

Abstract
Cannabinoids elicit complex hemodynamic responses in experimental animals that involve both peripheral and central sites. Centrally administered cannabinoids have been shown to predominantly cause pressor response. However, very little is known about the mechanism of the cannabinoid receptor 1 (CB1R)-centrally evoked pressor response. In this review, we provided an overview of the contemporary knowledge regarding the cannabinoids centrally elicited cardiovascular responses and the possible underlying signaling mechanisms. The current review focuses on the rostral ventrolateral medulla (RVLM) as the primary brainstem nucleus implicated in CB1R-evoked pressor response.
Keywords: Cannabinoids, ERK1/2, nNOS, PI3K, Cardiovascular, RVLM
Cannabinoids
Cannabinoids are heterogeneous group of compounds that target cannabinoid receptors: CB1 and CB2. These compounds include the naturally occurring Δ9-tetra-hydrocannabinol (Δ9-THC), isolated from the plant Cannabis sativa (marijuana), endogenous compounds known as endocannabinoids (ECs), as well as other synthetic compounds. Since at least 2000 B.C., the plant Cannabis has been long used for recreational and medical purposes. Δ9-THC, Cannabidiol (CBD), and cannabinol are the most abundant natural cannabinoids active at CB1 and CB2 receptors, but only Δ9-THC has an equal affinity for both CB1 and CB2 receptors [1,2]. The first endogenous ligand for both cannabinoid receptors [2], anandamide, is a derivative of arachidonic acid (arachidonoyl ethanolamide; AEA), which was isolated from pig brain in 1992 [3], and 2-arachidonoyl glycerol (2-AG) is another abundant ECs [4]. Most of the endogenous cannabinoids discovered so far are agonists except the inverse agonist virodhamine [5]. The high affinity non-eicosanoid cannabinoids CP55940 and the amino-alkyl-indole cannabinoid WIN55,212-2 were developed by Pfizer and Sterling Winthrop, respectively. SR141716A and AM251 are selective antagonists for the CB1R, while SR144528 is selective for the CB2R [2,6]. Notably, most of the synthetic compounds are highly lipophilic and water insoluble except for O-1057, which is highly water soluble and possesses comparable potency as CP55940 [7]. Hemopressin, a short peptide identified in rat brain, has been recently categorized as inverse cannabinoid agonist [8,9].
Cannabinoid receptor 1
It is now known that cannabinoids exert their actions mainly via two subtypes of G-protein-coupled receptors (GPCRs): CB1 and CB2. Additional non-CB1, non-CB2 established GPCRs, such as GPR55 and GPR18, are also targeted by these compounds (e.g. anandamide, virodhamine, CP559440, and AM251 but not WIN55,212-2) [10–14]. Our review focuses on the CB1R, which is found primarily in the CNS, including the cardiovascular regulatory nuclei in the brainstem. The CB1 receptor, a 473-amino-acid protein, was first cloned from a rat cerebral cortex cDNA library [15] and a human brainstem library [16], which maintains the essential topographical features for a G-protein-coupled receptor (GPCR) of (i) seven hydrophobic transmembrane domain regions that extend through the plasma membrane; (ii) three extracellular loops; (iii) three intracellular loops; (iv) an extracellular N-terminal; (v) and an intracellular C-terminal [17].
CB1R signaling
Activation of CB1R triggers several downstream effectors including inhibition of adenylyl cyclase, stimulation of inwardly rectifying potassium channels, inhibition of N- and P/Q-type voltage-dependent calcium channels, and activation of mitogen-activated protein kinase (MAPK) pathway. Cannabinoids acting via CB1R reduce cAMP production by inhibiting adenylyl cyclase [18–20] which is antagonized by cannabinoid antagonists SR141716A and LY320135 [21]. These effects are mediated via inhibitory G-protein (Gαi/o) because they were blocked by Gαi/o-selective pertussis toxin in mammalian brain and in cultured neuronal cells [18–20]. Many other CB1R-mediated physiological functions are G-protein Gαi/o mediated [19,22,23]. However, the diverse, sometimes opposing, CB1R-evoked physiological functions that are not completely attributable to simply lowering intracellular cAMP levels, have led to investigations of the role of other non-Gαi/o signaling mechanisms [24]. In this line, recent studies have linked CB1R coupling to activation of Gαq/11 or Gαs. It is possible that heterodimerization between the CB1R and other receptor(s) contribute, at least partly, to this divergent signal transduction. This notion is supported by the reported interaction between CB1R and other co-localized receptors e.g. dopamine D2R, which resulted in accumulation of cAMP [25,26]. Second, CB1R behaves as a Gαq/11-G-protein-coupled receptor in cultured hippocampal neurons and trabecular meshwork cells [24,27]. Further, the findings that heterodimerization between CB1R and OX1R resulted in enhanced Gαq/11-dependent OX1R signaling in presence of CB1R [28].
Retrograde CB1R-mediated signaling
CB1R is located mostly presynaptically, thus playing crucial roles in controlling the release of neurotransmitters at both excitatory and inhibitory synapses. Upon depolarization, the postsynaptically released endocannabinoids activate presynaptic CB1R, which in turn modulates the release of various neurotransmitters [23,29]. For example, WIN55,212-2 inhibited GABA release from presynaptic terminals in cultured hippocampal or ventromedial medulla (RVM) neurons following postsynaptic depolarization [30,31]. The latter effect was completely abolished in presence of selective CB1 receptor antagonists. This phenomenon is termed depolarization-induced suppression of inhibition (DSI). Findings from cerebellar Purkinje cells support the possibility that postsynaptically released endocannabinoids act as retrograde secondary messengers at both inhibitory as well as excitatory synapses because following depolarization, the released endocannabinoids, which stimulate presynaptic CB1R, ultimately suppress presynaptic calcium-induced glutamate release [32]. The latter phenomenon is termed depolarization-induced suppression of excitation or (DSE). Both CB1R mediated DSE and DSI are considered key mechanisms for many of the central effects of endogenous and exogenous cannabinoids.
Cardiovascular effects of cannabinoids
The cardiovascular responses to cannabinoids are complex and are dependent on the state of the studied animals (conscious vs. anaesthetized) and the route of administration (systemic vs. central) [33–38].
Systemic CB1R-evoked cardiovascular effects
In anesthetized animals, systemically administered cannabinoids elicit predominantly hypotension and bradycardia. These effects are mediated peripherally through prejunctional inhibition of sympathetic outflow and vagal stimulation resulting in reduction in BP and HR, respectively [39–42]. Systemic administration of THC, anandamide, or WIN55,212-2 elicited tri-phasic effects on BP in anesthetized rats: (i) an initial brief hypotensive phase, secondary to a bradycardic response, which was blocked by atropine pretreatment or vagotomy; (ii) a transient pressor response due to direct vasoconstriction; (iii) a more predominant depressor phase. The prolonged depressor phase was mediated via peripheral sympathoinhibition because it was attenuated by cervical spinal transection and blockade of α-adrenoceptors [39–43]. Interestingly, recent studies have suggested that, in addition to the direct vasoconstrictor action discussed above, the transient pressor response evoked by systemic cannabinoids in anaesthetized animals might involve central mechanisms [44,45]. However, the cardiovascular responses of systemically administered cannabinoids in conscious animals are quite different. The prolonged depressor response (phase III) is absent following systemically injected anandamide or WIN55,212-2 which, in contrast, cause predominate pressor responses along with bradycardia in conscious rats [36,37]. The elicited pressor response by systemic WIN55,212-2 in conscious animals is centrally mediated because it was attenuated by ganglion blockade [37]. Importantly, in humans, acute administration of cannabinoid is associated with tachycardia and a pressor response [46–48].
Central CB1R-evoked cardiovascular effects
Centrally administered cannabinoids predominantly elicit sympathoexcitation/pressor responses. Studies have elucidated the involvement of various brainstem nuclei in the cardiovascular responses elicited by central CB1R activation, e.g. Nucleus Tractus Solitarii (NTS) and the rostral ventrolateral medulla (RVLM) [39,49–52].
