

Abstract
Hops (Humulus lupulus L. Cannabaceae) is an economically important crop, that has drawn more attention in recent years due to its potential pharmaceutical applications. Bitter acids (prenylated polyketides) and prenylflavonoids are the primary phytochemical components that account for hops resins medicinal value. We have previously reported on utilizing untargeted NMR and MS metabolomics for analysis of 13 hops cultivars, revealing for differences in α- versus β-bitter acids composition in derived resins. In this study, effect of ratios of bitter α- to β-acids in hop resins to cytotoxicity of hop resins was investigated. In vitro cell culture assays revealed that β-acids were more effective than α-acids in growth inhibition of PC3 and HT29 cancer cell lines. Nevertheless, hop resins enriched in β-acids showed comparable growth inhibition patterns to α-enriched resins and suggesting that bioactivity may not be easily predicted by metabolomics and/or gross metabolic profiling in hops.
Keywords: Humulus lupulus L., Metabolomics, Humulones, Lupulones, Anticancer activity, Hops
Introduction
The hop plant (Humulus lupulus L., Cannabaceae) is an economically important crop cultivated in most temperate zones of the world for its female inflorescences, commonly referred to as “hop cones” or “hops”. The bitter, resinous substance produced in the glandular hairs of the strobiles (lupulin glands) is used in brewing, baking and as cattle feed for its bacteriostatic action and preservative qualities [1]. In addition, it is used in pharmaceutical applications. The resin is used as a mild sedative in European phytotherapy, and hop has been investigated for its potent estrogenic and, more recently, potential cancer chemopreventive activities [2,3]. Major class of secondary metabolites in hop lupulin glands include hop bitter acids which exhibit interesting effects on human health [4]. The hop bitter acids are resinous alicyclic phenolic acids, classified as α-acids (humulones) and β-acids (lupulones). The main α-acids are humulone, cohumulone, and adhumulone; the corresponding β-acids are lupulone, colupulone, and adlupulone (Fig. 1A). The β-acids differ structurally from the α-acids by having one extra isoprenyl group. Furthermore, hop resin contain terpenes and isoprenylated flavonoids [5–7]. There are at least 200 different hop varieties grown and cultivated worldwide and it is of increasing interest to develop accurate methods for hop characterization that could be used to classify hop from different geographical origins or countries. We have recently reported on the use of LC–MS and NMR for the metabolic fingerprinting of hop. This comparative untargeted approach revealed for compositional differences in α/β bitter acids among hop cultivars [7]. Our objective from this study was to further investigate whether differences in α- and β-bitter acids composition in hop resins could influence its cytotoxic effect. A total of 13 chemically well-characterized hop resins were tested for growth inhibition effect against (mutated androgen dependent) prostate (PC3) and (androgen independent) colon (HT-29) cancer cell lines along with standard mix of α- and β-acids. HCA multivariate data analysis was also used as an additional exploratory tool to assess the heterogeneity and relationship between the different hop cultivars.
Fig. 1.
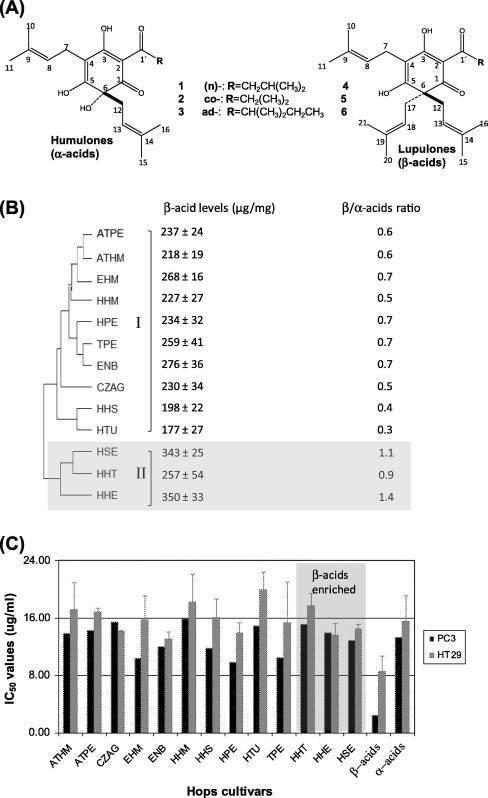
Hop α/β bitter acids composition in comparison to its cytotoxic effect. (A) Chemical structures of humulones (α-bitter acids) and lupulones (β-bitter acids) series detected in hop resin. (B) Hierarchical cluster analysis (HCA) of hop cultivars based on group average cluster analysis of its biochemical profile as the analytical data showing clustering of cultivars in 2 major groups mostly influenced by its β-bitter acids levels (μg/ml) and β/α-acid ratios. Data are mean ± SE from three independent measurements. Grey box highlights β-acids enriched resins (HSE, HHE & HHT) from other cultivars. (C) Cytotoxicity data of hops resin extracts and pure α- and β-bitter acids against human prostate (PC3) and colon (HT29) cancer cell lines (IC50 values expressed in μg/ml). Detailed description of bitter acids standards composition is provided under materials and methods. Data are mean ± SE from four independent experiments. Note the grey box highlighting β-enriched resins (HSE, HHE & HHT) showed no significant difference in its IC50 values from other samples.
