

Abstract
Background
Intracrinology is the study of the intracellular actions, regulation, trafficking, and interactions of extracellular signaling peptides/proteins.
Methods
We describe the development of intracrine biology since the term was defined in 1984.
Results
Intracrine biology plays a role in many normal and pathological processes and represents a fertile field for the development of novel therapeutics.
Conclusion
Although 30 years old, the field of intracrinology is only now becoming widely accepted. Intracrine principles can be applied to the investigation of physiological processes and to the development of new therapies.
Keywords: Angiotensin II, cytoplasmic and nuclear receptors, intracellular signaling peptides and proteins, intracrine, intracrinology, renin-angiotensin systems
INTRODUCTION
In 1984, this laboratory introduced the term intracrine, meaning the action of a peptide hormone within a cell as opposed to acting at cell surface receptors. Intracrine action could involve the action of a hormone or other extracellular signaling peptide/protein in its cell of synthesis or in target cells after internalization. This notion grew out of studies by our group and others of the intracellular trafficking and actions of the vasoactive peptide angiotensin II. In time, it became clear that peptides/proteins, not usually thought of as hormones, served as extracellular signaling molecules and could also act in the intracellular space: these varied factors displayed intracrine functionality. Surprisingly, included among these proteins were growth factors, cytokines, transcription factors, DNA binding proteins, and enzymes.1-9
We developed the basic principles of intracrine action based on observing the functionality of various intracrines.10-43 For example, many intracrines upregulate either their own synthesis or the synthesis of elements of their signaling cascades. Intracrines can operate in positive feedback loops—that is, in what has been termed a feed-forward mode. For example, angiotensin II action at the nucleus can upregulate the transcription of renin and angiotensinogen. Investigators have reported that high glucose levels stimulate the synthesis of intracellular angiotensin II and establish a feed-forward loop in which angiotensin II upregulates angiotensinogen.35,38 This feed-forward loop appears to play a role in diabetes-related pathology and in particular diabetic cardiomyopathy. Also, in some cases, intracrines can travel to target cells, be internalized, and act in