

Abstract
Task-switch costs and in particular the switch-cost asymmetry (i.e., the larger costs of switching to a dominant than a non-dominant task) are usually explained in terms of trial-to-trial carry-over of task-specific control settings. Here we argue that task switches are just one example of situations that trigger a transition from working-memory maintenance to updating, thereby opening working memory to interference from long-term memory. We used a new paradigm that requires selecting a spatial location either on the basis of a central cue (i.e., endogenous control of attention) or a peripheral, sudden onset (i.e., exogenous control of attention). We found a strong cost asymmetry that occurred even after short interruptions of otherwise single-task blocks (Exp. 1-3), but that was much stronger when participants had experienced the competing task under conditions of conflict (Exp. 1-2). Experiment 3 showed that the asymmetric costs were due to interruptions per se, rather than to associative interference tied to specific interruption activities. Experiment 4 generalized the basic pattern across interruptions varying in length or control demands and Experiment 5 across primary tasks with response-selection conflict rather than attentional conflict. Combined, the results support a model in which costs of selecting control settings arise when (a) potentially interfering memory traces have been encoded in long-term memory and (b) working-memory is forced from a maintenance mode into an updating mode (e.g., through task interruptions), thereby allowing unwanted retrieval of the encoded memory traces.
Keywords: Attention, Executive Control, Task Switching
The way we interact with the world is contingent on abstract control settings. These settings specify which external or internal information is currently relevant and how to act upon it in order to achieve one's goals. From research with the task-switching paradigm, in which people are prompted to switch between predefined task rules on a trial-by-trial basis, we know that it is difficult to flexibly change between task or control settings (for reviews see Monsell, 2003; Logan, 1993; Kiesel, Steinhauser, Wendt, Falkenstein, Jost, et al., 2010; Vandierendonck, Liefooghe, & Verbruggen, 2010). From this research we can also derive two fundamentally different accounts of how exactly these obstacles to flexible change arise. By the first, and intuitively most appealing account, costs of switching between tasks or control settings come from the direct clash between the residue of the most-recently used and the currently relevant task setting (e.g., Allport, Styles, & Hsieh, 1984; Gilbert & Shallice, 2002; Yeung & Monsell, 2003a,b). In contrast, the second account holds that interference between competing task settings is not the result of carry-over from the most-recent past, but rather reflects the long-term memory (LTM) knowledge base about the space of tasks involved in a particular context (e.g., Bryck & Mayr, 2008; Mayr, 2009; Wazsak, Hommel, & Allport, 2003).1 In the work described here, we examine which of these two accounts is better suited to explain the costs of selecting and changing control settings. For this purpose, we focus mainly on the important, yet understudied problem of selecting between endogenous versus exogenous control over spatial attention. Before we elaborate on our choice of control settings, we first develop our general theoretical and empirical approach.
Switch-Cost Asymmetry and Carry-Over of Task Sets
A benchmark result in the task-switching literature is the so-called switch-cost asymmetry. When people switch between a dominant task, such as Stroop word naming and a competing, non-dominant task, such as Stroop color naming, switch costs are larger when transitioning from the hard, non-dominant to the easy, dominant task than the other way round (e.g., Allport, Styles & Hsieh, 1984). This phenomenon is important here because carry-over models of task switching seem to be able to explain it in a straightforward manner: Non-dominant tasks require a particularly strong attentional setting to survive against the competition from the dominant task and this strong setting is carried forward into the next trial where it needs to be overcome when switching back to the dominant task. In contrast, the dominant task requires only weak support from a task setting and therefore relatively speaking, less change in control settings is required when switching from the dominant to the non-dominant task. Critically, for the carry-over account to work, trial-to-trial switching between the two competing tasks is a necessary condition for obtaining a switch cost asymmetry (Gilbert & Shallice, 2002; Yeung & Monsell, 2003a,b).
Even though this model adequately accounts for the basic finding of the asymmetry in switch costs, there is also some initial evidence that directly contradicts the carry-over account. Obviously, the carry-over account can explain the task-selection cost asymmetry only for cases in which the alternative task was performed in the immediately preceding trial—otherwise there would be no opportunity for carry-over. However, Bryck and Mayr (2008) have shown that a cost asymmetry can be obtained even in the absence of a task-switch transition (see also Allport & Wylie, 2000). This finding, which will be elaborated below, is important because it indicates that opportunity for trial-to-trial carry-over is not a necessary condition for the cost asymmetry to arise.
LTM Encoding and Interruption-based Interference
A key tenet of our account is that interference comes not from the most recent past (i.e., the previous trial), but from any kind of previous experiences with the competing tasks that are stored in long-term memory. One long-term memory model that is particularly well-equipped to handle the influence of past task experiences is memory instance theory (Hintzman, 1986; Logan, 1988). To fully explain task-selection costs, this theory needs to be augmented through additional assumptions about factors that affect encoding and retrieval of memory instances.
Automatic Encoding and Retrieval of Selection Instances
Logan (1988) has proposed that during episodes of selecting an appropriate response in a given task, selection-relevant features are automatically encoded and stored as integrated instances in LTM. Future, similar selection episodes can trigger automatic retrieval of these memory traces. Depending on the degree of match between the retrieved and current response demands this can then either lead to processing benefits or costs. While Logan (1988, 1990) had originally examined the encoding and retrieval of relatively simple contextual features, such as the location of a task-relevant object, more recent work has demonstrated that also abstract control settings (i.e., task sets) are automatically encoded in LTM. This is an important extension of instance theory because it can explain how even supposedly high-level, executive processes can come under automatic, memory-driven control (e.g., Crump & Logan, 2010; Mayr & Bryck , 2005, 2006; Verbruggen & Logan, 2010). In fact, there is evidence that in task-switching situations LTM retrieval of past selection instances can play a substantial role. For example, using picture-naming/word-reading tasks, Wazsak et al. (2003) showed that switch costs to the dominant word-reading task were substantially larger with picture-word constellations that had been also used in the picture-naming task—even if that experience occurred over 100 trials in the past (see also Mayr & Bryck, 2005; Bryck & Mayr, 2008).
Interruptions Trigger Interference/Retrieval
The assumption of automatically encoded memory instances alone does not explain the cost asymmetry. We need additional assumptions that explain why such interference may be particularly strong when switching to the dominant task. Biologically plausible models suggest that working memory can take on two qualitatively distinct modes, one geared towards short-term information maintenance, the other enabling updating of current working memory content. The maintenance mode supports preserving the current representation in a robust manner, thus allowing little effect of interference from the environment or LTM. In contrast, during the updating mode working memory is open towards external or internal influences, thus allowing a context-influenced search for new, stable representations (e.g., Durstewitz, Kelc, & Güntürkün, 1999, O'Reilley, 2006). In this state, the system should be maximally sensitive to interference from selection instances that are related to the current stimulus situation.
Given that in the maintenance mode working memory is shielded from information that does not fit to the current representation there is a danger of behavioral rigidity. Therefore, even in the maintenance mode the system needs to remain sensitive to low-level signals or events indicating that a change may be necessary. For example, abrupt onsets (e.g., Theeuwes, Kramer, Hahn, & Irwin, 1998), a perceptual change in task cue (Mayr, 2006), presence of information-processing conflict (Botvinick, Braver, Barch, Carter, & Cohen, 2001), or signals that have been linked with the need for change via associative learning (O'Reilley, 2006), can all switch working memory into an updating state. This also implies that the maintenance mode should be relatively effective as long as one can focus on an overlearned, or otherwise dominant task, and as long as none of the just-mentioned events occur. In this case, little to no interference from competing non-dominant tasks should be expected. However once disrupted, the system needs to go through an updating process before the maintenance mode can be re-established. During this phase, even performance of a dominant task is highly sensitive to interference from potential memory instances involving the competing task. In contrast, while performing non-dominant tasks, “hard-wired” or overlearned, competing response tendencies can produce low-level signals that can trigger updating attempts (Botvinick et al., 2001). Thus here, the distinction between updating and maintenance is less crisp and the costs of re-establishing the non-dominant task (i.e., after a switch) may be small, relative to the relatively pure difference between updating and maintenance for the dominant task.
According to this explanation, the back-and-forth switching between dominant and non-dominant tasks is not a necessary condition for the cost-asymmetry to arise. Instead, the following two conditions are necessary. Condition 1: Subjects working on the dominant task need to have experience with the competing, non-dominant task so that LTM contains potentially interfering memory traces. Condition 2: There need to be events that interrupt maintenance of the dominant task set so that an updating process becomes necessary, which in turn allows competing LTM traces to interfere with ongoing processing. In the standard switching paradigm, subjects constantly experience both tasks and each task switch enforces an updating operation. However in theory, any exogenous or endogenous event that interrupts maintenance should suffice to produce a cost asymmetry as long as LTM contains memory traces from competing tasks.
In the earlier-mentioned experiments reported by Bryck and Mayr (2008), the two above conditions were met, without requiring subjects to switch between tasks in a trial-by-trial manner. Most relevant here is Experiment 3: Subjects in the experimental group performed alternating pure-task blocks of either only Stroop color or word naming. Thus, within the session, participants in the experimental group experienced both tasks, without ever directly transitioning between them (i.e., Condition 1). We also varied the response-stimulus interval randomly between 50 and 5000 ms. The idea behind this manipulation was that long delays increase the probability of losing the current maintenance state and as a consequence trigger an updating process to re-establish the relevant task from LTM (i.e., Condition 2). As a control we used groups in which participants worked either only with Stroop color or Stroop word task blocks throughout the entire session. As predicted, we found that in the long-RSI condition, Stroop color naming RTs were particularly long, but only for participants who had experienced both tasks. This constitutes a cost-asymmetry pattern in the absence of an actual switch in task.
Selecting Between Endogenous and Exogenous Control of Attention
Typically, the switch-cost asymmetry is assessed using task combinations with mutual response conflict, such as Stroop word reading and color naming. In all, except for one of the experiments we report here (Experiment 5), we instead decided to focus on conflict between endogenous and exogenous control of spatial attention.
The distinction between exogenous and endogenous control is an important one in the study of attention. Experimentally, endogenous control is typically induced through symbolic cues (e.g., central arrow) and it is relatively slow-acting, and often effortful. In contrast, exogenous control allows fast orienting responses to sudden onsets in the visual field, requires no effort, but is also difficult or even impossible to resist (e.g., Jonides, 1981; Müller & Rabbitt, 1989; Posner, 1980). While these two modes of attentional control (and the differences between them) have been studied extensively, the question we address here, namely how we select between exogenous and endogenous control, has not been addressed explicitly.
What makes the examination of these two modes of control particularly useful in the current context is the fact that we can assume a very strong dominance asymmetry between exogenous and endogenous control. Exogenously controlled attention is a reflex-like, hard-wired process that strongly interferes with endogenous control, but should be unaffected by stimuli associated with endogenous control of attention. At least this is what we expect as long as subjects reside in the maintenance mode. However, during the updating mode (i.e., after recovering from an interruption), even exogenous control of attention may become vulnerable. It would be a particularly striking and novel result if we can create an experimental situation in which people have difficulties reacting appropriately to the very same stimuli that under typical circumstances elicit reflex-like responses. According to our model, such a situation should occur when people are both forced into an updating mode and when LTM contains memory traces from competing tasks (i.e., the endogenous control task). More generally, we believe that the contrast between endogenous and exogenous control of attention establishes a particularly crisp, empirical dissociation between updating (i.e., re-establishing exogenous control after interruptions) and maintenance (i.e., responding to abrupt onset stimuli once the exogenous control mode has been re-established).
The Current Research
In the earlier mentioned Bryck and Mayr (2008) study the distinction between updating and maintenance states was relatively subtle and not in all cases significant. Therefore, instead of using a manipulation in terms of long versus short inter-trial delays, we occasionally interrupted pure, dominant or non-dominant task blocks with a third, “neutral” task.2 The idea here is that recovery from such an interruption necessarily requires a working memory updating process. In contrast, in the absence of such interruptions maintenance and shielding against interference should be maximized, at least while performing the dominant, exogenous task.
