

Abstract
Like language, semantic memory is productive: It extends itself through self-derivation of new information through logical processes such as analogy, deduction, and induction, for example. Though it is clear these productive processes occur, little is known about the time course over which newly self-derived information becomes incorporated into semantic knowledge. In the present research, we used event-related potentials (ERPs) to examine this dynamic process. Subjects were presented with separate but related facts that when integrated with one another, supported generation of new information (Integration facts). After two 400 ms presentations, P600 responses to Integration facts differed from responses to Novel facts and did not differ from responses to Well-known facts, suggesting that the newly self-derived information had been incorporated into the knowledge base. The finding of rapid transition from newly self-derived to well-known helps explain the richness of semantic memory. By implication, it also may contribute to the absence of episodic information specifying when and where semantic contents were acquired.
Semantic memory is an open system to which new information is added regularly. Some new entries come about through direct observation or through explicit learning or instruction. Others are self-derived, through logical processes such as analogy (e.g., Gentner, 1983), deduction (e.g., Johnson-Laird & Byrne, 1991), and induction (e.g., Holland, Holyoak, Nisbett, & Thagard, 1989). Semantic memory also can be elaborated by self-derivation of new knowledge through integration of information acquired in separate yet related learning episodes. For example, children presented with separate passages of text in which they learn that “dolphins live in groups called pods,” and “dolphins talk by clicking and squeaking” subsequently generate the novel understanding that “pods talk by clicking and squeaking” (Bauer, King, Larkina, Varga, & White, 2012; Bauer & San Souci, 2010; Bauer, Varga, King, Nolen, & White, in press; Varga & Bauer, 2013). As a result of the productive nature of semantic memory, the boundaries of the conceptual knowledge network extend far beyond what has been directly learned. Indeed, this feature of semantic memory may help explain the absence of episodic information specifying when and where its contents were acquired—some entries in the semantic knowledge network were never directly experienced and thus there is no event to locate in time and place. Though we know these elaborative processes occur, we know little about the time course over which newly self-derived information becomes incorporated into the semantic knowledge base. In the present research, we used event-related potentials (ERPs) to examine the dynamic process of transition of information from newly self-derived to well-known.
Productive self-derivation of new knowledge is pervasive in adults (e.g., Johnson-Laird, 2006) and children (e.g., Moshman, 2004) alike. In some cases, the newly-derived information may be fleeting, such as when separate premises are combined in the service of comprehending a text (e.g., Yule, 2010). For example, the premises “Mary entered the room” and “She turned,” can be integrated to derive the information that it was Mary who turned. This type of self-derivation is fast-acting—happening on the order of milliseconds—and the resulting inference likely survives in working memory only long enough to ensure adequate comprehension (McKoon & Ratcliff, 1992). In other cases, the products of self-derivation may become integrated into the knowledge base and endure over time. In scientific reasoning, for example, separate empirically derived observations may be combined to reach logical conclusions. The observations that “the movement of tectonic plates produces jagged mountains,” and “the Appalachians are jagged mountains,” may lead to the logical conclusion that “the Appalachians were produced by the movement of tectonic plates.” In such cases, information that was neither directly observed nor explicitly learned may become part of the more-or-less permanent store of semantic or conceptual knowledge.
To date, the transition of information from newly self-derived to known has been assumed but not observed. Because they are measures of information processing in real time, ERPs are ideally suited to examination of this transition. ERPs are scalp-recorded electrical activity that reflect the summation of the postsynaptic potentials of large populations of neurons activated in response to specific stimuli. Several ERP components are sensitive to the status of semantic knowledge and thus may illuminate the process of transition from newly self-derived to known. One especially robust effect is the N400 (Kutas & Hillyard, 1980): a pronounced change in neural activity 300–500 ms after stimulus onset, that is maximal at 400 ms (for reviews, see for e.g., Federmeier & Laszlo, 2009; Kutas & Federmeier, 2011). When the stimulus is novel, the response is a marked negative deflection, relative to the baseline (i.e., pre-stimulus) level of neural activity. When the stimulus is well-known, the neural response is a substantially smaller change. Across stimulus classes, when stimuli are repeated, the amplitude of the N400 decreases (e.g., Besson & Kutas, 1993), presumably reflecting facilitated semantic processing resulting from the previous presentation of the item (e.g., Olichney, Van Petten, Paller, Salmon, Iragui, & Kutas, 2000).
Neural activity both before and after the N400 also can be revealing of the knowledge status of stimuli. The P200—a positive deflection 150–300 ms after stimulus onset—is sensitive to comparisons between sensory inputs and stored memory representations and generally is larger when they match (e.g., Evans & Federmeier, 2007; Federmeier, Mai, & Kutas, 2005; Wlotko & Federmeier, 2007). The P200 thus can be expected to differentiate novel from known information.
A third component, termed the P600 (e.g., Rugg, Roberts, Potter, Pickles, & Nagy, 1991) or Late Positive Component (e.g., Olichney, Morris, Ochoa, Salmon, Thal, Kutas, et al., 2002), reflects ongoing processing of a stimulus. It is marked by positive activity beginning around 500 ms and extending for several hundred ms, peaking around 600 ms (e.g., Coulson, King, & Kutas, 1998; Hagoort, Brown, & Osterhout, 1999). The P600 is associated with a variety of processes, including language comprehension and production (Hagoort et al., 1999; Osterhout & Holcomb, 1993), reading music (e.g., Patel, Gibson, Ratner, Besson, & Holcomb, 1998), and abstract pattern completion (e.g., Lelekov-Boissard & Dominey, 2002). In these contexts, the P600 is thought to reflect the effort associated with extracting meaning from stimuli, especially stimuli that are unexpected or improbable (Hagoort, 2003; Osterhout & Holcomb, 1993). Positive activity that peaks around 600 ms also is reflective of the status of contents in long-term memory (e.g., Besson, Kutas, & Van Petten, 1992; Olichney et al., 2000, 2002). Upon initial presentation of a stimulus, a larger P600 (or LPC) is associated with better subsequent memory (e.g., Rugg, Mark, Walla, Schloerscheidt, Birch, & Allan, 1998). Like the N400, the P600 changes with stimulus repetition. Unlike the N400, the effect is different for different stimulus classes. When the repeated stimulus is semantically predictable from prior context (i.e., such as a sentence or category label), a reduction in the size of the P600 is observed. When the repeated stimulus is not semantically predictable, a smaller or no change in the P600 is apparent (e.g., Besson et al., 1992; Olichney et al., 2000, 2002; Van Petten, Kutas, Kluender, Mitchiner, & McIsaac, 1991). Van Petten and colleagues (1991) have interpreted this patterns to indicate that the first presentation initiates an attempt to retrieve relevant contents from long-term memory, resulting in a high amplitude response. The second presentation is associated with a decrease in the amplitude of the P600 to the extent that the initial retrieval attempt was successful, rendering the relevant material more readily accessible upon the second presentation, thus requiring less effortful processing of the stimulus the second time. Due to the apparent sensitivity of the P600 to the existing contents of memory, it may be especially informative of a change in knowledge state from novel to known.