The NTS
The NTS is located in the brainstem flanked on each side of the fourth ventricle and consists of groups of cells in a column-like structure dorsal to the RVLM and represents the first relay station in the baroreflex arc. Upon stimulation, the NTS elicits a reduction in the BP, HR, and sympathetic outflow [53,54]. The most cardiovascular-relevant part of the NTS is located at the most caudal part of the NTS, which contains synapses from chemo and aortic baroreceptor processes that contact with secondary order neurons within the NTS [55,56]. The latter communicate either directly or indirectly through third order neurons with other nuclei including RVLM, hypothalamus or CVLM [57–60]. Functionally, activation of cardiovascular afferents (chemo or baroreceptors) enhances the release of excitatory amino-acid L-glutamate within the NTS [54], which prompts the excitation of NTS-projections to other baroreflex arc nuclei e.g. RVLM and CVLM. Several reports have shown important roles for activation of CB1R in the NTS in blood pressure regulation [50–52,61]. For examples, activation of NTS cannabinoid receptors by anandamide enhanced baroreflex-mediated sympathoinhibition, at least partly, via presynaptic inhibition of GABA release [52,62].
The RVLM
In this review, attention has been focused on the RVLM, which plays pivotal role in central control of cardiovascular function [63–65]. The RVLM is the final supraspinal site within the central nervous system that integrates multitudes of influences on blood pressure (BP) from higher brain regions such as paraventricular nucleus, lateral hypothalamus, and periaqueductal gray [64,66]. The RVLM is of high significance in controlling BP since bilateral lesioning of the RVLM leads to a profound fall in BP [59]. The RVLM is located in the ventral part of the brainstem, lateral to the inferior olive, caudal to the facial nucleus, and ventral to the nucleus ambiguous [59,67]. It is heterogeneous in composition and contains multiple cell groups that are different in their neurochemical phenotype (e.g. rostroventrolateralis, gigantocellular nucleus, and paragigantocellularis lateralis [68–71]. Within the RVLM, the adrenergic group C1 neurons, alternatively known as adrenergic neurons, are defined based on their expression of phenylethanolamine-n-methyltransferase (PNMT) [72,73]. The rostral C1 subgroup contains barosensitive neurons which project to the spinal cord [74,75] and provides tonic excitatory inputs to the sympathetic preganglionic neurons [76,77]. Beside catecholamine-containing neurons in RVLM [78], a wide variety of neurotransmitters and receptors are present in the RVLM including substance P [79], neuropeptide Y [80], enkephalin [80,81], adenosine receptors (A2A) [82], P2X receptors [83], Angiotensin II AT1 receptors [84], imidazoline I1 receptors [85,86], α2A adrenergic receptors [87,88], cannabinoid CB1 receptors [89,90], CB2 receptors [91], and mu-opioid receptors [92,93]. The RVLM is a crucial brainstem nucleus for the tonic generation of sympathetic nerve activity [59,60]. Activation of specific neurons within the RVLM causes an increase in BP by increasing peripheral resistance and cardiac output via released catecholamines [94–97]. In addition to cardiovascular control, specific neurons within the RVLM are involved in nociception [98,99] and breathing [100]. Intracisternal (i.c) administration [101–103] or intra-RVLM microinjection [90,104] of cannabinoids such as WIN55,212-2 or CP-55940 elicited a pressor response and caused increases in sympathetic nerve activity, plasma norepinephrine and blood pressure, in conscious and anesthetized animals, and these responses were attenuated by pretreatment with the CB1R antagonists SR171416A or AM251. The significant increase in tyrosine hydroxylase immunoreactive neurons (TH-ir) expressing c-Fos, a marker of neuronal activity, following i.c. WIN55,212-2 provided direct in vivo evidence that central CB1R-evoked pressor response involves activation of RVLM-catecholaminergic neurons [102], which was abrogated by CB1R antagonist AM251.
Centrally elicited hemodynamic effects of CB1R in conscious Sprague Dawley rats
In our recent studies, we sought to elucidate the mechanisms implicated in the central CB1R-evoked sympathoexcitation/pressor response [102,104,105]. In pursuit of this goal, we characterized the centrally mediated cardiovascular effects of central CB1R activation in conscious Sprague Dawley rats. We have confirmed the expression of CB1R (protein) in the RVLM by detecting the two bands at 64 and 53 kDa, which represent the N-glycosylated and non-glycosylated forms of CB1R, respectively (unpublished data) [106].
We reported that i.c. administration of WIN55,212-2 elicited dose-dependent pressor responses and increased NE plasma levels, denoting an increase in central sympathetic tone in conscious rats [102], which agrees with findings in experimental animals discussed above [39,101,103], and reflects similar responses observed in humans [47,48]. Similar pressor response was observed following microinjection of WIN55,212-2, for the first time, in the RVLM of conscious freely moving rats [104]. These studies were conducted in conscious rats to circumvent the negative impact of anesthesia that was shown to dramatically compromise cannabinoid-evoked hemodynamic responses [36–38].
We demonstrated in our studies that the cardiovascular, biochemical, and molecular responses elicited by WIN55,212-2 were CB1R mediated. This is important because (i) WIN55,212-2, which is routinely used in cannabinoid research, can also bind to CB2R [107,108]; (ii) both CBR subtypes are expressed in the brain [89,109], including the brainstem [90]. The ability of the selective CB1R antagonist AM251 [39,101,103] to virtually abolish the pressor, biochemical and neurochemical responses elicited by i.c. WIN55,212-2 clearly implicates the CB1R in the observed responses. It is important to note, however, that the lack of change in blood pressure, as well as other neurochemical responses, following AM251 administration argues against the involvement of central CB1R signaling in tonic control of blood pressure in conscious rats [102,104,105].
Signaling mechanisms involved in CB1R-evoked pressor response in the RVLM
Role of ERK1/2-PI3K/Akt signaling pathway
Cannabinoids are highly potent activators of extracellular-signal regulated kinase 1/2 (ERK1/2), which was evident in stably transfected Chinese hamster ovary cells expressing human CB1R. This effect was (i) abrogated by SR141716A; (ii) sensitive to pertussis toxin; (iii) and independent of the cannabinoid-induced inhibition of cAMP production [110]. The pivotal role of PI3K/Akt and ERK1/2 as potential downstream molecular mediators of the central CB1R-mediated sympathoexcitation/pressor response as suggested by multiple lines of evidence was demonstrated recently [105]. Central administration of WIN55,212-2 (i.c.) significantly elevated pERK1/2 in the NTS and RVLM [105]. The involvement of any CB2R role in these responses was precluded because of the abrogation of the WIN55,212-2-mediated cardiovascular and neurochemical responses by MEK-ERK1/2 inhibition (PD98059) and attenuation of the concomitant activation of ERK1/2 pathway by pretreatment with the selective CB1R antagonist AM251 (i.c.). In view of the crucial role of brainstem pERK1/2 signaling in central control of blood pressure, previous studies from our laboratory [82,86] and others [111–113] suggest that brainstem ERK1/2 plays a bi-directional role in central regulation of blood pressure. For example, in both normotensive and hypertensive rats, inhibition of RVLM ERK1/2 phosphorylation gradually lowered blood pressure [111], and its rapid activation plays pivotal role in the angiotensin II-mediated pressor response [113,114]. In contrast, we have previously shown that RVLM MEK-ERK1/2 signaling activation underlies the central α2A adrenergic or imidazoline evoked acute hypotensive response [82,86].
Studies on the neuroprotective and/or anti-oncogenic effects of cannabinoids via PI3K/Akt signaling pathway have yielded controversial results. First, intraperitoneal injection of Δ9-THC activated PI3K/Akt pathway in mouse hippocampus, striatum, and cerebellum via a mechanism that was ERK1/2-independent [115]. Second, THC-mediated anti-cancer effect in human prostate cells involved PI3K/AKT and ERK1/2 signaling pathway activation [116]. On the other hand, it was demonstrated in multiple cancer cell lines that CB1R activation down regulates both PI3K/Akt and ERK1/2 signaling pathway [117,118]. Based on the molecular findings from our studies, we concluded that the effect of WIN55,212-2 on PI3K/Akt may contribute to the enhancement of ERK1/2 phosphorylation because in the presence of the PI3K/Akt inhibitor wortmannin, WIN55,212-2-induced ERK1/2 phosphorylation was exacerbated [105]. Additionally, PD98059, MEK-ERK1/2 inhibitor, alone or in the presence of WIN55,212-2 had no effect on brainstem pAkt phosphorylation levels.