While the model we describe above can explain in principle how a cost asymmetry might arise in the absence of opportunity for trial-to-trial carry-over, it remains under-specified in important ways. In particular we had stated that “sufficient experience” with alternative tasks is necessary to create potentially competing memory traces. However, we do not know what exactly constitutes such sufficient experience. For example, Bryck and Mayr (2008) had speculated that encoding of non-dominant task LTM traces may be a function of how much attention is devoted to performing that task. In turn, the amount of attention devoted to the task may be a function of the presence of conflict from alternative tasks during the encoding situation. In other words, experience with the competing task alone may not be sufficient. Rather, such experience may require the presence of conflict (see also Verguts & Notebaert, 2009). Therefore, in Experiments 1 and 2, we will explore the role of conflict during encoding in some detail.
An important aspect of our model is the “structural” hypothesis that there is something special about the abstract category of “interruptions of the maintenance state” that creates opportunity for interference. Therefore, in Experiment 3 we attempted to rule out an alternative possibility, namely that associative interference (akin to the fan effect) between specific interrupting activities and competing tasks is the main source of the between-task interference.
Finally, in Experiments 4 and 5 we attempted to generalize the critical pattern of results along two dimensions. In Experiment 4, we manipulated the control demands of the interruption task. In Experiment 5, we exchanged the exogenous/exogenous attention tasks for a pair of tasks with mutual response conflict.
Experiment 1
Figure 1 presents our basic paradigm that pits endogenous and exogenous control of attention against each other. In this, as in all other experiments subjects only performed pure bocks of either the endogenous or the exogenous control task. No matter what the task, subjects had to make a left/right key press to the letter L or R shown within one of the six stimulus frames in a large circular array (i.e., the target circle). In the center of that array there was a much smaller arrangement of cue circles, corresponding to the large circular array. During the response-stimulus interval each of these cue circles was shown in red. With stimulus onset, all but one of the peripheral small circles turned white, leaving the one remaining red, small circle as a central cue. In addition, as a sudden-onset stimulus, an additional, large red circle could appear between two of the regularly arranged stimulus circles. On high-conflict trials, the display contained information associated with the currently irrelevant control mode (both the sudden onset and the central cue were presented) whereas on low-conflict trials only the currently relevant information was presented (either the sudden onset or the central cue). In pilot work, we found that switching between endogenous and exogenous control on a trial-by-trial manner indeed leads to a strong switch-cost asymmetry.3
Figure 1.
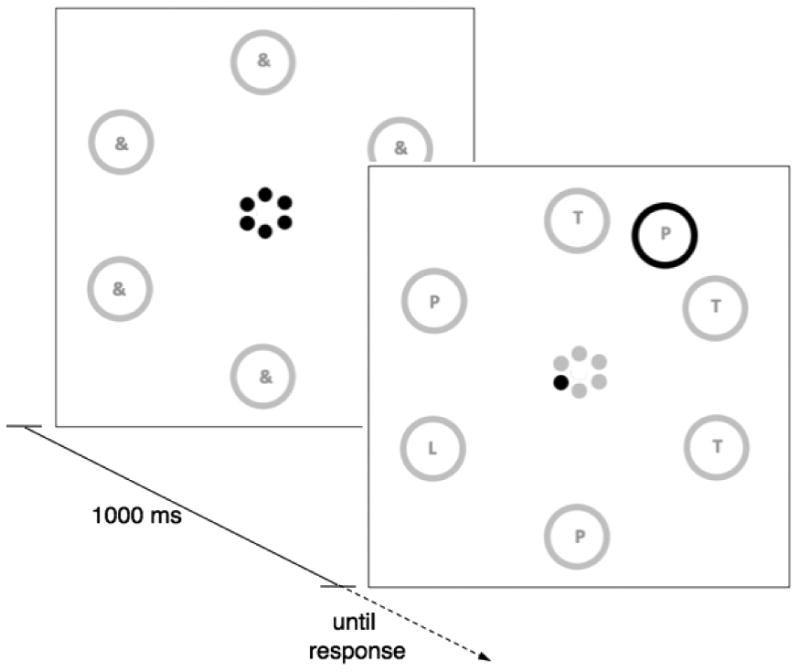
Stimulus setup used in the present experiments. Shown is an endogenous-task trial where participants use the center cue to locate the outer circle with the response-relevant information (i.e., L vs. R), while ignoring the sudden-onset stimulus in the periphery (if presented). For exogenous-task trials, subjects would have to locate the sudden-onset stimulus while ignoring the central-cue information (if presented). Stimuli are not drawn to scale; gray was presented as white on black background, black was presented as red.
The first prediction we tested in Experiment 1 is that a cost asymmetry can be obtained even when there is no trial-to-trial switching between competing tasks. Therefore the critical experimental group alternated between pure endogenous and pure exogenous 80-trial blocks. Performance in these blocks was interrupted occasionally (p=.25) by math equation trials (see Figure 2). On these trials, a math equation was presented instead of the regular displays and participants had to respond with a correct/incorrect judgment. After an interruption trial, the block continued with the main task relevant in that block. We assumed that on trials that follow an interruption a process of (re-)updating the current task set needs to happen, which in turn allows interference from the competing task. Thus, for post-interruption trials we predicted a cost asymmetry. Once updating has occurred, subject should experience little ambiguity about which task is currently relevant, thus allowing robust maintenance. Therefore, on these maintenance trials (i.e., all trials following post-interruption trials prior to the next interruption) we expected to see little evidence of interference, at least for the dominant task. With the presence of interruption events one critical condition for the cost asymmetry is met, as these allow interference from LTM during the post-interruption updating operation. A second condition is that participants actually had an opportunity to form LTM memory traces about both types of tasks/control settings. Therefore, aside from the experimental groups, which alternated between endogenous and exogenous blocks, we included as controls two groups of subjects which either only worked on endogenous or only on exogenous tasks throughout the entire experiment. These conditions allowed us to obtain baseline estimates of the size of the post-interruption costs and interference effects when no LTM traces of the competing task were available.
Figure 2.
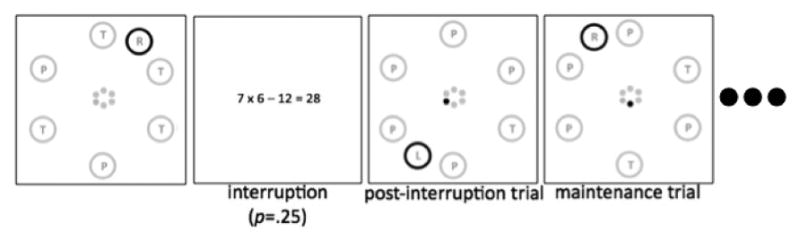
Example sequence of exogenous-task trials. Both the post-interruption trial and the maintenance trial contain conflict from the endogenous stimulus, whereas the pre-interruption trial contains no conflict.
The second prediction we wanted to test is that it is not just experience with competing tasks that drives encoding of interfering LTM traces, but that the experienced selection episodes need to include high levels of conflict. In the critical condition described so far, half of the trials contained conflict from the alternate task (i.e., a singleton distractor for the exogenous task and an endogenous cue for the exogenous task). We added two additional between-subject conditions, one of which contained 50% conflict trials for the exogenous task blocks, but no conflict trials for the endogenous blocks and the other contained conflict trials for the endogenous task, but not for the exogenous task. If experience of conflict is a necessary condition for encoding of interfering LTM traces then we expect the elimination of the cost asymmetry here, in particular for the condition without endogenous-task conflict.
Methods
Participants
Eighty students the University of Oregon participated in exchange for course credits.
Stimuli
Participants were seated 50 cm from the computer display. Figure 1 presents the basic stimulus setup used across all experiments. Six circular stimulus frames (diameter of each circle = 13 mm = 1.5 degrees) were presented on a virtual circle (diameter = 14 cm = 16 degrees) around the screen's center. These circles where always presented in white on a black background. Within each circle the “&” symbol or the letters L, R, P, T could be presented in white, size 12 Times font. An additional, sudden-onset circle of the same size could appear between two of the regular circle positions. This circle was always presented in red and could also contain the letters L, R, P, or T also in white, size 12 Times font. At the center of the screen there were six smaller “cue circles”, (diameter of each circle= 4 mm = .5 degrees). These were arranged in a way that mirrored the larger set of stimulus circles (diameter of the central cue circle= 14 mm = 1.6 degrees), such that for each position in the larger stimulus circle, there was a corresponding, smaller cue circle. The smaller cue circles could be presented either in white or red.
Tasks
Conditional on specific conditions, participants were asked to perform either only the “endogenous” or the “exogenous” task throughout a particular block. In exogenous, single-task blocks, each trial began with a 1000 ms inter-trial period in which the large peripheral circles all contained the “&” symbol and all central cue circles were red. Next the response-relevant stimulus was presented in form of a sudden-onset circle, containing either the letter “L” or “R”. Participants had to press the left-arrow key for the letter “L” and the right-arrow key for the letter “R”. The large stimulus circles contained either the letters “P” or “T”, to which no response was assigned. Depending on condition, no-conflict or conflict trials were presented. For no-conflict trials, all of the red cue circles were replaced by white circles. In 50% of trials, one of the cue circles remained red (i.e., conflict trials). These stimuli were presented until a response was executed.
The stimulus sequence for endogenous, single-task blocks differed from that of exogenous blocks only in that on each trial a red cue circle was left standing during presentation of the response-relevant stimulus. This circle pointed to one of the peripheral large circles, which contained either an “L” or an “R” to which participants could respond (whereas all other circles contained “P”s or “T”s). In addition, on 50% of trials the red sudden onset circle was presented, containing either a “P” or a “T” (i.e., conflict trials) whereas on the other 50% no such stimulus was shown (i.e., no-conflict trials).
Subjects only worked with endogenous or exogenous single-task blocks. The exact combination of tasks and the presence of conflict were manipulated across between-subject conditions. Twenty participants each were randomly assigned to one of four conditions. The between-subject control condition was further divided into two groups of 10 subjects each. The “pure endo” group performed only the endogenous task throughout the entire experimental session whereas the “pure exo” group performed only the exogenous task. Conflict from the non-relevant task was presented randomly with p=.5.
In the main experimental condition, the “exo/endo” condition, participants alternated between endogenous and exogenous task blocks. Conflict from the currently irrelevant task could occur with probability of p=.5. The “exo/endo–noconflict” condition was identical to the exo/endo condition, only that while performing the endogenous task, subjects never experienced conflict from exogenous stimuli. Finally, the “exo–noconflict/endo” condition was again identical to the exo/endo condition, except that subjects never experienced endogenous-task conflict while performing the exogenous task.
In addition, in all blocks single-task performance was interrupted by a math task. For these trials, the standard stimulus display disappeared and instead, an equation of the type “7*8-24=32” was shown, positioned at the center of the screen (Times font, size=24). Problems were constrained to produce solutions in the positive range. Participants used the arrow keys to indicate whether the equation was correct or incorrect (left key=incorrect, right key=correct). The probability of correct equations was p=.5. Incorrect equations were off by +/-1 or 2. Immediately after responding the next endogenous or exogenous-task stimulus display appeared. For each trial, the probability of a number task was p=.25, with the constraint that two number trials could not occur consecutively. In case of either primary-task or interruption-task errors a short error tone occurred.
Procedure
In the between-subject control condition, subjects began with one 80-trial practice block; in the remaining conditions with alternating task blocks, subjects began with two 80-trial practice blocks, one for each task and with the order counterbalanced across subjects. Practice blocks were in all aspects identical to the actual test blocks. Then followed eight additional blocks, either of the same task (in the between-subject control condition) or alternating between the two tasks. For the alternating condition, onscreen instructions prior to each block indicated the currently relevant task.
Results and Discussion
We excluded all error trials and non-math trials with RTs larger than 4000 ms. Trials after errors were also excluded, no matter whether they were primary task errors or errors in the interruption task. In no condition of the primary tasks did error rates exceed 3.9% and in no instance did the pattern of error effects counteract the pattern of RTs. Therefore, in our analysis we focus on RTs, but we do present the error results in Figure 3 along with the RT results. For the interruption task, the mean error rate was 11.89% (SD=8.76) and the mean RT was 3667 ms (SD=1008).
Figure 3.