In the present research, we capitalized on the sensitivity of ERPs to the status of semantic knowledge to observe the transition from novel to known of newly self-derived information. In two experiments, we arranged self-derivation of information by providing subjects with separate pieces of information that could be integrated with one another to yield new knowledge (Bauer et al., 2012, in press; Bauer & San Souci, 2010; Varga & Bauer, 2013). For example, when presented with the information that “apple seeds are called pips,” and “pips contain the compound cyanide,” subjects could be expected to generate the new information that “apple seeds contain the compound cyanide.” In Experiment 1a, we recorded ERPs as subjects processed facts derived through integration. We predicted that as the new information was processed, it would transition from being treated as novel (small P200, large N400, large P600) to known (reduced P600). As summarized above, each of these ERP components has been found to be sensitive to the status of the contents of memory. In the present research, we adopted the novel approach of using them as temporal markers of change in the knowledge status of information. Experiment 1b was an ERP control experiment to eliminate priming as an explanation for the observed pattern. In Experiment 2, we used a behavioral paradigm to verify that the integration facts were newly self-derived, based on integration of separate sources from a prior learning episode.
Experiment 1a
Method
Subjects
Subjects were 30 adults (M = 20.5 years, range 18.4–23.7 years) enrolled as undergraduate students at Emory University. An additional 4 subjects were eliminated from analysis due to an insufficient number of trials (see Procedure). Written informed consent was obtained prior to beginning the procedure. Subjects earned course credit or a $15 gift card for participation. All procedures were approved by the Emory University IRB.
Materials
For the encoding phase, the materials were 120 sentences, each 5–15 words long. Eighty sentences conveyed information that subjects were likely to know and which, based on Virginia state academic standards, were expected to have been acquired in elementary school (“a year has 365 days”), middle school (“a hexagon has six sides”), high school (“the Ninety Five Theses were the catalyst for the Protestant Reformation”), or college (“Beta Thalassemia is caused by a hemoglobin defect”). The remaining 40 sentences featured 20 pairs of related facts that subjects were unlikely to know (“the world’s largest biome is the boreal forest” and “the boreal forest is the Taiga”). When integrated with one another, each pair of related facts permitted self-generation of a novel fact (“the world’s largest biome is the Taiga”). At no time were subjects informed that any sentences were related and related sentences were separated by a minimum of 10 unrelated sentences.
For the test phase, the stimuli were 80 sentences, 5–9 words in length. None of the sentences had been previously presented. There were four categories of sentences: (a) well-known facts (Well-known; e.g., “The Eifel Tower is in France”; N = 20); (b) facts unlikely to be known, but which could be derived through integration of separate facts presented during encoding (Integration; e.g., “the world’s largest biome is the Taiga”; N = 20); (c) novel control facts unrelated to any previously encountered (Novel-non-integration; e.g., “Venus is the only planet that rotates clockwise”; N = 20); and (d) novel control facts derived from separate premises to which subjects had not been exposed during encoding (Novel-integration; e.g., “Kool-Aid was invented in Nebraska,” derived from “Kool-Aid originally was called Fruit-Smack,” and “Fruit-Smack was invented in Nebraska”; N = 20). The Novel-integration facts were included to control for the possibility that ERP responses to novel and Integration stimuli might differ due to uncontrolled features of facts derived through integration. Preliminary analyses revealed no differences between the two types of novel control facts (see Results). Therefore, they were collapsed into the Novel category used in all analyses. The likely knowledge status of the facts used in the test phase (i.e., Well-known, Integration, Novel) initially was verified through pilot testing and subsequent to the ERP test, was checked for each individual subject.
Procedure
At the beginning of the session, subjects were advised that we were interested in judgments of when information should be learned. Subjects then read each of 120 sentences presented on a computer (Figure 1, Panel A). Throughout the encoding task, to ensure attention to the information, subjects judged when in school the facts in the sentences should be learned: elementary, middle, high school, or college. At no time were subjects informed that any of the sentences were related. Related sentences were separated by a minimum of 10 unrelated sentences (M = 44).
Figure 1.

During encoding, sentences were presented one at a time on a computer. Duration of presentation was based on the number of characters in the sentence and ranged from 3–7 sec (100 ms per character). After reading each sentence, subjects verbally indicated when in school the fact in the sentence would be learned. Sentences were presented in one of 12 random orders, each used approximately equally often (Panel A). During the ERP test (Experiment 1), new sentences were presented one word at a time. ERPs were time locked to the target word, which always was the last word in the sentence. After each sentence, subjects used a game controller to determine when the fact would be learned. Trials were presented in one of 4 random orders, each used approximately equally often (Panel B). During behavioral testing (Experiment 2), new sentences were presented one word at a time. The final word was omitted and followed by a decision screen with four choices, one of which was correct. Subjects registered their choices with a game-controller. Trials were presented in one of 8 random orders, each used approximately equally often (Panel C).
Following the encoding task, subjects were told that we wanted to observe brain activity as they made the learning judgments. They were fitted with an Advanced Neuro Technology (A.N.T.) WaveGuard cap with 32 electrodes positioned according to an adaptation of the International 10–20 system (Jasper, 1958). Application required an average of 20 min. Once the cap was fitted, subjects were seated approximately 90 cm in front of a monitor. They were instructed to sit as still as possible, look at the center of the screen, and blink only between trials.
ERPs were collected as subjects read each of 80 sentences. Each word of the sentences was presented for 400 ms, followed by an inter-stimulus interval of 200 ms (Figure 1, Panel B). Prior to the test proper, subjects completed and correctly answered four practice questions, thereby demonstrating the ability to read and comprehend the sentences in the rapid serial visual presentation. ERPs were time locked to presentation of the target word, which always was the sentence-final word. The target word was followed by a blank screen for 1400 ms. Data were recorded for 100 ms before stimulus onset and 1800 ms after stimulus onset. After reading the full sentence, subjects used a game-controller to indicate when the fact in the sentence should be learned. To permit observation of anticipated change over time in the neural status of the self-derived information, each sentence was presented twice. All sentences were presented once (80 trials) before any was repeated (80 trials: total trials = 160).
Electrodes were referenced online to mathematically averaged mastoids. Data were sampled at 256 Hz continuously using ASA computer software (A.N.T. Software B.V., Enschede, The Netherlands). Impedances were consistently below 10 kΩ, and generally below 5 kΩ.