Consistent with a diverse physiological role of PI3K/Akt-ERK1/2 pathway, we showed that a dose-related reduction in pAkt phosphorylation levels in the NTS and RVLM contributes to the i.c. WIN55,212-2-evoked pressor response [105]. In support of this conclusion are the findings that the inhibition of Akt phosphorylation in the NTS and RVLM preceded the peak WIN55,212-2-evoked pressor response (5 min). Our Western blot findings are consistent with reported findings that CB1R activation resulted in down-regulation of the PI3K/Akt signaling [105,117,118]. However, others have shown that CB1R activation up-regulated PI3K/Akt signaling in U373 MG human astrocytoma cells [119], hippocampal slices [120], and in vivo [115]. Nonetheless, further support for a causal role for the observed inhibition in Akt phosphorylation in the brainstem in the central CB1R-mediated pressor response are the findings that pharmacological inhibition of brainstem PI3K-Akt signaling (wortmannin) significantly enhanced the WIN55,212-2 evoked dose-related pressor response [105]. Interestingly, the latter study reported an increase in Akt phosphorylation elicited by WIN55,212-2 following CB1R blockade with AM251 in the NTS but not in the RVLM. This finding clearly highlights differences between neurochemical responses elicited by CB1R activation in the RVLM vs. NTS.
CB1R enhances RVLM nNOS-NO signaling pathway
The well-documented role of NOS-NO signaling in the RVLM regulation of autonomic function has led us to investigate whether nNOS-NO plays a significant role in the central CB1R-mediated pressor response [104,121–123]. We reported that intra-RVLM WIN55,212-2 microinjection elicited dose-dependent increases in real-time RVLM NO and blood pressure; NO was measured by in vivo electrochemistry and is possibly nNOS-generated because: (i) parallel to the WIN55,212-2 dose-dependent enhancement of NO release, we detected a significant increase in nNOS phosphorylation in the WIN55,212-2-treated RVLM compared to the contra-lateral side (control); (ii) i.c. WIN55,212-2 increased the number of nNOS-ir neurons expressing c-Fos, denoting an increase in the activity of nNOS expressing neurons; (iii) these neurochemical responses were abolished following selective CB1R blockade (AM251) or prior inhibition of nNOS phosphorylation (NPLA) [104]; (iv) only RVLM nNOS, but not eNOS or iNOS, derived NO is implicated in centrally evoked hypertension [123]. Because ERK1/2 dependent phosphorylation of RVLM nNOS is implicated in sympathoexcitation [124–126], the interesting possibility exists that CB1R-mediated nNOS activation might be downstream to MEK-ERK1/2 activation, which ultimately results in CB1R-mediated pressor response.
CB1R downregulates brainstem GABAergic transmission
It is highly likely that central CB1R-elicited sympathoexcitation is mediated via indirect modulation of presympathetic neurons in the brainstem whose activity is regulated by an array of tonic excitatory and inhibitory inputs [90,127]. Notably, CB1R modulates synaptic transmission of both inhibitory (GABA) and excitatory (glutamate) neurotransmitters [23,29,128,129]. Interestingly, stimulation of central GABAA receptors (muscimol) caused the following: (i) abolished the CB1R-evoked pressor response and the elevation in plasma NE; (ii) attenuated the WIN55,212-2 evoked increase in the activity (c-Fos) of catecholamine (TH-ir) [102]. These findings are consistent with reported in vitro findings that demonstrated CB1R-evoked inhibition of GABAergic transmission in cultured rostral ventromedial medulla (RVM) neurons [31]. Yet, in the NTS, studies have demonstrated a controversial role for CB1R-mediated presynaptic modulation of excitatory (glutamate) and inhibitory (GABA) neurotransmitters. Anandamide increased baroreflex-mediated sympathoinhibition in the NTS, presumably, via presynaptic inhibition of GABA release because the response was reversed in presence of the GABAAR antagonist [52].
Conclusions
As summarized in Fig. 1, the present review highlights the molecular mechanisms implicated in the predominant sympathoexcitatory effect of brainstem CB1R activation in conscious rats. CB1R stimulation enhanced neuronal activity of presympathetic neurons in the RVLM (c-Fos/TH-ir ratio). Furthermore, PI3K/Akt-ERK1/2 signaling in the brainstem seems to differentially contribute, at least in part, to the sympathoexcitatory responses elicited by the central CB1R activation in conscious rats. The discussed studies demonstrated that CB1R activation in the RVLM elicits down-regulation of PI3K/Akt pathway along with the pressor response, which was supported by the exacerbation of WIN55,212-2 evoked hemodynamic responses when PI3K/Akt was inhibited by wortmannin. By contrast, the CB1R-evoked sympathoexcitation was associated with enhanced ERK1/2 activity in the brainstem. Further, suppressing ERK1/2 signaling abolished the central CB1R-evoked pressor response. Finally, CB1R activation in the RVLM enhanced neuronal nitroxidergic activity (nNOS-NO) essential for the central regulation of cardiovascular function. These latter neuronal responses may be linked to the modulation of brainstem GABAergic neurotransmission and subsequently to the central CB1R-evoked sympathoexcitatory and pressor response. It is imperative to note that this overview highlights important signaling networks implicated in the modulation of blood pressure caused by central CB1R activation in normotensive rats. The neurochemical and molecular responses discussed above might be different under pathophysiological conditions and might, therefore, lead to different cardiovascular outcomes. Therefore, future studies on the role of central CB1R signaling in animal models of human diseases are warranted.
Fig. 1.
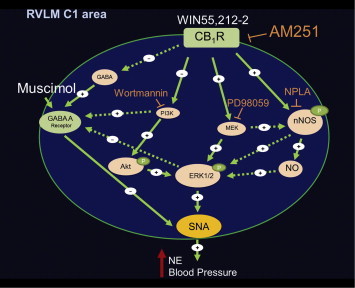
Schematic presentation of signaling mechanisms in the rostral ventrolateral medulla (RVLM) catecholaminergic C1 area, underlying central CB1R-mediated pressor response. In conscious freely moving rats, central CB1R activation (WIN55,212-2) increases blood pressure, plasma norepinephrine (NE), sympathetic neuronal activity (SNA) [102], enhances ERK1/2 and nNOS phosphorylation (NO production) and reduces Akt phosphorylation in the RVLM [104,105]. AM251 (CB1R antagonist); NPLA (nNOS inhibitor); PD98059 (MEK-ERK1/2 inhibitor) or muscimol (GABAA receptor agonist) attenuated CB1R (WIN55,212-2)-evoked pressor response [102,104,105]. In contrast, wortmannin (PI3K-Akt inhibitor) exaggerated WIN55,212-2 response [105]. The proposed model system is further supported by our neurochemical and pharmacological findings following intracisternal or intra-RVLM microinjection of the CB1R agonist WIN55,212-2 [104,105]. Solid arrows indicate signaling based on reported in vivo findings, while dashed arrows indicate proposed signaling based on reported in vitro findings, but not tested in this model (see text for details).
Conflict of interest
The authors have declared no conflict of interest.
Acknowledgments
The studies conducted in the authors’ lab were supported, in part, by NIH Grant 2R01 AA07839-19 (Abdel A. Abdel-Rahman), and by predoctoral fellowship provided by the Egyptian Cultural and Educational Bureau (Badr M. Ibrahim).
Biographies

Dr. Badr Ibrahim is an assistant professor of Pharmacology and Toxicology at El-Minia Faculty of Pharmacy, Egypt. He has completed his postdoctoral training and earned his PhD from the Department of Pharmacology and Toxicology, at East Carolina University, North Carolina, USA. Dr. Ibrahim’s research focused on elucidating the cellular and molecular mechanisms underlying the central cannabinoid receptor-mediated hypertensive effect in unrestrained conscious rats. The numerous national and international awards Dr. Ibrahim had received, as a graduate student, lend further support to his contributions to the field of cardiovascular neuropharmacology.

Dr. Abdel-Rahman is Distinguished Professor of Pharmacology and Vice Chair of the Department of Pharmacology and Toxicology, Brody School of Medicine at East Carolina University, Greenville, NC, USA. He published over 120 refereed scientific papers in addition to 10 education-related articles. His research findings have been published in top journals in his discipline and cited hundreds of times in scientific literature. Dr. Abdel-Rahman research deals with neural control of circulation and neurobiology of hypertension. Two National Institutes of Health grants fund his research. In the first project his research team investigates the effect of ethanol on neuronal pathways that control blood pressure and cardiac reflexes. The second project deals with the neuroprotective and cardioprotective actions of estrogen and how concurrent alcohol use might compromise these beneficial physiological effects of estrogen.
In addition to his contributions to research, Dr. Abdel-Rahman has been active as a member of many scientific societies for the past 30 years and has been named a Fellow of the American Heart Association. Dr. Abdel-Rahman also served as President of the East Carolina University Neuroscience Chapter in addition to his services editor/associate editor and reviewer for a number of scientific journals. He has also served as a member of review boards (study sections) of the National Institutes of Health and the American Heart Association.