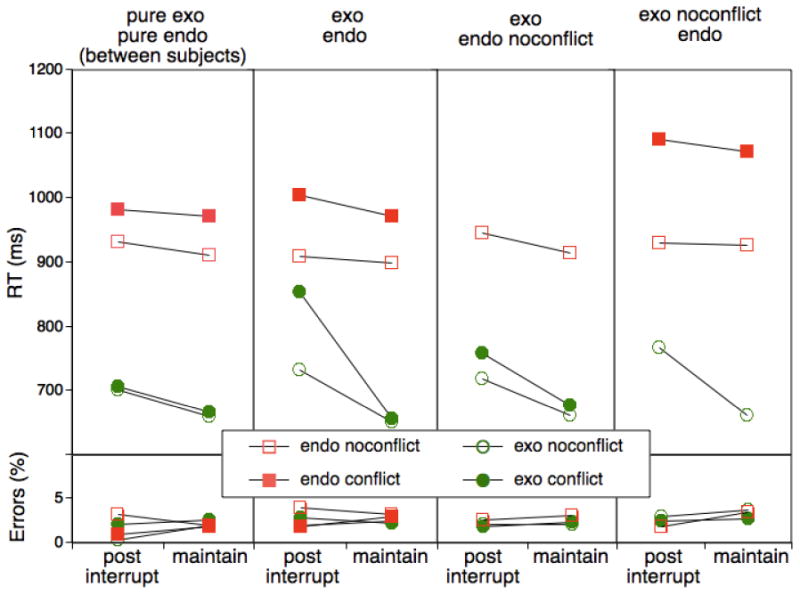
Mean RTs as a function of trial type (post-interruption vs. maintenance) and presence of conflict as a function of block context in Experiment 1. In the “pure-exo/pure-endo” condition, participants either performed only the endogenous or only the exogenous task throughout the experiment and conflict could occur in either task. In the “exo/endo” condition they performed the endogenous and the exogenous condition in alternating blocks and conflict could occur for either task. In the “exo/endo-noconflict-only” condition participants performed both tasks in alternating blocks, but conflict could occur only for the exogenous task. In the “exo-noconflict-only/endo” condition participants performed both tasks in alternating blocks, but conflict could occur only for the endogenous task.
Figure 3 shows RTs and errors as a function of task, conflict level, post-interruption versus maintenance trials, and each of the four conditions. We first examined the primary experimental condition, in which subjects alternated between endogenous and exogenous control, with conflict possible across all trials (exo/endo). Overall, the results show a cost-asymmetry pattern with large post-interruption effects for the exogenous task and relatively small effects for the endogenous task, Task × Interruption: F(1,19)= 32.71, MSE=4561.63, p<.001. This pattern occurred in the absence of an immediate transition between the endogenous and the exogenous task and therefore it cannot be explained in terms of a trial-to-trial carry-over effect between the endogenous and the exogenous task. In addition, this interaction was modulated by the Conflict factor, F(1,19)= 5.83, MSE=3464.79, p<.03, reflecting the fact that the cost asymmetry was 71 ms for no-conflict trials, but 165 ms for conflict trials. Specifically, there was almost no conflict effect for the exogenous task on maintenance trials, M=5 ms, t(19)<.8, compared to a very large effect for the exogenous task on post-interruption trials, M=121 ms, t(19)=4.20, p<.001. For the endogenous task, the corresponding difference was much smaller, M=74 ms, t(19)=7.78, p<.001, vs. M=101 ms, t(19)=4.16, p<.01, and not reliable, F(1,19)= 1.69, MSE=2142.55, p>.2. It may be premature to infer from this that there actually was no increase in the conflict effect as a function of interruption for the endogenous task. The post-interruption trials were less frequent than maintenance trials and this may have made it difficult to detect more subtle differences. However, there can be little question that the combined effect of conflict and interruptions was much larger for the exogenous than for the endogenous task. Overall, this pattern is consistent with the prediction that for the exogenous task the maintenance mode effectively shields against LTM interference, whereas the updating mode creates a situation of strong vulnerability to such interference.
We can also examine ask to what degree the large cost asymmetry persists throughout an entire 80-trial block. It would be consistent with a long-term memory effect if we see some leveling off of the effect as new, context-appropriate memory traces are added within a given block. When adding a block-half factor (i.e., comparing the first 40 trials with the second 40 trials of each block) we find a weak tendency towards a modulation of the Task × Interrupt interaction, F(1,19)=2.81, MSE=7758.64, p=.11. The cost asymmetry during the first half of 165 ms is reduced to 95 ms for the second half, but remains reliable throughout, all Fs>17.2.
In principle it is possible that the large cost asymmetry we observed in the exo/endo condition arises not from the competition between the exogenous and the endogenous task, but instead results from the switch between the math task and either of the two primary tasks. If this were the case then we should see a similar cost asymmetry even when comparing the pure exo and the pure endo control conditions. As Figure 3 indicates, this is clearly not the case. While there may be a small cost asymmetry for the control condition, it is by an order of magnitude smaller than for the exo/endo condition. To validate this observation statistically, we compared the exogenous-task condition from the exo/endo group with the pure exogenous, single-task group and the endogenous-task condition from the exo/endo group with the pure endogenous, single-task group. For the exogenous task, the Group × Interruption × Conflict effect was reliable, F(1,28)=6.54, MSE=3357.13, p<.02. As the figure shows, RTs were essentially identical for the maintenance trials, while for interruption trials RTs were generally increased and there was a substantial conflict effect for the exo/endo condition.
For the endogenous-task trials, the only reliable effect involving the Group contrast, was a Group × Conflict interaction, F(1,28)=7.00, MSE=2254.34, p<.02, indicating that generally, the conflict effect was larger in the exo/endo than in the pure-endo condition. Finally, we also checked whether within the two single-task conditions there was any indication of a cost asymmetry. However the Task (which is here a between-subject variable) × Interruption interaction was far from reliable, F(1,18)=.13, nor was the three-way interaction that also included the Conflict factor, F(1,18)=.32. Thus, clearly the large cost asymmetry observed in the exo/endo condition results from the fact that two competing primary tasks occur within the same context and is not a simply consequence of switching between the math and either of the primary tasks.
The exogenous-task condition proved particularly susceptible to the combined effect of interruptions and experience with the alternate task. An important follow-up question is to what degree it is the experience with the alternate task itself that is responsible for this pattern or whether the interfering LTM traces are particularly potent when encoded during a conflict situation. The latter pattern would be consistent with the idea that conflict boosts encoding of LTM traces. To examine this question, we had included the exo/endo–noconflict condition in which the endogenous task blocks were presented exclusively without exogenous conflict. As Figure 3 suggests, the cost asymmetry, which due to the “incomplete” design with the missing conflict trials for the endogenous task could be evaluated for the no-conflict trials only, was substantially reduced relative to the condition with the all-conflict condition, F(1,38)=6.02, MSE=1077.04, p<.02. More importantly, for exogenous-task trials, the interaction between the Interruption and Conflict factors was reliably larger in the exo/endo than in the exo/endo–noconflict condition, F(1,38)=7.52, MSE=2856.74, p<.01. Combined, this pattern suggests that while the cost-asymmetry is not completely contingent on the presence of conflict during encoding the alternate task, such conflict does boost interference to a substantial degree.
For sake of completeness, we had also included a group that experienced both conflict and no-conflict trials in the endogenous task, but only no-conflict trials in the exogenous task. Given that here participants had experience with the endogenous task in the presence of exogenous conflict, we again expected a clear cost-asymmetry pattern, which however could be evaluated only for the no-conflict trials (again because of the “incomplete” design). In fact, the cost asymmetry for this condition was highly reliable, F(1,19)=42.45, MSE=1445.88, p<.001. As for the endogenous condition, there was a highly reliable conflict effect, F(1,19)=32.46, MSE=15152.01, p<.001, but neither the interruption effect, F(1,19)=.22, nor the interaction with the conflict factor, F(1,19)=.30, were reliable. This pattern was similar to that for the corresponding conditions in the all-conflict condition, with the one exception that the overall conflict effect was larger when conflict was only experienced in the endogenous condition, F(1,19)=5.48, MSE=9311.01, p<.05. This difference was not expected. However, we note that comparisons with the equivalent conditions in Experiments 2 and 3 indicate that this effect may have less to do with a particularly large endogenous-task conflict effect in the exo–noconflict/endo condition than with an unusually small effect in the exo/endo condition of this experiment.
Experiment 2
A key result of the previous experiment was that the cost-asymmetry after interruptions was particularly strong when the non-dominant, endogenous task had to be performed under conditions of conflict. We believe that this result is critical to understanding the cost-asymmetry. After all, a key difference between the dominant and the non-dominant task is that, per definition, processing in the dominant task suffers much less conflict. Thus, the presence of conflict is a necessary condition for the encoding of the very memory traces that are responsible for the post-interruption costs when performing the dominant task. However, this raises the additional question what it is about experiencing conflict from the exogenous task while performing the non-dominant task that is responsible for strong cost asymmetry.
In this context it is important to note that on post-interruption trials the non-dominant task also interferes with performing the dominant task. If it were conflict itself that drives the post-interruption selection costs then it is not obvious why this type of conflict would not counteract the cost asymmetry. Thus, possibly there is something special about the conflict from a dominant task that produces particularly strong memory traces, while conflict from the endogenous task is less effective in this regard.
However, there may be a simpler account. While performing the non-dominant task the maintenance mode is much less effective in shielding processing from the competing-task conflict than when performing the dominant task. Thus, for the non-dominant task, participants experience conflict both on post-interruption and maintenance trials whereas for the dominant task they experience conflict only on post-interruption trials, whereas conflict is effectively blocked on maintenance trials. In other words, the mere number of trials with high, experienced conflict is much smaller for the dominant than for the non-dominant task. Thus, maybe it is simply the greater frequency of experienced conflict from the exogenous to the endogenous task than from the endogenous to the exogenous task that drives the asymmetric cost.
In Experiment 2, we attempted to test this frequency-of-experienced-conflict hypothesis. The critical condition was identical to the experimental condition from Experiment 1 where conflict could occur for both the dominant and the non-dominant task, except for one critical change: Conflict from the exogenous task while performing the endogenous task was limited to post-interruption trials and never occurred on maintenance trials. Ideally, this should mimic the situation for the dominant task, where experienced conflict is also limited to post-interruption trials. Thus, if the frequency-of-experienced-conflict hypothesis is correct, we should see a marked reduction of the cost asymmetry in this condition compared to a situation in which conflict can occur on all trials.
We used in this experiment two control conditions, which also allowed us to replicate the central results from Experiment 1. The first was the exo/endo condition from Experiment 1, for which we again expect the fullblown cost asymmetry. The second was the exo/endo-noconflict condition, for which we again expect to see only a small asymmetry. For the third condition in which non-dominant task conflict was limited to post-interruption trials, we expect performance to be similar to exo/endo-noconflict condition, assuming the frequency-of-experienced-conflict hypothesis is correct. If, however there is something special about conflict suffered from the dominant task that is responsible for the interfering memory traces then the pattern for the new condition with exogenous conflict limited to post-interruption trials, should be more similar to the standard, exo/endo condition.
Methods
Participants
A total of 60 students of the University of Oregon participated in exchange for course credits in this experiment.
Design
Stimuli, tasks, and procedures were identical to those used in Experiment 2, except for the differences mentioned below. Subjects were randomly assigned to three conditions, two of which were exact replications of Experiment 2 conditions: 1) The exo/endo condition with a p=.5 of conflict for both the endogenous and the endogenous task. 2) The exo/endo-noconflict condition with a p=.5 of conflict for the exogenous task, but p=0 conflict for the endogenous task. In the third, the experimental condition there was a p=.5 of conflict for the exogenous task and for the post-interruption trials of the endogenous task, but a p=0 of conflict for the maintenance trials of the endogenous task. Participants were randomly assigned to the three different conditions.
Results
We used the same trial exclusion criteria as in the previous experiments. In no condition of the primary task did error rates exceed 3.6% and in no instance did the pattern of error effects counteract the pattern of RTs. Therefore, we again focus only on RTs here, but present error results in Figure 4. For the interruption task, the mean error rate was 14.45% (SD=9.69) and the mean RT was 4787 ms (SD=1761).
Figure 4.