Material and methods
Plant material
The 13 different hop resins included in this study were provided by Hopsteiner (Mainburg, Germany). All information on collected samples and their origin is recorded in Table 1. Resins were obtained by standard extraction with ethanol. A detailed description of the resin preparation is given by [8].
Table 1.
Origins of hops cultivars harvested in 2009.
| Cultivar | Region, country | Abbreviation |
|---|---|---|
| Hallertau Perle | Hallertau, Germany | HPE |
| Hallertau Hallertauer Tradition | Hallertau, Germany | HHT |
| Hallertau Hersbrucker | Hallertau, Germany | HHE |
| Hallertau Herkules | Hallertau, Germany | HHS |
| Hallertau Hallertauer Magnum | Hallertau, Germany | HHM |
| Hallertau Spalter Select | Hallertau, Germany | HSE |
| Hallertau Hallertauer Taurus | Hallertau, Germany | HTU |
| Elbe-Saale Magnum | Elbe-Saale, Germany | EHM |
| Elbe-Saale Northern Brewer | Elbe-Saale, Germany | ENB |
| Tettnang Perle | Tettnang, Germany | TPE |
| Mühlviertel Magnum | Mühlviertel, Austria | ATHM |
| Mühlviertel Perle | Mühlviertel, Austria | ATPE |
| Saaz Agnus | Saaz, Czech Republic | CZAG |
Chemicals and reagents
Standard for α-acids mixture (30.06% cohumulone and 69.93% humulone + adhumulone) and β-acids mixtures (47.95% colupulone and 52.05% lupulone + adlupulone) were provided by Hopsteiner (Mainburg, Germany). All other chemicals and standards were provided by Sigma Aldrich (St. Louis, MO, USA).
HCA analysis of LC/MS dataset
Quantified hop metabolites performed using XCMS data analysis software [9] was imported into the R 2.9.2 software package using custom-written procedures for hierarchical clustering analysis (HCA) to visualize for general clustering trends. Absolute LC/MS peak area values were autoscaled (the mean area value of each feature throughout all samples was subtracted from each individual feature area and the result divided by the standard deviation) prior to clustering analysis. This provides similar weights for all the variables. Detailed description on bitter acids analysis and quantification methodology is provided in [7].
Cell lines and culture conditions
Human prostate cancer cell line, PC3, was obtained from the Deutsche Sammlung von Mikroorganismen und Zellkulturen, Braunschweig, (DMSZ ACC# 465) and the colon cancer cell line, HT29, was obtained from the medical immunology department at Martin Luther-Universität Halle-Wittenberg (Prof. Seliger). The cells were grown as monolayers in adherent cell lines and were routinely cultured in RPMI (Roswell Park Memorial Institute) 1640 supplemented with 10% heat-inactivated fetal bovine serum (FBS) and 1% l-glutamine in 75 cm2 polystyrene flasks (Corning Life Sciences, UK) and maintained at 37 °C in a humidified atmosphere with 5% CO2.