Following the ERP task, subjects were given a list of the 80 facts presented in the ERP task (i.e., 20 Well-Known, 20 Integration, 40 Novel) and asked to indicate whether they knew each fact before participating in the study. Well-known facts that subjects indicated they did not know prior to the study were eliminated from data analysis, as were Integration and Novel facts that subjects indicated they had known prior to the study (M = 11% facts eliminated per subject).
Data Reduction
Raw EEG data were individually bandpass filtered with half-power cutoffs of 0.1 and 30Hz, and a roll-off of 24 decibels/octave. Data then were processed with independent component analysis using EEGLAB 6.03b (Delorme & Makeig, 2004) running under Matlab 7.7.0 (MathWorks, Natick, MA, USA) to remove eye-blink and saccade artifacts. The EEG data for each participant were individually baseline corrected, and trials containing amplitudes that exceeded ±150 μV (typically caused by excessive movement or muscle activity) were rejected. Participants with 10 or more usable trials in each condition per presentation were included (4 participants were excluded). The numbers of trials included in analyses by condition and presentation (first, second) are provided in Table 1, Panel A. Across participants, we created separate grand averages for trials in the Well-known, Integration, and Novel conditions.
Table 1.
Experiment 1a: Descriptive Statistics (Mean, Standard Deviation, Range) for the Number of Trials Included in Analyses (Panel A) and Descriptive Statistics (Mean, Standard Deviation, Standard Error) for Mean Amplutides for the P200, N400, and P600 Event-Related Potential (ERP) Components for Each Condition for the Centro-parietal Cluster (P3/P4, CP1/CP2, CP5/CP6) (Panel B)
| ERP Component and Stimulus Condition | Presentation
|
|||||
|---|---|---|---|---|---|---|
| First | Second | |||||
| Panel A: Number of trials included in analyses
| ||||||
| M | SD | Range | M | SD | Range | |
|
| ||||||
| Well-known | 19.07 | 2.29 | 10–20 | 18.43 | 2.21 | 10–20 |
| Pseudo-Integration | 16.33 | 2.90 | 10–20 | 15.90 | 2.72 | 10–20 |
| Novel | 33.13 | 4.78 | 24–40 | 32.90 | 5.18 | 23–40 |
|
| ||||||
| Panel B: Mean amplitude (Amplitude μV)
| ||||||
| M | SD | StE | M | SD | StE | |
|
| ||||||
| P200
| ||||||
| Well-known | 3.80 | 3.83 | 0.70 | 2.06 | 2.79 | 0.51 |
| Integration | 2.87 | 3.23 | 0.59 | 2.96 | 3.45 | 0.63 |
| Novel | 1.68 | 3.18 | 0.58 | 3.10 | 2.85 | 0.52 |
|
| ||||||
| N400
| ||||||
| Well-known | 2.46 | 3.18 | 0.58 | 0.63 | 3.07 | 0.56 |
| Integration | 0.92 | 3.56 | 0.65 | 0.35 | 3.07 | 0.56 |
| Novel | −0.72 | 2.90 | 0.53 | 1.25 | 3.01 | 0.55 |
|
| ||||||
| P600
| ||||||
| Well-known | 3.64 | 3.07 | 0.56 | 1.39 | 2.52 | 0.46 |
| Integration | 3.61 | 3.01 | 0.55 | 2.10 | 2.79 | 0.51 |
| Novel | 3.43 | 2.79 | 0.51 | 3.56 | 2.85 | 0.52 |
To examine P200, N400, and P600 effects, we parsed the data into time windows of 150–300 ms, 300–500 ms, and 500–900 ms, respectively. Because all of these effects have been observed to be maximal at centro-parietal sites (Coulson et al., 1998; Federmeier et al., 2005; Hagoort et al., 1999; Van Petten, Coulson, Rubin, Plante, & Parks, 1999; Wlotko & Federmeier, 2007), we selected a centro-parietal cluster as the focus of investigation (electrode sites P3/P4, CP1/CP2, CP5/CP6; Figure 2). For all effects, the dependent measure was mean amplitude. Significant main effects were examined using posthoc tests, Bonferroni corrected.
Figure 2.
Representation of the montage of electrodes with the location of the centro-parietal cluster (P3/P4, CP1/CP2, CP5/CP6) outlined.
Results
Preliminary Analyses
Separate analyses of variance for each ERP component (P200, N400, P600) and each presentation (first, second) revealed no statistically significant differences between the two types of novel control facts (Novel-non-integration and Novel-integration). Specifically, at the first presentation, the F(1, 29) values for the P200, N400, and P600 were 1.43, 1.34, and 2.60, respectively; p values = .24, .26, and .12, respectively. At the second presentation, the F(1, 29) values for the P200, N400, and P600 were 0.00, 0.13, and 0.00, respectively; p values = .99, .72, and .99, respectively. Because of lack of evidence of meaningful differences in response between the two type of novel control facts, they were collapsed into the Novel category used in all analyses.
Main Analyses
Neural activity differed for Well-known, Integration, and Novel facts. Reflecting change in the status of the newly-integrated information from novel to known, the pattern was different the first (P200 and N400 effects) versus the second (P600 effects) time the facts were presented. Though we focused analyses on the centro-parietal cluster (Figure 2), as reflected in Figure 3, the patterns were observed across the scalp (analyses at other sites are available from the authors). Descriptive statistics at the centro-parietal cluster are provided in Table 1, Panel B.
Figure 3.

Scalp ERP of central and posterior sites that demonstrate widespread patterns of P200 and N400 in the first presentation (Panel A) and P600 in the second presentation (Panel B). The centro-parietal cluster (electrodes P3/P4, CP1/CP2, CP5/CP6) was the focus of analysis.
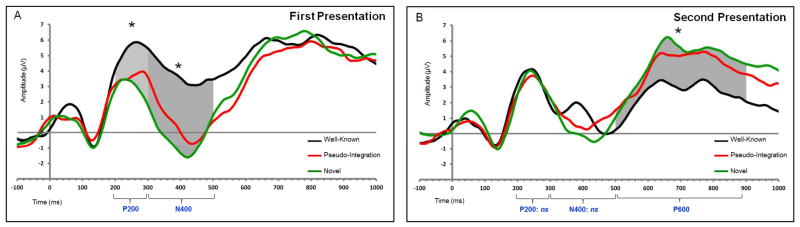
At the first 400 ms presentation of the stimuli (Figure 4, Panel A), P200 and N400 effects were apparent: F(2, 58) = 8.94, p < .001, pη2 = .236, and F(2, 58) = 16.17, p < .001, pη2 = .358, respectively. Reflecting the match between the sensory input and stored memory representations (i.e., familiar information: e.g., Evans & Federmeier, 2007; Federmeier et al., 2005), the P200 was largest for Well-known facts, and differed significantly from the response to Novel facts (p = .002). The P200 to Integration facts was intermediate and did not differ from either Well-known or Novel. The N400 revealed differences across all three conditions (ps = .032). For Novel facts, the response was a marked negative deflection relative to baseline that differed significantly from the response to Well-known facts. Integration facts—being neither entirely novel nor well-known—produced an N400 that was intermediate between Well-known and Novel. The patterns are reflective of differential accessibility of information in long-term stores (e.g., Kutas & Federmeier, 2000). High amplitude responses in the window corresponding to the P600 (500–900 ms) did not differ across conditions. The consistent pattern of high amplitude response likely is indicative of subjects’ attempts to retrieve relevant contents from long-term memory (e.g., Van Petten et al., 1991).