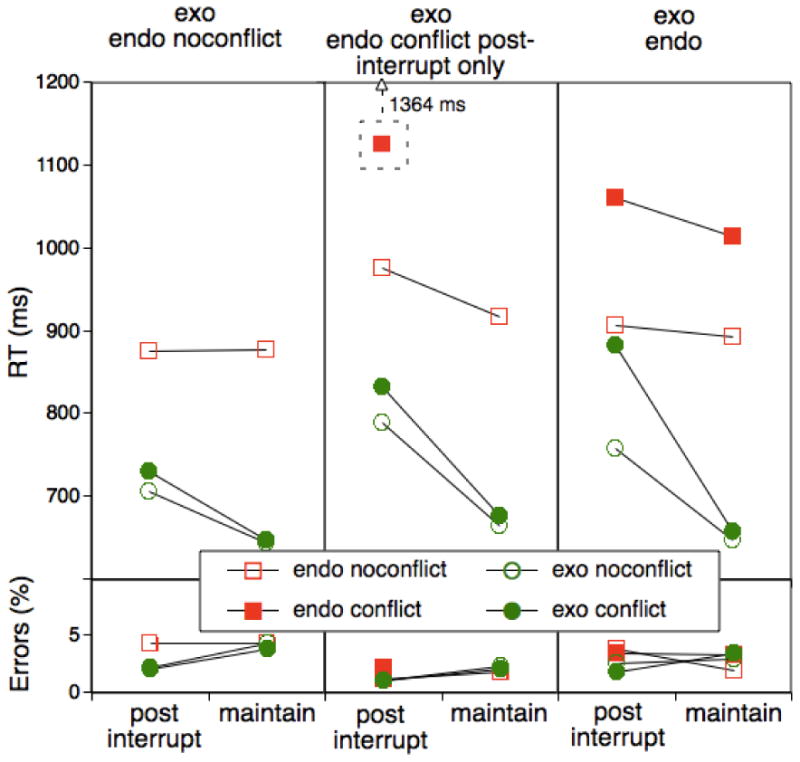
Mean RTs as a function of trial type (post-interruption vs. maintenance) and presence of conflict as a function of block context in Experiment 2. The “exo/endo-noconflict-only” and the “exo/endo” conditions were identical to the corresponding conditions in Experiment 1. The “exo/endo-conflict-postinterrupt-only” condition was identical to the “exo/endo” condition, however for endogenous trials conflict could occur only on post-interruption trials.
Figure 4 shows the pattern of RT and error results for each of the three conditions as a function of task, interruption (post-interruption vs. maintenance), and conflict. First, note that the pattern for the all-conflict and the exogenous-conflict-only conditions was very similar to the two corresponding conditions in Experiment 1. Thus, we replicated the basic pattern of an interruption-based cost asymmetry that is dependent on experience with conflict in the endogenous task. This conclusion is confirmed in the statistical analyses. When comparing the exo/endo and the exo/endo-noconflict group, we found a highly significant Group × Task × Interruption interaction, F(1,38)=8.06, p<.01, MSE=11288.99, and a significant Group × Task × Interruption × Conflict interaction, F(1,38)=9.68, p<.01, MSE=2136.51.
Regarding the new condition with endogenous-task conflict only for post-interruption trials, we first need to note that RTs in the endogenous, post-interruption, conflict trials were almost 300 ms larger than in the corresponding trials from the exo/endo condition (see also Experiment 2). Likely, this is due to the fact that in this condition, conflict is a rare event that occurs only on post-conflict trials and that therefore is particularly disruptive (e.g., Tzelgov, Henik, & Berger, 1992). We will return to potential implications of this effect below.
The most important result for this condition is that the pattern of RTs of task-specific interruption effects was more similar to the exo/endo-noconflict condition than to the exo/endo condition. Note, that this is somewhat obscured by the fact that there were larger task-unspecific post-interruption costs in this group. To confirm this impression, we first compared the corresponding no-conflict conditions for both tasks between the exogenous-conflict-only and the experimental group. In general, there was a significantly larger post-interruption main effect for the experimental group than for the exogenous-conflict-only group, F(1,38)=6.31, p<.02, MSE=6064.78, however there was no trace of the critical Task × Interruption interaction, F(1,38)<.2. We can also compare the exogenous conditions across these two groups. Again, we found a Group × Interruption interaction here, F(1,38)=6.14, p<.02, MSE=7340.83, but the Group × Interruption × Conflict interaction was not reliable, F(1,38)<.1.
Finally, we can also compare the experimental group with the all-conflict group. Here, we do see a reliable Group × Interruption × Conflict interaction, F(1,38)=4.72, p<.05, MSE=3540.66, suggesting that in the exo/endo group there was greater conflict on exogenous, post-interruption trials than in the experimental group.
As in the previous experiment, we again checked to what degree the cost asymmetry in the exo/endo condition was persistent within the 80-trial blocks. As in the previous experiment, there was a tendency towards a reduced asymmetry in the second half of the block, F(1,19)=3.19, p>.07, MSE=7340.83 (1st half=182 ms, 2nd half=110 ms), however the critical interaction was reliable for both halves, F(1,19)>23.23.
In general, these results suggest that frequency of experienced conflict is at least one critical factor behind the cost asymmetry observed in the all-conflict conditions in Experiment 1 and the current experiment. However, we need to ask at this point to what degree these conclusions need to be qualified by the unusually long RTs in endogenous, post-interruption, high-conflict trials (see Figure 4). Arguably, if amount of conflict were critical then the strong conflict that was experienced on these trials should have also led to particularly strong interference on exogenous-task, post-interruption trials. We did find that participants had larger post-interruption costs in a task-unspecific manner—which possibly is due to the experience of very high conflict on some post-interruption trials. However, there was no specific effect on conflict trials that would qualify our main conclusions. If anything the large RTs in the endogenous, post-interruption, high-conflict trials ensured that our experimental condition produced a rather conservative test of the idea that frequency of conflict instances is more critical than the experience of conflict per se.
Experiment 3
Experiments 1 and 2 clearly confirmed our predictions: Recovery from interruptions produced a strong cost asymmetry in the absence of actual switches between competing tasks and this effect was particularly pronounced when the competing task was experienced frequently in conditions of high conflict. The main purpose of this experiment was to further examine the role of interruptions in eliciting the cost asymmetry. From the perspective of our model, the effect of interruptions is structural in nature. It enforces an updating operation, which in turn creates an unconditional opening for any memory traces associated with the current context, wanted or unwanted, to influence processing. However, there is an alternative possibility. Across alternating blocks, the specific interruption task (e.g., solving math equations) may become linked with either of the two possible tasks that can potentially follow the interruption task via associative learning. Thus, after concluding a math trial, there may be two learned associations in place, one to the endogenous task and the other to the exogenous task and a time-consuming, controlled retrieval process may be necessary to determine the currently relevant task. To examine this possibility we used two different interruption tasks in Experiment 3. The first was the math task, identical to the one used in Experiments 1 and 2. The second task involved solving simple anagrams (i.e., the “word task”). In the critical condition there was a consistent mapping between interruption task and block type (i.e., 2:2 mapping), such that for half of the subjects the math task would be only coupled with the exogenous task and the word task only with the endogenous task (and the other way round for the other half of the subjects). We compared this condition to one in which each participant was exposed to only one interruption task for both endogenous and exogenous blocks (i.e., 1:2 mapping). If learned associations matter then the cost-asymmetry pattern should be present only in the group with the inconsistent 1:2 mapping. However, if we obtain the cost asymmetry even when type of interruption is consistently mapped to block type then this would suggest that interference is due to the structural effect of interruptions rather than to specific associations.
In this experiment, we also wanted to rule out another possible alternative explanation for the interruption-triggered cost asymmetry. In Experiments 1 and 2, the interruption-task stimuli were presented centrally, which is the same area on the screen where also the cue for the endogenous task was shown. This overlap in location may have biased participants towards the center of the screen while recovering from the interruption, thus giving priority to the endogenous task cues. The fact that the cost asymmetry was absent for the single-task conditions or much reduced when the endogenous task was experienced without conflict (in both of which the interruption task was also presented centrally) indicates that the positioning of the interruption task could not be the sole explanation. However, it is possible that this served as a mitigating factor. Therefore, in Experiment 3 we presented the interruption task at random locations on the screen, avoiding positions closer than 6 degrees to the center.
Methods
Participants
A total of 40 students of the University of Oregon participated in exchange for course credits in this experiment.
Tasks and Design
The same tasks and stimuli were used as in the preceding experiments. However, we added as a second interruption an anagram task. Participants were shown four or five-letter anagrams along with two letters just below to the left and the right of the anagram (Times font, size=24). Subjects had to press either the left or right key to indicate which of these was the first letter of a legal word that could be formed with the anagram letters. Anagrams were selected from a pool of 140 possible words. Words could be used more than once per experiment, but only after all other words had been used. Another difference from the preceding experiments was that interruption task stimuli both for the math and the anagram task were presented in random positions within an area that was at least 6 degrees, but no more than 9 degrees away from the center of the screen.
Half of the subjects were randomly assigned to the 1:2 mapping group, which worked either only with the math task or only with the anagram task (i.e., 10 subjects each). The other half of the subjects was assigned to the 2:2 mapping group, for which both the math and the anagram task were presented randomly. Otherwise, the endogenous and exogenous tasks were identical to the exo/endo conditions in Experiments 1 and 2 with 50% conflict in either the endogenous or the exogenous task.
Results and Discussion
We used the same trial exclusion criteria as in the previous experiments. Again, in no condition of the primary task did error rates exceed 3.0% and in no instance did the pattern of error effects counteract the pattern of RTs. Therefore, we focus our reporting of analyses on RTs only, but we do present errors in Figure 5 along with RTs.
Figure 5.
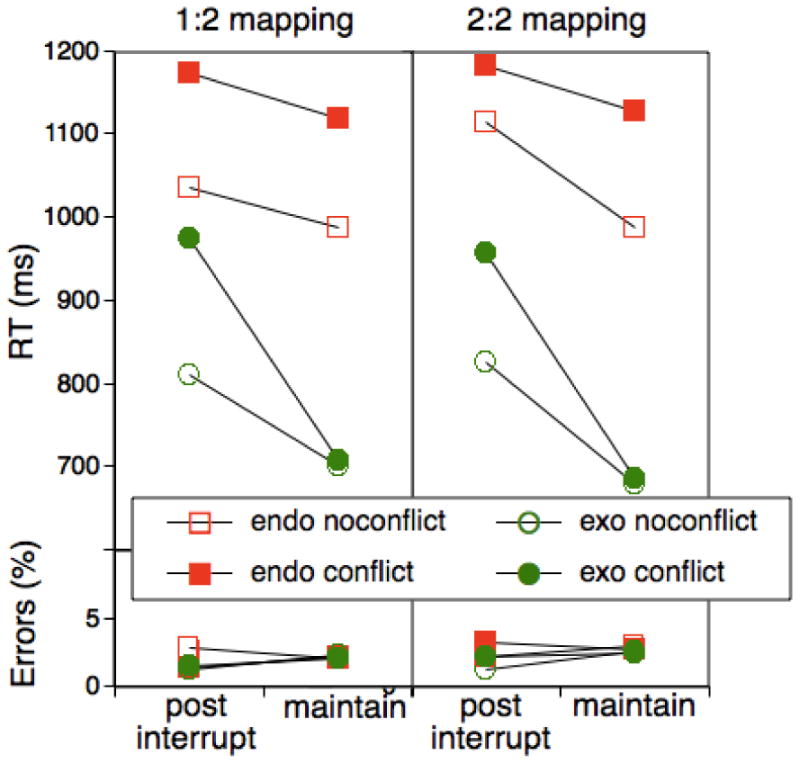
Mean RTs as a function of trial type (post-interruption vs. maintenance) and presence of conflict as a function of mapping between interruption tasks and primary tasks (1:2 mapping versus 2:2 mapping) for Experiment 3.
The mean error rate for the math task was 11.1% (SD=6.7%) in the group that only performed the math task and 15.5% (SD=12.4%) in the group that performed both interruption tasks. The corresponding values for the anagram task were 9.8% (SD=11.4) and 6.1% (SD=3.4). Mean RTs for the math task were 4486 ms (SD=1496) for the math-only group and 4979 ms (SD=2118) for the mixed group. The corresponding RTs for the anagram task were 2759 ms (SD=789) and 2478 ms (SD=1028).