Cytotoxicity assay
Cells were plated at a density of 1 × 104/well in 96-well plates. They were allowed to attach to the plate for 24 h. After 24 h, the media were replaced with RPMI media containing resin extracts. Four concentrations of each resins were tested (1, 5, 10 and 20 μg/ml). Resins were initially dissolved in DMSO at a concentration of 2 mg/ml and further diluted with RPMI medium. The DMSO concentration in the assay did not exceed 0.1% and was not cytotoxic to the tumor cells. After 72 h, the growth medium was taken out and 100 μl of XTT-solution (2,3-bis (2-methoxy-4-nitro-5-sulfophenyl)-5-[(phenylamino) carbonyl]-2H-tetrazolium hydroxide) (Roche Applied Science, Mannheim, Germany) was added to each well, and plates were incubated at 37 °C for another 4 h at a (final concentration 0.3 mg/ml). Absorbance was measured at 490 nm against a reference wavelength at 650 nm using a microplate reader (Beckman Coulter, DTX 880 Multimode Reader). The mean of four experiments for each dose was used to calculate the IC50 and repeated in 2 passages for each cancer cell line. Digitonin was used as a standard cytotoxic agent with an IC50 value of 1.7 μg/ml. IC50 values were calculated with GraphPad Prism version 5 software, using sigmoidal dose–response function.
Results and discussion
Hierarchical cluster analysis (HCA) of hop resins
The major goal of this study was to investigate the effect of hop bitter acids compositional differences on hop resin cytotoxic effect. To accomplish this goal, we have selected a study group of resins derived from 13 hop cultivars, 10 of which originated from Germany and 3 others originated from Austria and the Czech Republic (Table 1). Previous LC–MS and NMR analyses revealed the compositional differences in α and β-bitter acids among resins derived from different hop cultivars [7]. Hop cultivars HHS, HTU & HHM were found to be enriched in α-acids, whereas higher levels of β-acids were determined in cultivar HHE, and to less extent in HSE and HHT as confirmed by absolute metabolites quantification [7].
Hierarchical cluster analysis (HCA) is an unsupervised data analysis method that allows interpretation of metabolites results in a fairly intuitive graphical way without prior knowledge of the sample composition. In this study, HCA of the different hop resins based on quantified data from a total of 29 metabolites described by Farag et al. [7] was used as an additional exploratory tool to assess the heterogeneity among the different hop cultivars. HCA showed two clear major clusters, of 10 and 3 cultivars (Fig. 1B), referred to as groups I and II, respectively. Clustering pattern was mostly influenced by differences in bitter acids levels among cultivars as revealed from density of corresponding signal in the heatmap plot (data not shown). Inspection of group II showed that HHE, HHT and HSE cultivars are more closely related as in both lupulone type β-acids were enriched (260–350 μg/mg resin dry wt.) and in agreement with PCA results [7]. Cluster I included all other cultivars having a lower levels in β-acids reaching an average of 170 to 260 μg/mg resin dry wt. Interestingly, both ATHM and ATPE cultivars that were collected from the same geographical area/region (Austria) clustered together (Fig. 1B) suggesting that in hop, geographical origin can be reflected in its bitter acids composition.
It is hypothesized that in natural plant systems, metabolic pathways contain biosynthetic modules, which lead to the formation of metabolites (groups of metabolites whose production is co-regulated and biosynthetically linked). To test whether such co-regulated metabolite modules do exist in hops and produce this myriad of phytochemicals, HCA analysis was also performed (Supplementary Fig. 1) with Pearson correlation coefficients calculated for all pairs of 29 metabolites identified in our analysis [7]. The HCA results and ‘correlation heatmaps’ clearly show the existence of modules of co-regulated metabolites in hops, supporting the hypothesis that biosynthetic modules do indeed exist in natural plant systems [10]. Metabolites within the same module had abundance patterns across cultivars that were highly correlated with each other, and they had similar relationships to other compounds. Importantly, one of the many co-regulated metabolite modules that were readily detected in this clustering analysis contained the unprenylated flavonoids i.e. quercetin and kaempferol (module B) whereas prenylated metabolites belonged to separate metabolite modules. The presence of α-bitter acids in a separate module (module C) from β-forms (module D) suggests the presence of a distinct isoprenyl transferase in hops catalyzing for the attachment of an extra isoprenyl group (Fig. 1) as the last step of β-acids biosynthesis. Cloning and or functional characterization of isoprenyl transferases involved in bitters acids biosynthesis has yet to be achieved. It should be noted that β-bitter acids oxidative products i.e. cohulupone and hulupone were clustered in a separate metabolite (module A) along with other unknown metabolites. This is the first report on metabolites modules existence in hop plants.
