

Abstract
Hydrogen peroxide (H2O2) is an endothelium-derived hyperpolarizing factor. Since opposing vasoactive effects have been reported for H2O2 depending on the vascular bed and experimental conditions, this study was performed to assess whether H2O2 acts as a vasodilator in the rat mesenteric artery and, if so, to determine the underlying mechanisms. H2O2 elicited concentration-dependent relaxation in mesenteric arteries precontracted with norepinephrine. The vasodilatory effect of H2O2 was reversed by treatment with dithiothreitol. H2O2-elicited vasodilation was significantly reduced by blocking 4-aminopyridine (4-AP)-sensitive Kv channels, but it was resistant to blockers of big-conductance Ca2+-activated K+ channels and inward rectifier K+ channels. A patch-clamp study in mesenteric arterial smooth muscle cells (MASMCs) showed that H2O2 increased Kv currents in a concentration-dependent manner. H2O2 speeded up Kv channel activation and shifted steady state activation to hyperpolarizing potentials. Similar channel activation was seen with oxidized glutathione (GSSG). The H2O2-mediated channel activation was prevented by glutathione reductase. Consistent with S-glutathionylation, streptavidin pull-down assays with biotinylated glutathione ethyl ester showed incorporation of glutathione (GSH) in the Kv channel proteins in the presence of H2O2. Interestingly, conditions of increased oxidative stress within MASMCs impaired the capacity of H2O2 to stimulate Kv channels. Not only was the H2O2 stimulatory effect much weaker, but the inhibitory effect of H2O2 was unmasked. These data suggest that H2O2 activates 4-AP-sensitive Kv channels, possibly through S-glutathionylation, which elicits smooth muscle relaxation in rat mesenteric arteries. Furthermore, our results support the idea that the basal redox status of MASMCs determines the response of Kv currents to H2O2.
Electronic supplementary material
The online version of this article (doi:10.1007/s00424-014-1513-3) contains supplementary material, which is available to authorized users.
Keywords: H2O2, Kv channel, Mesenteric artery, S-glutathionylation, Oxidative stress
Introduction
Reactive oxygen species (ROS) are detrimental to biological processes and contribute to disease conditions such as inflammation, ischemia–reperfusion injury, atherosclerosis, diabetes mellitus, and hypertension. However, some ROS like hydrogen peroxide (H2O2) may act as physiological signaling molecules and contribute to biologically beneficial processes [34, 72].
H2O2 has been suggested to be an endothelium-derived hyperpolarizing factor (EDHF) [69], a major component of endothelium-dependent relaxation in resistance-sized arteries [70]. The cellular and molecular mechanisms by which H2O2 elicits vasodilation remain to be determined, although smooth muscle hyperpolarization seems to be required [58]. The vascular smooth muscle cells functionally express four different K+ channels [4, 33, 43, 45, 49], namely, voltage-gated (Kv), Ca2+-activated (KCa), ATP-sensitive (KATP), and inward rectifier K+ (Kir) channels. The K+ channels are thought to play an important role in maintaining the membrane potential of vascular myocytes [45] and have been implicated in H2O2-induced smooth muscle relaxation. However, there is no general agreement on the effects of H2O2 on K+ channels in smooth muscle. Several studies have identified KCa channels as putative targets that are activated in the process of H2O2-induced vasodilation [81], while some other groups indicate that H2O2 induced a vasorelaxation through opening of ATP-dependent K+ (KATP) channels [74]. The Kv channel is also suggested to be a target of H2O2, which activates Kv channels in coronary and pulmonary vascular smooth muscles [44, 54, 57]. However, Kv channels are inhibited by H2O2 in ductus arteriosus smooth muscles [2]. Studies using cloned Kv1.5, a major component of Kv current in coronary arteries, show that H2O2 increases Kv1.5 current for voltages < +20 mV but decreases it for high depolarizing voltages [12]. It is still uncertain whether H2O2 acts as a vasodilator. Studies have reported H2O2 as a vasoconstrictor [30, 67] and vasodilator [29, 73], or both [18, 38]. These differences may depend on experimental design and the specific vascular bed or vessel being studied [42, 47].
H2O2 is a small stable molecule carrying no charge, which allows it to readily cross membranes and travel freely to targets within cells [25]. Various cellular modifications occur with the increase in H2O2, and increasing evidence suggests that S-glutathionylation predominates in myocytes because glutathione (GSH) is the most abundant, low molecular mass, reducing equivalent [59]. H2O2 is capable of oxidizing the thiol groups of cysteine residues to form disulfide bonds with GSH (S-glutathionylation). S-glutathionylated proteins result from thiol/disulfide exchange between protein thiols and oxidized form of glutathione (GSSG) or S-glutathionylated protein. Modulation of protein activity by S-glutathionylation is a newly recognized posttranslational regulatory mechanism [20, 21]. This process results in major changes to protein conformation and function [22]. The Kir6.1/SUR2B channel and Cav1.2 channel are subject to S-glutathionylation induced by H2O2 [63, 71, 78].
In a previous study, we demonstrated that in mesenteric arteries, the 4-aminopyridine (4-AP)-sensitive Kv currents play a critical role in the regulation of smooth muscle resting membrane potential (Em) and vascular tone [3, 66]. In this study, we examined the hypothesis that H2O2 relaxes rat mesenteric arteries by S-glutathionylation-dependent activation of 4-AP-sensitive Kv channels. We performed studies using an approach of combined molecular biology, electrophysiology, and isometric organ chamber mechanics. Our results show that H2O2 enhanced the activity of 4-AP-sensitive Kv channels, possibly through S-glutathionylation, leading to vasorelaxation in the mesenteric artery. We also present evidence to show that Kv channels under conditions of persistent oxidative stress were not activated, but rather inhibited by the addition of H2O2, suggesting that H2O2 may act as a vasoconstrictor under certain pathological conditions.
Methods
Tissue and cell preparation
Male Sprague–Dawley (SD) rats (9–11 weeks old) were used for the experiments. All experiments were conducted in accordance with the National Institutes of Health guidelines for the care and use of animals, and the Institutional Animal Care and Use Committee of Konkuk University approved this study. Rats were euthanized by exposure to a rising concentration of carbon dioxide or exsanguinated by cutting the carotid arteries under deep ketamine–xylazine anesthesia. Single-cell suspensions of mesenteric arterial smooth muscle cells (MASMCs) were prepared as described previously [3]. Briefly, the second- to fourth-order branches of the superior mesenteric arteries were carefully removed and placed in normal Tyrode (NT) solution (143 mM NaCl, 5.4 mM KCl, 0.33 mM NaH2PO4, 1.8 mM CaCl2, 0.5 mM MgCl2, 5 mM hydroxyethyl piperazineethanesulfonic acid (HEPES), and 11 mM glucose, adjusted to pH 7.4 with NaOH). The arteries were cut into small pieces and transferred to a digestion solution. The tissue was first digested for 15 min in Ca2+-free normal NT solution containing 1 mg/mL papain (Sigma Chemical, St. Louis, MO, USA), 1 mg/mL bovine serum albumin, and 1 mg/mL dithiothreitol. The nominally Ca2+-free NT was prepared by omitting 1.8 mM CaCl2 from the NT solution. Subsequently, the tissue sample was incubated for 25 min in a second digestion solution, in which 3 mg/mL collagenase (Wako, Osaka, Japan) replaced papain. Following enzyme treatment, the cells were isolated by gentle agitation with a fire-polished glass pipette in Ca2+-free NT solution.
Solutions and drugs
NT was used as the bathing solution for the patch-clamp experiments. The pipette internal solution contained 140 mM KCl, 5 mM NaCl, 5 mM MgATP, 10 mM HEPES, and 10 mM 1,2-bis(aminophenoxy)ethane-N,N,N′,N′-tetraacetic acid, adjusted to pH 7.2 with KOH. Bicarbonate-buffered physiological salt solution (PSS) was used as the bath solution for the organ chamber mechanics experiments. The PSS was composed of 136.9 mM NaCl, 5.4 mM KCl, 1.5 mM CaCl2, 1.0 mM MgCl2, 23.8 mM NaHCO3, and 0.01 mM EDTA. All chemicals, including H2O2, GSH, and GSSG, were purchased from Sigma.