Footnotes
Peer review under responsibility of Cairo University.

References
- 1.Munro S., Thomas K.L., Abu-Shaar M. Molecular characterization of a peripheral receptor for cannabinoids. Nature. 1993;365(6441):61–65. doi: 10.1038/365061a0. [DOI] [PubMed] [Google Scholar]
- 2.Felder C.C., Joyce K.E., Briley E.M., Mansouri J., Mackie K., Blond O. Comparison of the pharmacology and signal transduction of the human cannabinoid CB1 and CB2 receptors. Mol Pharmacol. 1995;48(3):443–450. [PubMed] [Google Scholar]
- 3.Devane W., Hanus L., Breuer A., Pertwee R., Stevenson L., Griffin G. Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science. 1992;258(5090):1946–1949. doi: 10.1126/science.1470919. [DOI] [PubMed] [Google Scholar]
- 4.Sugiura T., Kodaka T., Nakane S., Miyashita T., Kondo S., Suhara Y. Evidence that the cannabinoid CB1 receptor is a 2-arachidonoylglycerol receptor. J Biol Chem. 1999;274(5):2794–2801. doi: 10.1074/jbc.274.5.2794. [DOI] [PubMed] [Google Scholar]
- 5.Porter A.C., Sauer J.-M., Knierman M.D., Becker G.W., Berna M.J., Bao J. Characterization of a novel endocannabinoid, virodhamine, with antagonist activity at the CB1 receptor. J Pharmacol Exp Ther. 2002;301(3):1020–1024. doi: 10.1124/jpet.301.3.1020. [DOI] [PubMed] [Google Scholar]
- 6.Rinaldi-Carmona M., Barth F., Millan J., Derocq J.-M., Casellas P., Congy C. SR 144528, the first potent and selective antagonist of the CB2 cannabinoid receptor. J Pharmacol Exp Ther. 1998;284(2):644–650. [PubMed] [Google Scholar]
- 7.Pertwee R.G., Gibson T.M., Stevenson L.A., Ross R.A., Banner W.K., Saha B. O-1057, a potent water-soluble cannabinoid receptor agonist with antinociceptive properties. Br J Pharmacol. 2000;129(8):1577–1584. doi: 10.1038/sj.bjp.0703245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Dodd G.T., Mancini G., Lutz B., Luckman S.M. The peptide hemopressin acts through CB1 cannabinoid receptors to reduce food intake in rats and mice. J Neurosci. 2010;30(21):7369–7376. doi: 10.1523/JNEUROSCI.5455-09.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Heimann A.S., Gomes I., Dale C.S., Pagano R.L., Gupta A., de Souza L.L. Hemopressin is an inverse agonist of CB1 cannabinoid receptors. Proc Nat Acad Sci. 2007;104(51):20588–20593. doi: 10.1073/pnas.0706980105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Lauckner J.E., Jensen J.B., Chen H.Y., Lu H.C., Hille B., Mackie K. GPR55 is a cannabinoid receptor that increases intracellular calcium and inhibits M current. Proc Nat Acad Sci USA. 2008;105(7):2699–2704. doi: 10.1073/pnas.0711278105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Jarai Z., Wagner J.A., Varga K., Lake K.D., Compton D.R., Martin B.R. Cannabinoid-induced mesenteric vasodilation through an endothelial site distinct from CB1 or CB2 receptors. Proc Nat Acad Sci USA. 1999;96(24):14136–14141. doi: 10.1073/pnas.96.24.14136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kapur A., Zhao P.W., Sharir H., Bai Y.S., Caron M.G., Barak L.S. Atypical responsiveness of the orphan receptor GPR55 to cannabinoid ligands. J Biol Chem. 2009;284(43):29817–29827. doi: 10.1074/jbc.M109.050187. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Ryberg E., Larsson N., Sjogren S., Hjorth S., Hermansson N.O., Leonova J. The orphan receptor GPR55 is a novel cannabinoid receptor. Br J Pharmacol. 2007;152(7):1092–1101. doi: 10.1038/sj.bjp.0707460. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Waldeck-Weiermair M., Zoratti C., Osibow K., Balenga N., Goessnitzer E., Waldhoer M. Integrin clustering enables anandamide-induced Ca(2+) signaling in endothelial cells via GPR55 by protection against CB1-receptor-triggered repression. J Cell Sci. 2008;121(10):1704–1717. doi: 10.1242/jcs.020958. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Matsuda L.A., Lolait S.J., Brownstein M.J., Young A.C., Bonner T.I. Structure of a cannabinoid receptor and functional expression of the cloned cDNA. Nature. 1990;346(6284):561–564. doi: 10.1038/346561a0. [DOI] [PubMed] [Google Scholar]
- 16.Gerard C.M., Mollereau C., Vassart G., Parmentier M. Molecular cloning of a human cannabinoid receptor which is also expressed in testis. Biochem J. 1991;279(Pt 1):129–134. doi: 10.1042/bj2790129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Howlett A.C. The CB1 cannabinoid receptor in the brain. Neurobiol Dis. 1998;5(6):405–416. doi: 10.1006/nbdi.1998.0215. [DOI] [PubMed] [Google Scholar]
- 18.Bidaut-Russell M., Devane W.A., Howlett A.C. Cannabinoid receptors and modulation of cyclic AMP accumulation in the rat brain. J Neurochem. 1990;55(1):21–26. doi: 10.1111/j.1471-4159.1990.tb08815.x. [DOI] [PubMed] [Google Scholar]
- 19.Howlett A.C., Qualy J.M., Khachatrian L.L. Involvement of Gi in the inhibition of adenylate cyclase by cannabimimetic drugs. Mol Pharmacol. 1986;29(3):307–313. [PubMed] [Google Scholar]
- 20.Childers S.R., Fleming L., Konkoy C., Marckel D., Pacheco M., Sexton T. Opioid and cannabinoid receptor inhibition of adenylyl cyclase in braina. Ann NY Acad Sci. 1992;654(1):33–51. doi: 10.1111/j.1749-6632.1992.tb25954.x. [DOI] [PubMed] [Google Scholar]
- 21.Felder C.C., Joyce K.E., Briley E.M., Glass M., Mackie K.P., Fahey K.J. LY320135, a novel cannabinoid CB1 receptor antagonist, unmasks coupling of the CB1 receptor to stimulation of cAMP accumulation. J Pharmacol Exp Ther. 1998;284(1):291–297. [PubMed] [Google Scholar]
- 22.Howlett A.C., Mukhopadhyay S. Cellular signal transduction by anandamide and 2-arachidonoylglycerol. Chem Phys Lipids. 2000;108(1–2):53–70. doi: 10.1016/s0009-3084(00)00187-0. [DOI] [PubMed] [Google Scholar]
- 23.Piomelli D. The molecular logic of endocannabinoid signalling. Nat Rev Neurosci. 2003;4(11):873–884. doi: 10.1038/nrn1247. [DOI] [PubMed] [Google Scholar]
- 24.Lauckner J.E., Hille B., Mackie K. The cannabinoid agonist WIN55,212-2 increases intracellular calcium via CB1 receptor coupling to Gq/11 G proteins. Proc Nat Acad Sci USA. 2005;102(52):19144–19149. doi: 10.1073/pnas.0509588102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kearn C.S., Blake-Palmer K., Daniel E., Mackie K., Glass M. Concurrent stimulation of cannabinoid CB1 and dopamine D2 receptors enhances heterodimer formation: a mechanism for receptor cross-talk? Mol Pharmacol. 2005;67(5):1697–1704. doi: 10.1124/mol.104.006882. [DOI] [PubMed] [Google Scholar]
- 26.Glass M., Felder C.C. Concurrent stimulation of cannabinoid CB1 and dopamine D2 receptors augments cAMP accumulation in striatal neurons: evidence for a Gs linkage to the CB1 receptor. J Neurosci. 1997;17(14):5327–5333. doi: 10.1523/JNEUROSCI.17-14-05327.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.McIntosh B.T., Hudson B., Yegorova S., Jollimore C.A.B., Kelly M.E.M. Agonist-dependent cannabinoid receptor signalling in human trabecular meshwork cells. Br J Pharmacol. 2007;152(7):1111–1120. doi: 10.1038/sj.bjp.0707495. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Ellis J., Pediani J.D., Canals M., Milasta S., Milligan G. Orexin-1 receptor-cannabinoid CB1 receptor heterodimerization results in both ligand-dependent and -independent coordinated alterations of receptor localization and function. J Biol Chem. 2006;281(50):38812–38824. doi: 10.1074/jbc.M602494200. [DOI] [PubMed] [Google Scholar]
