

Abstract
Niche complementarity in resource use has been proposed as a key mechanism to explain the positive effects of increasing plant species richness on ecosystem processes, in particular on primary productivity. Since hardly any information is available for niche complementarity in water use, we tested the effects of plant diversity on spatial and temporal complementarity in water uptake in experimental grasslands by using stable water isotopes. We hypothesized that water uptake from deeper soil depths increases in more diverse compared to low diverse plant species mixtures. We labeled soil water in 8 cm (with 18O) and 28 cm depth (with ²H) three times during the 2011 growing season in 40 temperate grassland communities of varying species richness (2, 4, 8 and 16 species) and functional group number and composition (legumes, grasses, tall herbs, small herbs). Stable isotope analyses of xylem and soil water allowed identifying the preferential depth of water uptake. Higher enrichment in 18O of xylem water than in ²H suggested that the main water uptake was in the upper soil layer. Furthermore, our results revealed no differences in root water uptake among communities with different species richness, different number of functional groups or with time. Thus, our results do not support the hypothesis of increased complementarity in water use in more diverse than in less diverse communities of temperate grassland species.
Introduction
Many results from experimental biodiversity research support the hypothesis that increased plant species richness has positive effects on several aspects of ecosystem functioning [1–4], such as plant biomass production aboveground [5–8], whereas the underlying mechanisms for these positive effects are not yet fully understood [9]. One frequently proposed explanation is niche complementarity [10, 11], assuming that partitioning of resources such as light, nutrients or water reduces competitive interactions among the species of a mixture. Consequently, resource exploitation at the community level is more complete, resulting in greater productivity compared to a monoculture or low diverse mixture. Partitioning of belowground resources might be achieved spatially via different root distribution patterns or temporally because of differences in phenology among species [12, 13]. However, experimental evidence, particularly for the resource water, is still sparse.
Furthermore, the hypothesis of complementary resource use was mainly tested indirectly, for instance by comparing aboveground biomass production in mixtures with values expected from monocultures [14–16] or by interpreting a more complete filling of available biotope space, i.e., soil depth and volume, indicated by increased vertical root biomass distribution with increasing species richness as greater complementarity [17–19]. In addition, complementary water use was suggested based on increased evapotranspiration rates in plant mixtures with increasing species richness [20] or based on lower δ13C values in mixtures compared to monocultures [21]. Although water is an important resource for plant performance, there is, to our knowledge, a lack of direct measurements to assess water partitioning in mixtures and to test complementarity in water use with increasing species richness under field conditions.
Stable water isotopes have often been applied to directly estimate the water source used by plants (e.g., water of different soil depths or even fog) and were used in many studies aiming to explain coexistence of plants in different natural ecosystems [22–27]. Potential water sources of co-occurring species were identified by comparing the natural abundance of oxygen or hydrogen isotopes in xylem water and soil water of different depths. As no isotopic fractionation occurs during water uptake, the isotopic signal of the xylem water reflects the signal of a plant’s water source [28, 29]. In herbaceous plants, it has been shown that the isotopic signal of xylem water in the root crown was the best indicator of the water source [30]. However, natural abundance analyses rely on a pronounced isotopic profile of soil water, which is often not given under field conditions [31]. More unequivocal results can be obtained by enriching the soil water at different depths with different stable water isotopes [32, 33].
Thus, we carried out a labeling experiment in the Jena Experiment [34] and applied water enriched in stable water isotopes (oxygen and hydrogen) at two different depths three times during the growing season 2011. The Jena Experiment is a large grassland biodiversity experiment with communities of varying species richness and functional group number, based on a pool of 60 temperate grassland species which greatly differ in their functional characteristics (grasses, legumes, tall herbs, small herbs). Based on the niche complementarity theory, we expected (i) uptake of water from distinctly different soil layers with increasing species richness or functional group number, and (ii) functional characteristics, i.e., functional group identity, to explain spatial and seasonal variations in water uptake patterns.
Material and Methods
Study site
The Jena Experiment is a large temperate grassland biodiversity experiment located in the floodplain of the Saale river near the city of Jena (Germany, 50°55’N, 11°35’E, 130 m a.s.l.), which was established in 2002 on a former arable field. Since the Jena Experiment field site is not subjected to nature conservation and our study did not involve endangered or protected species, no specific permission was required. The soil is a Eutric Fluvisol developed from up to 2 m thick fluvial sediments. Mean annual precipitation is 587 mm, mean annual temperature is 9.3°C [35]. The Jena Experiment consists of 82 plots with different plant species number (1, 2, 4, 8, 16 and 60 species) and functional group richness (1, 2, 3 and 4 functional groups), from a species pool of 60 species assigned to four plant functional groups (grasses, legumes, small herbs and tall herbs). The experimental plots are arranged in four blocks to account for a gradient in soil texture, ranging from sandy loam to silty clay with increasing distance from the river. All plots are weeded three times per year (April, June and September) and mown two times per year (June, September) to mimic the management of extensive hay meadows.
Tracer application and field sampling
The tracer experiment was conducted at the start of the growing season (April) and during the regrowth after the first and the second mowing (June and September) 2011. The experiment was carried out on a subset of 40 plots, covering a species richness gradient with well-established 2, 4, 8 and 16 plant species mixtures (ten replicates per species richness level, list of mixtures in S1 Table). These plots were selected to be equally distributed among the experimental blocks and to cover a balanced distribution of all functional group richness levels in each species richness level. At each plot, three subplots were established (44 cm × 56 cm), each for one of the three labeling campaigns of the tracer experiment.