Electrophysiological recordings
We used the conventional whole-cell configuration of the patch-clamp technique [3] to record membrane currents and Em. EPC8 (HEKA, Mahone Bay, Nova Scotia, Canada) patch-clamp amplifier with a DAQPad-6070E interface (National Instrument, Austin, TX, USA) or an Axopatch 200B patch-clamp amplifier with a DigiData 1200 interface (Axon Instruments, Foster City, CA, USA) was used. Data were digitized with custom-built software (R-clamp, by Dr. SY Ryu) or with pClamp6 software (Axon Instruments) at a sampling rate of 1–10 kHz. The data were low-pass filtered at 1 kHz and saved for analysis. Voltage pulse generation was also controlled by R-clamp software and pClamp6. Patch pipettes were pulled from borosilicate capillary tubes (Clark Electromedical Instruments, Pangbourne, UK) using a puller (PP-83; Narishige, Tokyo, Japan). We used patch pipettes with a resistance of 2–4 MΩ when filled with the abovementioned pipette solution. Recordings were started at least 7 min after establishing the whole-cell configuration to allow adequate cell dialysis of the pipette solution. All experiments were carried out at room temperature (20–25 °C).
Kv currents were elicited by depolarizing steps between −60 and +50 mV (200 ms duration) from a holding potential of −70 mV. Tetraethylammonium (TEA, 1 mM) was added to all bath solutions during the patch-clamp study to prevent activation of big-conductance KCa (BKCa) channels. Additionally, specific activation of the Kv current was confirmed using 4-AP at the end of the experiments. Conductance–voltage (G–V) relationships were plotted using steady state current amplitudes divided by driving force (Em–Erev, where Erev is the reversal potential of the Kv current). The normalized conductance was fit using Origin 6.0 software to the Boltzmann equation.
Organ chamber isometric contraction measurements
The mesenteric arterial rings were mounted vertically on two L-shaped stainless steel wires in a 3-mL tissue chamber for the tension measurements. One wire was attached to a micromanipulator and the other to an isometric force transducer (FT03; Grass, West Warwick, RI, USA). Changes in isometric force were digitally acquired at 1 Hz with a PowerLab data acquisition system (AD Instruments, Colorado Springs, CO, USA). Resting tension was set to 1 g using the micromanipulator. After a 60-min equilibration under resting tension in a tissue chamber filled with PSS, the rings were sequentially exposed to 70 mM KCl–PSS (10 min) and PSS (15 min) three times for stabilization. The high KCl (70 mM)–PSS was prepared by replacing NaCl with equimolar KCl in PSS. Bath solutions were thermostatically controlled at 37 °C and were continuously saturated with a mixture of 95 % O2 in 5 % CO2 to achieve pH 7.4.
Western blot
Primary cultures of rat MASMCs between six and ten passages were used for Western blot. MASMCs were isolated from SD rats and cultured in Dulbecco’s modified Eagle’s medium (DMEM) containing 10 % fetal bovine serum (FBS) and 1 % penicillin–streptomycin. The cells were grown to 80 % confluence and starved in DMEM without FBS for 12 to 24 h prior to experiments. After starvation, cells were treated with H2O2 for 10 min at 37 °C. The cells were then washed twice with phosphate-buffered saline (PBS) and lysed using RIPA buffer (TNT Research, Seoul, South Korea). Samples were run on an 8 % SDS–polyacrylamide nonreducing gel and then transferred to a polyvinylidene fluoride (PVDF) membrane (Millipore, Bedford, MA, USA). Rabbit primary antibodies against Kv1.2, Kv1.5, and Kv 2.1 (1:500; Alomone Lab) and secondary antibodies conjugated with horseradish peroxidase were used in the Western blot (1:2,000; Cell Signaling Technology, Danvers, MA, USA). Signals were visualized using Las-4000 (Fuji Film, Tokyo, Japan).
Streptavidin pull-down assay
The culture medium was replaced with fresh medium 2 h before experiments. Biotinylated glutathione ethyl ester (BioGEE; 100 μM; Invitrogen, Carlsbad, CA, USA) was added to the medium and incubated for 1 h, followed by H2O2 (0.1 or 10 mM) challenges for 10 min. Biotin–GSH-conjugated proteins were pulled down using Dynabeads streptavidin according to the methods provided by Invitrogen. Dynabeads streptavidin was washed thrice with PBS before conjugation with biotin. Samples were then mixed with beads and incubated at room temperature with gentle rotation for 30 min. A magnet was used to separate the biotinylated molecule–bead complex. The supernatant containing unlabeled proteins was discarded, and the pellet was resuspended, followed by washes with PBS. The biotinylated molecule–bead complex was boiled with loading buffer for 7 min for Western blotting.
Data analysis
The Origin 6.0 software (Microcal Software, Inc., Northampton, MA, USA) was used for data analysis. Activation kinetics was calculated by fitting the data to a single exponential decay function. The time course of current inactivation was also fit to a single exponential function. The results are shown as mean ± standard error. Paired or independent Student’s t tests were used to test for significance, and p < 0.05 was regarded as significant. We performed one-way repeated measures ANOVA and Holm–Sidak test in order to examine the statistical significance of data shown in Fig. 3b and one-way ANOVA and Tukey’s test for Fig. 5c using SigmaPlot 12.5.
Fig. 3.

H2O2 shifts G–V curves for Kv channels to the left and speeds up channel activation. a G–V curves for KV channels, before and after H2O2 (0.1–10 mM) treatment. Smooth curves were fitted using the Boltzmann function. b V 1/2 obtained from the Boltzmann function is plotted against H2O2 concentration. One-way repeated measures ANOVA and Holm–Sidak test (α = 0.01, p < 0.05). Statistically distinct groups are indicated by letters. c Representative traces at +40 mV show that H2O2 (10 mM) accelerates Kv current activation. Activation of KV currents was fit with a single exponential function. Fits to original traces and time constant (τ) values are shown. d Time constants are presented as a function of the pulse potential in control (filled black square) and following application of 10 mM H2O2 (filled red circle) (*p < 0.05; **p < 0.01; ***p < 0.001 vs. control)
Fig. 5.
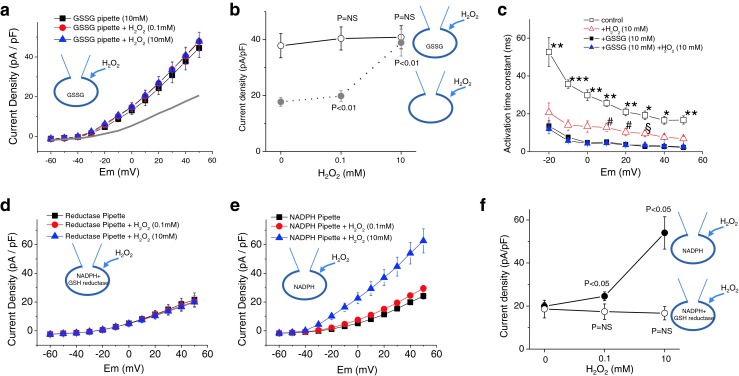
GSSG increases Kv currents in rat MASMCs. a Effects of adding GSSG in the pipette solution on Kv currents and on the action of H2O2. Averaged I–V curves before (filled black square) and after bath application of 0.1 mM H2O2 (filled red circle) or 10 mM H2O2 (filled blue triangle) in cells with GSSG in the pipette are shown. Gray line indicates the averaged I–V relationship with control pipette solution (redrawn from Fig. 2b for comparison). H2O2 was added cumulatively (n = 12). Note that Kv currents with pipette solution containing GSSG (filled black square) are much larger than those with control pipette solution (gray line). b Comparative data of Kv current densities measured at +40 mV for H2O2 effects in control pipette solution (filled gray circle; n = 21) with that in the pipette containing 10 mM GSSG (empty circle; n = 12). p < 0.01 vs. basal condition (control pipette solution); p = NS vs. basal condition (GSSG in the pipette). c Time constants recorded with GSSG pipette are presented as a function of the pulse potential before (filled black square) and after application of 10 mM H2O2 (filled blue triangle) (n = 8). Time constants recorded with control pipette (empty square and empty triangle) are redrawn from Fig. 3d for comparison (n = 16). One-way ANOVA and Tukey’s test, *p < 0.05; **p < 0.01; ***p < 0.001 vs. all other groups; # p < 0.05 vs. GSSG pipette groups; § p < 0.05 vs. GSSG alone. d, e Effects of GSH reductase (0.2 units/mL) plus NADPH (d) on Kv currents and on the action of H2O2. NADPH alone had no effect (e). Averaged I–V curves before (filled black square) and after bath application of 0.1 mM H2O2 (filled red circle) and 10 mM H2O2 (filled blue triangle) are shown. f Comparative data between Kv current densities measured at +40 mV for H2O2 effects in the pipette containing NADPH alone (filled black circle; n = 11) and those in the pipette containing NADPH plus GSH reductase (empty circle; n = 11). p = NS vs. basal condition (NADPH plus GSH reductase)
Results
H2O2 causes relaxation of the precontracted mesenteric arterial rings by redox-dependent alterations
We used isometric organ chamber mechanics to examine whether H2O2 relaxes rat mesenteric arteries. We used arterial rings without intact endothelium. The arterial rings were precontracted with norepinephrine (NE) (1 μM). H2O2 induced a concentration-dependent relaxation in the precontracted mesenteric arterial rings (Fig. 1a, c, d). We determined whether a thiol-specific reducing agent, DL-dithiothreitol (DTT), could reverse H2O2-induced relaxation. Pretreatment with 1 mM DTT almost completely prevented the relaxation by 1 mM H2O2 (Fig. 1b, c). The addition of DTT to vascular rings, in the absence of contractile agonist, did not affect resting tension (0.23 ± 0.19 vs. 0.22 ± 0.19 g before and after adding 1 mM DTT, respectively; n = 8). These data indicate that the effect of DTT is specific for H2O2-induced relaxation and suggest that thiol groups in smooth muscle are targets of H2O2 signaling. TEA was used to assess the contribution of the K+ channels to H2O2-induced smooth muscle relaxation. At a concentration of 1 mM, TEA is reported to be relatively specific for BKCa channels and has little effect on voltage-dependent K+ channels [65]. One millimolar TEA (Fig. 1d; n = 6) did not significantly inhibit the relaxation caused by H2O2 (compared with control; n = 6). BaCl2 (100 μM), a blocker of Kir, did not affect the H2O2-induced relaxation either (Fig. 1d). We attempted similar experiments with 10 mM 4-AP, a known Kv channel blocker; 4-AP significantly attenuated H2O2-induced relaxation (Fig. 1d, n = 6). These data suggest that 4-AP-sensitive Kv channels mediate the vasodilation by H2O2 in the mesenteric artery.
