi: 10.1093/brain/awr363. [DOI] [PubMed] [Google Scholar]
- 27.Hoeft F, Meyler A, Hernandez A, Juel C, Taylor-Hill H, Martindale JL, McMillon G, Kolchugina G, Black JM, Faizi A, et al. Functional and morphometric brain dissociation between dyslexia and reading ability. Proc Nat Acad Sci USA. 2007;104:4234–4239. doi: 10.1073/pnas.0609399104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Livingstone MS, Rosen GD, Drislane FW, Galaburda AM. Physiological and anatomical evidence for a magnocellular defect in developmental dyslexia. Proc Natl Acad Sci U S A. 1991;88:7943–7947. doi: 10.1073/pnas.88.18.7943. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Eden GF, VanMeter JW, Rumsey JM, Maisog JM, Woods RP, Zeffiro TA. Abnormal processing of visual motion in dyslexia revealed by functional brain imaging. Nature. 1996;382:66–69. doi: 10.1038/382066a0. [DOI] [PubMed] [Google Scholar]
- 30**.Olulade OA, Napoliello EM, Eden GF. Abnormal visual motion processing is not a cause of dyslexia. Neuron. 2013;79:180–190. doi: 10.1016/j.neuron.2013.05.002. This study showed how use of a younger, ability-matched control group can revise the interpretation of a brain difference in dyslexia. Abnormal activations in response to visual motion occurred in children with dyslexia relative to age-matched typical readers, but activations were equal to ability-matched typically reading younger children. These findings suggest that activation differences in dyslexic children reflected differences in reading experience and not the etiology of dyslexia. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31*.Krafnick AJ, Flowers DL, Luetje MM, Napoliello EM, Eden GF. An investigation into the origin of anatomical differences in dyslexia. J Neurosci. 2014;34:901–908. doi: 10.1523/JNEUROSCI.2092-13.2013. This paper addressed the question of what brain structure differences in children are a cause versus a consequence of dyslexia. Contrary to the many differences observed between dyslexic and age-matched control groups, differences between dyslexic and ability-matched younger groups included only gray matter in right precentral gyrus, and no white matter differences. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Fisher SE, Francks C. Genes, cognition and dyslexia: learning to read the genome. Trends Cogn Sci. 2006;10:250–257. doi: 10.1016/j.tics.2006.04.003. [DOI] [PubMed] [Google Scholar]
- 33.Leppänen PHT, Hämäläinen JA, Guttorm TK, Eklund KM, Salminen H, Tanskanen A, Torppa M, Puolakanaho A, Richardson U, Pennala R, Lyytinen H. Infant brain responses associated with reading-related skills before school and at school age. Neurophysiol Clin. 2012;42:35–41. doi: 10.1016/j.neucli.2011.08.005. [DOI] [PubMed] [Google Scholar]
- 34.van Zuijen TL, Plakas A, Maassen BAM, Maurits NM, van der Leij A. Infant ERPs separate children at risk of dyslexia who become good readers from those who become poor readers. Dev Sci. 2013;16:554–563. doi: 10.1111/desc.12049. [DOI] [PubMed] [Google Scholar]
- 35.Molfese D. Predicting dyslexia at 8 years of age using neonatal brain responses. Brain Lang. 2000;72:238–245. doi: 10.1006/brln.2000.2287. [DOI] [PubMed] [Google Scholar]
- 36*.Neuhoff N, Bruder J, Bartling J, Warnke A, Remschmidt H, Müller-Myhsok B, Schulte-Körne G. Evidence for the late MMN as a neurophysiological endophenotype for dyslexia. PLoS One. 2012;7:e34909. doi: 10.1371/journal.pone.0034909. The authors studied children with dyslexia and their unaffected siblings in order to examine genetic effects on the amplitude of the mismatch negativity ERP component. This approach breaks ground on characterizing the variety of phenotypes in dyslexia based on new insights into their biological bases. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37**.Raschle NM, Zuk J, Gaab N. Functional characteristics of developmental dyslexia in left-hemispheric posterior brain regions predate reading onset. Proc Natl Acad Sci USA. 2012;109:2156–2161. doi: 10.1073/pnas.1107721109. This is the first study showing that there are anatomically specific differences in brain function in pre-reading kindergartners at familial risk for dyslexia even prior to formal reading instruction in school. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Raschle NM, Chang M, Gaab N. Structural brain alterations associated with dyslexia predate reading onset. Neuroimage. 2011;57:742–749. doi: 10.1016/j.neuroimage.2010.09.055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Black JM, Tanaka H, Stanley L, Nagamine M, Zakerani N, Thurston A, Kesler S, Hulme C, Lyytinen H, Glover GH, et al. Maternal history of reading difficulty is associated with reduced language-related gray matter in beginning readers. Neuroimage. 2012;59:3021–3032. doi: 10.1016/j.neuroimage.2011.10.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Saygin ZM, Norton ES, Osher DE, Beach SD, Cyr AB, Ozernov-Palchik O, Yendiki A, Fischl B, Gaab N, Gabrieli JDE. Tracking the roots of reading ability: white matter volume and integrity correlate with phonological awareness in prereading and early-reading kindergarten children. J Neurosci. 2013;33:13251–13258. doi: 10.1523/JNEUROSCI.4383-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Ramus F, Szenkovits G. What phonological deficit? Q J Exp Psychol. 2008;61:129–141. doi: 10.1080/17470210701508822. [DOI] [PubMed] [Google Scholar]