Figure 4.

In response to the first 400 ms presentation of the stimuli, analysis of the centro-parietal cluster (P3/P4, CP1/CP2, CP5/CP6) revealed P200 (light shading) and N400 (dark shading), effects. Topographic maps reflect mean amplitudes to Well-known, Integration, and Novel facts at 250 ms (P200) and 400 ms (N400) post stimulus onset (Panel A). In response to the second 400 ms presentation of the stimuli, analysis of the centro-parietal cluster revealed dampened (and nonsignificant) P200 and N400 effects. The P600 (shaded area) differed for Novel relative to Well-known and Integration facts. Topographic maps reflect mean amplitudes at 750 ms post stimulus onset (Panel B). In both panels, ERP waveforms are from electrode site Cz. The raw EEG data were bandpass filtered with half-power cutoffs of 0.1 and 30 Hz; analyses were conducted on these data. For presentation purposes, the data were refiltered with half-power cutoffs of 0.1 and 10 Hz.
The second 400 ms presentation of the stimuli revealed change in the knowledge status of the newly-integrated information (Figure 4, Panel B). The P200 and N400 responses were less pronounced, presumably because all of the stimuli were more familiar (e.g., Van Petten et al., 1991). Neither the P200 nor the N400 differed by condition. In contrast, beginning 500 ms after stimulus onset and continuing through 900 ms, responses differed as a function of stimulus type: F(2, 58) = 6.72, p = .002, pη2 = .188. Responses to both Well-known and Integration facts were significantly smaller than those to Novel facts (ps = .009 and .034, respectively). Critically, responses to Well-known and Integration facts did not differ. The lower amplitude responses in the Well-known and Integration conditions are consistent with the suggestion that in both conditions, the initial retrieval attempt (on the first presentation) was successful, rendering the relevant material more readily accessible upon the second presentation, thus requiring less effortful processing of the stimulus the second time. In contrast, the Novel stimuli remained relatively unfamiliar, resulting in a high amplitude P600 (e.g., Besson et al., 1992; Olichney et al., 2000, 2002; Van Petten et al., 1991).
Additional support for the suggestion that the P600 reflected a change in knowledge status of the newly self-generated information over time comes from comparison of the amplitudes of the effect at the first versus the second presentation. As suggested by inspection of Figure 4, Panels a and b, across presentations, the amplitude of the response to the Integration facts decreased significantly, t(29) = 2.97, p = .006, indicating that fewer resources were devoted to processing of the stimuli on the second presentation. The same pattern was apparent for the Well-known stimuli: t(29) = 3.56, p < .001. In contrast, the amplitude of the response to the Novel facts did not differ between presentations: t(29) = −0.23, p = .817.
Discussion
The novel approach of using well-established indices of semantic and mnemonic processing as temporal markers of change revealed transition of the knowledge status of new information over time and experience. At the first presentation of the stimuli, neural responses to newly-derived Integration facts held an intermediate status that initially (200–300 ms) did not differ from either Well-known or Novel facts, but by 400 ms, were a category unto themselves. The second time the Integration facts were encountered, neural responses to them no longer differed from responses to Well-known facts (in any time window); responses to both Well-known and Integration facts differed from Novel facts (500–900 ms). A decrease in the amplitude of the P600 from Presentation 1 to Presentation 2 is further evidence of a change in knowledge status of the facts in the Integration condition.
Both the (a) change in pattern of responding from the P200 to the N400 (first presentation, when all of the sentences were new: none had been presented in the encoding phase); and (b) the change in pattern of responding in the P600 window on the first versus the second presentation, are consistent with the suggestion that new facts derived through integration had transitioned from novel to known. However, an alternative interpretation is that neural responses in the Integration condition were the result of priming. In this experiment, the sentence-final words to which ERPs were time-locked had appeared in the encoding phase. For example, the word “Taiga” in the integration fact “the world’s largest biome is the Taiga,” also had been presented in one of the pair of related sentences during the encoding phase: “the boreal forest is the Taiga.” This raises the possibility that the basis for the change in neural response was priming resulting from the prior exposure to the sentence-final word, rather than familiarity born of self-derivation through integration. We addressed this possibility in Experiment 1b.
Experiment 1b
Method
Subjects
Subjects were 21 adults (M = 19.8 years, range 18.6–23.1 years) drawn from the same population as Experiment 1a; none had participated in Experiment 1a. Subjects’ participation was recognized in the same way as in Experiment 1a.
Materials, Procedure, and Data Reduction
The sole change from Experiment 1a to 1b was to the nature of the 20 pairs of related facts presented during encoding; all other stimuli and test and data reduction procedures were identical. The encoding phase of Experiment 1a featured 20 pairs of related facts that when integrated with one another, permitted derivation of new Integration facts. In the present experiment, new sentences were developed that were related to the original stimuli, but which could not be combined with the original stimuli to derive the Integration facts (Pseudo-Integration facts). For example, in the pair of related sentences—“the world’s largest biome is the boreal forest” and “the boreal forest is the Taiga”—the second stem fact was replaced with “soil in the Taiga is mildly acidic.” The result was that the target words to be used in ERP testing appeared in the encoding sentences (as they had in Experiment 1a) but the pairs of related sentences did not support derivation of the Integration facts (i.e., when combined, the stem-fact sentences “the world’s largest biome is the boreal forest” and “soil in the Taiga is mildly acidic” do not support derivation of the novel integration fact that “the world’s largest biome is the Taiga”). Across subjects, sentences were used approximately equally often in the Pseudo-Integration and Novel-integration control conditions (a methodological precaution inadvertently neglected in Experiment 1a). The facts used in ERP testing were the same as in Experiment 1a. If the apparent transition from novel to well-known observed in Experiment 1a was the result of priming of the target word from exposure in the encoding phase, then the pattern of response to the Pseudo-Integration facts in Experiment 1b should be the same as that to the Integration facts in Experiment 1a.