Fig. 1.

Effects of H2O2 on NE-precontracted mesenteric arterial rings under control and DTT-pretreated conditions. a Isometric tension recordings showing the effects of increasing concentrations of H2O2 (0.1–10 mM) on mesenteric arteries precontracted with NE. b Pretreatment with DTT inhibited the H2O2-induced relaxation. c H2O2-induced relaxation under control and DTT-pretreated conditions. The H2O2 effect was blocked by DTT (*p < 0.05 vs. control condition). d Summary of the effects of various potassium current blockers on the H2O2-induced relaxation (***p < 0.001 vs. control)
Effects of H2O2 on Kv currents in rat MASMCs
To examine whether H2O2 activated 4-AP-sensitive Kv currents, we recorded Kv currents using the conventional whole-cell, patch-clamp technique with depolarizing voltage steps as described previously [3] (Fig. 2). Cells were held at −70 mV to remove voltage-dependent channel inhibition, and membrane potential was stepped from −60 to +50 mV in 10-mV increments. Cumulative application of H2O2 superfusion (5 min at each concentration) increased Kv currents (Fig. 2a). Addition of 10 mM 4-AP to the bath reduced current magnitude below the baseline level, indicating that 4-AP-sensitive Kv channels are responsible for the outward current. H2O2-induced Kv current modulation was concentration-dependent; 0.1, 1, and 10 mM H2O2 induced an increase of Kv current amplitude at +40 mV to 11.4 ± 2.9, 47.5 ± 7.7, and 127.4 ± 23.8 %, respectively (Fig. 2b, c). Analysis of I–V relationship also indicated that the effect of H2O2 on current becomes significant at −20 mV. The degree of activation of the steady state Kv currents by H2O2 was large in the negative voltage range compared to those at potentials positive to 0 mV (Fig. 2c). This indicates that H2O2 can act as a potent modulator of the Kv channel function in rat MASMCs within the range of physiologically relevant voltages. H2O2, even at 10 mM, did not result in any nonspecific effects due to cellular damage. In all cells tested, neither access resistance nor leak current was significantly altered (Fig. 2a, b).
Fig. 2.

H2O2 increases Kv currents in MASMCs in a concentration-dependent manner. a Representative current traces illustrating the effect of H2O2 (0.1–10 mM) on whole-cell Kv currents. Deactivating tail currents were observed at −40 mV, following various test potentials, indicative of Kv channel activation. Application of 4-AP (10 mM) reduced current below the baseline level. b Current–voltage (I–V) relationship with H2O2 treatment (0.1–10 mM) (n = 19 MASMCs; **p < 0.01 vs. control; ***p < 0.001 vs. control). c H2O2-induced activation of Kv current. Amplitudes of Kv currents were measured at +40 mV. The magnitude of activation with various concentrations of H2O2 was plotted against Em (n = 19 for each concentration). Insets show the indicated graph with expanded scale
We then examined whether H2O2 influences voltage dependence of activation and activation kinetics of the putative Kv current (Fig. 3). We noticed that the conductance–voltage (G–V) curves of the Kv channel were significantly shifted to more negative potentials after H2O2 application (Fig. 3a), i.e., the channel was now activated at a more hyperpolarized potential. V 1/2 (midpoint of the G–V curve) in control and 0.1, 1, and 10 mM H2O2 were 7.6 ± 1.0, 2.7 ± 1.4, −3.1 ± 1.3, and −1.7 ± 1.8 mV, respectively (n = 14; Fig. 3b). The slope factors were unaffected (Fig. 3a). H2O2 also had an important effect on the time course of activation of the Kv channel (Fig. 3c). For example, for a −20-mV depolarizing pulse, the time constant of activation decreased by 60 %. Similar effects were observed for all depolarizing pulses tested (n = 16, Fig. 3d), suggesting that H2O2 speeds up the activation of Kv currents. The data from Figs. 2 and 3 (i.e., 4-AP sensitivity, voltage dependence, and kinetics) suggest that the currents modulated by H2O2 are 4-AP-sensitive KV currents.
Biochemical evidence for the Kv channel S-glutathionylation by H2O2
To reveal the mechanism(s) by which H2O2 increases the voltage sensitivity and activation kinetics of 4-AP-sensitive Kv channels, we examined whether the Kv channel proteins from rat MASMCs can be glutathionylated after exposure to H2O2. It has been reported that Kv1.2 and Kv1.5 channels are molecular identities of 4-AP-sensitive Kv channels in vascular smooth muscle cells. In addition, a recent study reported that Kv2.1 and Kv 9.3 channels are oxygen-sensitive K+ channels in the pulmonary artery. Since a previous study confirmed the presence of Kv1.2, Kv1.5, and Kv2.1 channel proteins in isolated MASMCs by immunocytochemistry [75], we examined the S-glutathionylation of these channel proteins by using a streptavidin pull-down assay. Primarily cultured rat MASMCs were loaded with BioGEE (250 μM) for 1 h, followed by H2O2 (0.1–10 mM) challenge for 10 min, as described previously [82]. Strong Kv1.2 (75 kDa), Kv1.5 (67 kDa), and Kv2.1 (95 kDa) bands were detected in the whole-cell lysates (Fig. 4, lower panel). If BioGEE was incorporated into channel proteins, streptavidin beads should pull down the channel protein–BioGEE complex, which would be further detected by channel protein antibodies in a Western blot. In contrast, if the channel proteins were not glutathionylated, the binding of the channel protein to BioGEE should decrease, resulting in a weaker band or even no band in the Western blot. In the streptavidin pull-down experiments, the immunoreactivity of Kv2.1 was significantly increased in the cell lysate pretreated with H2O2, compared to control cells (Fig. 4, upper panel). After streptavidin pull-down, immunoreactivities of Kv1.2 and Kv1.5 were not detectable. Western blotting of whole lysates verified Kv1.2 and Kv1.5 protein expression. Similar results were obtained using immunoprecipitation with anti-GSH, followed by immunoblot with antibodies against Kv1.2, Kv1.5, and Kv2.1 (data not shown).
Fig. 4.

S-glutathionylation of the Kv 2.1 channel after exposure to H2O2. a Kv 2.1 channels were detected only from samples that were obtained from the cells treated with both BioGEE and H2O2 in the streptavidin pull-down assay. In the streptavidin pull-down assay, Kv1.2 and Kv1.5 channels were not detected even from samples that were obtained from cells treated with both BioGEE and H2O2. b Kv channel subunits were detected through conventional Western blot using rat mesenteric arterial smooth muscle cell (MASMC) primary cultures pretreated with various concentrations of H2O2 and untreated control. Band density did not change with H2O2 treatment. The results are representative examples of three independent experiments
S-glutathionylation mediates activation of Kv currents by H2O2
GSSG causes S-glutathionylation [20]. We examined whether GSSG directly increased Kv currents. We established the conventional whole-cell configuration to deliver GSSG to the cytosol, as done in a previous study [68]. Current recordings usually started 10 min after the whole-cell configuration was made. Intracellular loading of 10 mM GSSG via a patch pipette significantly increased the Kv current (Fig. 5a). At +40 mV, Kv current densities in the absence and presence of GSSG were 17.7 ± 1.5 pA/pF (n = 19) and 37.8 ± 4.3 pA/pF (n = 21, p < 0.01), respectively (Fig. 5b). Furthermore, with GSSG in the pipette, subsequent application of H2O2 had no effect on Kv current (Fig. 5a, b). We then tested whether GSSG would also induce a negative shift in the activation curves for Kv channels. We found that G–V curves of the Kv channel were significantly shifted to more negative potentials after GSSG application. V 1/2 in the presence of GSSG was −1.9 ± 2.2 mV (n = 12), and it was not significantly different from that induced by 10 mM H2O2 (−1.7 ± 1.8 mV, n = 14). GSSG also had an effect on the time course of activation (n = 8, Fig. 5c). In the presence of GSSG, the time constant of activation decreased, and the change was even larger than that observed with 10 mM H2O2. More importantly, when 10 mM H2O2 was added to the GSSG-treated cells, we observed no further change in the G–V curve (V 1/2; −1.0 ± 1.6 mV, n = 12) and activation kinetics (Fig. 5c). These data indicate that GSSG mimicked and occluded the effects of H2O2, implying that S-glutathionylation appears to occur in the Kv channel during H2O2 application leading to the activation of the channel activity.