Hop resin cytotoxic effect in relation to α/β bitter acid levels
Increasing evidence in the literature points towards the marked cytotoxic effect found for hop bitter acids in addition to the well established effects of xanthohumol and 8-prenylnaringenin [11,4]. Analyses of resins in the current study show that xanthohumol was present at much lower levels compared to bitter acids (ca. 30 ug/mg resin dry wt.) whereas 8-prenylnaringenin was found at trace levels. Recently, both humulone and lupulone isolates were found to inhibit cancer cell growth with lupulone exhibiting lower IC50 values against lung and breast cancer cell lines [12]. Cytotoxic effect in hop is mediated via angiogenesis, inducing apoptosis, and by increasing the expression of cytochrome P450 detoxification enzyme [4]. Our objective was to investigate cytotoxic effect of hop resins derived from the 13 cultivars with different α-/β-acids composition as revealed from our HCA analysis. The 13 hop resins were tested for growth inhibition effect against (mutated androgen dependent) prostate (PC3) and (androgen independent) colon (HT29) cancer cell lines along with standard mix of α- and β-acids. IC50 values exhibited are presented in Fig. 1C. All hop resin extracts inhibited cancer cell growth with comparable IC50 values ranging from 9 to 20 μg/ml for HT29 and PC3 cell lines. Both α/β bitter acid standards exhibited high cytotoxic activities against both PC3 and HT29 cells. However, IC50 values for β-acids (lupulones: PC3, 2.4 μg/ml; HT29, 8.1 μg/ml) were significantly lower than the α acids (humulones: PC3, 13.2 μg/ml; HT29, 15.5 μg/ml). These results were in agreement with previous results [12]. It is unclear though, whether differences are based on difference in lipophilicity due to extra isoprenyl group in β-acids (and thus cell uptake), or rather a specific target effect. Interestingly, prenylation of apigenin and liquiritigenin flavonoids was found to enhance its cytotoxicity against rat H4IIE hepatoma and C6 glioma cells [13]. Nevertheless, cultivars such as HSE, HHE and HHT enriched in β-acids (clustered separately in group II, Fig. 1B) did not exhibit a lower IC50 values against both cell lines compared with other cultivars. This might be attributed to the small differences between α- and β-acids levels among cultivars. In addition, other compounds of higher specific activity i.e. xanthohumol and 8-prenyl naringenin (8-PN) present in the hop matrix [14–16] may act additively or synergistically, and may eventually be more relevant for the cytotoxic or specificity effects of hop resin extracts than just the high concentrations (75% resin content) of bitter acids. These results suggest that anticancer activity may not be easily predicted by multivariate data analysis of metabolites composition in hop. In contrast to the compound related differences, there is a consistent differentiation according to the cell type, with PC3 being more sensitive than HT29, which is opposite to the common response of these cell lines on cytotoxic compounds. The origin of this trend is unclear, but PC3 cells are more responsive to steroid mimetics as found in hop. In conclusion, our results confirm the potential health benefits of hops in chemoprevention, however no correlation could be observed between differences in hop bitter acids composition and anticancer effect among hop cultivars and suggesting that bioactivity may not be easily predicted by bitter acids profiling in hops.
Acknowledgments
We are grateful to Hopsteiner Inc. (and here especially Dr. Martin Biendl, Halletau and Harald Schwarz) for hop resins. Dr. M.A. Farag thanks the Alexander von Humboldt foundation, Germany for financial support.
Footnotes
Peer review under responsibility of Cairo University.