About five days before starting the tracer application, plant and soil samples were collected 10 cm next to the study plots to identify the natural abundance of 18O and 2H (later referred to as background samples). Using a soil auger of 1 cm diameter (Eijkelkamp, The Netherlands), soil background samples were taken at one plot per species richness level in each of the four blocks in 0–10, 10–20, 20–30 and 30–40 cm soil depth, resulting in a total of 64 background samples per campaign. Root crowns, the connection between above- and belowground tissues, of single plants were collected and immediately placed into 12 ml glass vials (Labco Limited, UK), sealed with a cap and parafilm, and frozen until cryogenic water extraction was carried out. In total, 49 root crown background samples, homogenously distributed along the species richness gradient and representing species of each functional group in each species richness level, were collected prior each campaign.
For the tracer experiment, labeled water (1H2 18O, Sigma-Aldrich, Germany, and 2H2O, Euriso-top, France) was injected at the same subplot (44 cm × 56 cm), but in different soil depths (1H2 18O at 8 cm and 2H2O at 28 cm depth). To achieve a homogenous tracer distribution within the subplots, injection points were arranged on a grid of seven horizontal lines, which had a distance of 8.7 cm. The injection points for the two depths were alternating along the lines with a distance of 10 cm. This resulted in 32 injection points for the upper and 31 injection points for the lower soil depth (Application scheme in S1 Fig.). Holes of 8 mm diameter were drilled down to the two target depths of 8 and 28 cm with a handheld automated drill during five days prior to labeling, and stabilized with wooden sticks.
The tracer solutions were created to achieve an enrichment of 400 ‰ for 18O (upper soil depth) and 800 ‰ for 2H (lower soil depth), based on the average soil water content of all plots measured a few days prior to labeling. Thus, the following tracer solutions were created and added to the soil water: 8’700 ‰ δ18O and 26’500 ‰ δ2H in April (18 to 19 April 2011), 12’100 ‰ δ18O and 33’000 ‰ δ2H in June (27 to 28 June 2011) and 15’500 ‰ δ18O and 39’000 ‰ δ2H in September (27 to 28 September 2011). The tracer solutions were applied at 20 subplots per day between 8 am and 4 pm. The respective tracer solution was applied at 3 cm (18O-enriched water) or 23 cm depth (2H-enriched water) within 30 min per depth, using a 3 mm diameter four-sideport needle connected by a silicon tube to a bottle top dispenser (Sartorius, Germany) put on a 1 L glass bottle. The injection depth differed from the drilled depth to prevent overflow out of the drilled holes in the upper depth during the injections, as the tracer solutions infiltrated into the soil rather slowly. Each hole received 2 ml of the respective tracer solution, resulting in a total of 64 ml for the upper depth and 62 ml for the lower depth per subplot. A funnel was placed around the injection hole to prevent contamination of the vegetation with the tracer solution during tracer application.
Exactly 48 h after finishing the labeling of each subplot (20 to 21 April, 29 to 30 June and 29 to 30 September 2011), root crowns of three to five individual plants of each species present per plot were collected, cleaned and pooled by plant species and subplot. Three soil samples were taken at each subplot with a soil auger of 1 cm diameter (Eijkelkamp, The Netherlands) in nine soil depths (0–3, 3–6, 6–10, 10–15, 15–20, 20–23, 23–26, 26–30 and 30–40 cm). One soil replicate was taken very close to an injection point for the upper soil depth, one very close to an injection point for the lower depth, and one in between injection points. Soil samples in each depth were pooled, resulting in nine soil samples per subplot, covering the top 40 cm. All plant and soil samples were immediately placed into 12 ml glass vials (Labco Limited, UK), sealed with a cap and parafilm, kept cool in a cooling box and transported to a freezer within two hours. Samples were kept frozen until cryogenic water extraction. In total, 360 soil samples were taken and analyzed at each labeling campaign. In addition 197 plant samples were taken in April, 192 in June, and 193 in September. Due to the low water content of some plant samples, only 148, 136 and 145 samples were analyzed for each campaign, respectively.
Laboratory analyses
Xylem water in root crowns and soil water were extracted for isotopic analyses using a cryogenic water extraction line [30] and measured with a TC/EA high-temperature conversion/elemental analyzer coupled with a DeltaplusXP isotope ratio mass spectrometer via a ConFlo III interface (Thermo-Finnigan, Bremen, Germany; see Werner et al. [36] for further information). Oxygen and hydrogen isotopic compositions of the water samples are given in δ notation measured as (RSample/RStandard)—1, and expressed in ‰. R is the ratio of 18O to 16O or 2H to 1H of the sample or the standard. Our standard was a working control standard, regularly calibrated against international standards (V-SMOW, SLAP, GISP). The overall precision of the measurements was ± 0.09 ‰ for δ18O and ± 0.37 ‰ for δ2H.
Data analyses
All statistical analyses and graphics were done with R 2.14.1 (R Development Core Team 2011). Mixed effects models were carried out by using the lmer function within the lme4 package [37]. Prior to analyses, all data were log transformed to meet the assumptions for mixed effects models that require normally distributed within-group errors. The maximum likelihood method was used to estimate the variance components. Block, plot identity (nested within block) and species identity were treated as random factors. Analyses were started from a null model containing the random factors. Fixed factors and interactions between the fixed factors were entered stepwise. Likelihood ratio tests (Χ 2) were applied to compare models and to test for a significant improvement of the model after adding the fixed effects.
To compare whether the δ18O or δ2H values in the xylem water of the samples taken after the labeling differ from the background samples, mixed-effect models were carried out, including sample type (i.e., back ground sample or labeled sample) as fixed factor separately for each labeling campaign.