To confirm that S-glutathionylation mediates the activation of Kv currents by H2O2, we examined the effect of glutathione reductase on the action of H2O2. Glutathione reductase reduces GSSG to GSH and prevents oxidation of GSH. Since NADPH is an indispensable cofactor for glutathione reductase activity, NADPH (1 mM) was applied to cells together with glutathione reductase. NADPH alone did not block the stimulatory effect of H2O2 on Kv currents (Fig. 5e). However, intracellular glutathione reductase completely abolished the stimulatory effect of H2O2 on Kv currents (Fig. 5d). Data summarized in Fig. 5f suggest that increased channel activity by H2O2 occurs because of direct modification of thiol groups on the Kv channel by GSSG in rat MASMCs.
Redox status determines the response of Kv current to H2O2
Recent studies showed that an increase of ROS is linked to hypertension [17]. Most endogenously produced ROS, including H2O2, are derived from mitochondrial respiration [23, 36], wherein 1–2 % of consumed oxygen is converted to superoxide radical and then to H2O2 [10, 15]. Since H2O2 treatment does not induce relaxation but contraction of vascular smooth muscle cells in hypertensive vessels [28], we hypothesized that the increased basal H2O2 preoccupied the activation mechanism of Kv channels, thus rendering H2O2 treatment ineffective in activating Kv channels. To mimic the endogenously generated H2O2, we directly conveyed H2O2 into the cytosol by adding H2O2 in the patch pipette. The elevated intracellular level of H2O2 increased the Kv currents (Fig. 6a). At +40 mV, 0.1 and 10 mM H2O2 increased Kv current density up to 45.7 ± 4.8 pA/pF (n = 21, p < 0.01 vs. control pipette) and 43.8 ± 3.7 pA/pF (n = 21, p < 0.01 vs. control pipette), respectively (Fig. 6a). Notably, the concentration–response relationship was shifted to the left, implying that intracellular H2O2 is more effective in elevating Kv currents than extracellular H2O2. Under this high level of intracellular H2O2, the stimulatory effect of bath-applied H2O2 was completely abolished (Fig. 6b). Summarized data in Fig. 6c showed that the basal Kv current density under 0.1 mM of intracellular H2O2 was 45.7 ± 4.8 pA/pF (n = 21), and it was not enhanced but unaffected (43.9 ± 4.4 pA/pF, p > 0.05) or rather reduced (37.9 ± 4.6 pA/pF, p < 0.01) by subsequent bath application of 0.1 and 10 mM H2O2, respectively. These data suggest that high levels of basal H2O2 upregulates Kv currents through S-glutathionylation, which may keep acute exposure to H2O2 from regulating the Kv channels.
Fig. 6.
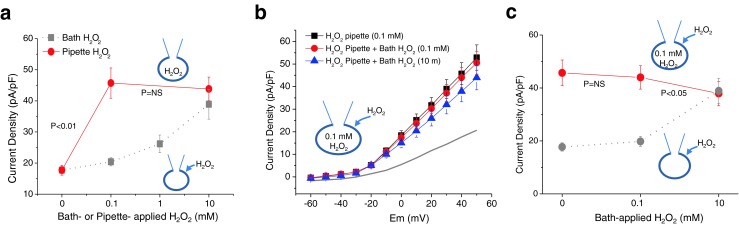
Effects of intracellular redox states on the capacity of H2O2 to stimulate Kv channels. a Intracellular application of H2O2 induced a concentration-dependent increase of Kv currents (filled red circle, n = 21). Compared to bath application (filled gray square, n = 21), the concentration–response relationship was shifted to the left when H2O2 was added intracellularly. Current densities of Kv channels were measured at +40 mV. b Effects of 100 μM H2O2 in the pipette solution on the action of H2O2. Averaged I–V curves before (filled black square) and after bath application of 0.1 mM H2O2 (filled red circle) and 10 mM H2O2 (filled blue triangle) are shown. Gray line indicates the averaged I–V relationship with control pipette solution (redrawn from Fig. 2a for comparison). Note that Kv currents with pipette solution containing 100 μM H2O2 (filled black square) are much larger than those with control pipette solution (gray line). c Comparative data between Kv current densities measured at +40 mV for H2O2 effects in control pipette solution (filled gray circle; n = 21) and those in the pipette containing 100 μM H2O2 (filled red circle; n = 21)
Discussion
It is still uncertain whether H2O2 acts as a vasodilator [55, 80]. There is even less certainty regarding the signal transduction and/or effector mechanism(s) that may be involved in the vascular response to H2O2. For example, it may involve the BKCa channel [81], KATP channel [40], or Kv channel [55]. The present study resolves some of this complexity by providing direct evidence for an effector molecule that can mediate H2O2-induced vasodilation through the Kv channel and proposes that S-glutathionylation underlies the stimulatory effect of H2O2 on Kv channels. Further experiments demonstrate that under oxidative conditions, Kv channels were not stimulated but rather inhibited by acute exposure to H2O2, suggesting that cellular redox status affected S-glutathionylation of the Kv channel and determined the response of the Kv channel to H2O2.
In the present study, H2O2 relaxed the rat mesenteric artery that was precontracted with an agonist. In contrast, H2O2 failed to relax arteries pretreated with DTT and 4-AP; 1 mM TEA and 100 μM BaCl2 did not affect the H2O2 response. The inability of H2O2 to relax arteries pretreated with 4-AP suggested that the relaxation response to H2O2 involved stimulation of the Kv channel, a mechanism that has also been suggested from patch-clamp/whole-cell studies employing other cell types [12, 54]. Here, we directly measured the effects of H2O2 on Kv channels in MASMCs. H2O2 increased 4-AP-sensitive Kv currents in a concentration-dependent manner. This was a result of change in the voltage dependence of activation; 10 mM H2O2 shifted voltage dependence of 4-AP-sensitive Kv channel conductance by ~7.6 mV (Fig. 3a, b), and the shift was concentration-dependent. The activation kinetics was also accelerated after exposure to H2O2 (Fig. 3c, d). It is worthy to note that unlike the role of BKCa channel in H2O2-induced dilation of coronary arteries and arterioles [5, 81], the BKCa channel did not contribute to the dilatory effect of H2O2 in rat mesenteric arteries (Fig. 1). This discrepancy may suggest that the regulatory mechanisms underlying vascular tone and the sensitivity of diverse K+ channels to H2O2 differ among different types of arteries. The 4-AP-sensitive Kv channels are expressed in high density in myocytes derived from rat mesenteric [75] and human pulmonary [26] arteries and are important targets of receptor agonists [3]. BKCa channels are the key determinant of coronary arterial tone [37].