The upper panel of Figure 5 presents RT results for the primary tasks as a function of task, interruption, and conflict, separately for the condition in which interruption task and primary tasks were either inconsistently (1:2) or consistently (2:2) mapped. As apparent, across both conditions the qualitative data pattern was largely similar to the one obtained for the corresponding exo/endo conditions in Experiments 1 and 2. The switch-cost asymmetry, that is the Task × Interruption interaction was highly significant, F(1,38)=55.88, MSE=5846.37, p<.001, and this effect was not modulated by the Mapping factor, F(1,38)=.25. As in the preceding experiments, the cost-asymmetry was further modulated by conflict, F(1,38)=26.07, MSE=5745.93, p<.001. In particular, there was generally no conflict effect for exogenous-task maintenance trials, while there was a substantial effect for interruption trials. Again, this interaction was not significantly modulated by the task mapping, F(1,38)=.51. As in the previous experiments, we checked whether the asymmetry pattern persisted across the entire block and found a numerical reduction of the asymmetry effect from 145 to 108 ms, that however was not significant, F(1,38)=1.81, MSE=7780.39, p<.15. 4
To conclude, with the present results we cannot rule out the presence of associative learning effects between consecutive tasks (e.g., Koch, 2001). Small effects of this kind may have been difficult to detect with our design. However, the results provide little reason to suspect that associative links between tasks play a major role in producing the interruption-specific, cost-asymmetry pattern. Therefore, they strengthen our structural hypothesis, namely that interruptions enforce an updating operation in the course of which interference through LTM traces can enter the selection process.
Different from the preceding experiments, we presented the interruption-task stimuli far from the screen's center to avoid any kind of bias favoring the central cue after an interruption. The qualitative pattern of effects was very similar to the one obtained in precious experiments. Therefore it is unlikely that the positioning of the interruption task on the screen played a major role in the pattern of costs.
Experiment 4
One limitation of our results thus far is that we used interruption tasks which themselves were fairly complex and required considerable attentional control. Maybe the pattern of post-interruption costs we had observed can be explained in terms of an aftereffect of immediately preceding, high control demands. Therefore, in this experiment, we used a variant of a spatial Stroop task requiring manual key responses as interruption events that allowed us to directly manipulate control demands. Specifically, one group of subjects performed the interruption task with low-demand instructions, where correct key responses were indicated through the dominant dimension (i.e., arrow directions). The second group of subjects performed the task with high-demand instructions. Here, correct key presses were indicated through arbitrary color-key assignment rules and the arrow direction produced potentially conflicting information. Also, different from the preceding experiments, the interruption events were not just single trials, but followed the same probabilistic “switch” rules as the primary tasks. Specifically, no matter what the current task type, there was a p=.2 probability that the next trial switched to the other task possible in that block. This also allowed us to examine to what degree the pattern of post-interruption costs depended in any critical manner on the number of intervening interruption events/trials.
Methods
Participants
A total of 40 students of the University of Oregon participated in exchange for course credits in this experiment.
Tasks and Design
The same primary tasks and stimuli were used as in the preceding experiments. However, a new interruption task was used that allowed a manipulation of response-selection demands. For these interruption trials, initially an empty stimulus box (6 degrees side length) appeared on the screen. After 1000 ms, an arrow (4.8 degrees length) appeared that pointed to one of the four corners of the box and was colored red, blue, green, or yellow. Depending on condition, subjects responded with their right-hand index finger by pressing keys on the numerical keypad that corresponded to the corners of the square (2, 3, 5, or 6). Subjects in the low-control condition were instructed to press the key that was compatible with the arrow direction. The color dimension was not explicitly mentioned to these subjects. For subjects in the high-control condition the correct key was indicated through arbitrary color-key assignments (red=upper left, green=upper right, yellow= lower left, blue=lower right). Arrow direction and the key indicated by the color were in conflict on 50% of interruption trials. Transitions between the primary task and the interruption task occurred with probability p=.2.
As in the critical experimental conditions of the preceding experiments, subjects alternated between pure endogenous and pure exogenous control blocks. The only difference was that we extended the length of blocks to 100 trials per block. Half of the subjects worked exclusively with the low-control interruption task, the other half with the high-control interruption task.
Results and Discussion
We used the same trial exclusion criteria as in the previous experiments. In this experiment relevant error results were obtained and will be reported alongside with RT results.
The mean RTs for the low-demand interruption task was 501 ms (SD=63). Mean RTs for the high-demand interruption task were 714 ms (SD=114) for compatible and 816 ms (SD=169) for incompatible trials. Corresponding error percentages were 0.7% (SD=.64), 1.4% (SD=2.2) and 5.9% (SD=3.0%). Thus, with these interruption tasks, we implemented a strong variation in control demands.
Figure 6 presents RT and error results for the primary tasks as a function of task, interruption, and conflict, separately for the low-demand and the high-demand interruption conditions. As apparent, across all conditions the qualitative RT data pattern was largely similar to the one obtained for the corresponding conditions from the previous experiments. For the analysis, we added as additional factor whether or not the last interruption episode was short (i.e., <=2 trial) versus long (>2 trials). With this categorization of interruption episodes, there was an about equal number of observations in each category. The switch-cost asymmetry, that is the Task × Interruption interaction was highly significant, F(1,38)=29.33, MSE=19629.69, p<.001, and this effect was not modulated by the type of interruption, F(1,38)=.07. Also, the cost-asymmetry was modulated by conflict, F(1,38)=5.63, MSE=13918.91, p<.03. In particular, there was generally no conflict effect for exogenous-task maintenance trials, while there was a substantial effect for interruption trials. Again, this interaction was not significantly modulated by interruption-task demands, F(1,38)=.04. Also the Task × Interruption interaction was not significantly modulated by whether the previous interruption episode was short or long, F(1,38)=.18, and there were no other significant effects associated with the length of the interruption.
Figure 6.
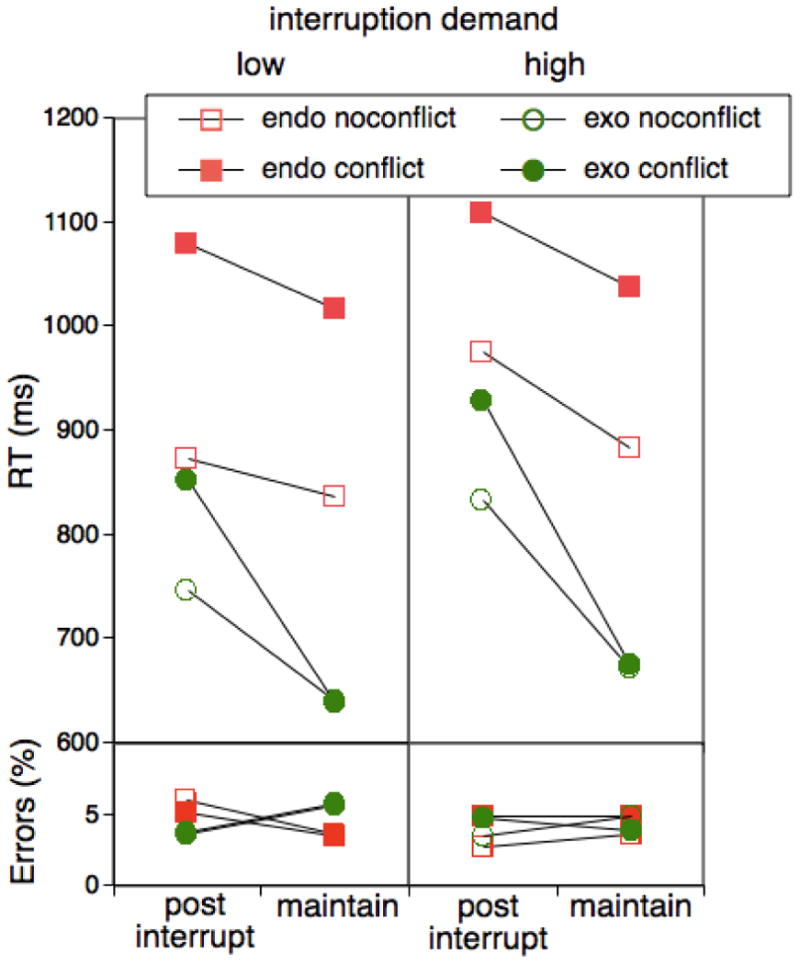
Mean RTs and errors as a function of task (exogenous vs. endogenous), trial type (post-interruption vs. maintenance), presence of conflict, and control demands of the interruption task for Experiment 4.
As in the previous experiment, there was a tendency for the cost asymmetry to decline between the first half (142 ms) and the second half of the block (98 ms), F(1,38)=3.52, MSE=3037.54, p>.06. However, the cost asymmetry was highly reliable for both block halves, Fs(1,38)>24.15.
For the high-demand condition, the pattern of errors was consistent with previous experiments in that it was not reliably affected by the experimental factors; in particular there was no trace of a cost asymmetry, F(1,19)=.01. However, in the low-demand condition, the cost-asymmetry pattern was opposite to that obtained on the level of RTs and the relevant Task × Interruption interaction as reliably modulated by the condition factor, F(1,38)=6.42, MSE=13.10, p<.02. In principle, this pattern could point to a speed-accuracy tradeoff. However, the size of the “reverse” error cost-asymmetry effect showed a zero correlation with the RT cost-asymmetry effect in either of the two between-subject conditions (low demand: r= –.01 ; high demand: r=–.01). Also, when repeating the RT analyses for the low-demand group after eliminating those subjects with an above-median reverse cost-asymmetry effect, there was still a highly reliable cost asymmetry, F(1,38)=3.52, MSE=2557.40, p<.01. Thus, while the unique pattern of error effects was certainly not predicted for the low-demand condition, there is no reason to assume that it qualifies the pattern of RT results.5 Therefore, the main result of this experiment was that neither the level of control demands during the interruption nor the length of the interruption influenced the pattern of post-interruption costs in a theoretically significant manner.
Experiment 5
So far, as our primary task pair we had juxtaposed endogenous versus exogenous control over spatial attention. In this final experiment we wanted to examine to what degree the basic pattern of results generalizes to a paradigm where conflict is generated during response selection, rather than during attentional-input selection. To this end, we replaced the endogenous versus exogenous spatial attention tasks with a spatial stroop task. Participants in the experimental group switched back and forth between blocks that either required a response to a word (UP, DOWN, LEFT, or RIGHT) presented in one of four locations, or execute a spatially compatible response to the location of the word. We also included two different control groups of subjects who either only performed the word or the location task throughout the experiment. As interruption events, we returned to the math tasks from Experiments 1-3, but presented at random locations (as in Experiment 3).
Method
Participants
A total of 60 students of the University of Oregon participated in exchange for course credits in this experiment.
Tasks and Design
On each trial, four squares (4 degrees side length) were presented at one of four locations, in a crosswise arrangement (8 degrees from the center of the screen). Additionally, a single word (UP, DOWN, LEFT, or RIGHT, presented in 24 point Helvetica font) appeared in the center of one of the squares. Subjects responded with their right-hand index finger by pressing either the 2 (bottom), 4 (left), 6 (right), or 8 (top) key on the numerical keyboard. In the word task, subjects were instructed to press the key that corresponded to the word that was displayed. In the location task, subjects' key responses were compatible with the spatial location of the word. Each block was 100 trials long. The response-stimulus interval was 1000 ms. The math interruption task was presented exactly as in Experiments 3.
Subjects were randomly selected into the experimental condition or in one of two single-task controls conditions In the experimental condition (N=20), subjects alternated between pure word and pure location blocks. In each of the two control conditions (N=20), subjects either performed only the word or only the location task throughout the experiment. As in the previous experiments, participants performed on initial practice block for each task that was identical to the following test blocks.
Results and Discussion
We used the same trial exclusion criteria as in the previous experiments. Initial analyses revealed that in this experiment, different results were obtained for the first versus the second half of each block. Therefore, Figure 7 shows the RT and error pattern separated by block half. Given that in this experiment, error patterns could reflect theoretically relevant response-conflict effects, we analyzed them here as well.