Enrichment of the xylem water was then identified by calculating the difference of δ18O or δ2H values of the samples taken after the labeling and the respective average value of the plant background samples for each labeling time. Finally, effects of species richness, number of functional groups and functional group identity (i.e., grasses, legumes, small herbs and tall herbs) on uptake of 18O- or 2H-enriched water were tested for each labeling campaign by adding the fixed factors in the following order: species richness (SR, log-linear), functional group richness (FR, linear), functional group identity (FG), and the interaction between SR and FG.
Results
δ18O and δ2H values of soil water
Soil water in a depth of 6 to 10 cm, where the 18O-enriched water was injected, displayed average δ18O values across all species richness levels of 65.5 ‰ (SD ± 39.45 ‰) in April, 106.7 ‰ (SD ± 44.14 ‰) in June, and 85 ‰ (SD ± 45.64 ‰) in September (Fig. 1), highly enriched compared to the respective background values (δ18OApril = -9.67 ‰ (SD ± 1.15 ‰), δ18OJune = -5.08 ‰ (SD ± 0.83 ‰) and δ18OSeptember = -2.79 ‰ (SD ± 1.98 ‰). Similarly, soil water in a depth of 26 to 30 cm, where the 2H-enriched water was added, showed on average δ2H values of 16.9 ‰ (SD ± 99.32 ‰) in April, 262.6 ‰ (SD ± 206.21 ‰) in June, and 144 ‰ (SD ± 171.22 ‰) in September, highly above the background values (δ2HApril = -110.27 ‰ (SD ± 7.45 ‰), δ2HJune = -93.14 ‰ (SD ± 10.24 ‰) and δ2HSeptember = -50.12 ‰ (SD ± 7.61 ‰)). Soil layers above the target depths were enriched as well (Fig. 1), Mainly for δ18O, enrichment of soil water in layers below the target depth was also found at some plots. However, two distinct soil layers imitating two different water sources were achieved at all three campaigns.
Figure 1. δ18O and δ2H values of soil water before and after the labeling.

δ18O (A, C, E) and δ2H (B, D, F) values of soil water labeled with 18O-enriched water in upper soil depths and with 2H-enriched water in lower soil depths. Grey bands illustrate depths of tracer application. Data are given for the natural background soil as well as after the labeling at three different times in 2011 (April: A-B, June: C-D, September: E-F) in each case pooled for all species richness levels. Values of soil water content (SWC) in 8, 16 and 32 cm are given as mean ± 1 SD for the 4-day labeling and harvest campaigns.
During the course of the growing season, background δ18O and δ2H values increased by about 7 ‰ and 60 ‰ in the target depth (6–10 cm for 18O and 26–30 cm for 2H), driven by enhanced soil water evaporation at higher temperatures and changes in the isotopic composition of precipitation [38, 39].
δ18O and δ2H values of xylem water
Xylem water after the labeling and pooled over all species richness levels displayed average δ18O values of 14.24 ‰ (SD ± 14.21 ‰) in April, 28.88 ‰ (SD ± 21.01 ‰) in June, and 30.4 ‰ (SD ± 24.1 ‰) in September, well above the corresponding background values of -8.5 ‰ (SD ± 1.5 ‰) in April, -4.78 ‰ (SD ± 1.39 ‰) in June, and -3.35 ‰ (SD ± 1.27 ‰) in September. The δ18O values of the xylem water after the labeling were significantly higher than the δ18O values of the xylem water of the background samples at all three times (April: Χ 2 = 209.82, P < 0.001; June: Χ 2 = 220.37, P < 0.001; September: Χ 2 = 227.16, P < 0.001, Fig. 2). In contrast, δ2H values in the xylem water of the plants after the labeling did not differ significantly from background samples in April (Χ 2 = 0.87, P = 0.350) and June (Χ 2 = 1.19, P = 0.276), but in September (Χ 2 = 65.75, P < 0.001). While δ2H values of the xylem water of labeled plants were -65.37 ‰ (SD ± 14.75 ‰) in April, -43.13 ‰ (SD ± 16.78 ‰) in June and -19.16 ‰ (SD ± 9.02 ‰) in September, δ2H values of background plants were -69.81 ‰ (SD ± 9.71) in April, -45.6 ‰ (SD ± 8.05 ‰) in June, and -29.71 ‰ (SD ± 7.53 ‰) in September (Fig. 2).
Figure 2. δ18O and δ2H values of xylem water before and after the labeling.