Previous studies have proposed a variety of molecular pathways that can be stimulated by H2O2. For example, guanylyl cyclase may underlie H2O2 relaxation of pulmonary arteries [11], while arachidonic acid (AA) may mediate a vasodilator effect of H2O2 in coronary arteries [5]. Recent studies have shown that H2O2 induces S-glutathionylation of the channel protein, thereby affecting channel activity [71, 77, 78]. Since the thiol-specific reducing agent DTT blocked the vasodilatory effect of H2O2 (Fig. 1), we considered it possible that S-glutathionylation of the Kv channel protein mediates the stimulatory effect of H2O2 on Kv currents in the mesenteric artery smooth muscle. We found that addition of H2O2 to MASMCs increased S-glutathionylation of the Kv2.1 channel protein dramatically (Fig. 4). It is well known that Kv2.1 expresses a slow-inactivating, TEA-resistant, and 4-AP-sensitive Kv current in rat and human mesenteric arteries [51]. However, we could not exclude a possible glutathionylation of Kv1.2 and Kv1.5 channel proteins since it might not have been detected due to differences in pull-down efficiency in each Kv channel after S-glutathionylation. To confirm the cause–effect relationship, we blocked S-glutathionylation by using GSH reductase. GSH reductase completely abolished the stimulatory effect of H2O2. Addition of exogenous GSSG directly stimulated channel activity. Similar to H2O2, GSSG alters channel function by speeding up the activation kinetics and shifting the voltage dependence of channel activation to the left. Bath application of H2O2 (10 mM), subsequent to maximal GSSG effect, induced no further change in the Kv channel. These results suggest that S-glutathionylation of the Kv channel protein mediates the stimulatory effect of H2O2 on the Kv channel and, consequently, the vasodilatory effects in the mesenteric artery. Interestingly, conditions of increased oxidative stress within smooth muscle cells impaired the capacity of exogenous H2O2 to stimulate Kv channels (Fig. 6). Not only was the H2O2 stimulatory effect much weaker, but also the inhibitory effect of H2O2 was unmasked. The molecular mechanism of how H2O2 inhibits Kv channel under oxidative condition is not yet known. However, it can be speculated that since S-glutathionylation of the Kv channel persists and the Kv channels are already maximally enhanced, signals such as cyclooxygenase are involved [5]. Taken together, S-glutathionylation of the Kv channel under elevated basal H2O2 levels may be involved in the development of the pathology of the hypertensive vessel. This concept is still speculative; therefore, further studies will be required to test this hypothesis.
In the present study, a high concentration of extracellular H2O2 is required to regulate KV channels. This argument holds for neurons. This can be reflected from the fact that, in the hippocampus, the IC50 value for extracellular H2O2 to affect postsynaptic potentials was nearly 6 mM [46]. In contrast to extracellular application, a low level of intracellular H2O2 is sufficient to elevate the Kv currents (Fig. 6). This difference suggests that either the permeability of the cell membrane to H2O2 is low [7, 8, 31] or the rate of H2O2 degradation is high near the cell membrane [39]. In addition, the difference in effects possibly indicates that the modulation of thiol groups takes place mainly on the intracellular side of the plasma membrane. This is further supported by the fact that there are no cysteines in the extracellular location of the Kv 2.1 channel. In Kv2.1 channels, 15 cysteines are present: four in a COOH-terminal domain, three in transmembrane core regions (S2 and S6), and the remaining eight in an NH2-terminal domain (Supplementary Fig. 1). Our electrophysiological data showed that the reaction of Kv2.1 with H2O2 or GSSG caused a pronounced increase in channel kinetics and left shift of steady state activation. However, all cysteines of Kv2.1 channels are located outside of S4, a central component of the voltage sensor. Given that the NH2 terminus has the largest number of cysteines and it has been shown to participate in channel gating [50], one or more cysteines located at the NH2 terminus might be involved in the effects of H2O2 on channel activation. Further studies are required to ascertain this.
Combined with the fact that endothelial cells can produce up to 500 μM H2O2 [27] and myoendothelial gap junction can be a pathway of H2O2 from the endothelium to the smooth muscle [16], our results suggest that endothelium-derived H2O2 can act as a relaxing factor in mammalian arteries. EDHFs are important factors controlling the vascular tone. Sobey [64] suggested that EDHFs play a major role in conditions of high blood pressure, arteriosclerosis, and diabetes by controlling potassium ion channels. The identity of the EDHFs differs depending on the animal species and type of arteries examined [24, 35, 41, 42, 52, 54]. The four major EDHF candidates are an electrical coupling through myoendothelial gap junctions, potassium ions (K+), cytochrome P450 metabolites of AA such as epoxyeicosatrienoic acid, and H2O2 [7, 31, 42, 76]. A study published in 1991, for the first time, suggested that H2O2 was an EDHF [6]: production of H2O2 by hyperpolarization of the endothelium, which consequently acts on vascular smooth muscle cells, causing relaxation of blood vessels. Subsequently, several studies have verified the hypothesis that H2O2 is an EDHF in animal and human arteries [35, 41, 42, 48, 76]. Although the contribution of EDHFs to vascular tone is not entirely clear, it is generally accepted that nitric oxide (NO) plays a dominant role in controlling the tone of large conduit blood vessels compared to EDHFs, whereas EDHF is more important in small-resistance blood vessels [14]. Consistent with this notion, we found that acetylcholine-induced endothelium-dependent vasodilation was largely inhibited by catalase in small mesenteric arteries. In contrast, acetylcholine-induced endothelium-dependent vasodilation was largely inhibited by an NO synthase blocker, but not by catalase in the aorta (Supplementary Fig. 2).
Posttranslational modifications (PTMs) are important mechanisms regulating ion channel functions. One of the classical PTMs is protein phosphorylation, and a large number of ion channels are regulated by phosphorylation through protein kinase A (PKA), PKC, and other protein kinases [19, 32, 60-62, 79]. A variety of different types of PTMs (e.g., ubiquitylation, SUMOylation, O-glycosylation/O-GlcNAcylation) exist and are discussed elsewhere [9, 13, 53, 56]. Among all these PTMs, redox-mediated PTM is an important category of PTMs that targets the thiol group of cysteine residues. Recently, redox-mediated PTMs are receiving increasing attention, as they are found in both physiological and pathological conditions, including oxidative stress. S-glutathionylation is a major redox-mediated thiol modulation mechanism, involving the addition of a GSH moiety to the protein. Oxidative stress and ROS facilitate S-glutathionylation. Over the past few years, S-glutathionylation has been increasingly observed in many ion channels such as voltage-gated calcium channels, the ryanodine receptor, and KATP channels, all of which contribute to critical cellular functions [1, 71, 77, 78]. Our results indicate that the Kv channel protein is significantly glutathionylated after exposure to H2O2 (Fig. 4). As delineated above, S-glutathionylation of the Kv channel resulted in an increase in the Kv currents in myocytes (Fig. 5). Alterations in thiol groups on proteins can alter function through structural changes in the channel protein. Since H2O2 changed the channel gating properties (Fig. 3), we propose that S-glutathionylation of the Kv2.1 channel protein causes a structural rearrangement of the channel that results in an increase in voltage sensitivity.
In conclusion, H2O2 relaxed rat mesenteric arteries by S-glutathionylation-dependent activation of Kv currents under physiological conditions. Our data suggest that S-glutathionylation of the Kv channel protein is, at least in part, an important and novel mechanism of 4-AP-sensitive Kv current activation by H2O2. Identifying the mechanisms underlying the vasoactive effects of H2O2 should increase our understanding of diseases where oxidative damage has been implicated such as in hypertension, atherosclerosis, and diabetes mellitus.
Electronic supplementary material
Below is the link to the electronic supplementary material.
(PDF 164 kb)
Acknowledgements
This research was supported by the Pioneer (2011-0027921) and Basic Science Research Program (NRF-2012046878 and NRF-2013025108) through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology.
Abbreviations
- 4-AP
4-Aminopyridine
- AA
Arachidonic acid
- BioGee
Biotinylated glutathione ethyl ester
- BKCa
Big-conductance Ca2+-activated K+
- DTT
DL-Dithiothreitol
- EDHF
Endothelium-derived hyperpolarizing factor
- Em
Membrane potential
- GSSG
Oxidized form of glutathione
- H2O2
Hydrogen peroxide
- KATP
ATP-sensitive K+
- KCa
Ca2+-activated K+
- Kir
Inward rectifier K+
- Kv
Voltage-gated K+
- MASMCs
Mesenteric arterial smooth muscle cells
- NO
Nitric oxide
- NT
Normal Tyrode
- PSS
Physiological salt solution
- ROS
Reactive oxygen species
- TEA
Tetraethylammonium
Contributor Information
Young Min Bae, Phone: +82-43-8403727, FAX: +82-43-8519329, Email: ymbae30@kku.ac.kr.
Hana Cho, Phone: +82-31-2996104, FAX: +82-31-2996129, Email: hanacho@skku.edu.