- 29.Freund T.F., Katona I., Piomelli D. Role of endogenous cannabinoids in synaptic signaling. Physiol Rev. 2003;83(3):1017–1066. doi: 10.1152/physrev.00004.2003. [DOI] [PubMed] [Google Scholar]
- 30.Ohno-Shosaku T., Maejima T., Kano M. Endogenous cannabinoids mediate retrograde signals from depolarized postsynaptic neurons to presynaptic terminals. Neuron. 2001;29(3):729–738. doi: 10.1016/s0896-6273(01)00247-1. [DOI] [PubMed] [Google Scholar]
- 31.Vaughan C.W., McGregor I.S., Christie M.J. Cannabinoid receptor activation inhibits GABAergic neurotransmission in rostral ventromedial medulla neurons in vitro. Br J Pharmacol. 1999;127(4):935–940. doi: 10.1038/sj.bjp.0702636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kreitzer A.C., Regehr W.G. Retrograde inhibition of presynaptic calcium influx by endogenous cannabinoids at excitatory synapses onto purkinje cells. Neuron. 2001;29(3):717–727. doi: 10.1016/s0896-6273(01)00246-x. [DOI] [PubMed] [Google Scholar]
- 33.Mendizabal V.E., Adler-Graschinsky E. Cannabinoids as therapeutic agents in cardiovascular disease: a tale of passions and illusions. Br J Pharmacol. 2007;151(4):427–440. doi: 10.1038/sj.bjp.0707261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Randall M.D., Kendall D.A., O’Sullivan S. The complexities of the cardiovascular actions of cannabinoids. Br J Pharmacol. 2004;142(1):20–26. doi: 10.1038/sj.bjp.0705725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Randall M.D., Harris D., Kendall D.A., Ralevic V. Cardiovascular effects of cannabinoids. Pharmacol Ther. 2002;95(2):191–202. doi: 10.1016/s0163-7258(02)00258-9. [DOI] [PubMed] [Google Scholar]
- 36.Stein E.A., Fuller S.A., Edgemond W.S., Campbell W.B. Physiological and behavioural effects of the endogenous cannabinoid, arachidonylethanolamide (anandamide), in the rat. Br J Pharmacol. 1996;119(1):107–114. doi: 10.1111/j.1476-5381.1996.tb15683.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Gardiner S.M., March J.E., Kemp P.A., Bennett T. Regional haemodynamic responses to the cannabinoid agonist, WIN 55212-2, in conscious, normotensive rats, and in hypertensive, transgenic rats. Br J Pharmacol. 2001;133(3):445–453. doi: 10.1038/sj.bjp.0704100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Lake K.D., Martin B.R., Kunos G., Varga K. Cardiovascular effects of anandamide in anesthetized and conscious normotensive and hypertensive rats. Hypertension. 1997;29(5):1204–1210. doi: 10.1161/01.hyp.29.5.1204. [DOI] [PubMed] [Google Scholar]
- 39.Niederhoffer N., Szabo B. Effect of the cannabinoid receptor agonist WIN55212-2 on sympathetic cardiovascular regulation. Br J Pharmacol. 1999;126(2):457–466. doi: 10.1038/sj.bjp.0702337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Varga K., Lake K.D., Huangfu D., Guyenet P.G., Kunos G. Mechanism of the hypotensive action of anandamide in anesthetized rats. Hypertension. 1996;28(4):682–686. doi: 10.1161/01.hyp.28.4.682. [DOI] [PubMed] [Google Scholar]
- 41.Lake K.D., Compton D.R., Varga K., Martin B.R., Kunos G. Cannabinoid-induced hypotension and bradycardia in rats mediated by CB1-like cannabinoid receptors. J Pharmacol Exp Ther. 1997;281(3):1030–1037. [PubMed] [Google Scholar]
- 42.Varga K., Lake K., Martin B.R., Kunos G. Novel antagonist implicates the CB1 cannabinoid receptor in the hypotensive action of anandamide. Eur J Pharmacol. 1995;278(3):279–283. doi: 10.1016/0014-2999(95)00181-j. [DOI] [PubMed] [Google Scholar]
- 43.Siqueira S.W., Lapa A.J., Ribeiro do Valle J. The triple effect induced by delta 9-tetrahydrocannabinol on the rat blood pressure. Eur J Pharmacol. 1979;58(4):351–357. doi: 10.1016/0014-2999(79)90305-4. [DOI] [PubMed] [Google Scholar]
- 44.Kwolek G., Zakrzeska A., Schlicker E., Gothert M., Godlewski G., Malinowska B. Central and peripheral components of the pressor effect of anandamide in urethane-anaesthetized rats.[see comment] Br J Pharmacol. 2005;145(5):567–575. doi: 10.1038/sj.bjp.0706195. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Malinowska B., Zakrzeska A., Kurz C., Göthert M., Kwolek G., Wielgat P. Involvement of central β2 adrenergic, NMDA and thromboxane A2 receptors in the pressor effect of anandamide in rats. Naunyn Schmiedebergs Arch Pharmacol. 2010;381(4):349–360. doi: 10.1007/s00210-010-0497-6. [DOI] [PubMed] [Google Scholar]
- 46.Benowitz N.L., Rosenberg J., Rogers W., Bachman J., Jones R.T. Cardiovascular effects of intravenous delta-9-tetrahydrocannabinol: autonomic nervous mechanisms. Clin Pharmacol Ther. 1979;25(4):440–446. doi: 10.1002/cpt1979254440. [DOI] [PubMed] [Google Scholar]
- 47.Foltin R.W., Fischman M.W., Pedroso J.J., Pearlson G.D. Marijuana and cocaine interactions in humans: cardiovascular consequences. Pharmacol Biochem Behav. 1987;28(4):459–464. doi: 10.1016/0091-3057(87)90506-5. [DOI] [PubMed] [Google Scholar]
- 48.Sidney S. Cardiovascular consequences of marijuana use. J Clin Pharmacol. 2002;42(90110):S64–S70. doi: 10.1002/j.1552-4604.2002.tb06005.x. [DOI] [PubMed] [Google Scholar]
- 49.Dean C. Cannabinoid and GABA modulation of sympathetic nerve activity and blood pressure in the dorsal periaqueductal gray of the rat. Am J Physiol Regul Integr Comp Physiol. 2011;301(6):R1765–72. doi: 10.1152/ajpregu.00398.2011. [DOI] [PubMed] [Google Scholar]
- 50.Seagard J.L., Hopp F.A., Hillard C.J., Dean C. Effects of endocannabinoids on discharge of baroreceptive NTS neurons. Neurosci Lett. 2005;381(3):334–339. doi: 10.1016/j.neulet.2005.02.044. [DOI] [PubMed] [Google Scholar]
- 51.Rademacher D.J., Patel S., Hopp F.A., Dean C., Hillard C.J., Seagard J.L. Microinjection of a cannabinoid receptor antagonist into the NTS increases baroreflex duration in dogs. Am J Physiol – Heart Circulat Physiol. 2003;284(5):H1570–H1576. doi: 10.1152/ajpheart.00772.2002. [DOI] [PubMed] [Google Scholar]
- 52.Seagard J.L., Dean C., Patel S., Rademacher D.J., Hopp F.A., Schmeling W.T. Anandamide content and interaction of endocannabinoid/GABA modulatory effects in the NTS on baroreflex-evoked sympathoinhibition. Am J Physiol – Heart Circulat Physiol. 2004;286(3):H992–1000. doi: 10.1152/ajpheart.00870.2003. [DOI] [PubMed] [Google Scholar]
- 53.Aicher S.A., Randich A. Antinociception and cardiovascular responses produced by electrical stimulation in the nucleus tractus solitarius, nucleus reticularis ventralis, and the caudal medulla. Pain. 1990;42(1):103–119. doi: 10.1016/0304-3959(90)91096-2. [DOI] [PubMed] [Google Scholar]
- 54.Miura M., Reis D.J. The role of the solitary and paramedian reticular nuclei in mediating cardiovascular reflex responses from carotid baro- and chemoreceptors. J Physiol. 1972;223(2):525–548. doi: 10.1113/jphysiol.1972.sp009861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Nomura S., Mizuno N. Central distribution of afferent and efferent components of the glossopharyngeal nerve: an HRP study in the cat. Brain Res. 1982;236(1):1–13. doi: 10.1016/0006-8993(82)90029-4. [DOI] [PubMed] [Google Scholar]