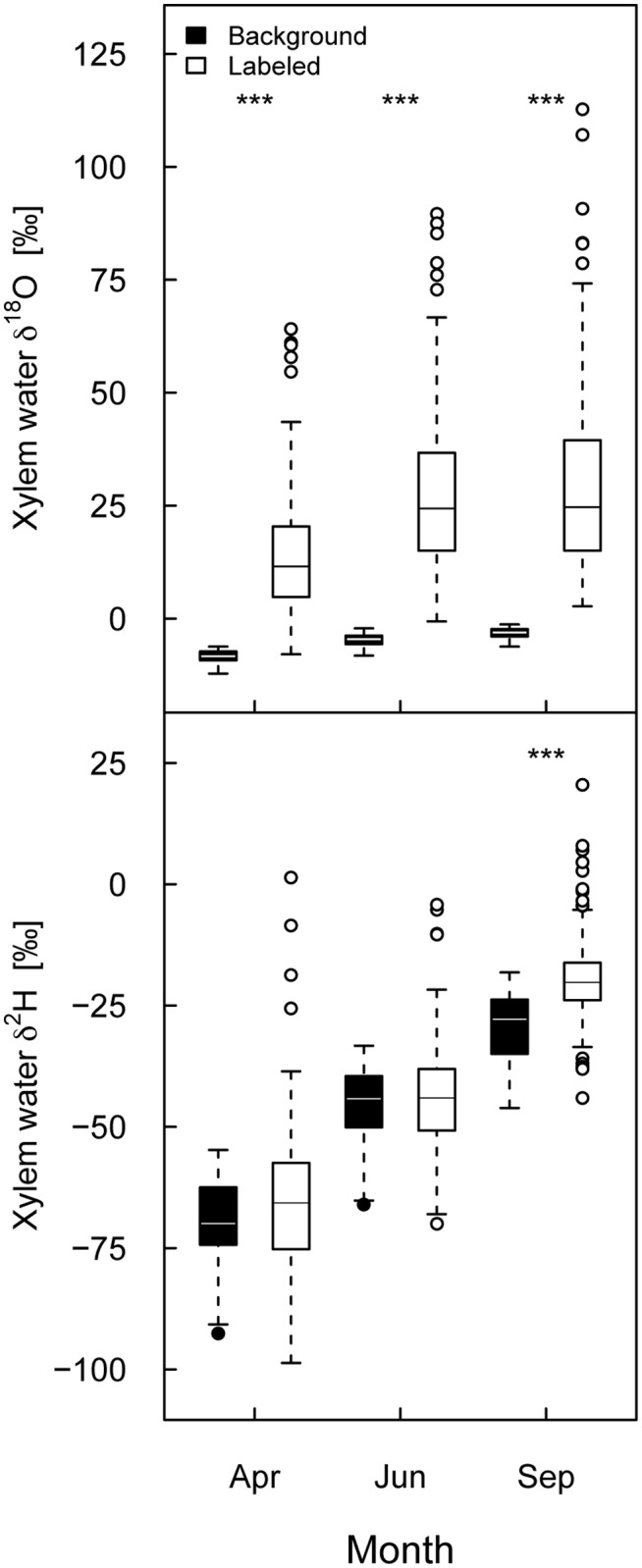
δ18O (top) and δ2H (bottom) values are given for the background samples and the samples taken after the labeling at three different times (April, June, September 2011) in each case pooled for all species richness levels. Outliers (at δ18O = 141.7 ‰ and δ2H = 101.3 ‰ in June and at δ18O = 187.3 ‰ in September) were removed for reasons of clarity. Results of the corresponding mixed-effects models are given in the running text. Significant differences among the background samples and the samples taken after labeling are indicated with asterisks (with *** referring to P < 0.001).
The enrichment of xylem water in 18O, i.e., the difference between the average seasonal background δ18O value and the δ18O values of the samples taken after the labeling, ranged between 22.74 ‰ and 33.75 ‰ during the growing season, in comparison to the much larger enrichment in the soil water that ranged between 75.12 ‰ and 111.78 ‰ at 6 to 10 cm soil depth. However, the enrichment of xylem water in 2H ranged only between 2.47 ‰ and 10.55 ‰, despite a very large enrichment in the corresponding target depth of 26 to 30 cm soil depth (127.17 ‰ to 355.69 ‰), clearly indicating preferential water uptake in the upper soil depth.
Enrichment of the xylem water in 18O or 2H was not affected by species richness or number of functional groups at any time (Table 1, Fig. 3). Functional groups only differed in their 18O enrichment in April, but not in June or September (PFG = 0.005, Table 1), with legumes displaying lower and small herbs slightly higher 18O enrichments compared to the other functional groups in April (legumes: δ18O = 11.61 ‰ (SD ± 8.14 ‰), small herbs: δ18O = 27.39 ‰ (SD ± 16.08 ‰), tall herbs: δ18O = 22.96 ‰ (SD ± 15.02 ‰), grasses: δ18O = 20.75 ‰ (SD ± 9.95 ‰). No difference among functional groups was found for 2H enrichment at any time.
Table 1. Summary of the mixed-effects model testing the effects of species richness, functional group number and functional group identity on xylem water enrichment in 18O and 2H (i.e., difference between samples taken after the labeling and the background samples).
| April | June | September | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| δ18O | δ2H | δ18O | δ2H | δ18O | δ2H | |||||||
| χ2 ratio | P | χ2 ratio | P | χ2 ratio | P | χ2 ratio | P | χ2 ratio | P | χ2 ratio | P | |
| Species richness (SR, log-linear) | 0.71 | 0.399 | 0.01 | 0.931 | 0.18 | 0.669 | 1.82 | 0.177 | 3.01 | 0.083 | 2.13 | 0.145 |
| Functional group richness (FR, linear) | 0.05 | 0.825 | 1.45 | 0.228 | 0.26 | 0.61 | 0.79 | 0.373 | 1.1 | 0.294 | 0.14 | 0.709 |
| Functional group identity (FG) | 12.94 | 0.005 | 3.79 | 0.285 | 6.11 | 0.106 | 2.13 | 0.546 | 2.62 | 0.453 | 5.82 | 0.121 |
| SR x FG | 0.19 | 0.801 | 1.6 | 0.66 | 2.37 | 0.499 | 1.73 | 0.63 | 0.85 | 0.837 | 2.58 | 0.461 |
Analyses were carried out for each campaign separately.
Figure 3. Enrichment of the xylem water in 18O or 2H along the sown species richness gradient.

Enrichment was calculated as differences in δ18O (A-C) and δ2H (D-F) values in the xylem water after the labeling compared to the corresponding background at three different times (April, June, September 2011) separately for each species richness level. Outliers (at δ18O = 146.5 ‰ and δ2H = 146.9 ‰ in June and at δ18O = 190.6 ‰ in September) were removed for reasons of clarity. Results of the corresponding mixed-effects models are given in Table 1.