those cells, including setting up feed-forward loops. This action results in a change of state of the target cells. Indeed, if the secretion of an intracrine ceases, the target cells remain in an altered state by virtue of the intracellular feed-forward loop. This has the result of producing a novel form of differentiation, one based on active feed-forward loops. If target cells in turn secrete intracrines to affect nearby cells, a wave of differentiation can propagate through a tissue. Again, intracellular feed-forward loops can maintain the differentiated state, even after secretion ceases. As an example, one can look to the homeodomain transcription factors that are intracrines. Each of these factors contains a cell penetration sequence that enables it to enter target cells. One such factor, when applied to pancreatic duct cells, enters the cells, upregulates its own synthesis, and converts the ductal cells into islet cells. It appears that intracrines can traffick between cells in a variety of ways including in the fluid phase following secretion, traveling in lipid bodies called exosomes that are frequently released by cells in a variety of circumstances, or potentially between cells in thin nanotubes. Many intracrines traffick to the nucleolus; these intracrines almost invariably are either angiogenic or antiangiogenic.13,14,16,17,22,24
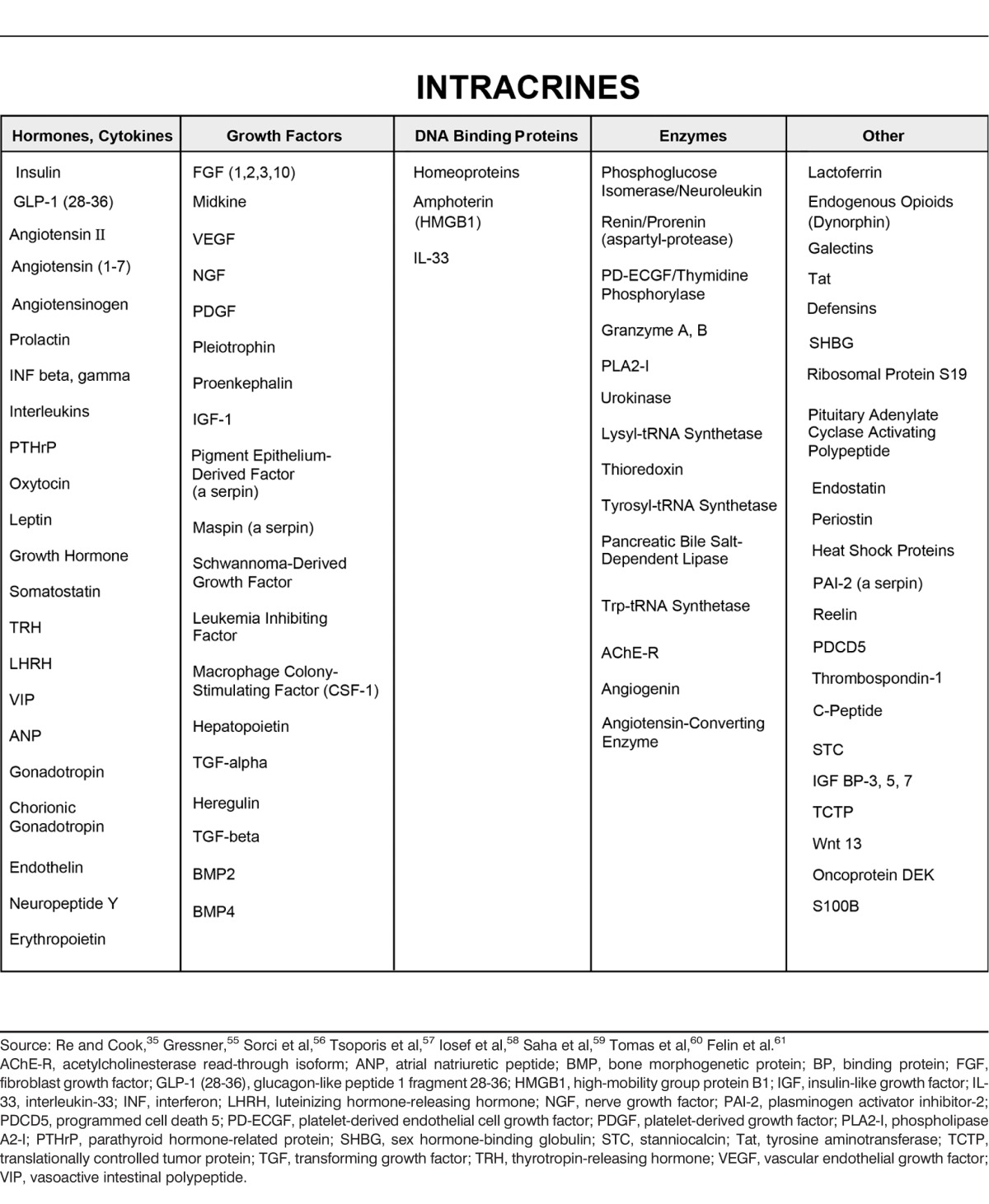
Although not all intracrines display all the characteristics of intracrine function, these notions form the basis of a coherent view of intracrine functionality, a view that surprisingly has been demonstrated to have a heuristic and predictive value. These ideas serve as the basis of a coherent branch of study, intracrinology. Here we discuss some of the findings and implications of this approach. This discussion cannot be comprehensive and, given space constraints, must be somewhat superficial in its treatment of the topic. Although only a few representative examples of the application of intracrinology can be discussed, much of this subject matter has been reviewed in more depth in prior publications. While the intracrine hypothesis originated at Ochsner Clinic Foundation, many scientists around the world have provided the data that support it. It is not possible to credit or reference all these talented people in a short article. References in this manuscript primarily arise from this institution, but these references cite the work of the many others who have made progress possible in this field (Table).1-54
Table.