References
- 1.Aracena-Parks P, Goonasekera SA, Gilman CP, Dirksen RT, Hidalgo C, Hamilton SL. Identification of cysteines involved in S-nitrosylation, S-glutathionylation, and oxidation to disulfides in ryanodine receptor type 1. J Biol Chem. 2006;281(52):40354–40368. doi: 10.1074/jbc.M600876200. [DOI] [PubMed] [Google Scholar]
- 2.Archer SL, Wu XC, Thebaud B, Moudgil R, Hashimoto K, Michelakis ED. O2 sensing in the human ductus arteriosus: redox-sensitive K+ channels are regulated by mitochondria-derived hydrogen peroxide. Biol Chem. 2004;385(3–4):205–216. doi: 10.1515/BC.2004.014. [DOI] [PubMed] [Google Scholar]
- 3.Bae YM, Kim A, Kim J, Park SW, Kim TK, Lee YR, Kim B, Cho SI. Serotonin depolarizes the membrane potential in rat mesenteric artery myocytes by decreasing voltage-gated K+ currents. Biochem Biophys Res Commun. 2006;347(2):468–476. doi: 10.1016/j.bbrc.2006.06.116. [DOI] [PubMed] [Google Scholar]
- 4.Bae YM, Park MK, Lee SH, Ho WK, Earm YE. Contribution of Ca2+-activated K+ channels and non-selective cation channels to membrane potential of pulmonary arterial smooth muscle cells of the rabbit. J Physiol. 1999;514(Pt 3):747–758. doi: 10.1111/j.1469-7793.1999.747ad.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Barlow RS, White RE. Hydrogen peroxide relaxes porcine coronary arteries by stimulating BKCa channel activity. Am J Physiol. 1998;275(4 Pt 2):H1283–H1289. doi: 10.1152/ajpheart.1998.275.4.H1283. [DOI] [PubMed] [Google Scholar]
- 6.Beny JL, von der Weid PY. Hydrogen peroxide: an endogenous smooth muscle cell hyperpolarizing factor. Biochem Biophys Res Commun. 1991;176(1):378–384. doi: 10.1016/0006-291X(91)90935-Z. [DOI] [PubMed] [Google Scholar]
- 7.Bienert GP, Moller AL, Kristiansen KA, Schulz A, Moller IM, Schjoerring JK, Jahn TP. Specific aquaporins facilitate the diffusion of hydrogen peroxide across membranes. J Biol Chem. 2007;282(2):1183–1192. doi: 10.1074/jbc.M603761200. [DOI] [PubMed] [Google Scholar]
- 8.Bienert GP, Schjoerring JK, Jahn TP. Membrane transport of hydrogen peroxide. Biochim Biophys Acta. 2006;1758(8):994–1003. doi: 10.1016/j.bbamem.2006.02.015. [DOI] [PubMed] [Google Scholar]
- 9.Bimboese P, Gibson CJ, Schmidt S, Xiang W, Ehrlich BE. Isoform-specific regulation of the inositol 1,4,5-trisphosphate receptor by O-linked glycosylation. J Biol Chem. 2011;286(18):15688–15697. doi: 10.1074/jbc.M110.206482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Boveris A, Chance B. The mitochondrial generation of hydrogen peroxide. General properties and effect of hyperbaric oxygen. Biochem J. 1973;134(3):707–716. doi: 10.1042/bj1340707. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Burke TM, Wolin MS. Hydrogen peroxide elicits pulmonary arterial relaxation and guanylate cyclase activation. Am J Physiol. 1987;252(4 Pt 2):H721–H732. doi: 10.1152/ajpheart.1987.252.4.H721. [DOI] [PubMed] [Google Scholar]
- 12.Caouette D, Dongmo C, Berube J, Fournier D, Daleau P. Hydrogen peroxide modulates the Kv1.5 channel expressed in a mammalian cell line. Naunyn Schmiedeberg’s Arch Pharmacol. 2003;368(6):479–486. doi: 10.1007/s00210-003-0834-0. [DOI] [PubMed] [Google Scholar]
- 13.Chandrasekhar KD, Lvov A, Terrenoire C, Gao GY, Kass RS, Kobertz WR. O-glycosylation of the cardiac I(Ks) complex. J Physiol. 2011;589(Pt 15):3721–3730. doi: 10.1113/jphysiol.2011.211284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Clark SG, Fuchs LC. Role of nitric oxide and Ca++-dependent K+ channels in mediating heterogeneous microvascular responses to acetylcholine in different vascular beds. J Pharmacol Exp Ther. 1997;282(3):1473–1479. [PubMed] [Google Scholar]
- 15.Cohen G. Enzymatic/nonenzymatic sources of oxyradicals and regulation of antioxidant defenses. Ann N Y Acad Sci. 1994;738:8–14. doi: 10.1111/j.1749-6632.1994.tb21784.x. [DOI] [PubMed] [Google Scholar]
- 16.Coleman HA, Tare M, Parkington HC. Endothelial potassium channels, endothelium-dependent hyperpolarization and the regulation of vascular tone in health and disease. Clin Exp Pharmacol Physiol. 2004;31(9):641–649. doi: 10.1111/j.1440-1681.2004.04053.x. [DOI] [PubMed] [Google Scholar]
- 17.Cruzado MC, Risler NR, Miatello RM, Yao G, Schiffrin EL, Touyz RM. Vascular smooth muscle cell NAD(P)H oxidase activity during the development of hypertension: effect of angiotensin II and role of insulin-like growth factor-1 receptor transactivation. Am J Hypertens. 2005;18(1):81–87. doi: 10.1016/j.amjhyper.2004.09.001. [DOI] [PubMed] [Google Scholar]
- 18.Cseko C, Bagi Z, Koller A. Biphasic effect of hydrogen peroxide on skeletal muscle arteriolar tone via activation of endothelial and smooth muscle signaling pathways. J Appl Physiol. 2004;97(3):1130–1137. doi: 10.1152/japplphysiol.00106.2004. [DOI] [PubMed] [Google Scholar]
- 19.Dai S, Hall DD, Hell JW. Supramolecular assemblies and localized regulation of voltage-gated ion channels. Physiol Rev. 2009;89(2):411–452. doi: 10.1152/physrev.00029.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Dalle-Donne I, Milzani A, Gagliano N, Colombo R, Giustarini D, Rossi R. Molecular mechanisms and potential clinical significance of S-glutathionylation. Antioxid Redox Signal. 2008;10(3):445–473. doi: 10.1089/ars.2007.1716. [DOI] [PubMed] [Google Scholar]
- 21.Dalle-Donne I, Rossi R, Colombo G, Giustarini D, Milzani A. Protein S-glutathionylation: a regulatory device from bacteria to humans. Trends Biochem Sci. 2009;34(2):85–96. doi: 10.1016/j.tibs.2008.11.002. [DOI] [PubMed] [Google Scholar]
- 22.Dalle-Donne I, Rossi R, Giustarini D, Colombo R, Milzani A. S-glutathionylation in protein redox regulation. Free Radic Biol Med. 2007;43(6):883–898. doi: 10.1016/j.freeradbiomed.2007.06.014. [DOI] [PubMed] [Google Scholar]
- 23.Dugan LL, Sensi SL, Canzoniero LM, Handran SD, Rothman SM, Lin TS, Goldberg MP, Choi DW. Mitochondrial production of reactive oxygen species in cortical neurons following exposure to N-methyl-D-aspartate. J Neurosci. 1995;15(10):6377–6388. doi: 10.1523/JNEUROSCI.15-10-06377.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Edwards G, Dora KA, Gardener MJ, Garland CJ, Weston AH. K+ is an endothelium-derived hyperpolarizing factor in rat arteries. Nature. 1998;396(6708):269–272. doi: 10.1038/24388. [DOI] [PubMed] [Google Scholar]
- 25.Finkel T. Oxygen radicals and signaling. Curr Opin Cell Biol. 1998;10(2):248–253. doi: 10.1016/S0955-0674(98)80147-6. [DOI] [PubMed] [Google Scholar]
- 26.Firth AL, Remillard CV, Platoshyn O, Fantozzi I, Ko EA, Yuan JX. Functional ion channels in human pulmonary artery smooth muscle cells: voltage-dependent cation channels. Pulm Cir. 2011;1(1):48–71. doi: 10.4103/2045-8932.78103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Gao YJ, Lee RM. Hydrogen peroxide is an endothelium-dependent contracting factor in rat renal artery. Br J Pharmacol. 2005;146(8):1061–1068. doi: 10.1038/sj.bjp.0706423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Garcia-Redondo AB, Briones AM, Beltran AE, Alonso MJ, Simonsen U, Salaices M. Hypertension increases contractile responses to hydrogen peroxide in resistance arteries through increased thromboxane A2, Ca2+, and superoxide anion levels. J Pharmacol Exp Ther. 2009;328(1):19–27. doi: 10.1124/jpet.108.144295. [DOI] [PubMed] [Google Scholar]