Discussion
With the present study, we tested if plant communities of increased species or functional group richness exhibit increased spatial or temporal complementarity in water use compared to low diverse communities. Our results suggest that the main water uptake was from the top soil layers in all mixtures at all times, indicated by a considerable enrichment of xylem water in 18O (applied to the top soil layer) but not in 2H (applied to the deeper soil layer). We found no evidence for increased water exploitation from deeper soil layers with increasing species richness or functional group richness nor effects of functional group identity on spatial or temporal exploitation of soil water. Increased 2H enrichment of xylem water in September indicated uptake from the 2H enriched soil layer, but was not affected by increasing species or functional group richness. Thus, our results do not support the hypothesis of complementary water use as explanation for a positive biodiversity-ecosystem functioning relationship, neither spatially nor temporally.
The approach of our study was to create two distinct soil layers highly enriched in 18O (top soil layer) or 2H (deeper soil layer) to imitate two different water sources, which we generally achieved. However, in some plots, soil layers above (and below) the target depths were enriched as well (Fig. 1), most likely due to slow soil infiltration and percolation of the tracer solution when injected into the holes. A further reason might be hydraulic lift, although this has not been shown in grasslands yet [40]. The enrichment of some soil water samples below the target depth might have been caused by soil cracks or earthworm holes [41]. Furthermore, the enrichment of soil and xylem water in 18O and 2H was lower than intended. Although we aimed to achieve a homogenous distribution of the tracer solution by evenly distributing injection holes across the plots, horizontal infiltration and distribution of the tracer within the soil seemed to have been limited. Therefore, soil samples taken close to an injection point and in between injection points and then pooled by layer for each plot probably reflected a mix of enriched and unenriched soil water available for root uptake. However, since the experimental set-up was identical for all species richness levels, no bias was introduced, rendering the conclusion of preferential water uptake from the top soil layers as very robust.
Our experimental results, based on direct measurements of soil water use, contradict earlier studies that inferred water complementarity based on indirect approaches. For instance, Caldeira et al. [21] studied soil moisture patterns and plant δ13C in Mediterranean grasslands of varying species richness and interpreted lower foliar δ13C values of plants growing in mixtures than in monocultures as a result of more complete water use due to higher stomatal conductance rates. Verheyen et al. [20] considered complementary water use as the underlying mechanism for increased evapotranspiration with increasing species richness obtained from canopy surface temperature measurements. Van Peer et al. [42] reported increased water consumption with increasing species richness in heat stressed, container-grown artificial grasslands based on soil moisture measurements. However, lower δ13C values and thus higher stomatal conductance rates can also be the result of low light levels due to higher community biomass, which could in turn increase community evapotranspiration and lower canopy temperature. Thus, these indirect approaches cannot be used to unequivocally disentangle cause and effects.
On the other hand, studies using stable isotopes to directly test water uptake among coexisting species found strong evidence for water partitioning, but typically in semi-arid ecosystems, where water availability is naturally limited [12, 22, 26, 32, 43–45]. However, none of these studies tested different species richness levels. Thus, spatial niche differentiation seems more likely to allow for coexistence when water in upper soil layers is scarce than under conditions when water is not a limiting resource (see soil water content given in Fig. 1). Our results are also in line with the stress-gradient hypothesis, assuming low levels of competitive interactions in plant communities under low levels of stress [46]. Under such conditions, water availability is closely linked to nutrient availability, which is higher in upper than in deeper soil layers of the Jena Experiment (personal communication Markus Lange, see also [47–49]), thus favoring the development of a shallow rooting system [50], even along a diversity gradient.
Furthermore, complementarity in belowground resources use (water, nutrients) is thought to result from an increased dissimilarity of rooting depths among species with increasing species richness. Hence, vertical root biomass distribution is expected to change in favor of increasing root biomass also in deeper soil layers with increasing species richness. However, Ravenek et al. [51] did not find any shifts in relative root distribution along the vertical soil profile with increasing species richness or in plots with different functional group composition. The ratio of standing root biomass among different soil layers remained quite similar, despite a significant increase in total standing root biomass at higher species richness levels. These results give further support for a lack of vertical niche differentiation with increasing species richness, but rather show preferential resource uptake from the upper soil layers independent of species or functional group richness.
Clear experimental evidence for complementarity is also scarce for other soil resources, e.g., nitrogen. In two grassland studies, both conducting 15N labeling experiments, neither spatial nor temporal complementarity of nitrogen uptake was found in more diverse grasslands compared to low diverse grasslands [52, 53]. In both studies, the main nitrogen uptake was from the top soil layer (upper 3 cm).
Ecosystem processes have been found to be highly influenced by the functional group composition rather than by species richness alone [54, 55]. Kahmen et al. [52] observed significant differences in nitrogen uptake among different functional groups (legumes, tall herbs, legumes, small herbs), irrespective of species richness level. In our study, differences in water uptake among functional groups were not significant except for April 2011, the very start of the growing season when growth commences. Based on information derived from the literature, small herbs are assumed to have shallower roots than tall herbs, grasses and legumes in the Jena Experiment [56, 57], but roots of most species cover the depths studied with our labeling approach and root characteristics vary greatly among species within functional groups. This variation may explain the lack of a consistent functional group effect on water uptake patterns in our experiment.
In conclusion, our results suggest no increased complementarity in water use with increasing species richness. The main water uptake from the top soil layer is consistent with observed rooting patterns as well as with results on nitrogen uptake found in other temperate grasslands. If complementarity in water use differs between systems adapted to low vs. high water availability remains to be seen. Furthermore, since plant species are often limited by multiple resources and differ in their resource requirements [10, 58], complementarity not only for a single resource, but for multiple resources might be the mechanism to explain the positive effects of high plant species richness on ecosystem processes.