Results and Discussion
Preliminary Analyses
As in Experiment 1a, separate analyses of variance for each ERP component (P200, N400, P600) and each presentation (first, second) revealed no statistically significant differences between the two types of novel control facts (Novel-non-integration and Novel-integration). Specifically, at the first presentation, the F(1, 20) values for the P200, N400, and P600 were 0.78, 2.49, and 1.39, respectively; p values = .39, .13, and .25, respectively. At the second presentation, the F(1, 20) values for the P200, N400, and P600 were 1.34, 0.11, and 1.99, respectively; p values = .26, .75, and .17, respectively. The two type of novel control facts were collapsed into the Novel category used in all analyses.
Main Analyses
The numbers of trials included in analyses by condition and presentation (first, second) are provided in Table 2, Panel A.
Table 2.
Experiment 1b: Descriptive Statistics (Mean, Standard Deviation, Range) for the Number of Trials Included in Analyses (Panel A) and Descriptive Statistics (Mean, Standard Deviation, Standard Error) for Mean Amplutides for the P200, N400, and P600 Event-Related Potential (ERP) Components for Each Condition for the Centro-parietal Cluster (P3/P4, CP1/CP2, CP5/CP6) (Panel B)
| ERP Component and Stimulus Condition | Presentation
|
|||||
|---|---|---|---|---|---|---|
| First | Second | |||||
| Panel A: Number of trials included in analyses
| ||||||
| M | SD | Range | M | SD | Range | |
|
| ||||||
| Well-known | 18.90 | 1.04 | 17–20 | 18.38 | 1.32 | 15–20 |
| Pseudo-Integration | 17.48 | 1.83 | 13–20 | 17.19 | 1.89 | 13–20 |
| Novel | 33.81 | 3.36 | 25–39 | 33.29 | 2.99 | 26–39 |
|
| ||||||
| Panel B: Mean amplitude (Amplitude μV)
| ||||||
| M | SD | StE | M | SD | StE | |
|
| ||||||
| P200
| ||||||
| Well-known | 4.03 | 3.21 | 0.70 | 2.42 | 2.15 | 0.47 |
| Pseudo-Integration | 3.22 | 2.93 | 0.64 | 2.78 | 2.43 | 0.53 |
| Novel | 1.81 | 2.66 | 0.58 | 2.42 | 2.43 | 0.53 |
|
| ||||||
| N400
| ||||||
| Well-known | 3.80 | 2.25 | 0.49 | 1.54 | 2.93 | 0.64 |
| Pseudo-Integration | 1.87 | 2.52 | 0.55 | 1.89 | 3.48 | 0.76 |
| Novel | 0.37 | 2.29 | 0.50 | 1.55 | 2.61 | 0.57 |
| P600 | ||||||
| Well-known | 4.73 | 3.30 | 0.72 | 2.31 | 2.20 | 0.48 |
| Pseudo-Integration | 5.23 | 3.57 | 0.78 | 3.97 | 3.25 | 0.71 |
| Novel | 4.41 | 3.07 | 0.67 | 4.45 | 2.89 | 0.63 |
As in Experiment 1a, neural activity differed in the Well-known and Novel fact conditions. However, the pattern of response to the Pseudo-Integration facts was different from that to the Integration facts in Experiment 1a. The pattern of response at the first presentation was consistent with a priming interpretation. In contrast, the pattern of response at the second presentation was not suggestive of a transition from novel to known, thereby eliminating priming as an explanation for that aspect of the pattern. Descriptive statistics for the P200, N400, and P600 ERP components at the centro-parietal cluster are provided in Table 2, Panel B.
At the first 400 ms presentation of the stimuli (Figure 5, Panel A), P200 and N400 effects were apparent: F(2, 40) = 8.443, p< .001, pη2 = .297, and F(2, 40) = 19.464, p < .001, pη2 = .493, respectively. As in Experiment 1a, the P200 was largest for Well-known facts, and differed significantly from the response to Novel facts (p = .002). The P200 to Pseudo-Integration facts was intermediate and differed from Novel (p = .033) but not from Well-known facts. Also as in Experiment 1a, the N400 revealed differences across all three conditions (ps < .032). Again as in Experiment 1a, amplitude in the P600 window did not differ by condition. Given the similarities in the patterns of responding to Pseudo-Integration (present experiment) and Integration (Experiment 1a) facts, we conclude that the change in response in the N400 relative to the P200 time window cannot be taken as evidence of a transition from novel to well-known but instead can be accounted for in terms of priming.
Figure 5.
In response to the first 400 ms presentation of the stimuli, analysis of the centro-parietal cluster (P3/P4, CP1/CP2, CP5/CP6) revealed P200 (light shading) and N400 (dark shading), effects (Panel A). In response to the second 400 ms presentation of the stimuli, analysis of the centro-parietal cluster revealed dampened P200 and N400 effects. The P600 (shaded area) differed for the Well-known relative to the Novel and Pseudo-Integration fact conditions which did not differ from one another. In both panels, ERP waveforms are from electrode site Cz. The raw EEG data were bandpass filtered with half-power cutoffs of 0.1 and 30 Hz; analyses were conducted on these data. For presentation purposes, the data were refiltered with half-power cutoffs of 0.1 and 10 Hz.
The second presentation of the stimuli revealed differentiation of the knowledge status of Well-known facts relative to those in the Pseudo-Integration and Novel conditions in the P600 window (Figure 5, Panel B). The pattern is not consistent with a priming explanation. The P200 and N400 responses were less pronounced, presumably because all of the stimuli were more familiar. Neither the P200 nor the N400 differed by condition.
The critical findings is that in the P600 window, the pattern was different from that observed in Experiment 1a. Examination of the main effect of condition, F(2, 40) = 5.073, p = .011, pη2 = .202, revealed significant differences between the Well-known and both the Pseudo-Integration (p = .026) and Novel (p = .008) fact conditions; the latter conditions did not differ from one another. Moreover, unlike in Experiment 1a, across presentations, the amplitude of the P600 in the Pseudo-Integration condition did not differ: t(20) = 1.21, p = .24. In contrast, the patterns for the Well-known and Novel conditions were the same as in Experiment 1a, such that a reduction in response amplitude was observed in the Well-known condition, t(20) = 4.01, p = .001, but not in the Novel condition, t(20) = −0.06, p = .95. Thus in this experiment, the facts in the Pseudo-Integration condition did not make the transition from novel to known. The pattern is inconsistent with interpretation of the P600 effect in Experiment 1a in terms of priming alone.