- 56.Seiders E.P., Stuesse S.L. A horseradish peroxidase investigation of carotid sinus nerve components in the rat. Neurosci Lett. 1984;46(1):13–18. doi: 10.1016/0304-3940(84)90191-5. [DOI] [PubMed] [Google Scholar]
- 57.Aicher S.A., Saravay R.H., Cravo S., Jeske I., Morrison S.F., Reis D.J. Monosynaptic projections from the nucleus tractus solitarii to C1 adrenergic neurons in the rostral ventrolateral medulla: comparison with input from the caudal ventrolateral medulla. J Comp Neurol. 1996;373(1):62–75. doi: 10.1002/(SICI)1096-9861(19960909)373:1<62::AID-CNE6>3.0.CO;2-B. [DOI] [PubMed] [Google Scholar]
- 58.Aicher S.A., Kurucz O.S., Reis D.J., Milner T.A. Nucleus tractus solitarius efferent terminals synapse on neurons in the caudal ventrolateral medulla that project to the rostral ventrolateral medulla. Brain Res. 1995;693(1–2):51–63. doi: 10.1016/0006-8993(95)00660-i. [DOI] [PubMed] [Google Scholar]
- 59.Dampney R.A., Czachurski J., Dembowsky K., Goodchild A.K., Seller H. Afferent connections and spinal projections of the pressor region in the rostral ventrolateral medulla of the cat. J Auton Nerv Syst. 1987;20(1):73–86. doi: 10.1016/0165-1838(87)90083-x. [DOI] [PubMed] [Google Scholar]
- 60.Ross C.A., Ruggiero D.A., Reis D.J. Projections from the nucleus tractus solitarii to the rostral ventrolateral medulla. J Comp Neurol. 1985;242(4):511–534. doi: 10.1002/cne.902420405. [DOI] [PubMed] [Google Scholar]
- 61.Van Sickle M.D., Oland L.D., Ho W., Hillard C.J., Mackie K., Davison J.S. Cannabinoids inhibit emesis through CB1 receptors in the brainstem of the ferret. Gastroenterology. 2001;121(4):767–774. doi: 10.1053/gast.2001.28466. [see comment] [DOI] [PubMed] [Google Scholar]
- 62.Chen C.-Y., Bonham A.C., Dean C., Hopp F.A., Hillard C.J., Seagard J.L. Retrograde release of endocannabinoids inhibits presynaptic GABA release to second-order baroreceptive neurons in NTS. Auton Neurosci. 2010;158(1–2):44–50. doi: 10.1016/j.autneu.2010.06.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Nassar N., Abdel-Rahman A.A. Central adenosine signaling plays a key role in centrally mediated hypotension in conscious aortic barodenervated rats. J Pharmacol Exp Ther. 2006;318(1):255–261. doi: 10.1124/jpet.105.100495. [DOI] [PubMed] [Google Scholar]
- 64.Dampney R.A., Polson J.W., Potts P.D., Hirooka Y., Horiuchi J. Functional organization of brain pathways subserving the baroreceptor reflex: studies in conscious animals using immediate early gene expression. Cell Mol Neurobiol. 2003;23(4–5):597–616. doi: 10.1023/A:1025080314925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Strack A.M., Sawyer W.B., Hughes J.H., Platt K.B., Loewy A.D. A general pattern of CNS innervation of the sympathetic outflow demonstrated by transneuronal pseudorabies viral infections. Brain Res. 1989;491(1):156–162. doi: 10.1016/0006-8993(89)90098-x. [DOI] [PubMed] [Google Scholar]
- 66.Guyenet P.G., Haselton J.R., Sun M.K. Sympathoexcitatory neurons of the rostroventrolateral medulla and the origin of the sympathetic vasomotor tone. Prog Brain Res. 1989;81:105–116. doi: 10.1016/s0079-6123(08)62002-6. [DOI] [PubMed] [Google Scholar]
- 67.Farlow D.M., Goodchild A.K., Dampney R.A. Evidence that vasomotor neurons in the rostral ventrolateral medulla project to the spinal sympathetic outflow via the dorsomedial pressor area. Brain Res. 1984;298(2):313–320. doi: 10.1016/0006-8993(84)91430-6. [DOI] [PubMed] [Google Scholar]
- 68.Andrezik J.A., Chan-Palay V., Palay S.L. The nucleus paragigantocellularis lateralis in the rat. Anat Embryol. 1981;161(4):355–371. doi: 10.1007/BF00316048. [DOI] [PubMed] [Google Scholar]
- 69.Villanueva L., de Pommery J., Menétrey D., Le Bars D. Spinal afferent projections to subnucleus reticularis dorsalis in the rat. Neurosci Lett. 1991;134(1):98–102. doi: 10.1016/0304-3940(91)90517-w. [DOI] [PubMed] [Google Scholar]
- 70.Watanabe S., Kitamura T., Watanabe L., Sato H., Yamada J. Projections from the nucleus reticularis magnocellularis to the rat cervical cord using electrical stimulation and iontophoretic injection methods. Anatom Sci Int. 2003;78(1):42–52. doi: 10.1046/j.0022-7722.2003.00038.x. [DOI] [PubMed] [Google Scholar]
- 71.Ruggiero D.A., Cravo S.L., Arango V., Reis D.J. Central control of the circulation by the rostral ventrolateral reticular nucleus: anatomical substrates. Prog Brain Res. 1989;81:49–79. doi: 10.1016/s0079-6123(08)61999-8. [DOI] [PubMed] [Google Scholar]
- 72.Ross C.A., Ruggiero D.A., Joh T.H., Park D.H., Reis D.J. Rostral ventrolateral medulla: selective projections to the thoracic autonomic cell column from the region containing C1 adrenaline neurons. J Comp Neurol. 1984;228(2):168–185. doi: 10.1002/cne.902280204. [DOI] [PubMed] [Google Scholar]
- 73.Jeske I., McKenna K.E. Quantitative analysis of bulbospinal projections from the rostral ventrolateral medulla: contribution of C1-adrenergic and nonadrenergic neurons. J Comp Neurol. 1992;324(1):1–13. doi: 10.1002/cne.903240102. [DOI] [PubMed] [Google Scholar]
- 74.Kanjhan R., Lipski J., Kruszewska B., Rong W. A comparative study of pre-sympathetic and Bötzinger neurons in the rostral ventrolateral medulla (RVLM) of the rat. Brain Res. 1995;699(1):19–32. doi: 10.1016/0006-8993(95)00814-7. [DOI] [PubMed] [Google Scholar]
- 75.Schreihofer A.M., Guyenet P.G. Identification of C1 presympathetic neurons in rat rostral ventrolateral medulla by juxtacellular labeling in vivo. J Comp Neurol. 1997;387(4):524–536. doi: 10.1002/(sici)1096-9861(19971103)387:4<524::aid-cne4>3.0.co;2-4. [DOI] [PubMed] [Google Scholar]
- 76.Guyenet P.G. The sympathetic control of blood pressure. Nat Rev Neurosci. 2006;7(5):335–346. doi: 10.1038/nrn1902. [DOI] [PubMed] [Google Scholar]
- 77.Guyenet P.G., Koshiya N., Huangfu D., Baraban S.C., Stornetta R.L., Li Y.-W. Chapter 8 role of medulla oblongata in generation of sympathetic and vagal outflows. In: Holstege R.B.G., Saper C.B., editors. Progress in brain research. Elsevier; 1996. pp. 127–144. [DOI] [PubMed] [Google Scholar]
- 78.Goodchild A.K., Phillips J.K., Lipski J., Pilowsky P.M. Differential expression of catecholamine synthetic enzymes in the caudal ventral pons. J Comp Neurol. 2001;438(4):457–467. doi: 10.1002/cne.1328. [DOI] [PubMed] [Google Scholar]
- 79.Pilowsky P., Minson J., Hodgson A., Howe P., Chalmers J. Does substance P coexist with adrenaline in neurones of the rostral ventrolateral medulla in the rat? Neurosci Lett. 1986;71(3):293–298. doi: 10.1016/0304-3940(86)90636-1. [DOI] [PubMed] [Google Scholar]
- 80.Polson J.W., Halliday G.M., McAllen R.M., Coleman M.J., Dampney R.A. Rostrocaudal differences in morphology and neurotransmitter content of cells in the subretrofacial vasomotor nucleus. J Auton Nerv Syst. 1992;38(2):117–137. doi: 10.1016/0165-1838(92)90232-6. [DOI] [PubMed] [Google Scholar]