Figure 7.
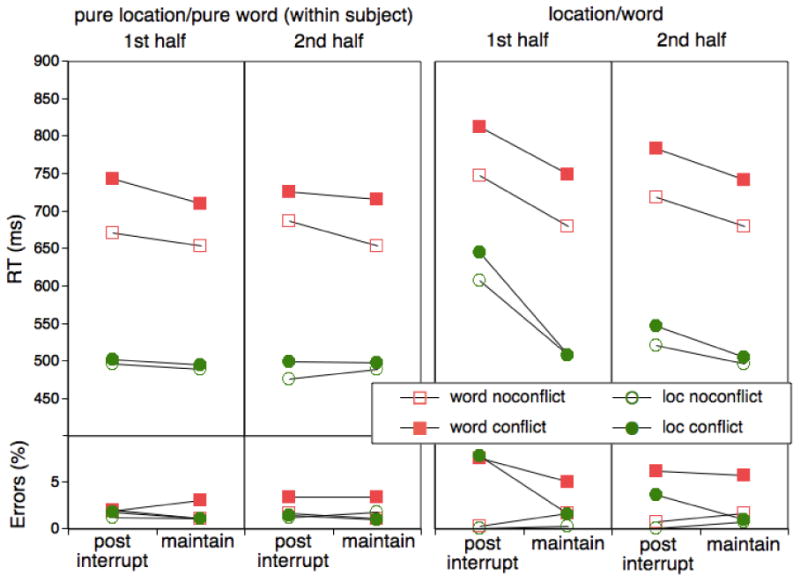
Mean RTs and errors as a function of task (spatial Stroop responses to locations vs. words), trial type (post-interruption vs. maintenance), presence of conflict, block half, and control vs. experimental conditions (pure-location/pure-word vs. alternating location/word) for Experiment 5.
Turning first to the experimental group, in the first block half there was a clear RT cost-asymmetry pattern, whereas in the second block half, overall interruption costs and the cost asymmetry pattern were much smaller. In the experimental group, the two-way interaction indicative of the cost asymmetry pattern did not quite reach the significance criterion, F(1,19)=3.19, MSE=3183.20, p>.09; the same is true for the additional modulation of this effect through the response-congruency factor, F(1,19)=3.28, MSE=1224.43, p<.09. However, the cost-asymmetry interaction was significantly modulated through the block-half factor, F(1,19)=5.36, MSE=3466.19, p<.04. In the first half, the cost asymmetry was reliable, F(1,19)=6.84, MSE=2386.41, p<.02, and the additional modulation of this effect by the response-conflict factor was almost reliable, F(1,19)=3.83, MSE=1038.90, p<.07. For the second half, both of these two effects were far from reliable, F<.5. In terms of errors, the theoretically critical Task × Interruption interaction approached significance, F(1,19)=3.97, MSE=10.26, p<.07, while the additional interaction with the Congruency factor was in the expected direction, but not significant, F(1,19)=1.85, MSE=14.45, p=.19.
By looking at the control groups, we can again assess to what degree the cost asymmetry present in the experimental group is an unspecific phenomenon rather than tied to experience with both types of tasks. As obvious from Figure 6, the cost asymmetry that is present in the experimental group (at least in the first half) is completely absent when comparing the two control groups. In fact, in an ANOVA comparing the two word-task group and the location-task group, there was a nearly reliable Group × Interruption interaction that in terms of direction was opposite to the cost asymmetry interaction in the experimental condition, F(1,38)=3.89, MSE=2357.60, p<.06.
Also, when comparing the dominant, location-task performance for the control and the experimental groups there was a highly reliable Group × Task × Interruption interaction, F(1,38)=20.54, MSE=5439.39, p<.01, that was further modulated by the block-half factor, F(1,38)=10.56, MSE=2506.01, p<.01, and in addition, by the response-congruency factor, F(1,38)=4.15, MSE=880.64, p<.05, with somewhat larger congruency effects for first-half, post-interruption trials. Furthermore, for the location task, there was no reliable difference between groups for maintenance trials (F1,38)<.6, suggesting that once recovery from interruptions was complete, subjects in the experimental condition were able to focus on the location task just as well as those in the control condition. For errors, it is evident that there is no hint of a post-interruption cost-asymmetry in the control condition.
All in all, the pattern we obtained with this task combination was similar to what we found for the endogenous/exogenous attentional control tasks (see also our previous results with the Stroop task (Bryck & Mayr, 2008). However, there were two qualifications. First, the effect was less persistent than in the previous experiments with a clear cost asymmetry in the first half that largely diminished in he second half. Accumulation of new memory traces during the first half of each block in the experimental condition may have counteracted the interference from the previous block with the competing task. We had found a tendency of a diminishing cost asymmetry also in the preceding experiments. Thus, at this point it would be premature to conclude that there is a qualitative difference in the persistence of interference between attentional-selection and the response-selection domain.
Second, in the preceding experiments with the endogenous/exogenous tasks the presence of conflicting information (in particular the presence/absence of endogenous task cues) typically had a reliable effect on the cost asymmetry. In the spatial Stroop task, the presence of response conflict had less of an additional effect on the dominant task over and above the effect of interruptions. One possible reason for this divergence is that in order to avoid contingencies between the irrelevant and the relevant dimensions (e.g., Melara & Algom, 2003) locations and word stimuli were selected randomly, leading to an average conflict probability of p=.75. In situations with high probability of conflict, conflict effects are often reduced, possibly because of a general tightening of control (Tzelgov et al., 1992). Another reason for a relatively small contribution of conflict trials to the cost asymmetry is that in this experiment, conflict effects were “diluted” across RTs and errors, whereas in the endogenous/exogenous task conflict effects affected RTs only. The error effects we did observe in Experiment 5 were in the same direction as RT effects, albeit only approaching the statistical significance criterion.
General Discussion
Our goal was to explore the conditions that make it difficult to select between competing control settings, specifically between endogenous and exogenously controlled attention. On a theoretical level we started out by proposing two conditions that have to be met so that subjects experience substantial selection costs. First, LTM needs to contain memory traces from earlier selection instances with the competing task. Second, these memory traces produce interference only once working memory is forced from a maintenance to an updating state, such as through strong bottom-up interference (as in the endogenous task), task switches, or during recovery from externally imposed interruptions (as in the exogenous task during post-interruption trials). From these assumptions we derived and empirically confirmed the prediction that the asymmetric selection costs (i.e., larger costs for dominant than for non-dominant control settings) arise after any interruption of ongoing processing. This is in contrast to predictions from the currently dominant “carry-over” account of task switching (e.g., Gilbert & Shallice, 2002), which cannot explain a selection cost asymmetry that arises from mere interruptions. Rather, for this account the trial-to-trial clash between competing task or control settings is a necessary condition for the selection cost asymmetry to arise.
Cost asymmetries in the absence of task switches between competing tasks have been reported occasionally in the past (Allport & Wylie, 2000; Bryck & Mayr, 2008). The current results go significantly beyond the existing evidence and allow us to both strengthen and fill in important details about our rather broad starting propositions:
First, the interruption-based cost asymmetry is fairly general. In particular, it occurs both in situations in which subjects need to select between competing attentional control settings (Exp. 1 to 4) and between competing stimulus-response mappings (Exp. 5; see also Bryck & Mayr, 2008)).
Second, in line with the LTM memory account, we document the role of experienced conflict on encoding of LTM traces. The cost asymmetry arises only when subjects have experienced conflict from the dominant task while performing the non-dominant task (Exp. 1).
Third, we also examined the role of interruptions in determining the cost asymmetry. Consistent with the idea that interruptions have a structural effect (i.e., in terms of enforcing an updating process during recovery from the interruption), we demonstrated that the cost asymmetry could not be explained in terms of task-specific associative mappings (Experiment 3). Also, consistent with the structural hypothesis we demonstrated that the type, the attentional control demands, or the duration of interruptions have at best very small effects on the cost asymmetry. In line with our general model this suggests that what matters is not the interruption activity itself, but simply the fact that interruptions elicit a working-memory updating process.
Fourth, an important aspect of the current work is that the interruption paradigm, combined with the exogenous/endogenous tasks provides a particularly clear empirical distinction between the updating mode (post-interruption trials) and the maintenance mode (exogenous-task maintenance trials). In standard task-switching situations it seems particularly difficult to bring subjects to adopt a pure maintenance mode, as the mere possibility of task switching may induce a tendency to update even on no-switch trials (e.g., Monsell & Mizon, 2006). A clear empirical distinction between updating and maintenance allows examining context and individual differences factors that could selectively affect these two modes of control and how people negotiate between them. We are currently using this paradigm to test the hypothesis that older adults are “chronic updaters” (Mayr, 2001; Spieler, Mayr, & LaGrone, 2006). Our initial results indicate that while young adults exhibit virtually no trace of conflict from the endogenous task in exogenous-task maintenance trials (e.g., as in the exo/endo condition in Figure 2), old adults continue to exhibit substantial costs and conflict effects across maintenance trials. This is consistent with the hypothesis that old adults find it difficult to revert to a full maintenance mode following an interruption.
Combined, our results provide a powerful challenge to models that emphasize the clash between the immediate past and the present as the main driver of task-selection costs. In the following we discuss both implications of our results, as well as remaining challenges.
Controlling Attentional Control
We found that adopting an exogenous attentional setting after an interruption produces a larger RT cost than adopting an endogenous attentional setting. As noted, this pattern is similar to the switch-cost asymmetry found for other dominant versus non-dominant task combinations in traditional task-switching situations. It is a defining criterion of exogenous control that it suffers no or only very little interference from signals associated with endogenous control. Thus, the present results suggest an important exception from this rule. Exogenous control appears fluent and interferenceresistant only once it is established and when it merely needs to be maintained across trials. In fact, what we know about the difference between exogenous and endogenous selection comes from studies using such “maintenance” conditions (i.e., pure blocks and no interruptions; e.g., Posner, 1980; Müller & Rabbitt, 1989). The current results show that the process of intentionally selecting an exogenous mode of control seems at least as vulnerable as the process of selecting endogenous settings, at least when LTM contains traces about competing, endogenous control settings.
A remaining open question is how exactly endogenous-task interference disrupts processing on post-interruption, exogenous-task trials. Responses to sudden-onset stimuli have been proposed to reflect an unconditional, reflex-like response (e.g., Theeuwes, 2004). Therefore it would be a particularly noteworthy (and for this notion damaging) result if exogenous-task selection costs arise because the potency of the exogenous stimulus to attract attention is reduced on post-interruption trials. Alternatively, it is also possible that the initial, exogenous pull of the exogenous stimulus remains intact and that it is only after visiting the exogenous stimulus that attention is (erroneously) brought back to inspect the central cue. We are currently investigating this important question by applying eye-tracking analyses to the exogenous/endogenous control paradigm.
Carry-Over versus LTM Retrieval
The LTM encoding/retrieval model of task selection that is supported by the current data has the potential of explaining traditional task-switching effects without invoking the need for passive, trial-to-trial carry-over of information. Such passive carryover is a hallmark of connectionist explanations for task-switch effects (Brown, Reynolds, & Braver, 2005; Gilbert & Shallice, 2002; Yeung & Monsell, 2003a,b). Obviously, such models cannot explain selection costs that arise in the absence of any switch from the competing task. Also, these results cannot be explained by the kind of hybrid carry-over/LTM retrieval model proposed by Wazsak et al. (2003, Wazsak, Hommel, & Allport, 2005). According to this account, interference does result—just as we assume—from LTM traces of earlier selection instances. However, it is the trial-to-trial carry-over of the no-longer relevant task representation (i.e., “task-set inertia”) that generates the vulnerability towards these LTM traces on switch trials. Instead, our results suggest that the a switch between competing tasks is only one instance of a broader category of events that lead to a working-memory updating state, which in turn allows interference from LTM traces to enter the system. Carry-over form the preceding trial is not a necessary part of this process.
Any recovery from interruptions involves a switch in task (albeit not necessarily from a competing task) and on the basis of the current results alone we cannot rue out that this switch is somehow responsible for the cost asymmetry. However, in Bryck and Mayr (2008) we did find a cost asymmetry even after 5 seconds unfilled delays between trials and in the absence of any switches in tasks. As we would argue, such delays increase the probability of loosing the current task set, which then triggers working memory updating.