As a whole, the findings of Experiment 1 support the conclusion that facts newly derived through integration made a rapid transition from novel to known. Importantly, however, neither protocol in Experiment 1 provided direct evidence that subjects actually derived the integration facts based on exposure to the pairs of related facts in the encoding phase. Experiment 2 was a behavioral test of this question.
Experiment 2
Method
Subjects
Subjects were 14 adults (M = 20.6 years, range 18.9–21.7 years) drawn from the same population as Experiment 1; none had participated in Experiment 1. Subjects’ participation was recognized in the same way as in Experiment 1.
Materials, Procedure, and Data Reduction
The encoding stimuli were 120 sentences, selected from the same pool as in Experiment 1a. Ninety sentences conveyed information subjects were likely to have acquired at different ages. The remaining 30 sentences conveyed information that subjects were unlikely to know. Within this category were 10 pairs of related sentences (N = 20 sentences) which, when integrated with one another, permitted derivation of new facts. The other 10 sentences conveyed one-half of the information necessary to derive a new fact through integration. That is, during encoding, subjects read one member of the pair of sentences, but not both. Across subjects, sentences were used approximately equally often in the paired (both sentences presented) and single (one, but not both, sentences presented) conditions.
For the test phase, the stimuli were 40 sentences, none of which had been previously presented. Ten sentences conveyed facts that subjects were likely to know (Well-known). Ten sentences conveyed facts that subjects were unlikely to have known prior to the encoding phase, but which could be derived through integration of separate facts presented during encoding (Integration). The remaining 20 sentences served as controls (Novel). Ten of the control facts were unknown and could not be derived through integration of information from the encoding phase. The final 10 sentences conveyed facts derived through integration, but for which subjects had been exposed to only one of the requisite pair of facts during encoding. Because subjects had been exposed to only one member of the pair from which these novel facts were derived, they were not expected to self-generate them. Because preliminary analyses revealed no differences between the two types of control facts [t(13) = 0.62, p = .544], they were collapsed into the Novel category for analysis.
After encoding, subjects engaged in buffer tasks for the amount of time required to apply the ERP cap in Experiment 1 (~20 min). The buffer tasks were a number of demographic and background/experience questionnaires. Subjects then took part in the same protocol as in Experiment 1, with two important exceptions. First, no ERPs were recorded. Second, whereas subjects in Experiment 1 were presented with declarative sentences (Figure 1, Panel B), subjects in Experiment 2 were presented with questions created by omitting the final word of each sentence (Figure 1, Panel C). The final word was replaced by a blank screen, followed by a decision screen with four choices, one of which was correct. The distractors were conceptually plausible answers that represented the same part of speech as the correct answer, and had the same tense ending, if applicable. For example, for “the world’s largest biome is the ____,” the responses were Taiga (which was correct), Tundra, Xeric, and Alpine. Subjects made their selections using a game controller. Subjects were presented with each of 40 test sentences that conveyed facts that were Well-known (N = 10), derived through Integration (N = 10), and Novel (N = 20). Each word of each test sentence was presented for 400 ms, followed by an inter-stimulus interval of 200 ms.
Finally, as in Experiment 1, subjects were given the 40 facts on which they were tested (Well-known, Integration, Novel) and asked to indicate whether they knew each fact before the study. We eliminated from analysis Well-known facts of which subjects did not have prior knowledge, and Integration and Novel facts of which they had prior knowledge (M = 7% facts eliminated per subject).
Results and Discussion
Figure 6 depicts the mean number of correct responses in the Well-known, Integration, and Novel conditions. Performance differed across conditions: F(2, 26) = 59.81, p < .001, pη2 = .821. Performance on Well-known facts was virtually flawless, whereas performance on the Novel facts did not differ from chance (25%). Subjects’ answered 56% of the Integration fact questions correctly. Performance in all three conditions differed reliably (Tukey, p < .01). Given that the subjects had not encountered any of the facts in the encoding phase, differential performance in the Integration and Novel conditions was due to the opportunity or lack thereof (respectively) for integration and thus self-derivation as a source of knowledge.
Figure 6.

Mean number of correct responses in the Well-known, Integration, and Novel conditions of behavioral testing in Experiment 2 (error bars reflect standard error).
Experiment 2 provides evidence that based on exposure to two related sentences, subjects derived a substantial number of integration facts. In contrast, based on exposure to only one of the sentences, recognition performance did not differ from chance. This lends strong support to the interpretation that the subjects in Experiment 1a derived new facts through integration and that the newly derived information rapidly achieved the status of known.
General Discussion
The present research afforded observation of the rapid transition of newly self-derived information from novel to known. In two experiments, subjects read large numbers of sentences, some of which featured facts they knew and others of which featured facts that were novel. The critical manipulation was that some of the novel facts could be integrated with one another to support self-derivation of new knowledge. At test, subjects were presented with entirely new sentences. Again, some sentences featured facts that subjects were expected to know whereas others were novel. Importantly, some of the novel facts could have been derived by the subjects through integration of related facts.
In the ERP Experiments 1a and 1b, there were three potential indices of the knowledge status of the new facts that subjects could potentially have derived through integration of related facts, namely, the P200 (150–300 ms after stimulus onset), the N400 (300–500 ms after stimulus onset), and the P600 (500–900 ms after stimulus onset). Each of the indices was examined in response to an initial and a second 400 ms presentation of the stimuli. In Experiment 1a, upon the initial presentation, the response to Integration stimuli changed between the P200 and the N400. The change was consistent with the suggestion that the Integration facts were assuming a status of “known.” Importantly, the priming control implemented in Experiment 1b cast doubt on that interpretation: a similar pattern of change in response was observed to Pseudo-Integration facts that did not support new knowledge derivation.
In contrast to the activity in the P200 and N400 windows, compelling evidence of the rapid transition of new information from novel to known was evident in the P600 window. In both Experiments 1a and 1b, upon the initial 400 ms presentation of the stimuli, positive, high amplitude responses were observed in all three conditions (Well-known, Integration/Pseudo-Integration, Novel). The responses could reasonably be interpreted as evidence of attempts to retrieve relevant contents from long-term memory (e.g., Van Petten et al., 1991). Importantly, upon the second 400 ms presentation of the stimuli, P600 responses differed across experiments and stimulus categories. In Experiment 1a, in which integration was possible, amplitudes were lower on the second relative to the first presentation of both the Well-known and the Integration stimuli; P600 amplitude in the two conditions did not differ, and both were different from the Novel condition (in which amplitudes did not decrease across presentations). In contrast, in Experiment 1b, the pattern of response to the Pseudo-Integration stimuli looked different from the pattern to Well-known stimuli; it was parallel to the response to Novel stimuli. Together, the ERP experiments support the interpretation that information newly derived through integration made a rapid transition from novel to known. Experiment 2 provided support for the interpretation that the basis for the rapid transition of information from novel to known was self-derivation of new knowledge through integration.