- 42**.Boets B, Op de Beeck HP, Vandermosten M, Scott SK, Gillebert CR, Mantini D, Bulthé J, Sunaert S, Wouters J, Ghesquière P. Intact but less accessible phonetic representations in adults with dyslexia. Science. 2013;342:1251–1254. doi: 10.1126/science.1244333. The authors reported comparable neural representations of phonemes in temporal-lobe auditory cortex, as revealed by MVPA, in controls and dyslexics; weakened structural and functional connectivity in inferior frontal and temporal regions points to inefficient access to phonological representations, rather than weakened representations, as the source of the PA deficit in dyslexia. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Kovelman I, Norton ES, Christodoulou JA, Gaab N, Lieberman DA, Triantafyllou C, Wolf M, Whitfield-Gabrieli S, Gabrieli JDE. Brain basis of phonological awareness for spoken language in children and its disruption in dyslexia. Cereb Cortex. 2012;22:754–764. doi: 10.1093/cercor/bhr094. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44**.He Q, Xue G, Chen C, Chen C, Lu Z-L, Dong Q. Decoding the neuroanatomical basis of reading ability: a multivoxel morphometric study. J Neurosci. 2013;33:12835–12843. doi: 10.1523/JNEUROSCI.0449-13.2013. This study examined a sample of over 400 college-age readers with the goal of identifying separable components of reading and their neural correlates. Using a multivariate analysis of gray matter volume, authors identified unique networks associated with phonological decoding, form-sound association, and naming speed. Notably, naming speed was associated with a distributed and bilateral set of brain regions. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Norton ES, Black JM, Stanley LM, Tanaka H, Gabrieli JDE, Sawyer C, Hoeft F. Functional neuroanatomical evidence for the double-deficit hypothesis of developmental dyslexia. Neuropsychologia. 2014;61:235–246. doi: 10.1016/j.neuropsychologia.2014.06.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Langer N, Benjamin C, Minas J, Gaab N. The neural correlates of reading fluency deficits in children. Cereb Cortex. 2013 doi: 10.1093/cercor/bht330. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Christodoulou JA, Del Tufo SN, Lymberis J, Saxler PK, Ghosh SS, Triantafyllou C, Whitfield-Gabrieli S, Gabrieli JDE. Brain bases of reading fluency in typical reading and impaired fluency in dyslexia. PLoS One. 2014;9:e100552. doi: 10.1371/journal.pone.0100552. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Giraud A-L, Poeppel D. Cortical oscillations and speech processing: emerging computational principles and operations. Nat Neurosci. 2012;15:511–517. doi: 10.1038/nn.3063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Lehongre K, Ramus F, Villiermet N, Schwartz D, Giraud AL. Altered low-gamma sampling in auditory cortex accounts for the three main facets of dyslexia. Neuron. 2011;72:1080–1090. doi: 10.1016/j.neuron.2011.11.002. [DOI] [PubMed] [Google Scholar]
- 50*.Lehongre K, Morillon B, Giraud A-L, Ramus F. Impaired auditory sampling in dyslexia: further evidence from combined fMRI and EEG. Front Hum Neurosci. 2013;7:454. doi: 10.3389/fnhum.2013.00454. Although temporal sampling and oscillatory deficits in dyslexia have been proposed before, the authors trace the problem to a reduced specialization for phoneme-level rhythms in left auditory cortex. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Bedo N, Ribary U, Ward LM. Fast dynamics of cortical functional and effective connectivity during word reading. PLoS One. 2014;9:e88940. doi: 10.1371/journal.pone.0088940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52*.White-Schwoch T, Kraus N. Physiologic discrimination of stop consonants relates to phonological skills in pre-readers: a biomarker for subsequent reading ability? Front Hum Neurosci. 2013;7:899. doi: 10.3389/fnhum.2013.00899. Examining 4-year-old children, the authors found that the auditory brainstem response to speech sounds was associated with phonological awareness skills. This is a promising potential early biomarker of dyslexia that should be studied longitudinally, and a potential window into how neural differences can manifest as reading problems. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Hasko S, Bruder J, Bartling J, Schulte-Körne G. N300 indexes deficient integration of orthographic and phonological representations in children with dyslexia. Neuropsychologia. 2012;50:640–654. doi: 10.1016/j.neuropsychologia.2012.01.001. [DOI] [PubMed] [Google Scholar]