Intracrines
INTRACRINOLGY AND THE CARDIOVASCULAR SYSTEM
The first evidence of intracellular peptide action was discovered when tritiated angiotensin II was injected into rats that were immediately killed and subjected to electron microscope autoradiography. Angiotensin II-associated radioactivity was found in the nucleus and mitochondria of the rat cells. Then specific angiotensin II receptors were described on cell nuclei and also in association with euchromatin. The binding of angiotensin II to the nuclei upregulated gene transcription and produced changes in nucleosome/chromatin structure consistent with enhanced transcriptional activity. A variety of laboratories reported angiotensin II AT-1 and AT-2 receptors on isolated nuclei, but it was only years later that electromicroscopic immunohistology confirmed angiotensin II in association with euchromatin.1-8,10,11 Collectively, these findings indicate that angiotensin II binds to several nuclear sites and regulates transcription.
Studies of fluorescently labeled AT-1 receptors demonstrated that angiotensin II binding to cell surface receptors could lead to their internalization and nuclear translocation. Moreover, a variety of models was employed to demonstrate that angiotensin II binding to nuclei in vivo could be associated with cellular proliferation or hypertrophy.11-18,23,28,54 The discovery of a renin moiety that was not secreted but was predicted to be synthesized in an active form, as opposed to a prohormone form that required activation, supported the notion that intracellular systems could generate physiologically active angiotensin II in the intracellular space.12,19,54 Indeed, high glucose was later shown to upregulate an intracellular renin-angiotensin system in cardiac myocytes and cardiac fibroblasts, leading to increased intracellular angiotensin II levels. This finding fit nicely with the previously demonstrated increased angiotensin II in cardiac myocytes from patients suffering from diabetic cardiomyopathy.17,18,28,54 Finally, a transgenic mouse line was developed that overexpressed angiotensin II intracellularly but not extracellularly.34,40 These animals became hypertensive and developed renal thrombotic microangiopathy. In this model, nuclear localization of internally synthesized angiotensin II was found, but the binding of the hormone to mitochondria was much more prevalent. Indeed, the binding of angiotensin II to 2 specific mitochondrial electron chain proteins was demonstrated, and the binding was associated with altered generations of reactive oxygen species, long assumed to play a role in angiotensin II-induced pathology. This observation pointed out that future studies of intracrine biology in general, and angiotensin II biology in particular, must take into account what we called noncanonical intracrine action––that is, an intracrine's action independent of its canonical action at its typical receptors.38 This in turn implied that total blockade of angiotensin II action could not be achieved with receptor blockers alone. Any therapeutic implications of this observation remain to be determined. Another line of investigation involved the direct physical introduction of angiotensin II into cardiac myocytes. Clear effects on cellular electrical conductance were observed, and these potentially have an important role to play in arrhythmogenesis.12,15,16,18-43
Although the study of intracrine angiotensin II led the way toward the study of intracrine processes, especially in the cardiovascular system, other intracrines also pointed the way. Parathyroid hormone-related protein (PTHrP), a hormone associated with the hypercalcemia of malignancy, is an intracrine. One form of the protein is retained in the intracellular space and acts at the nucleus. Another form of the protein is secreted to act at cell-surface receptors. In vascular smooth muscle cells, the binding of the hormone to the nucleus is associated with increased cellular proliferation, while the binding to the cell surface receptor is antiproliferative. This example points out some of the complexity of intracrine biology. Other studies showed the important role of intracrine loops in embryonic cardiac development. In this process, dynorphin B, for example, plays an important role.14,16,17,19,22,31,49
A variety of studies revealed that, like angiotensin II, other renin-angiotensin system components are intracrines. Angiotensinogen, (pro)renin, angiotensin-converting enzyme, and angiotensin (1-7) display intracrine features. Indeed, an intracellular renin-angiotensin system has been described in diabetic cardiomyocytes; this system displays intracrine feed-forward loops and participates in diabetic cardiomyopathy.18,19,22,27,28,54