The present research provided important new information indicative of rapid transition of information from newly self-derived to known. It also raised new questions about the processes involved and the nature of the resulting products. Perhaps chief among the process questions is the role of priming in new knowledge generation. Comparison of the results of Experiments 1a and 1b strongly suggests that priming was at least partially responsible for the neural responses in the P200 and N400 windows (150–300 ms and 300–500 ms after stimulus onset, respectively). The response patterns in these windows were highly similar in the two experiments, even though self-generation of new knowledge through integration of related facts was possible only in Experiment 1a. In contrast, between-experiment differences in the response patterns in the P600 window indicate that priming was not the source of the apparent transition of the newly-derived information from novel to know: the P600s to Integration and Well-known facts (Experiment 1a) were similar whereas the P600s to Pseudo-Integration and Well-known facts (Experiment 1b) were different. Elsewhere, we have argued that the self-generative process of derivation of new knowledge through integration involves two steps (Bauer et al., in press; Bauer & Varga, in press; Varga & Bauer, 2013). The first step is integration of the separate but related facts that are the platform from which new knowledge is derived; the presumed basis for integration is shared or common elements (e.g., mention of “the Taiga”). Integration of the separate units of information is necessary—but not sufficient—for the second step, which involves derivation of new information from the newly-integrated product. It is reasonable to assume that the spreading activation that is responsible for priming also is involved in the first of these steps, namely, integration of separate units based on shared elements. The additional processes associated with self-generation of new information seemingly unfold over a longer period of time, as reflected in amplitude in the 500–900 ms time window of the P600. Additional research will be required to further explicate the processes involved in self-generation of new knowledge through integration, and the time course over which the processes unfold.
The present research also raised questions about the nature of the mnemonic products resulting from integration of separate yet related units of information. A salient question in this regard is whether the newly-derived information persists in memory over time. As noted earlier, in some cases, self-generated information may be short-lived, surviving in working memory only long enough to ensure comprehension of text, for example (e.g., McKoon & Ratcliff, 1992; Yule, 2010). For newly self-generated information to become integrated into the knowledge-base, it must endure over time. Though the question has yet to be addressed in an adult sample, there is evidence that children 6 years of age retain newly self-generated information, at least over a 1-week delay. Specifically, in Varga and Bauer (2013), children 6 years of age were provided with separate passages of text that when integrated with one another, supported self-generation of new knowledge (i.e., Integration facts). When tested later in the session in which they heard the stories, the children produced the integration facts on 63% of the trials. When tested again 1 week later, they showed no evidence of forgetting: they recalled the facts on 62% of trials. It will be left to future research to determine whether adults also retain newly self-generated information over time.
The present research afforded a unique window on the broadly productive nature of semantic memory. It provided evidence that in addition to operations such as analogy, deduction, and induction, semantic memory can be extended through self-generation of new knowledge derived through integration of separate sources of information (see also Bauer et al., 2012, in press; Bauer & San Souci, 2010; Varga & Bauer, 2013). Moreover, it permitted observation of the rapid time course over which these productive processes unfold. The speed with which newly self-derived information is treated as well-known helps explain the richness of our knowledge base. We suggest that it also may help explain why semantic knowledge exists seemingly without reference to the time or place it was acquired. In contrast to episodic memory, entries of which are located in temporal and spatial context, semantic memories are timeless and placeless (Tulving, 1983). One explanation of the absence of time and place tags in semantic memory is that these differentiating features are forgotten after experience of the events that gave rise to the content. The present research suggests another potential explanation, namely, that some of the information in semantic memory never carried tags as to its origin because it was never directly experienced—we generated it ourselves. Based on the present research, we cannot test this hypothesis directly. Though we asked subjects whether they knew the test facts before beginning the experiment (and we eliminated any Integration or Novel facts for which subjects indicated prior knowledge), we did not ask subjects to indicate where or when they learned the newly-derived Integration facts, or whether they were aware of having self-generated them in the context of the experiment. Nor did we ask whether subjects had a subjective sense of familiarity of the information. In future research it would be informative to address these questions directly, thereby informing the phenomenological status of newly self-generated information.
In conclusion, the present research provided a unique perspective on the process of elaboration of semantic memory. ERPs revealed rapid transition of information from the status of newly self-derived to known. This process helps to explain the broadly productive nature of semantic memory. Though we could not address the question in the present research, we suggest that it also may account for the absence of episodic information specifying when and where its contents were acquired.
Acknowledgments
This research was supported by NICHD HD57291 and HD67359. We also thank Jessica King and Manas Winfield for help with data collection, Jacqueline Leventon for help with data reduction and analysis, other members of the Memory at Emory research group, and the participants.
References
- Bauer PJ, King JE, Larkina M, Varga NL, White EA. Characters and clues: Factors affecting children’s extension of knowledge through integration of separate episodes. Journal of Experimental Child Psychology. 2012;111:681–694. doi: 10.1016/j.ecp.2011.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bauer PJ, San Souci P. Going beyond the facts: Young children extend knowledge by integrating episodes. Journal of Experimental Child Psychology. 2010;107:452–465. doi: 10.1016/j.jecp.2010.05.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bauer PJ, Varga NL. The developmental cognitive neuroscience of memory: Implications for education. In: Tardif E, Doudin P-A, editors. Cognitive Neuroscience and Education. Brussels Press; (in press) To appear. [Google Scholar]