- 54.Hornickel J, Kraus N. Unstable representation of sound: a biological marker of dyslexia. J Neurosci. 2013;33:3500–3504. doi: 10.1523/JNEUROSCI.4205-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Szalkowski CE, Fiondella CG, Galaburda AM, Rosen GD, Loturco JJ, Fitch RH. Neocortical disruption and behavioral impairments in rats following in utero RNAi of candidate dyslexia risk gene Kiaa0319. Int J Dev Neurosci. 2012;30:293–302. doi: 10.1016/j.ijdevneu.2012.01.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Centanni TM, Booker AB, Sloan AM, Chen F, Maher BJ, Carraway RS, Khodaparast N, Rennaker R, Loturco JJ, Kilgard MP. Knockdown of the dyslexia-associated gene Kiaa0319 impairs temporal responses to speech stimuli in rat primary auditory cortex. Cereb Cortex. 2013 doi: 10.1093/cercor/bht028. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Giraud A-L, Ramus F. Neurogenetics and auditory processing in developmental dyslexia. Curr Opin Neurobiol. 2013;23:37–42. doi: 10.1016/j.conb.2012.09.003. [DOI] [PubMed] [Google Scholar]
- 58.Darki F, Peyrard-Janvid M, Matsson H, Kere J, Klingberg T. Three dyslexia susceptibility genes, DYX1C1, DCDC2, and KIAA0319, affect temporo-parietal white matter structure. Biol Psychiatry. 2012;72:671–676. doi: 10.1016/j.biopsych.2012.05.008. [DOI] [PubMed] [Google Scholar]
- 59.Maurer U, Bucher K, Brem S, Benz R, Kranz F, Schulz E, van der Mark S, Steinhausen H-C, Brandeis D. Neurophysiology in preschool improves behavioral prediction of reading ability throughout primary school. Biol Psychiatry. 2009;66:341–348. doi: 10.1016/j.biopsych.2009.02.031. [DOI] [PubMed] [Google Scholar]
- 60*.Bach S, Richardson U, Brandeis D, Martin E, Brem S. Print-specific multimodal brain activation in kindergarten improves prediction of reading skills in second grade. Neuroimage. 2013;82:605–615. doi: 10.1016/j.neuroimage.2013.05.062. This was the first study to combine fMRI and EEG/ERP in kindergarten to predict reading outcomes in 2nd grade. Both fMRI and ERP measures related to visual processing in the left occipito-temporal region predicted 2nd grade reading ability. In this small group of children, results suggest that brain imaging in kindergarten could significantly improve classification of future good versus poor readers. [DOI] [PubMed] [Google Scholar]
- 61.Hoeft F, McCandliss BD, Black JM, Gantman A, Zakerani N, Hulme C, Lyytinen H, Whitfield-Gabrieli S, Glover GH, Reiss AL, Gabrieli JDE. Neural systems predicting long-term outcome in dyslexia. Proc Nat Acad Sci USA. 2011;108:361–366. doi: 10.1073/pnas.1008950108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Myers CA, Vandermosten M, Farris EA, Hancock R, Gimenez P, Black JM, Casto B, Drahos M, Tumber M, Hendren R, Hulme C, Hoeft F. Structural changes in white matter are uniquely related to children’s reading development. Psychol Sci. 2014 doi: 10.1177/0956797614544511. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63*.Eicher JD, Gruen JR. Imaging-genetics in dyslexia: connecting risk genetic variants to brain neuroimaging and ultimately to reading impairments. Mol Genet Metab. 2013;110:201–212. doi: 10.1016/j.ymgme.2013.07.001. This review paper examined genetic contributions to reading ability, and in particular, to the reading brain. The authors highlight the importance of the brain as an endophenotype or link between genes and reading behavior, and the importance of subtypes for understanding the heterogeneity of dyslexia. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Wagner RK, Torgesen JK, Rashotte CA, Pearson N. The Comprehensive Test of Phonological Processing, Second Edition (CTOPP-2) Austin, Texas: Pro-Ed; 2013. [Google Scholar]
- 65.Robertson C, Salter W. The Phonological Awareness Test, Second Edition (PAT-2) East Moline, Illinois: LinguiSystems; 2007. [Google Scholar]
- 66.Wolf M, Denckla MB. RAN/RAS: Rapid Automatized Naming and Rapid Alternating Stimulus Tests. Austin, Texas: Pro-Ed; 2005. [Google Scholar]
- 67.Torgesen JK, Wagner RK, Rashotte CA. Test of Word Reading Efficiency, Second Edition (TOWRE-2) Austin, Texas: Pro-Ed; 2012. [Google Scholar]
- 68.Wiederholt JL, Bryant BR. Gray Oral Reading Tests, Fifth Edition (GORT-5) Austin, Texas: Pro-Ed; 2012. [Google Scholar]