Supporting Information
(DOCX)
(XLSX)
Acknowledgments
We are grateful to Hans de Kroon, Arthur Gessler, Liesje Mommer and Michael Scherer-Lorenzen for great support in designing the experiment as well as helping during the field campaigns. Furthermore we thank Thomas Schröder-Georgi, Alexandra Bähring, Anja Kahl, Carsten Jesch, Georg Doebel, Victor Malakov, Hannie de Caluwe, Jan Willem van der Paauw, Annemiek Smit-Teikstra and many students for their help during the preparation of the experiment, the labeling and the harvests. We acknowledge Anne Ebeling for the coordination of the Jena Experiment and the gardeners for maintaining the field. We also thank Rolf Siegwolf for helping out with 18O-labeled water, sample analyses, and him and Johanna Spiegel for useful discussions.
Data Availability
Data are available from the ETH Grassland Sciences Data Archive and can be obtained via data@usys.ethz.ch.
Funding Statement
This study was financially supported by the Swiss National Science Foundation (315230E-131194, www.snf.ch) and the German Science Foundation (FOR 456/1451, www.dfg.de). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1. Balvanera P, Pfisterer AB, Buchmann N, He J-S, Nakashizuka T, et al. (2006) Quantifying the evidence for biodiversity effects on ecosystem functioning and services. Ecology Letters 9: 1146–1156. 10.1111/j.1461-0248.2006.00963.x [DOI] [PubMed] [Google Scholar]
- 2. Isbell F, Calcagno V, Hector A, Connolly J, Harpole WS, et al. (2011) High plant diversity is needed to maintain ecosystem services. Nature 477: 199–202. 10.1038/nature10282 [DOI] [PubMed] [Google Scholar]
- 3. Schläpfer F, Schmid B (1999) Ecosystem effects of biodiversity: A classification of hypotheses and exploration of empirical results. Ecological Applications 9: 893–912. 10.1890/1051-0761(1999)009[0893:EEOBAC]2.0.CO;2 [DOI] [Google Scholar]
- 4. Allan E, Weisser W, Fischer M, Schulze E-D, Weigelt A, et al. (2013) A comparison of the strength of biodiversity effects across multiple functions. Oecologia: 1–15. [DOI] [PubMed] [Google Scholar]
- 5. Tilman D, Reich PB, Knops J, Wedin D, Mielke T, et al. (2001) Diversity and productivity in a long-term grassland experiment. Science 294: 843–845. 10.1126/science.1060391 [DOI] [PubMed] [Google Scholar]
- 6. Hector A, Schmid B, Beierkuhnlein C, Caldeira MC, Diemer M, et al. (1999) Plant diversity and productivity experiments in european grasslands. Science 286: 1123–1127. 10.1126/science.286.5442.1123 [DOI] [PubMed] [Google Scholar]
- 7. Marquard E, Weigelt A, Roscher C, Gubsch M, Lipowsky A, et al. (2009) Positive biodiversity–productivity relationship due to increased plant density. Journal of Ecology 97: 696–704. 10.1111/j.1365-2745.2009.01521.x [DOI] [Google Scholar]
- 8. Loreau M, Naeem S, Inchausti P, Bengtsson J, Grime JP, et al. (2001) Biodiversity and ecosystem functioning: current knowledge and future challenges. Science 294: 804–808. 10.1126/science.1064088 [DOI] [PubMed] [Google Scholar]
- 9. Hooper DU, Chapin FS, Ewel JJ, Hector A, Inchausti P, et al. (2005) Effects of biodiversity on ecosystem functioning: a consensus of current knowledge. Ecological Monographs 75: 3–35. 10.1890/04-0922 [DOI] [Google Scholar]
- 10. Tilman D, Lehman CL, Thomson KT (1997) Plant diversity and ecosystem productivity: Theoretical considerations. Proceedings of the National Academy of Sciences 94: 1857–1861. 10.1073/pnas.94.5.1857 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11. Loreau M, Hector A (2001) Partitioning selection and complementarity in biodiversity experiments. Nature 412: 72–76. 10.1038/35083573 [DOI] [PubMed] [Google Scholar]
- 12. Fargione J, Tilman D (2005) Niche differences in phenology and rooting depth promote coexistence with a dominant C4 bunchgrass. Oecologia 143: 598–606. 10.1007/s00442-005-0010-y [DOI] [PubMed] [Google Scholar]
- 13. Berendse F (1982) Competition between plant populations with different rooting depths. Oecologia 53: 50–55. 10.1007/BF00377135 [DOI] [PubMed] [Google Scholar]
- 14. Hector A, Bazeley-White E, Loreau M, Otway S, Schmid B (2002) Overyielding in grassland communities: testing the sampling effect hypothesis with replicated biodiversity experiments. Ecology Letters 5: 502–511. 10.1046/j.1461-0248.2002.00337.x [DOI] [Google Scholar]
- 15. Roscher C, Temperton VM, Scherer-Lorenzen M, Schmitz M, Schumacher J, et al. (2005) Overyielding in experimental grassland communities—irrespective of species pool or spatial scale. Ecology Letters 8: 419–429. 10.1111/j.1461-0248.2005.00736.x [DOI] [Google Scholar]
- 16. van Ruijven J, Berendse F (2005) Diversity–productivity relationships: Initial effects, long-term patterns, and underlying mechanisms. Proceedings of the National Academy of Sciences of the United States of America 102: 695–700. 10.1073/pnas.0407524102 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17. Mueller KE, Tilman D, Fornara DA, Hobbie SE (2013) Root depth distribution and the diversity–productivity relationship in a long-term grassland experiment. Ecology 94: 787–793. 10.1890/12-1399.1 [DOI] [Google Scholar]