Early on, we predicted that self-sustaining, feed-forward loops played a role in vascular endothelial growth factor (VEGF) biology in angiogenesis. Indeed, such a feed-forward process was described, albeit in myeloma cells and in hematopoietic stem-cell differentiation. However, even more intricate loops involving fibroblast growth factor 2 (FGF2), angiogenin, and VEGF were later shown to participate in a complex intracrine loop to drive angiogenesis. Angiogenin trafficking to the nucleolus was shown to be critically important in this process, and upregulation of ribosomal RNA synthesis played an important role. Interestingly, the nucleolar protein nucleolin also is located on the cell membrane and trafficks various angiogenic and antiangiogenic factors to the nucleolus. The potent antiangiogenic factor endostatin binds to nucleolin and trafficks to the nucleolus where it blocks nucleolin phosphorylation, thereby blocking angiogenesis. Most of the factors that traffick to the nucleolus are either angiogenic or antiangiogenic. Collectively, these results point to a powerful role of intracrine action at the nucleolus in the process of angiogenesis.12,22,24,25,29,44-46
INTRACRINE BIOLOGY AND DEVELOPMENT
Several lines of evidence have been developed that indicate the important role of intracrine biology in development.31,32,44-49 First, it was discovered that homeodomain protein DNA-binding transcription factors leave cells, traffick to nearby cells, are internalized, traffick to the nucleus, and regulate transcription. This system has been described in the case of retina development. When applied to cultured bile duct cells, the homeodomain transcription factor PDX-1 can be internalized, traffick to the nucleus, upregulate its own synthesis, and produce a transformation of the target cell to an islet cell phenotype complete with the synthesis of insulin. In addition to its potential application to the repopulation of beta cells in patients with diabetes, this finding points to intracrine biology's important role in development. The sequence in the homeodomain transcription factor antennapedia that is responsible for its cell uptake has been employed as a cell-penetrating peptide that when fused to a peptide introduces that peptide into target cells. Intracrine-derived cell-penetrating peptides are currently being used in the study of the intracellular biology of a variety of nonintracrine peptides, and in the future they may provide novel therapeutic opportunities.31,32
The endogenous opiate dynorphin drives positive intracrine feedback loops that regulate cardiac stem cell differentiation and thus acts in an intracrine fashion to direct cardiac embryogenesis.31,32,49 Neurons secrete the read-through isoform of acetylcholinesterase (AChE-R) that is the precursor of a fragment generated in the extracellular space. This fragment is then internalized by neurons and has the effect of enhancing fear-conditioned memory. The same sequence of events occurs in hematopoietic stem cells. In this case, there is clear evidence that the internalized fragment upregulates AChE-R synthesis by target cells, resulting in a feed-forward positive loop, and antisense to AChE-R blocks differentiation of the stem cells.31,32,47,48
It is also possible to suggest future applications of intracrine biology to regenerative medicine. Retroviral-mediated transduction of fibroblasts with 4 genes (Oct3/4, Sox2, c-Myc, and Klf4) can reprogram the cells to an embryonic stem cell–like state.31,32 This is of interest because Oct3/4 encodes a homeodomain transcription factor and can likely be internalized by target cells. Sox2 encodes a protein containing a high mobility group motif, and because there is evidence that the high-mobility group box 1 protein functions in an intracrine fashion, Sox2 protein may also be internalized by cells. Thus, 2 of the 4 transducing factors potentially could be delivered without the use of retroviruses. Moreover, the transcription factor Nanog can also be involved in reprogramming. Because it is a homeodomain protein, it likely can be applied extracellularly and function. Because retroviral transduction can be associated with viral integration in the genome and subsequent tumor formation, the direct application of proteins would appear to be considerably safer. The additional transducing factors could potentially be coupled with cell-penetrating peptides to avoid the use of viral transduction.31,32 The first steps of the process have already been reported.50 While speculative, these ideas suggest the potential of the intracrine paradigm.