The LTM-interference/updating account can explain both the clearly LTM-induced cost asymmetry studied here and the traditional switch cost asymmetry (see also, Mayr, 2008; Mayr, 2009; Wazsak et al., 2003). Therefore, according to Occam's razor any inferences about an additional process (such as carry-over) should be based on strong evidence that the trial-to-trial switching situation is in some way unique and cannot be handled by the LTM-interference/updating account alone.
One result that at first sight seems to support the carry-over model comes from Yeung, Nystrom, Aronson, and Cohen (2006). Participants switched between tasks that required subjects to attend either to word or to face stimuli. After task switches there was greater activity in neural areas associated with the currently irrelevant task than after non-switch trials. Also, the degree of this irrelevant-task activity predicted RT switch costs. These results are consistent with the carry-over account, but not necessarily inconsistent with the LTM interference account. With only two tasks it is impossible to tell to what degree there would have been also heightened activity on no-switch trials—only to a lesser degree than on switch trials. It is not at all incompatible with an LTM model when interference is greater for a more recently used alternate task (e.g., on switch trials) than for a less recently used task (e.g., on no-switch trials; for related findings see Bryck & Mayr, 2008). In fact, Wylie, Javitt, and Foxe (2004) looked for activation in neural areas associated with task dimensions that had been performed in a previous block of trials, but were not relevant on the current single-task block. Consistent with the LTM model, they found increased task-irrelevant activity (compared to a control situation in which the irrelevant task had not yet been experienced), even though no immediate switching between tasks was involved.
We are aware of one set of studies that proposed a specific version of a carry-over model, and also made an explicit effort of ruling out an LTM account of the cost asymmetry. Yeung and Monsell (2003a) had presented a connectionist model that explains different patterns of cost asymmetry as resulting from the combination of carryover of task activation and the relative amounts of control activation required when tasks either do or do not directly compete with each other. Yeung and Monsell (2003b) extended this work to test the carry-over (a.k.a. task-set inertia) against the LTM (a.k.a., associative priming) account. Participants had to switch between two initially unfamiliar tasks (i.e., alphabet arithmetic and judging whether a letter and a number both contained curves or not). However, each switching block was preceded by a single-task practice block that was supposed to selectively strengthen one of the two tasks. Across the experiment, practice blocks alternated between the two tasks. The authors proposed that the associative priming account predicts that it should be particularly hard to switch to the most recently non-practiced task because that would require countering the interference from the most recently practiced task. In contrast, the carry-over account predicts larger costs when switching to the recently practiced task because more control was necessary for the recently unpracticed task on the pre-switch trial, which in turn should make it harder to switch away from that task (due to carry-over). The results were largely consistent with the latter prediction. However, there were also aspects of these results that are inconsistent with the interpretation that the observed cost asymmetry was due to inertia of either high-control or a low-control task settings across trials. Specifically, there was little evidence that the relatively short practice blocks (i.e., 32 trials) actually affected relative task dominance. In fact, no-switch RTs were largely similar across recently practiced and unpracticed tasks. Therefore it is not clear to what degree this actually constituted a traditional switch-cost asymmetry, which is defined in terms of larger switch costs to a dominant/easy than to a non-dominant/hard task.
An alternative interpretation of the pattern reported by Yeung and Monsell (2003b) is that the larger switch costs to the practiced task reflect the effect of “inappropriate transfer” between the single-task blocks and the task-switching blocks. It may be harder to switch to the most recently practiced task (i.e., task A) exactly because switch operations were not necessarily associated with this task during the interspersed task-A practice block. In contrast, task B had last been used in a switching context (i.e., the switching block that preceded the last single-task block). Thus, at this point we do not know to what degree the pattern reported in Yeung and Monsell (2003b) truly reflects a switch-cost asymmetry associated with relative differences in dominance between tasks. Whether or not the LTM account will turn out to be fully sufficient to explain task-switch costs, our results do show an important category of asymmetric costs for which the carry-over account clearly cannot provide a sufficient explanation.
As mentioned earlier, our finding of large selection costs in the absence of task switches are not without precedence. For example, several researchers have shown that a simple restart of a continuous run of tasks produces substantial costs (e.g., Gopher, Armony, & Greenshpan, 2000, Altman, 2007; Allport & Wylie, 2000; Lien, & Ruthruff, 2004). Based on our account, these costs arise because the need for a restart enforces an updating process, including costly re-retrieval of the current task set. There are probably many factors that can elicit such updating operations, such as forced breaks (Gopher et al., 2000), high probability of task switches (e.g., Mayr, 2007), or errors and the experience of conflict (e.g., Botvinick et al., 2001).
We do not want to preclude the possibility that trial-to-trial transitions between task/control settings have unique characteristics that are not present for LTM retrieval effects. In fact, when also considering stimulus-response repetition effects across task repetitions versus changes, usually a characteristic cost-benefit pattern arises. Specifically, costs are largest when there is partial overlap (e.g., cost for task changes with stimulus or response repetitions is larger than when everything changes). Hommel (2004) has suggested that this partial-mismatch pattern reflects aftereffects of integrated “event files” that bring all relevant codes for a specific selection instance together into an executable state and that have to be “unpacked” if specific codes need to be reused on the next trial. Using a rule-switching paradigm, Mayr and Bryck (2005) looked for such a partial-mismatch pattern both for trial-to-trial transitions and for the effect of long-term memory traces on current processing. Interestingly, while the first yielded the nonmonotonic, partial-mismatch pattern, LTM effects were characterized by monotonic, similarity-based effects (the greater the match between past and current traces the larger the effects). Thus, there seem to be qualitative differences in the way in which the most recent and the less recent past influence processing. The exact cognitive/neural basis for these differences are currently not well understood. Clearly, this is a theoretically important issue that deserves further investigation.
Conflict and LTM Encoding/Retrieval
According to our results, presence of conflict modulates the cost asymmetry at two points. First, and not surprisingly, across all experiments the cost asymmetry was increased (albeit not quite significantly so in Exp. 5) when stimuli associated with the non-dominant task (i.e., the central cues) were present while performing the dominant task. This result is consistent with findings in the standard task-switching paradigm (e.g., Yeung & Monsell, 2003a) according to which the cost asymmetry is modulated through the presence of stimulus and/or response-related conflict. From the LTM perspective, this can be explained by assuming that the endogenous stimulus serves as a particularly powerful retrieval cue for the currently irrelevant (endogenous) task.
Second, and theoretically more interesting is the fact that the presence of the exogenous stimulus (i.e., the sudden onset) while performing the endogenous task is apparently a necessary condition for the encoding of the encoding memory traces that later lead to selection costs (Experiment 1 and 2). This result is consistent with the idea that conflict boosts in a general manner attention on the currently relevant processes (i.e., attending to the endogenous cue), which in turn promotes encoding of memory instances of that particular selection episode (Bryck & Mayr, 2008; Verguts & Noetebaert, 2009).
Another possible interpretation of this result is that when exposed to an exogenous stimulus during the endogenous task, subjects encode a “suppression response” along with this stimulus (for a similar explanation of negative priming effects, see Rothermund, Wentura, & DeHouver, 2005). This suppression response is then retrieved during the post-interruption trials in the exogenous task and interferes with the now appropriate orienting towards the exogenous stimulus. In future experiments it will be important to distinguish between these accounts. For example, if conflict generally boosts encoding of memory traces, we should find the cost asymmetry even when that conflict is elicited in a different way (e.g., through incongruent flanker stimuli) than through the exogenous stimuli.
In Experiment 2 we also tested a condition in which experience with conflict during the endogenous task was limited to post-interruption trials. We did this with the goal to roughly equate the conflict experience during the endogenous task to that during the exogenous task, where subjects are able to effectively filter out the interfering information from the endogenous task during maintenance trials. Indeed, we found that in this situation the cost asymmetry was reduced. Thus, it seems that in the standard situation, frequent experiences with conflict during the endogenous task are responsible for the large post-interruption costs. Obviously, this type of a positive relationship between amount of interference and frequency of encoding opportunities is consistent with an LTM account, such as instance theory (e.g., Logan, 1988).
The results from this experiment also suggest a possible reason why the cost asymmetry occurs in the first place: The effective filtering during maintenance of the exogenous control setting prevents the encoding of potentially interfering memory traces, whereas the relatively ineffective filtering in the endogenous task allows the encoding of such traces. This leads to a testable prediction: In situations in which filtering is disrupted or for individuals with filtering problems, the cost asymmetry should be weakened or even eliminated. Interestingly, in the above-mentioned initial results with older adults (who show no efficient maintenance/filtering in the exogenous task) we actually do find an absence of a clear cost asymmetry between the exogenous and the endogenous task.
We have not yet completely resolved the question how exactly selection costs in general and more specifically the cost asymmetry arise. However, our results clearly indicate that aspects of the initial encoding situation (e.g., type and frequency of specific selection instances), critically determine the potency of LTM traces, which then eventually lead to the costs of selecting between competing control settings.
When and Why are Interruptions Harmful?
It is a truism that interruptions of ongoing activities harm fluent and effective performance. However, we currently do not have a full understanding of when and why exactly interruptions—an omnipresent reality in most real-world work environments—actually do have negative effects.
One thing we do know is that at least after interruptions of cumulative tasks (i.e., where one needs to take off exactly where one stopped before the interruption) there is a time cost in terms of re-establishing the current task context in working memory (e.g., Altman & Trafton, 2007). The current results point to an additional factor. If our explanation of the cost-asymmetry is correct then every recovery from an interruption will force the system into an updating state during which it is vulnerable to alternative paths of action. Take for example a typical, complex work situation with multiple tasks that need to be performed before the day is done. These additional demands may have little effect while one is absorbed in the currently prioritized task. However, once pulled away (e.g., through the email inbox signal) the return to that task may easily go astray because that requires crossing a choice point during which the system is temporarily open to all alternative paths of action that are currently activated in LTM. Thus, one potential danger of interruptions may lie in increasing the number of these choice points, a hypothesis that can be tested empirically and that may have important practical consequences for how to operate in or design multi-tasking environments.
Conclusion
The current work allows two broad conclusions. First, while exogenous control of attention may be fast and effortless, the process of intentionally adopting such a control setting produces larger behavioral costs than when adopting an endogenous control setting. Second, our pattern of results suggests that at least two things need to come together to produce interference when adopting an exogenous task setting: the presence of interfering LTM traces and an updating working memory mode, as triggered for example while recovering from an interruption. We propose that this model also provides a more general explanation of the types of costs regularly obtained in task-switching situations than the assumption of trial-to-trial carry-over between competing tasks or control settings.
Highlights.
Subjects performed automatic or controlled tasks in alternating pure blocks.
We found large post-interruption costs, in particular for automatic tasks.
These large costs were eliminated in the absence of experience with competing tasks.
Interruption-induced updating opens working memory to long-term memory interference.
Results challenge the idea that task-switch costs reflect carry-over from last trial.
Footnotes
A separate discussion in the literature deals with the question to what degree task switching requires a special “reconfiguration process” (e.g., Allport & Wylie, 2000; Monsell, 2003). We ignore this issue here because it is largely orthogonal to the question where interference originates. An important purpose of such a configuration process could be to implement a task-specific attentional filter that could counteract any type of interference, no matter whether it originates from the previous trial or from LTM.
The term “neutral” should not be taken too literally as there is no such thing as a truly neutral task. What we refer to here is that the interruption task does not induce the kind of between-task conflict that is critical for standard task-switch costs. In particular, note that the interruption task shares no stimulus aspects with any of the two primary tasks, thus eliminating any ambiguity about whether or not the interruption task should be performed on a particular trial.
The pilot experiment used the exact same stimulus presentation procedures as in Experiment 1, but without the interruption task, with conflict present on each trial, and with tasks randomly cued through visually presented verbal cues occurring either 200 ms or 700 ms into the 1000 ms response-stimulus interval. The words “central” or “inside” were used as cues for the endogenous task and the words “surround” or “outside” were used as cues for the exogenous task. To avoid cue-priming effects (e.g., Mayr & Kliegl, 2003), cues were never repeated (following Monsell & Masson, 2006). Performance was assessed in four 80-trial blocks with a switch rate of 33%. We found a strong switch-cost asymmetry: RT switch costs were exactly twice as large for the exogenous, M=208 ms (SD=192) as for the endogenous task, M=104 (SD=133), F(1,19)=8.70, p<.01, MSE=6232.32, and this effect was not modulated through the cue-stimulus interval. There were no reliable error effects.