- 81.Boone J.B., Jr., Corry J.M. Proenkephalin gene expression in the brainstem regulates post-exercise hypotension. Brain Res Mol Brain Res. 1996;42(1):31–38. doi: 10.1016/s0169-328x(96)00105-2. [DOI] [PubMed] [Google Scholar]
- 82.Nassar N., Abdel-Rahman A.A. Brainstem phosphorylated extracellular signal-regulated kinase 1/2-nitric-oxide synthase signaling mediates the adenosine A2A-dependent hypotensive action of clonidine in conscious aortic barodenervated rats. J Pharmacol Exp Ther. 2008;324(1):79–85. doi: 10.1124/jpet.107.129692. [DOI] [PubMed] [Google Scholar]
- 83.Ralevic V. P2 receptors in the central and peripheral nervous systems modulating sympathetic vasomotor tone. J Auton Nerv Syst. 2000;81(1–3):205–211. doi: 10.1016/s0165-1838(00)00139-9. [DOI] [PubMed] [Google Scholar]
- 84.Potts P.D., Allen A.M., Horiuchi J., Dampney R.A. Does angiotensin II have a significant tonic action on cardiovascular neurons in the rostral and caudal VLM? Am J Physiol – Regulat Integrat Comp Physiol. 2000;279(4):R1392–R1402. doi: 10.1152/ajpregu.2000.279.4.R1392. [DOI] [PubMed] [Google Scholar]
- 85.Zhang J., Abdel-Rahman A.A. The hypotensive action of rilmenidine is dependent on functional N-methyl-D-aspartate receptor in the rostral ventrolateral medulla of conscious spontaneously hypertensive rats. J Pharmacol Exp Ther. 2002;303(1):204–210. doi: 10.1124/jpet.102.037333. [DOI] [PubMed] [Google Scholar]
- 86.Zhang J., Abdel-Rahman A.A. Mitogen-activated protein kinase phosphorylation in the rostral ventrolateral medulla plays a key role in imidazoline (i1)-receptor-mediated hypotension. J Pharmacol Exp Ther. 2005;314(3):945–952. doi: 10.1124/jpet.105.087510. [DOI] [PubMed] [Google Scholar]
- 87.El-Mas M.M., Abdel-Rahman A.A. Differential modulation by estrogen of alpha2-adrenergic and I1-imidazoline receptor-mediated hypotension in female rats. J Appl Physiol. 2004;97(4):1237–1244. doi: 10.1152/japplphysiol.00401.2004. [DOI] [PubMed] [Google Scholar]
- 88.Li G., Wang X., Abdel-Rahman A.A. Neuronal norepinephrine responses of the rostral ventrolateral medulla and nucleus tractus solitarius neurons distinguish the I1- from the alpha2-receptor-mediated hypotension in conscious SHRs. J Cardiovasc Pharmacol. 2005;46(1):52–62. doi: 10.1097/01.fjc.0000162773.54915.52. [DOI] [PubMed] [Google Scholar]
- 89.Herkenham M., Lynn A.B., Johnson M.R., Melvin L.S., de Costa B.R., Rice K.C. Characterization and localization of cannabinoid receptors in rat brain: a quantitative in vitro autoradiographic study. J Neurosci. 1991;11(2):563–583. doi: 10.1523/JNEUROSCI.11-02-00563.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Padley J.R., Li Q., Pilowsky P.M., Goodchild A.K. Cannabinoid receptor activation in the rostral ventrolateral medulla oblongata evokes cardiorespiratory effects in anaesthetised rats. Br J Pharmacol. 2003;140(2):384–394. doi: 10.1038/sj.bjp.0705422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Van Sickle M.D., Duncan M., Kingsley P.J., Mouihate A., Urbani P., Mackie K. Identification and functional characterization of brainstem cannabinoid CB2 receptors. Science. 2005;310(5746):329–332. doi: 10.1126/science.1115740. [DOI] [PubMed] [Google Scholar]
- 92.Drake C.T., Aicher S.A., Montalmant F.L., Milner T.A. Redistribution of mu-opioid receptors in C1 adrenergic neurons following chronic administration of morphine. Exp Neurol. 2005;196(2):365–372. doi: 10.1016/j.expneurol.2005.08.012. [DOI] [PubMed] [Google Scholar]
- 93.Aicher S.A., Kraus J.A., Sharma S., Patel A., Milner T.A. Selective distribution of mu-opioid receptors in C1 adrenergic neurons and their afferents. J Comp Neurol. 2001;433(1):23–33. doi: 10.1002/cne.1122. [DOI] [PubMed] [Google Scholar]
- 94.Guertzenstein P.G., Silver A. Fall in blood pressure produced from discrete regions of the ventral surface of the medulla by glycine and lesions. J Physiol. 1974;242(2):489–503. doi: 10.1113/jphysiol.1974.sp010719. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Feldberg W., Guertzenstein P.G. Vasodepressor effects obtained by drugs acting on the ventral surface of the brain stem. J Physiol. 1976;258(2):337–355. doi: 10.1113/jphysiol.1976.sp011423. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Guertzenstein P.G. Vasodepressor and pressor responses to drugs topically applied to the ventral surface of the brain stem. J Physiol. 1972;224(2):84P–85P. [PubMed] [Google Scholar]
- 97.McAllen R.M., Dampney R.A. The selectivity of descending vasomotor control by subretrofacial neurons. Prog Brain Res. 1989;81:233–242. doi: 10.1016/s0079-6123(08)62013-0. [DOI] [PubMed] [Google Scholar]
- 98.Karlsson G.A., Preuss C.V., Chaitoff K.A., Maher T.J., Ally A. Medullary monoamines and NMDA-receptor regulation of cardiovascular responses during peripheral nociceptive stimuli. Neurosci Res. 2006;55(3):316–326. doi: 10.1016/j.neures.2006.04.002. [DOI] [PubMed] [Google Scholar]
- 99.Javanmardi K., Parviz M., Sadr S.s., Keshavarz M., Minaii B., Dehpour A.R. Involvement of N-methyl-D-aspartate receptors and nitric oxide in the rostra ventromedial medulla in modulating morphine pain-inhibitory signals from the periaqueductal grey matter in rats. Clin Exp Pharmacol Physiol. 2005;32(7):585–589. doi: 10.1111/j.1440-1681.2005.04234.x. [DOI] [PubMed] [Google Scholar]
- 100.Nattie E.E., Li A.H. Fluorescence location of RVLM kainate microinjections that alter the control of breathing. J Appl Physiol. 1990;68(3):1157–1166. doi: 10.1152/jappl.1990.68.3.1157. [DOI] [PubMed] [Google Scholar]
- 101.Niederhoffer N., Szabo B. Cannabinoids cause central sympathoexcitation and bradycardia in rabbits. J Pharmacol Exp Ther. 2000;294(2):707–713. [PubMed] [Google Scholar]
- 102.Ibrahim B.M., Abdel-Rahman A.A. Role of brainstem GABAergic signaling in central cannabinoid receptor evoked sympathoexcitation and pressor responses in conscious rats. Brain Res. 2011;1414:1–9. doi: 10.1016/j.brainres.2011.07.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Pfitzer T., Niederhoffer N., Szabo B. Central effects of the cannabinoid receptor agonist WIN55212-2 on respiratory and cardiovascular regulation in anaesthetised rats. Br J Pharmacol. 2004;142(6):943–952. doi: 10.1038/sj.bjp.0705874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Ibrahim B.M., Abdel-Rahman A.A. Enhancement of rostral ventrolateral medulla neuronal nitric-oxide synthase-nitric-oxide signaling mediates the central cannabinoid receptor 1-evoked pressor response in conscious rats. J Pharmacol Exp Ther. 2012;341(3):579–586. doi: 10.1124/jpet.112.192369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Ibrahim B.M., Abdel-Rahman A.A. Differential modulation of brainstem PI3K/Akt and ERK1/2 signaling underlies WIN55,212-2 centrally-mediated pressor response in conscious rats. J Pharmacol Exp Ther. 2012;340(1):11–18. doi: 10.1124/jpet.111.186858. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Song C., Howlett A. Rat brain cannabinoid receptors are N-linked glycosylated proteins. Life Sci. 1995;56(23-24):1983–1989. doi: 10.1016/0024-3205(95)00179-a. [DOI] [PubMed] [Google Scholar]