- 18. Dimitrakopoulos PG, Schmid B (2004) Biodiversity effects increase linearly with biotope space. Ecology Letters 7: 574–583. 10.1111/j.1461-0248.2004.00607.x [DOI] [Google Scholar]
- 19. von Felten S, Schmid B (2008) Complementarity among species in horizontal versus vertical rooting space. Journal of Plant Ecology 1: 33–41. 10.1093/jpe/rtm006 [DOI] [Google Scholar]
- 20. Verheyen K, Bulteel H, Palmborg C, Olivié B, Nijs I, et al. (2008) Can complementarity in water use help to explain diversity–productivity relationships in experimental grassland plots? Oecologia 156: 351–361. 10.1007/s00442-008-0998-x [DOI] [PubMed] [Google Scholar]
- 21. Caldeira MC, Ryel RJ, Lawton JH, Pereira JS (2001) Mechanisms of positive biodiversity–production relationships: insights provided by δ13C analysis in experimental Mediterranean grassland plots. Ecology Letters 4: 439–443. 10.1046/j.1461-0248.2001.00238.x [DOI] [Google Scholar]
- 22. Ehleringer J, Phillips S, Schuster WF, Sandquist D (1991) Differential utilization of summer rains by desert plants. Oecologia 88: 430–434. 10.1007/BF00317589 [DOI] [PubMed] [Google Scholar]
- 23. Ehleringer JR, Dawson TE (1992) Water uptake by plants: perspectives from stable isotope composition. Plant, Cell & Environment 15: 1073–1082. 10.1111/j.1365-3040.1992.tb01657.x [DOI] [Google Scholar]
- 24. Gordon DR, Rice KJ (1992) Partitioning of space and water between two california annual grassland species. American Journal of Botany 79: 967–976. 10.2307/2444905 [DOI] [Google Scholar]
- 25. Grieu P, Lucero DW, Ardiani R, Ehleringer JR (2001) The mean depth of soil water uptake by two temperate grassland species over time subjected to mild soil water deficit and competitive association. Plant and Soil 230: 197–209. 10.1023/A:1010363532118 [DOI] [Google Scholar]
- 26. Nippert JB, Knapp AK (2007) Soil water partitioning contributes to species coexistence in tallgrass prairie. Oikos 116: 1017–1029. 10.1111/j.0030-1299.2007.15630.x [DOI] [Google Scholar]
- 27. Hoekstra NJ, Finn JA, Lüscher A (2014) The effect of drought and interspecific interactions on the depth of water uptake in deep- and shallow-rooting grassland species as determined by δ18O natural abundance. Biogeosciences Discuss 11: 4151–4186. 10.5194/bgd-11-4151-2014 [DOI] [Google Scholar]
- 28. Dawson TE, Mambelli S, Plamboeck AH, Templer PH, Tu KP (2002) Stable isotopes in plant ecology. Annual Review of Ecology and Systematics 33: 507–559. 10.1146/annurev.ecolsys.33.020602.095451 [DOI] [Google Scholar]
- 29. White JWC, Cook ER, Lawrence JR, Wallace S B (1985) The D/H ratios of sap in trees: Implications for water sources and tree ring D/H ratios. Geochimica et Cosmochimica Acta 49: 237–246. 10.1016/0016-7037(85)90207-8 [DOI] [Google Scholar]
- 30. Barnard RL, de Bello F, Gilgen AK, Buchmann N (2006) The δ18O of root crown water best reflects source water δ18O in different types of herbaceous species. Rapid Communications in Mass Spectrometry 20: 3799–3802. 10.1002/rcm.2778 [DOI] [PubMed] [Google Scholar]
- 31. Allison GB, Barnes CJ, Hughes MW, Leaney FWJ (1983) Effect of climate and vegetation on oxygen-18 and deuterium profiles in soils. Isotope Hydrology, IAEA, Vienna. pp. 105–122. [Google Scholar]
- 32. Kulmatiski A, Beard KH, Verweij RJT, February EC (2010) A depth-controlled tracer technique measures vertical, horizontal and temporal patterns of water use by trees and grasses in a subtropical savanna. New Phytologist 188: 199–209. 10.1111/j.1469-8137.2010.03338.x [DOI] [PubMed] [Google Scholar]
- 33. Ogle K, Wolpert RL, Reynolds JF (2004) Reconstructing plant root area and water uptake profiles. Ecology 85: 1967–1978. 10.1890/03-0346 [DOI] [Google Scholar]
- 34. Roscher C, Schumacher J, Baade J, Wilcke W, Gleixner G, et al. (2004) The role of biodiversity for element cycling and trophic interactions: an experimental approach in a grassland community. Basic and Applied Ecology 5: 107–121. 10.1078/1439-1791-00216 [DOI] [Google Scholar]
- 35. Kluge G, Müller-Westermeier G (2000) Das Klima ausgewählter Orte der Bundesrepublik Deutschland: Jena. Berichte des Deutschen Wetterdienstes 213 Offenbach/Jena. [Google Scholar]
- 36. Werner RA, Bruch BA, Brand WA (1999) ConFlo III—an interface for high precision δ13C and δ15N analysis with an extended dynamic range. Rapid Communications in Mass Spectrometry 13: 1237–1241. [DOI] [PubMed] [Google Scholar]
- 37. Bates D, Maechler M, Bolker B (2011) lme4: Linear mixed-effects models using S4 classes. R package version 0.999375–42. [Google Scholar]
- 38. Dansgaard W (1964) Stable isotopes in precipitation. Tellus 16: 436–468. 10.1111/j.2153-3490.1964.tb00181.x [DOI] [Google Scholar]