The math task was considerably harder than the anagram task, both in terms of RTs and error rates. Thus, it is worth checking to what degree the observed cost asymmetry may be limited to the difficult math task, but is less apparent for the easier anagram task. A clean contrast is only possible for the 1:2 mapping group where half of the subjects worked only with the math task and the other half worked only with the anagram task. We included interruption task as a between-subject factor along with the within-subject factors of the main analysis. The only effect that involved the between-subject factor was a four-way interaction between interruption task, task (i.e., endogenous vs. exogenous), interruption, and conflict that approached significance, F(1,18)=3.72, MSE=4918.26, p=.07. For subjects who were interrupted through the anagram task, the critical cost asymmetry was somewhat larger than for subjects who were interrupted by the math task. While we cannot conclude from this result that the observed asymmetry is larger for easier tasks, it certainly provides no indication that it is limited to difficult interruptions.
In interpreting accuracy effects it is important to keep in mind that in the exogenous/endogenous paradigm response selection difficulty was not correlated with input selection difficulty (i.e., exogenous/endogenous conflict). In fact, high selection demands on the input side (e.g., due to increased interference from the endogenous cue on post-interruption trials) may produce slack time for response-selection difficulties to dissipate. In the current paradigm such response-selection difficulties can arise for the exogenous task in form of a Simon-type effect as subjects had to make a left-right response to a stimulus that appears either on the left or the right side of the screen. Further, subjects may have greater difficulties with warding off such Simon-type interference in the low-demand condition due to contextual carry-over from an interruption task that required responding to the location of a stimulus. In fact, when we repeated the main RT and error analyses after eliminating those trials that should produce the largest Simon-type effects (e.g., when the exogenous stimulus occurs either on the leftmost or the rightmost side of the stimulus circle), the difference between attentional demand conditions on the error cost-asymmetry was no longer reliable, F(1,38)=2.90, MSE=17.36, p=.1, while the RT cost asymmetry remained highly reliable in either of the two conditions, Fs(1,38)>11.19.
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Allport A, Wylie G. Task-switching, stimulus–response bindings, and negative priming. In: Monsell S, Driver JS, editors. Attention and performance XVIII: Control of cognitive processes. Cambridge, MA: MIT Press; 2000. pp. 35–70. [Google Scholar]
- Altman EM. Comparing switch costs: Alternating runs and explicit cuing. Journal of Experimental Psychology: Learning, Memory, and Cognition. 2007;33:475–483. doi: 10.1037/0278-7393.33.3.475. [DOI] [PubMed] [Google Scholar]
- Altman EM, Trafton JG. Timecourse of recovery from interruption: Data and a model. Psychological Bulletin & Review. 2007;14:1079–1084. doi: 10.3758/bf03193094. [DOI] [PubMed] [Google Scholar]
- Botvinick MM, Braver TS, Barch DM, Carter CS, Cohen JD. Conflict monitoring and cognitive control. Psychological Re- view. 2001;108:624–652. doi: 10.1037//0033-295x.108.3.624. [DOI] [PubMed] [Google Scholar]
- Bryck R, Mayr U. Task selection cost asymmetry without task switching. Psychonomic Bulletin and Review. 2008;15:128–134. doi: 10.3758/pbr.15.1.128. [DOI] [PubMed] [Google Scholar]
- Crump MJC, Logan GD. Contextual control over task-set retrieval. Attention, Perception and Psychophysics. 2010;72:2047–2053. doi: 10.3758/bf03196681. [DOI] [PubMed] [Google Scholar]
- Durstewitz D, Kelc M, Güntürkün O. A neurocomputational theory of dopaminergic modulation of working memory functions. The Journal of Neuroscience. 1999;19:2807–1822. doi: 10.1523/JNEUROSCI.19-07-02807.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gilbert S, Shallice T. Task switching: A PDP model. Cognitive Psychology. 2002;44:297–337. doi: 10.1006/cogp.2001.0770. [DOI] [PubMed] [Google Scholar]
- Gopher D, Armony L, Greenshpan Y. Switching tasks and attention policies. Journal of Experimental Psychology: General. 2000;129:308–339. doi: 10.1037//0096-3445.129.3.308. [DOI] [PubMed] [Google Scholar]
- Hintzman D. “Schema abstraction” in a multiple-trace memory model. Psychological Review. 1986;93:411–428. [Google Scholar]
- Hommel B. Event files: Feature binding in and across perception and action. Trends in Cognitive Sciences. 2004;8:494–500. doi: 10.1016/j.tics.2004.08.007. [DOI] [PubMed] [Google Scholar]
- Jonides J. Voluntary versus automatic control over the mind's eye. In: Long J, Baddeley A, editors. Attentionand performance IX. Hillsdale, NJ: Erlbaum; 1981. pp. 187–203. [Google Scholar]
- Kiesel A, Steinhauser M, Wendt M, Falkenstein M, Jost K, Philipp AM, Koch I. Control and interference in task switching—A review. Psychological Bulletin. 2010;136:849. doi: 10.1037/a0019842. [DOI] [PubMed] [Google Scholar]
- Koch I. Automatic and intentional activation of task sets. Journal of Experimental Psychology: Learning, Memory, and Cognition. 2001;27:1474–1486. doi: 10.1037//0278-7393.27.6.1474. [DOI] [PubMed] [Google Scholar]
- Lien MC, Ruthruff E. Task switching in a hierarchical task structure: Evidence for the fragility of the task repetition benefit. Journal of Experimental Psychology: Learning, Memory, and Cognition. 2004;30:697–713. doi: 10.1037/0278-7393.30.3.697. [DOI] [PubMed] [Google Scholar]
- Logan GD. Toward an instance theory of automatization. Psychological Review. 1988;95:492–527. [Google Scholar]
- Logan GD. Repetition priming and automaticity: Common un- derlying mechanisms? Cognitive Psychology. 1990;22:1–35. [Google Scholar]
- Mayr U. Age differences in the selection of mental sets: The role of inhibition, stimulus ambiguity, and response-set overlap. Psychology and Aging. 2001;16:96–109. doi: 10.1037/0882-7974.16.1.96. [DOI] [PubMed] [Google Scholar]
- Mayr U. What matters in the cued task-switching paradigm: Tasks or cues? Psychonomic Bulletin and Review. 2006;5:794–799. doi: 10.3758/bf03193999. [DOI] [PubMed] [Google Scholar]
- Mayr U, Bryck RL. Sticky rules: Integration between abstract rules and specific actions. Journal of Experimental Psychology: Learning, Memory, & Cognition. 2005;31:337–350. doi: 10.1037/0278-7393.31.2.337. [DOI] [PubMed] [Google Scholar]
- Mayr U, Bryck RL. Outsourcing Control to the Environment: Effects of Stimulus/Response Locations on Task Selection. Psychological Research. 2006;71:107–116. doi: 10.1007/s00426-005-0039-x. [DOI] [PubMed] [Google Scholar]
- Melara RD, Algom D. Driven by information: a tectonic theory of Stroop effects. Psychological Review. 2003;110:422. doi: 10.1037/0033-295x.110.3.422. [DOI] [PubMed] [Google Scholar]
- Monsell S. Task switching. Trends in Cognitive Sciences. 2003;7:134–140. doi: 10.1016/s1364-6613(03)00028-7. [DOI] [PubMed] [Google Scholar]
- Monsell S, Mizon GA. Can the task-cueing paradigm measure an “endogenous” task-set reconfiguration process? Journal of Experimental Psychology: Human Perception and Performance. 2006;32(3):493–516. doi: 10.1037/0096-1523.32.3.493. [DOI] [PubMed] [Google Scholar]
- Müller HJ, Rabbitt PMA. Reflexive and Voluntary Orienting of Visual Attention: Time Course of Activation and Resistance to Interruption. Journal of Experimental Psychology: Human Perception and Performance. 1989;15:315–330. doi: 10.1037//0096-1523.15.2.315. [DOI] [PubMed] [Google Scholar]
- O'Reilly RC. Biologically Based Computational Models of High-Level Cognition. Science. 2006;314:91–94. doi: 10.1126/science.1127242. [DOI] [PubMed] [Google Scholar]
- Posner MI. Orienting of attention. Quarterly Journal of Experimental Psychology. 1980;32:3–25. doi: 10.1080/00335558008248231. [DOI] [PubMed] [Google Scholar]
- Rothermund K, Wentura D, De Houwer J. Retrieval of incidental stimulus-response associations as a source of negative priming. Journal of Experimental Psychology: Learning, Memory, and Cognition. 2005;31:482. doi: 10.1037/0278-7393.31.3.482. [DOI] [PubMed] [Google Scholar]
- Spieler D, Mayr U, LaGrone S. Outsourcing cognitive control to the environment: Adult-age differences in the use of task cues. Psychonomic Bulletin and Review. 2006;5:787–793. doi: 10.3758/bf03193998. [DOI] [PubMed] [Google Scholar]
- Theeuwes J. Top-down search strategies cannot override attentional capture. Psychonomic Bulletin & Review. 2004;11:65–70. doi: 10.3758/bf03206462. [DOI] [PubMed] [Google Scholar]
- Theeuwes J, Kramer AF, Hahn S, Irwin DE. Our eyes do not always go where we want them to go: capture of the eyes by new objects. Psychological Science. 1998;9:379–385. [Google Scholar]
- Tzelgov J, Henik A, Berger J. Controlling Stroop effects by manipulating expectations for color words. Memory & Cognition. 1992;20:727–735. doi: 10.3758/bf03202722. [DOI] [PubMed] [Google Scholar]
- Vandierendonck A, Liefooghe B, Verbruggen F. Task switching: Interplay of reconfiguration and interference. Psychological Bulletin. 2010;136:601–626. doi: 10.1037/a0019791. [DOI] [PubMed] [Google Scholar]
- Verbruggen F, Logan GD. Automaticity of cognitive control: Goal priming in response-inhibition paradigms. Journal of Experimental Psychology: Learning, Memory, and Cognition. 2009;35:1381–1388. doi: 10.1037/a0016645. [DOI] [PubMed] [Google Scholar]
- Verguts T, Notebaert W. Adaptation by binding: A learning account of cognitive control. Trends in Cognitive Sciences. 2009;13:252–257. doi: 10.1016/j.tics.2009.02.007. [DOI] [PubMed] [Google Scholar]
- Waszak F, Hommel B, Allport A. Task-switching and long-term priming: Role of episodic stimulus–task bindings in task- shift costs. Cognitive Psychology. 2003;46:361–413. doi: 10.1016/s0010-0285(02)00520-0. [DOI] [PubMed] [Google Scholar]
- Waszak F, Hommel B, Allport A. Interaction of task readiness and automatic retrieval in task switching: Negative priming and competitor priming. Memory & Cognition. 2005;33:595–610. doi: 10.3758/bf03195327. [DOI] [PubMed] [Google Scholar]
- Wylie GR, Javitt DC, Foxe JJ. Don't think of a white bear: An fMRI investigation of the effects of sequential instructional sets on cortical activity in a task-switching paradigm. Human brain mapping. 2004;21:279–297. doi: 10.1002/hbm.20003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yeung N, Monsell S. Switching between tasks of unequal familiarity: The role of stimulus-attribute and response-set selection. Journal of Experimental Psychology: Human Perception and Performance. 2003a;29:455–469. doi: 10.1037/0096-1523.29.2.455. [DOI] [PubMed] [Google Scholar]
- Yeung N, Monsell S. Effects of recent practice on task switching. Journal of Experimental Psychology: Human Perception and Performance. 2003b;29:919–436. doi: 10.1037/0096-1523.29.5.919. [DOI] [PubMed] [Google Scholar]
- Yeung N, Nystrom LE, Aronson JA, Cohen JD. Between-task competition and cognitive control in task switching. The Journal of Neuroscience. 2006;26:1429–1438. doi: 10.1523/JNEUROSCI.3109-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]