- 107.Griffin G., Atkinson P.J., Showalter V.M., Martin B.R., Abood M.E. Evaluation of cannabinoid receptor agonists and antagonists using the guanosine-5′-O-(3-[35S]thio)-triphosphate binding assay in rat cerebellar membranes. J Pharmacol Exp Ther. 1998;285(2):553–560. [PubMed] [Google Scholar]
- 108.Showalter V.M., Compton D.R., Martin B.R., Abood M.E. Evaluation of binding in a transfected cell line expressing a peripheral cannabinoid receptor (CB2): identification of cannabinoid receptor subtype selective ligands. J Pharmacol Exp Ther. 1996;278(3):989–999. [PubMed] [Google Scholar]
- 109.Viscomi M.T., Oddi S., Latini L., Pasquariello N., Florenzano F., Bernardi G. Selective CB2 receptor agonism protects central neurons from remote axotomy-induced apoptosis through the PI3K/Akt Pathway. J Neurosci. 2009;29(14):4564–4570. doi: 10.1523/JNEUROSCI.0786-09.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Bouaboula M., Poinot-Chazel C., Bourrie B., Canat X., Calandra B., Rinaldi-Carmona M. Activation of mitogen-activated protein kinases by stimulation of the central cannabinoid receptor CB1. Biochem J. 1995;312(Pt 2):637–641. doi: 10.1042/bj3120637. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Seyedabadi M., Goodchild A.K., Pilowsky P.M. Differential role of kinases in brain stem of hypertensive and normotensive rats. Hypertension. 2001;38(5):1087–1092. doi: 10.1161/hy1101.096054. [DOI] [PubMed] [Google Scholar]
- 112.Lin Y.Z., Matsumura K., Tsuchihashi T., Fukuhara M., Fujii K., Iida M. Role of ERK and Rho kinase pathways in central pressor action of urotensin II. J Hypertens. 2004;22(5):983–988. doi: 10.1097/00004872-200405000-00021. [DOI] [PubMed] [Google Scholar]
- 113.Chan S.H., Wang L.-L., Tseng H.-L., Chan J.Y. Upregulation of AT1 receptor gene on activation of protein kinase C[beta]/nicotinamide adenine dinucleotide diphosphate oxidase/ERK1/2/c-fos signaling cascade mediates long-term pressor effect of angiotensin II in rostral ventrolateral medulla. J Hypertens. 2007;25(9):1845–1861. doi: 10.1097/HJH.0b013e328217b286. [DOI] [PubMed] [Google Scholar]
- 114.Chan S.H.H., Hsu K.-S., Huang C.-C., Wang L.-L., Ou C.-C., Chan J.Y.H. NADPH oxidase-derived superoxide anion mediates angiotensin II-induced pressor effect via activation of p38 mitogen-activated protein kinase in the rostral ventrolateral medulla. Circ Res. 2005;97(8):772–780. doi: 10.1161/01.RES.0000185804.79157.C0. [DOI] [PubMed] [Google Scholar]
- 115.Ozaita A., Puighermanal E., Maldonado R. Regulation of PI3K/Akt/GSK-3 pathway by cannabinoids in the brain. J Neurochem. 2007;102(4):1105–1114. doi: 10.1111/j.1471-4159.2007.04642.x. [DOI] [PubMed] [Google Scholar]
- 116.Sanchez M.G., Ruiz-Llorente L., Sanchez A.M., Diaz-Laviada I. Activation of phosphoinositide 3-kinase/PKB pathway by CB(1) and CB(2) cannabinoid receptors expressed in prostate PC-3 cells. Involvement in Raf-1 stimulation and NGF induction. Cell Signal. 2003;15(9):851–859. doi: 10.1016/s0898-6568(03)00036-6. [DOI] [PubMed] [Google Scholar]
- 117.Ellert-Miklaszewska A., Kaminska B., Konarska L. Cannabinoids down-regulate PI3K/Akt and Erk signalling pathways and activate proapoptotic function of Bad protein. Cell Signal. 2005;17(1):25–37. doi: 10.1016/j.cellsig.2004.05.011. [DOI] [PubMed] [Google Scholar]
- 118.Greenhough A., Patsos H.A., Williams A.C., Paraskeva C. The cannabinoid delta(9)-tetrahydrocannabinol inhibits RAS-MAPK and PI3K-AKT survival signalling and induces BAD-mediated apoptosis in colorectal cancer cells. Int J Cancer. 2007;121(10):2172–2180. doi: 10.1002/ijc.22917. [DOI] [PubMed] [Google Scholar]
- 119.Galve-Roperh I., Rueda D., Gomez del Pulgar T., Velasco G., Guzman M. Mechanism of extracellular signal-regulated kinase activation by the CB(1) cannabinoid receptor. Mol Pharmacol. 2002;62(6):1385–1392. doi: 10.1124/mol.62.6.1385. [DOI] [PubMed] [Google Scholar]
- 120.Derkinderen P., Valjent E., Toutant M., Corvol J.C., Enslen H., Ledent C. Regulation of extracellular signal-regulated kinase by cannabinoids in hippocampus. J Neurosci. 2003;23(6):2371–2382. doi: 10.1523/JNEUROSCI.23-06-02371.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Martins-Pinge M.C., Araujo G.C., Lopes O.U. Nitric oxide-dependent guanylyl cyclase participates in the glutamatergic neurotransmission within the rostral ventrolateral medulla of awake rats. Hypertension. 1999;34(4):748–751. doi: 10.1161/01.hyp.34.4.748. [DOI] [PubMed] [Google Scholar]
- 122.Mayorov D.N. Nitric oxide synthase inhibition in rostral ventrolateral medulla attenuates pressor response to psychological stress in rabbits. Neurosci Lett. 2007;424(2):89–93. doi: 10.1016/j.neulet.2007.07.007. [DOI] [PubMed] [Google Scholar]
- 123.Martins-Pinge M.C., Garcia M.R., Zoccal D.B., Crestani C.C., Pinge-Filho P. Differential influence of iNOS and nNOS inhibitors on rostral ventrolateral medullary mediated cardiovascular control in conscious rats. Auton Neurosci-Basic Clin. 2007;131(1–2):65–69. doi: 10.1016/j.autneu.2006.07.004. [DOI] [PubMed] [Google Scholar]
- 124.Chan J.Y.H., Chan S.H.H., Chang A.Y.W. Differential contributions of NOS isoforms in the rostral ventrolateral medulla to cardiovascular responses associated with mevinphos intoxication in the rat. Neuropharmacology. 2004;46(8):1184–1194. doi: 10.1016/j.neuropharm.2004.01.017. [DOI] [PubMed] [Google Scholar]
- 125.Chan S.H., Sun E.Y., Chang A.Y. Extracellular signal-regulated kinase 1/2 plays a pro-life role in experimental brain stem death via MAPK signal-interacting kinase at rostral ventrolateral medulla. J Biomed Sci. 2010;17:17. doi: 10.1186/1423-0127-17-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Chan J.Y.H., Chan S.H.H., Li F.C.H., Tsai C.Y., Cheng H.L., Chang A.Y.W. Phasic cardiovascular responses to mevinphos are mediated through differential activation of cGMP/PKG cascade and peroxynitrite via nitric oxide generated in the rat rostral ventrolateral medulla by NOS I and II isoforms. Neuropharmacology. 2005;48(1):161–172. doi: 10.1016/j.neuropharm.2004.08.012. [DOI] [PubMed] [Google Scholar]
- 127.Pilowsky P.M., Goodchild A.K. Baroreceptor reflex pathways and neurotransmitters: 10 years on. J Hypertens. 2002;20(9):1675–1688. doi: 10.1097/00004872-200209000-00002. [DOI] [PubMed] [Google Scholar]
- 128.Drew G.M., Mitchell V.A., Vaughan C.W. Glutamate spillover modulates GABAergic synaptic transmission in the rat midbrain periaqueductal grey via metabotropic glutamate receptors and endocannabinoid signaling. J Neurosci. 2008;28(4):808–815. doi: 10.1523/JNEUROSCI.4876-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Jelsing J., Galzin A.-M., Guillot E., Pruniaux M.-P., Larsen P.J., Vrang N. Localization and phenotypic characterization of brainstem neurons activated by rimonabant and WIN55,212-2. Brain Res Bull. 2009;78(4–5):202–210. doi: 10.1016/j.brainresbull.2008.10.014. [DOI] [PubMed] [Google Scholar]