Appendix A. Supplementary material
References
- 1.Zhang X.Z., Liang X.M., Xiao H.B., Xu Q. Direct characterization of bitter acids in a crude hop extract by liquid chromatography–atmospheric pressure chemical ionization mass spectrometry. J Am Soc Mass Spectr. 2004;15:180–187. doi: 10.1016/j.jasms.2003.09.014. [DOI] [PubMed] [Google Scholar]
- 2.Chadwick L.R., Nikolic D., Burdette J.E., Overk C.R., Bolton J.L., van Breemen R.B. Estrogens and congeners from spent hops (Humulus lupulus) J Nat Prod. 2004;67:2024–2032. doi: 10.1021/np049783i. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Lamy V., Roussi S., Chaabi M., Gosse F., Schall N., Lobstein A. Chemopreventive effects of lupulone, a hop beta-acid, on human colon cancer-derived metastatic SW620 cells and in a rat model of colon carcinogenesis. Carcinogenesis. 2007;28:1575–1581. doi: 10.1093/carcin/bgm080. [DOI] [PubMed] [Google Scholar]
- 4.Van Cleemput M., Cattoor K., De Bosscher K., Haegeman G., De Keukeleire D., Heyerick A. Hop (Humulus lupulus)-derived bitter acids as multipotent bioactive compounds. J Nat Prod. 2009;72:1220–1230. doi: 10.1021/np800740m. [DOI] [PubMed] [Google Scholar]
- 5.Arraez-Roman D., Cortacero-Ramirez S., Segura-Carretero A., Contreras J.A.M.L., Fernandez-Gutierrez A. Characterization of the methanolic extract of hops using capillary electrophoresis–electrospray ionization-mass spectrometry. Electrophoresis. 2006;27:2197–2207. doi: 10.1002/elps.200500714. [DOI] [PubMed] [Google Scholar]
- 6.Li H.J., Deinzer M.L. Structural identification and distribution of proanthocyanidins in 13 different hops. J Agric Food Chem. 2006;54:4048–4056. doi: 10.1021/jf060395r. [DOI] [PubMed] [Google Scholar]
- 7.Farag M.A., Porzel A., Schmidt J., Wessjohann L. Metabolite profiling and fingerprinting of commercial cultivars of Humulus lupulus L. (hop): a comparison of MS and NMR methods in metabolomics. Metabolomics. 2012;8:492–507. [Google Scholar]
- 8.Biendl M. Hops and health uses – Effects – History; 2008.
- 9.Smith C.A., Want E.J., O’Maille G., Abagyan R., Siuzdak G. XCMS: processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal Chem. 2006;78:779–787. doi: 10.1021/ac051437y. [DOI] [PubMed] [Google Scholar]
- 10.Xie Z.Z., Ma X.Q., Gang D.R. Modules of co-regulated metabolites in turmeric (Curcuma longa) rhizome suggest the existence of biosynthetic modules in plant specialized metabolism. J Exp Bot. 2009;60:87–97. doi: 10.1093/jxb/ern263. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Chen W.J., Lin J.K. Mechanisms of cancer chemoprevention by hop bitter acids (Beer aroma) through induction of apoptosis mediated by fas and caspase cascades. J Agric Food Chem. 2004;52:55–64. doi: 10.1021/jf034737u. [DOI] [PubMed] [Google Scholar]
- 12.Tyrrell E., Archer R., Skinner G.A., Singh K., Colston K., Driver C. Structure elucidation and an investigation into the in vitro effects of hop acids on human cancer cells. Phytochem Lett. 2010;3:17–23. [Google Scholar]
- 13.Watjen W., Weber N., Lou Y.J., Wang Z.Q., Chovolou Y., Kampkotter A. Prenylation enhances cytotoxicity of apigenin and liquiritigenin in rat H4IIE hepatoma and C6 glioma cells. Food Chem Toxicol. 2007;45:119–124. doi: 10.1016/j.fct.2006.08.008. [DOI] [PubMed] [Google Scholar]
- 14.Vogel S., Heilmann J. Synthesis, cytotoxicity, and antioxidative activity of minor prenylated chalcones from Humulus lupulus. J Nat Prod. 2008;71:1237–1241. doi: 10.1021/np800188b. [DOI] [PubMed] [Google Scholar]
- 15.Deeb D., Gao X., Jiang H., Arbab A., Dulchavsky S., Gautam S. Growth inhibitory and apoptosis-inducing effects of xanthohumol, a prenylated chalone present in hops, in human prostate cancer cells. Anticancer Res. 2010;30:3333–3339. [PMC free article] [PubMed] [Google Scholar]
- 16.Delmulle L., Bellahcene A., Dhooge W., Comhaire F., Roelens F., Huvaere K. Anti-proliferative properties of prenylated flavonoids from hops (Humulus lupulus L.) in human prostate cancer cell lines. Phytomedicine. 2006;13:732–734. doi: 10.1016/j.phymed.2006.01.001. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.