- 39. Clark ID, Fritz P (1997) Environmental isotopes in hydrology (1st edition): CRC Press. [Google Scholar]
- 40. Volaire F, Barkaoui K, Norton M (2014) Designing resilient and sustainable grasslands for a drier future: Adaptive strategies, functional traits and biotic interactions. European Journal of Agronomy 52, Part B: 81–89. 10.1016/j.eja.2013.10.002 [DOI] [Google Scholar]
- 41. Fischer C, Roscher C, Jensen B, Eisenhauer N, Baade J, et al. (2014) How Do Earthworms, Soil Texture and Plant Composition Affect Infiltration along an Experimental Plant Diversity Gradient in Grassland? PLoS ONE 9: e98987 10.1371/journal.pone.0098987 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42. Van Peer L, Nijs I, Reheul D, De Cauwer B (2004) Species richness and susceptibility to heat and drought extremes in synthesized grassland ecosystems: compositional vs. physiological effects. Functional Ecology 18: 769–778. 10.1111/j.0269-8463.2004.00901.x [DOI] [Google Scholar]
- 43. Dodd MB, Lauenroth WK, Welker JM (1998) Differential water resource use by herbaceous and woody plant life-forms in a shortgrass steppe community. Oecologia 117: 504–512. 10.1007/s004420050686 [DOI] [PubMed] [Google Scholar]
- 44. Moreno-Gutiérrez C, Dawson TE, Nicolás E, Querejeta JI (2012) Isotopes reveal contrasting water use strategies among coexisting plant species in a Mediterranean ecosystem. New Phytologist 196: 489–496. 10.1111/j.1469-8137.2012.04276.x [DOI] [PubMed] [Google Scholar]
- 45. Casper BB, Jackson RB (1997) Plant competition underground. Annual Review of Ecology and Systematics 28: 545–570. 10.1146/annurev.ecolsys.28.1.545 [DOI] [Google Scholar]
- 46. Bertness MD, Callaway R (1994) Positive interactions in communities. Trends in Ecology & Evolution 9: 191–193. 10.1016/0169-5347(94)90088-4 [DOI] [PubMed] [Google Scholar]
- 47. Oelmann Y, Buchmann N, Gleixner G, Habekost M, Roscher C, et al. (2011) Plant diversity effects on aboveground and belowground N pools in temperate grassland ecosystems: Development in the first 5 years after establishment. Global Biogeochemical Cycles 25: GB2014. [Google Scholar]
- 48. Bessler H, Oelmann Y, Roscher C, Buchmann N, Scherer-Lorenzen M, et al. (2012) Nitrogen uptake by grassland communities: contribution of N2 fixation, facilitation, complementarity, and species dominance. Plant and Soil 358: 301–322. 10.1007/s11104-012-1181-z [DOI] [Google Scholar]
- 49. Jobbágy E, Jackson R (2001) The distribution of soil nutrients with depth: Global patterns and the imprint of plants. Biogeochemistry 53: 51–77. 10.1023/A:1010760720215 [DOI] [Google Scholar]
- 50. Schenk HJ, Jackson RB (2002) The global biogeography of roots. Ecological Monographs 72: 311–328. 10.1890/0012-9615(2002)072[0311:TGBOR]2.0.CO;2 [DOI] [Google Scholar]
- 51. Ravenek JM, Bessler H, Engels C, Scherer-Lorenzen M, Gessler A, et al. (2014) Long-term study of root biomass in a biodiversity experiment reveals shifts in diversity effects over time. Oikos 10.1111/oik.01502. [DOI] [Google Scholar]
- 52. Kahmen A, Renker C, Unsicker SB, Buchmann N (2006) Niche complementarity for nitrogen: an explanation for the biodiversity and ecosystem functioning relationship? Ecology 87: 1244–1255. 10.1890/0012-9658(2006)87[1244:NCFNAE]2.0.CO;2 [DOI] [PubMed] [Google Scholar]
- 53. von Felten S, Hector A, Buchmann N, Niklaus PA, Schmid B, et al. (2009) Belowground nitrogen partitioning in experimental grassland plant communities of varying species richness. Ecology 90: 1389–1399. 10.1890/08-0802.1 [DOI] [PubMed] [Google Scholar]
- 54. Hooper DU, Vitousek PM (1997) The effects of plant composition and diversity on ecosystem processes. Science 277: 1302–1305. 10.1126/science.277.5330.1302 [DOI] [Google Scholar]
- 55. Tilman D, Knops J, Wedin D, Reich P, Ritchie M, et al. (1997) The influence of functional diversity and composition on ecosystem processes. Science 277: 1300–1302. 10.1126/science.277.5330.1300 [DOI] [Google Scholar]
- 56. Gubsch M, Roscher C, Gleixner G, Habekost M, Lipowsky A, et al. (2011) Foliar and soil δ15N values reveal increased nitrogen partitioning among species in diverse grassland communities. Plant, Cell & Environment 34: 895–908. 10.1111/j.1365-3040.2011.02287.x [DOI] [PubMed] [Google Scholar]
- 57. Roscher C, Schumacher J, Gubsch M, Lipowsky A, Weigelt A, et al. (2012) Using plant functional traits to explain diversity–productivity relationships. PLoS ONE 7: e36760 10.1371/journal.pone.0036760 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58. Harpole WS, Tilman D (2007) Grassland species loss resulting from reduced niche dimension. Nature 446: 791–793. 10.1038/nature05684 [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
(DOCX)
(XLSX)
Data Availability Statement
Data are available from the ETH Grassland Sciences Data Archive and can be obtained via data@usys.ethz.ch.