- 29.Hatoum OA, Binion DG, Miura H, Telford G, Otterson MF, Gutterman DD. Role of hydrogen peroxide in ACh-induced dilation of human submucosal intestinal microvessels. Am J Physiol Heart Circ Physiol. 2005;288(1):H48–H54. doi: 10.1152/ajpheart.00663.2004. [DOI] [PubMed] [Google Scholar]
- 30.Heinle H. Vasoconstriction of carotid artery induced by hydroperoxides. Arch Int Physiol Biochim. 1984;92(4):267–271. doi: 10.3109/13813458409071166. [DOI] [PubMed] [Google Scholar]
- 31.Henzler T, Steudle E. Transport and metabolic degradation of hydrogen peroxide in Chara corallina: model calculations and measurements with the pressure probe suggest transport of H2O2 across water channels. J Exp Bot. 2000;51(353):2053–2066. doi: 10.1093/jexbot/51.353.2053. [DOI] [PubMed] [Google Scholar]
- 32.Kamp TJ, Hell JW. Regulation of cardiac L-type calcium channels by protein kinase A and protein kinase C. Circ Res. 2000;87(12):1095–1102. doi: 10.1161/01.RES.87.12.1095. [DOI] [PubMed] [Google Scholar]
- 33.Kim SH, Bae YM, Sung DJ, Park SW, Woo NS, Kim B, Cho SI. Ketamine blocks voltage-gated K(+) channels and causes membrane depolarization in rat mesenteric artery myocytes. Pflugers Arch. 2007;454(6):891–902. doi: 10.1007/s00424-007-0240-4. [DOI] [PubMed] [Google Scholar]
- 34.Kozlowska H, Baranowska M, Gromotowicz A, Malinowska B. Endothelium-derived hyperpolarizing factor (EDHF): potential involvement in the physiology and pathology of blood vessels. Postepy Hig Med Dosw. 2007;61:555–564. [PubMed] [Google Scholar]
- 35.Lacza Z, Puskar M, Kis B, Perciaccante JV, Miller AW, Busija DW. Hydrogen peroxide acts as an EDHF in the piglet pial vasculature in response to bradykinin. Am J Physiol Heart Circ Physiol. 2002;283(1):H406–H411. doi: 10.1152/ajpheart.00007.2002. [DOI] [PubMed] [Google Scholar]
- 36.Liu Y, Fiskum G, Schubert D. Generation of reactive oxygen species by the mitochondrial electron transport chain. J Neurochem. 2002;80(5):780–787. doi: 10.1046/j.0022-3042.2002.00744.x. [DOI] [PubMed] [Google Scholar]
- 37.Liu Y, Gutterman DD. The coronary circulation in diabetes: influence of reactive oxygen species on K+ channel-mediated vasodilation. Vasc Pharmacol. 2002;38(1):43–49. doi: 10.1016/S1537-1891(02)00125-8. [DOI] [PubMed] [Google Scholar]
- 38.Lucchesi PA, Belmadani S, Matrougui K. Hydrogen peroxide acts as both vasodilator and vasoconstrictor in the control of perfused mouse mesenteric resistance arteries. J Hypertens. 2005;23(3):571–579. doi: 10.1097/01.hjh.0000160214.40855.79. [DOI] [PubMed] [Google Scholar]
- 39.Ma HP. Hydrogen peroxide stimulates the epithelial sodium channel through a phosphatidylinositide 3-kinase-dependent pathway. J Biol Chem. 2011;286(37):32444–32453. doi: 10.1074/jbc.M111.254102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Marvar PJ, Hammer LW, Boegehold MA. Hydrogen peroxide-dependent arteriolar dilation in contracting muscle of rats fed normal and high salt diets. Microcirculation. 2007;14(8):779–791. doi: 10.1080/10739680701444057. [DOI] [PubMed] [Google Scholar]
- 41.Matoba T, Shimokawa H, Kubota H, Morikawa K, Fujiki T, Kunihiro I, Mukai Y, Hirakawa Y, Takeshita A. Hydrogen peroxide is an endothelium-derived hyperpolarizing factor in human mesenteric arteries. Biochem Biophys Res Commun. 2002;290(3):909–913. doi: 10.1006/bbrc.2001.6278. [DOI] [PubMed] [Google Scholar]
- 42.Matoba T, Shimokawa H, Nakashima M, Hirakawa Y, Mukai Y, Hirano K, Kanaide H, Takeshita A. Hydrogen peroxide is an endothelium-derived hyperpolarizing factor in mice. J Clin Invest. 2000;106(12):1521–1530. doi: 10.1172/JCI10506. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Michelakis ED, Rebeyka I, Wu X, Nsair A, Thebaud B, Hashimoto K, Dyck JR, Haromy A, Harry G, Barr A, Archer SL. O2 sensing in the human ductus arteriosus: regulation of voltage-gated K+ channels in smooth muscle cells by a mitochondrial redox sensor. Circ Res. 2002;91(6):478–486. doi: 10.1161/01.RES.0000035057.63303.D1. [DOI] [PubMed] [Google Scholar]
- 44.Michelakis ED, Thebaud B, Weir EK, Archer SL. Hypoxic pulmonary vasoconstriction: redox regulation of O2-sensitive K+ channels by a mitochondrial O2-sensor in resistance artery smooth muscle cells. J Mol Cell Cardiol. 2004;37(6):1119–1136. doi: 10.1016/j.yjmcc.2004.09.007. [DOI] [PubMed] [Google Scholar]
- 45.Nelson MT, Quayle JM. Physiological roles and properties of potassium channels in arterial smooth muscle. Am J Physiol. 1995;268(4 Pt 1):C799–C822. doi: 10.1152/ajpcell.1995.268.4.C799. [DOI] [PubMed] [Google Scholar]
- 46.Nistico R, Piccirilli S, Cucchiaroni ML, Armogida M, Guatteo E, Giampa C, Fusco FR, Bernardi G, Nistico G, Mercuri NB. Neuroprotective effect of hydrogen peroxide on an in vitro model of brain ischaemia. Br J Pharmacol. 2008;153(5):1022–1029. doi: 10.1038/sj.bjp.0707587. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Nowicki PT, Flavahan S, Hassanain H, Mitra S, Holland S, Goldschmidt-Clermont PJ, Flavahan NA. Redox signaling of the arteriolar myogenic response. Circ Res. 2001;89(2):114–116. doi: 10.1161/hh1401.094367. [DOI] [PubMed] [Google Scholar]
- 48.Palen DI, Ouhtit A, Belmadani S, Lucchesi PA, Matrougui K. Hydrogen peroxide acts as relaxing factor in human vascular smooth muscle cells independent of map-kinase and nitric oxide. Front Biosci. 2006;11:2526–2534. doi: 10.2741/1987. [DOI] [PubMed] [Google Scholar]
- 49.Park WS, Son YK, Ko EA, Ko JH, Lee HA, Park KS, Earm YE. The protein kinase C inhibitor, bisindolylmaleimide (I), inhibits voltage-dependent K+ channels in coronary arterial smooth muscle cells. Life Sci. 2005;77(5):512–527. doi: 10.1016/j.lfs.2004.10.073. [DOI] [PubMed] [Google Scholar]
- 50.Pascual JM, Shieh CC, Kirsch GE, Brown AM. Contribution of the NH2 terminus of Kv2.1 to channel activation. Am J Physiol. 1997;273(6 Pt 1):C1849–C1858. doi: 10.1152/ajpcell.1997.273.6.C1849. [DOI] [PubMed] [Google Scholar]
- 51.Patel AJ, Lazdunski M, Honore E. Kv2.1/Kv9.3, a novel ATP-dependent delayed-rectifier K+ channel in oxygen-sensitive pulmonary artery myocytes. EMBO J. 1997;16(22):6615–6625. doi: 10.1093/emboj/16.22.6615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Quyyumi AA, Ozkor M. Vasodilation by hyperpolarization: beyond NO. Hypertension. 2006;48(6):1023–1025. doi: 10.1161/01.HYP.0000250965.03934.15. [DOI] [PubMed] [Google Scholar]
- 53.Rengifo J, Gibson CJ, Winkler E, Collin T, Ehrlich BE. Regulation of the inositol 1,4,5-trisphosphate receptor type I by O-GlcNAc glycosylation. J Neurosci. 2007;27(50):13813–13821. doi: 10.1523/JNEUROSCI.2069-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Rogers PA, Chilian WM, Bratz IN, Bryan RM, Jr, Dick GM. H2O2 activates redox- and 4-aminopyridine-sensitive Kv channels in coronary vascular smooth muscle. Am J Physiol Heart Circ Physiol. 2007;292(3):H1404–H1411. doi: 10.1152/ajpheart.00696.2006. [DOI] [PubMed] [Google Scholar]
- 55.Rogers PA, Dick GM, Knudson JD, Focardi M, Bratz IN, Swafford AN, Jr, Saitoh S, Tune JD, Chilian WM. H2O2-induced redox-sensitive coronary vasodilation is mediated by 4-aminopyridine-sensitive K+ channels. Am J Physiol Heart Circ Physiol. 2006;291(5):H2473–H2482. doi: 10.1152/ajpheart.00172.2006. [DOI] [PubMed] [Google Scholar]
- 56.Rougier JS, Albesa M, Abriel H. Ubiquitylation and SUMOylation of cardiac ion channels. J Cardiovasc Pharmacol. 2010;56(1):22–28. doi: 10.1097/FJC.0b013e3181daaff9. [DOI] [PubMed] [Google Scholar]