- Bauer PJ, Varga NL, King JE, Nolen AM, White EA. Semantic elaboration through integration: Hints both facilitate and inform the process. Journal of Cognition and Development. doi: 10.1080/15248372.2013.849707. (in press) [DOI] [Google Scholar]
- Besson M, Kutas M. The many facets of repetition: A cued-recall and event-related potential analysis of repeating words in same versus different sentence contexts. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1993;19:1115–1133. doi: 10.1037/0278-7393.19.5.1115. [DOI] [PubMed] [Google Scholar]
- Besson M, Kutas M, Van Petten C. An event-related potential (ERP) analysis of semantic congruity and repetition effects in sentences. Journal of Cognitive Neuroscience. 1992;4:132–149. doi: 10.1162/jocn.1992.4.2.132. [DOI] [PubMed] [Google Scholar]
- Coulson S, King JW, Kutas M. Expect the unexpected: Event-related brain response to morphosyntactic violations. Language and Cognitive Processes. 1998;13:21–58. doi: 10.1080/016909698386582. [DOI] [Google Scholar]
- Delorme A, Makeig S. EEGLAB: an open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. Journal of Neuroscience Methods. 2004;134:9–21. doi: 10.1016/j.jneumeth.2003.10.009. [DOI] [PubMed] [Google Scholar]
- Evans KM, Federmeier KD. The memory that’s right and the memory that’s left: Event-related potentials reveal hemispheric asymmetries in the encoding and retention of verbal information. Neuropsychologia. 2007;45:1777–1790. doi: 10.106/j.neuropsychologia.2006.12.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Federmeier KD, Mai H, Kutas M. Both sides get the point: Hemispheric sensitivities to sentential constraint. Memory & Cognition. 2005;33:871–886. doi: 10.3758/BF03193082. [DOI] [PubMed] [Google Scholar]
- Federmeier KD, Laszlo S. Time for meaning: Electrophysiology provides insights into the dynamics of representation and processing in semantic memory. In: Ross BH, editor. Psychology of Learning & Motivation. Vol. 51. Burlington: Academic Press; 2009. pp. 1–44. [DOI] [Google Scholar]
- Gentner D. Structure-mapping: A theoretical framework for analogy. Cognitive Science. 1983;7:155–170. doi: 10.1016/S0364-0213(83)80009-3. [DOI] [Google Scholar]
- Hagoort P. How the brain solves the binding problem for language: A neurocomputational model of syntactic processing. Neuroimage. 2003;20:S18–S29. doi: 10.1016/j.neuroimage.2003.09.013. [DOI] [PubMed] [Google Scholar]
- Hagoort P, Brown CM, Osterhout L. The neurocognition of syntactic processing. In: Brown CM, Hagoort P, editors. The neurocognition of language. Oxford, UK: Oxford University Press; 1999. pp. 273–316. [Google Scholar]
- Holland JH, Holyoak KJ, Nisbett RE, Thagard RR. Induction: Processes of inference, learning, and discovery. Cambridge, MA: Cambridge University Press; 1989. [Google Scholar]
- Jasper HH. The ten-twenty electrode system of the International Federation. Electroencephalography & Clinical Neurophysiology. 1958;10:371–375. [PubMed] [Google Scholar]
- Johnson-Laird PN. How we reason. Oxford, UK: Oxford University Press; 2006. [Google Scholar]
- Johnson-Laird PN, Byrne RMJ. Deduction. Hillsdale, NJ: Erlbaum; 1991. [Google Scholar]
- Kutas M, Federmeier KD. Electrophysiology reveals semantic memory use in language comprehension. Trends in Cognitive Sciences. 2000;4:463–470. doi: 10.1016/S1364-6613(00)01560-6. [DOI] [PubMed] [Google Scholar]
- Kutas M, Federmeier KD. Thirty years and counting: Finding meaning in the N400 component of the event-related brain potential (ERP) Annual Review of Psychology. 2011;62:621–647. doi: 10.1146/annurev.psych.093008.131123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kutas M, Hillyard SA. Reading senseless sentences: Brain potentials reflect semantic incongruity. Science. 1980;207:203–205. doi: 10.1126/science.7350657. [DOI] [PubMed] [Google Scholar]
- Lelekov-Boissard T, Dominey PF. Human brain potentials reveal similar processing of non-linguistic abstract structure and linguistic syntactic structure. Clinical Neurophysiology. 2002;32:72–84. doi: 10.1016/s0987-7053(01)00291-x. [DOI] [PubMed] [Google Scholar]
- McKoon G, Ratcliff R. Inferences during reading. Psychological Review. 1992;99:440–466. doi: 10.1037/0033-295X.99.3.440. [DOI] [PubMed] [Google Scholar]
- Moshman D. From inference to reasoning: The construction of rationality. Thinking & Reasoning. 2004;10:221–239. doi: 10.1080/13546780442000024. [DOI] [Google Scholar]
- Olichney JM, Morris SK, Ochoa C, Salmon DP, Thal LJ, Kutas M, Iragui VJ. Abnormal verbal event related potentials in mild cognitive impairment and incipient Alzheimer’s disease. Journal of Neurology, Neurosurgery, & Psychiatry. 2002;73:377–384. doi: 10.1136/jnnp.73.4.377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Olichney JM, Van Petten S, Paller KA, Salmon DP, Iragui VJ, Kutas M. Word repetition in amnesia: Electrophysiological measures of impaired and spared memory. Brain. 2000;123:1948–1963. doi: 10.1093/brain/123.9.1948. [DOI] [PubMed] [Google Scholar]
- Osterhout L, Holcomb PJ. Event-related potentials and syntactic anomaly: Evidence of anomaly detection during the perception of continuous speech. Language and Cognitive Processes. 1993;8:413–437. doi: 10.1080/01690969308407584. [DOI] [Google Scholar]
- Patel AD, Gibson E, Ratner J, Besson M, Holcomb PJ. Processing syntactic relations in language and music: An event-related potential study. Journal of Cognitive Neuroscience. 1998;10:717–733. doi: 10.1162/jocn.1998.10.2.717. [DOI] [PubMed] [Google Scholar]
- Rugg MD, Roberts RC, Potter DD, Pickles CD, Nagy ME. Event-related potentials related to recognition memory: Effects of unilateral temporal lobectomy and temporal lobe epilepsy. Brain. 1991;114:2313–2332. doi: 10.1093/brain.114.5.2313. [DOI] [PubMed] [Google Scholar]
- Rugg MD, Mark RE, Walla P, Schloerscheidt AM, Birch CS, Allan K. Dissociation of the neural correlates of implicit and explicit memory. Nature. 1998;392:595–598. doi: 10.1038/33396. [DOI] [PubMed] [Google Scholar]
- Tulving E. Elements of episodic memory. Oxford, UK: Oxford University Press; 1983. [Google Scholar]
- Van Petten C, Coulson S, Rubin S, Plante E, Parks M. Time course of word identification and semantic integration in spoken language. Journal of Experimental Psychology: Learning, Memory, and Cognition. 1999;25:394–417. doi: 10.1037/0278-7393.25.2.394. [DOI] [PubMed] [Google Scholar]
- Van Petten C, Kutas M, Kluender R, Mitchiner M, McIsaac H. Fractionating the word repetition effect with event-related potentials. Journal of Cognitive Neuroscience. 1991;3:131–150. doi: 10.1162/jocn.1991.3.2.131. [DOI] [PubMed] [Google Scholar]
- Varga NL, Bauer PJ. Effects of delay on 6-year-old children’s self-generation and retention of knowledge through integration. Journal of Experimental Child Psychology. 2013;115:326–341. doi: 10.1016/j.jecp.2013.01.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wlotko EW, Federmeier KD. Finding the right word: Hemispheric asymmetries in the use of sentence context information. Neuropsychologia. 2007;45:3001–3014. doi: 10.1016/j.neuropsychologia.2007.05.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yule G. The study of language. Cambridge, UK: Cambridge University Press; 2010. [Google Scholar]