- 57.Saitoh S, Zhang C, Tune JD, Potter B, Kiyooka T, Rogers PA, Knudson JD, Dick GM, Swafford A, Chilian WM. Hydrogen peroxide: a feed-forward dilator that couples myocardial metabolism to coronary blood flow. Arterioscler Thromb Vasc Biol. 2006;26(12):2614–2621. doi: 10.1161/01.ATV.0000249408.55796.da. [DOI] [PubMed] [Google Scholar]
- 58.Sato A, Sakuma I, Gutterman DD. Mechanism of dilation to reactive oxygen species in human coronary arterioles. Am J Physiol Heart Circ Physiol. 2003;285(6):H2345–H2354. doi: 10.1152/ajpheart.00458.2003. [DOI] [PubMed] [Google Scholar]
- 59.Schafer FQ, Buettner GR. Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple. Free Radic Biol Med. 2001;30(11):1191–1212. doi: 10.1016/S0891-5849(01)00480-4. [DOI] [PubMed] [Google Scholar]
- 60.Shi Y, Chen X, Wu Z, Shi W, Yang Y, Cui N, Jiang C, Harrison RW. cAMP-dependent protein kinase phosphorylation produces interdomain movement in SUR2B leading to activation of the vascular KATP channel. J Biol Chem. 2008;283(12):7523–7530. doi: 10.1074/jbc.M709941200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Shi W, Cui N, Shi Y, Zhang X, Yang Y, Jiang C. Arginine vasopressin inhibits Kir6.1/SUR2B channel and constricts the mesenteric artery via V1a receptor and protein kinase C. Am J Physiol Regul Integr Comp Physiol. 2007;293(1):R191–R199. doi: 10.1152/ajpregu.00047.2007. [DOI] [PubMed] [Google Scholar]
- 62.Shi Y, Wu Z, Cui N, Shi W, Yang Y, Zhang X, Rojas A, Ha BT, Jiang C. PKA phosphorylation of SUR2B subunit underscores vascular KATP channel activation by beta-adrenergic receptors. Am J Physiol Regul Integr Comp Physiol. 2007;293(3):R1205–R1214. doi: 10.1152/ajpregu.00337.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Shi WW, Yang Y, Shi Y, Jiang C. K(ATP) channel action in vascular tone regulation: from genetics to diseases. Sheng Li Xue Bao. 2012;64(1):1–13. [PMC free article] [PubMed] [Google Scholar]
- 64.Sobey CG. Potassium channel function in vascular disease. Arterioscler Thromb Vasc Biol. 2001;21(1):28–38. doi: 10.1161/01.ATV.21.1.28. [DOI] [PubMed] [Google Scholar]
- 65.Somers MJ, Mavromatis K, Galis ZS, Harrison DG. Vascular superoxide production and vasomotor function in hypertension induced by deoxycorticosterone acetate-salt. Circulation. 2000;101(14):1722–1728. doi: 10.1161/01.CIR.101.14.1722. [DOI] [PubMed] [Google Scholar]
- 66.Sung DJ, Noh HJ, Kim JG, Park SW, Kim B, Cho H, Bae YM. Serotonin contracts the rat mesenteric artery by inhibiting 4-aminopyridine-sensitive Kv channels via the 5-HT2A receptor and Src tyrosine kinase. Exp Mol Med. 2013;45:e67. doi: 10.1038/emm.2013.116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Suvorava T, Lauer N, Kumpf S, Jacob R, Meyer W, Kojda G. Endogenous vascular hydrogen peroxide regulates arteriolar tension in vivo. Circulation. 2005;112(16):2487–2495. doi: 10.1161/CIRCULATIONAHA.105.543157. [DOI] [PubMed] [Google Scholar]
- 68.Suzuki YJ, Cleemann L, Abernethy DR, Morad M. Glutathione is a cofactor for H2O2-mediated stimulation of Ca2+-induced Ca2+ release in cardiac myocytes. Free Radic Biol Med. 1998;24(2):318–325. doi: 10.1016/S0891-5849(97)00227-X. [DOI] [PubMed] [Google Scholar]
- 69.Takaki A, Morikawa K, Murayama Y, Yamagishi H, Hosoya M, Ohashi J, Shimokawa H. Roles of endothelial oxidases in endothelium-derived hyperpolarizing factor responses in mice. J Cardiovasc Pharmacol. 2008;52(6):510–517. doi: 10.1097/FJC.0b013e318190358b. [DOI] [PubMed] [Google Scholar]
- 70.Takaki A, Morikawa K, Tsutsui M, Murayama Y, Tekes E, Yamagishi H, Ohashi J, Yada T, Yanagihara N, Shimokawa H. Crucial role of nitric oxide synthases system in endothelium-dependent hyperpolarization in mice. J Exp Med. 2008;205(9):2053–2063. doi: 10.1084/jem.20080106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Tang H, Viola HM, Filipovska A, Hool LC. Ca(v)1.2 calcium channel is glutathionylated during oxidative stress in guinea pig and ischemic human heart. Free Radic Biol Med. 2011;51(8):1501–1511. doi: 10.1016/j.freeradbiomed.2011.07.005. [DOI] [PubMed] [Google Scholar]
- 72.Thakali K, Davenport L, Fink GD, Watts SW. Pleiotropic effects of hydrogen peroxide in arteries and veins from normotensive and hypertensive rats. Hypertension. 2006;47(3):482–487. doi: 10.1161/01.HYP.0000201540.91234.8f. [DOI] [PubMed] [Google Scholar]
- 73.Thomas G, Ramwell P. Induction of vascular relaxation by hydroperoxides. Biochem Biophys Res Commun. 1986;139(1):102–108. doi: 10.1016/S0006-291X(86)80085-7. [DOI] [PubMed] [Google Scholar]
- 74.Wei EP, Kontos HA, Beckman JS. Mechanisms of cerebral vasodilation by superoxide, hydrogen peroxide, and peroxynitrite. Am J Physiol. 1996;271(3 Pt 2):H1262–H1266. doi: 10.1152/ajpheart.1996.271.3.H1262. [DOI] [PubMed] [Google Scholar]
- 75.Xu C, Lu Y, Tang G, Wang R. Expression of voltage-dependent K(+) channel genes in mesenteric artery smooth muscle cells. Am J Physiol. 1999;277(5 Pt 1):G1055–G1063. doi: 10.1152/ajpgi.1999.277.5.G1055. [DOI] [PubMed] [Google Scholar]
- 76.Yada T, Shimokawa H, Hiramatsu O, Kajita T, Shigeto F, Goto M, Ogasawara Y, Kajiya F. Hydrogen peroxide, an endogenous endothelium-derived hyperpolarizing factor, plays an important role in coronary autoregulation in vivo. Circulation. 2003;107(7):1040–1045. doi: 10.1161/01.CIR.0000050145.25589.65. [DOI] [PubMed] [Google Scholar]
- 77.Yang Y, Shi W, Chen X, Cui N, Konduru AS, Shi Y, Trower TC, Zhang S, Jiang C. Molecular basis and structural insight of vascular K(ATP) channel gating by S-glutathionylation. J Biol Chem. 2011;286(11):9298–9307. doi: 10.1074/jbc.M110.195123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Yang Y, Shi W, Cui N, Wu Z, Jiang C. Oxidative stress inhibits vascular K(ATP) channels by S-glutathionylation. J Biol Chem. 2010;285(49):38641–38648. doi: 10.1074/jbc.M110.162578. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Yang Y, Shi Y, Guo S, Zhang S, Cui N, Shi W, Zhu D, Jiang C. PKA-dependent activation of the vascular smooth muscle isoform of KATP channels by vasoactive intestinal polypeptide and its effect on relaxation of the mesenteric resistance artery. Biochim Biophys Acta. 2008;1778(1):88–96. doi: 10.1016/j.bbamem.2007.08.030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Yogi A, Callera GE, Hipolito UV, Silva CR, Touyz RM, Tirapelli CR. Ethanol-induced vasoconstriction is mediated via redox-sensitive cyclo-oxygenase-dependent mechanisms. Clin Sci (Lond) 2010;118(11):657–668. doi: 10.1042/CS20090352. [DOI] [PubMed] [Google Scholar]
- 81.Zhang DX, Borbouse L, Gebremedhin D, Mendoza SA, Zinkevich NS, Li R, Gutterman DD. H2O2-induced dilation in human coronary arterioles: role of protein kinase G dimerization and large-conductance Ca2+-activated K+ channel activation. Circ Res. 2012;110(3):471–480. doi: 10.1161/CIRCRESAHA.111.258871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Zimmermann AK, Loucks FA, Schroeder EK, Bouchard RJ, Tyler KL, Linseman DA. Glutathione binding to the Bcl-2 homology-3 domain groove: a molecular basis for Bcl-2 antioxidant function at mitochondria. J Biol Chem. 2007;282(40):29296–29304. doi: 10.1074/jbc.M702853200. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(PDF 164 kb)