INTRACRINE BIOLOGY AND CANCER
Given the important role played by intracrine biology in growth and development and given the fact that growth factors such as FGF2, VEGF, and insulin-like growth factor 1 are intracrines, it is not surprising that intracrines play a role in the development and progression of cancer. For example, angiogenesis is an important component of cancer growth, and the important role played by FGF2, VEGF, and angiogenin acting in concert has already been discussed in the cardiovascular system section. Angiogenin has been detected in the nuclei of breast cancer cells, and pharmacologic inhibition of angiogenin trafficking to the nucleus blunts cancer cell proliferation. The presence of angiogenin in the nuclei of breast cancer cells strongly suggests a role for angiogenin in cancer physiology, just as the intracrine VEGF loop in myeloma cells previously described indicates a role for VEGF in that disorder. Additionally, the existence of the same feed-forward loops in cancer cells and vascular cells led us to suggest the notion of intracrine reciprocity, whereby intracrines in the vasculature or other tissues can spill over and strengthen intracrine loops in cancer cells and vice versa. Similarly, intracrine biology can predict the response of some cancers to therapy. For example, the epidermal growth factor receptor 1 (HER1) is a receptor for a variety of intracrine growth factors such as the epidermal growth factor (EGF) and neuregulin. HER1 is a therapeutic target in breast cancer, and antibodies targeting the receptor as well as tyrosine-kinase inhibitors blocking its signaling have been successfully used in the clinic. However, not infrequently drug resistance develops, and this resistance was initially ascribed solely to receptor mutation. Given the intracrine nature of the growth factors involved, we predicted that upregulation of the intracrine HER1 ligands could produce resistance by allowing the intracellular system to signal, unaffected by antibodies working solely at the cell surface. Similarly, upregulation of the intracrine system could bypass the HER1 tyrosine-kinase inhibitor. Soon thereafter, this kind of resistance was demonstrated, pointing to the need for an expanded approach to interrupting the EGF growth network.14,29,32,38,44-46,51
Additionally, the intracrine view suggests novel approaches to cancer therapy. For example, the nucleolar protein nucleolin is intimately involved in the regulation of ribosomes, but it also serves as a cell surface protein that shuttles intracrines from the cell surface to the nucleolus. Nucleolin is upregulated on the surface of a variety of cancer cell types. The interruption of this shuttle pathway in endothelial cells inhibits angiogenesis, and the interruption of the nucleolin shuttle or other trafficking pathways to the nucleus inhibits the growth of various cancer cells. For example, the drug neamine inhibits the trafficking of angiogenin to the nuclei of breast and prostate cancer cells and inhibits their proliferation.24,25,29,52,53
Another potential area of interest is the notion of conditional stem cells. As we have discussed, intracrines play an important role in stem cell regulation. The existence of cancer stem cells has been proposed. These cancer stem cells are proposed to be slow growing but immortal cells that can repopulate tumors after chemotherapy. However, very few cancer cells display traditional stem cell intracrines, and few if any express all needed factors. Thus, we suggested that a reciprocity could occur in which cancer stem cells lacking 1 or another intracrine factor could take up an intracrine factor from a neighboring cell and possibly return a different factor to be internalized by the neighboring cell.24,25,31,32 Thus, 2 conditional cancer stem cells would become fully functional. The interaction of these conditional cancer stem cells is analogous to intracrine reciprocity, and although this kind of intracrine interaction has not yet been demonstrated, it potentially could provide new targets for cancer therapy.21,24,25,31,32,52,53
INTRACRINE BIOLOGY IN OTHER DISORDERS
Intracrine functionality can be found in multiple physiologic processes, leading us to establish the principles of what might be called intracrine pharmacology.29 The interruption of intracrine synthesis, extracellular trafficking, intracellular trafficking, canonical signaling, and noncanonical signaling are targets for the development of therapies for a variety of disorders. Indeed, some currently available drugs such as endostatin have been shown to act through intracrine pathways in target cells. Endostatin colocalizes with nucleolin on the surface of endothelial cells and trafficks with it to the nucleolus. There it inhibits the phosphorylation of nucleolin that is necessary for cell proliferation. Endostatin is, therefore, antiangiogenic, but it must traffick to the nucleolus to block angiogenesis.24,25,29 The principles of intracrine pharmacology can be applied to the development of therapies for the conditions already discussed and to other processes that involve intracrine action.35,37-39 For example, the process of cellular senescence is complex and involves a senescence secretory response on the part of senescent cells.32 This response involves the intercellular trafficking of intracrines, including trafficking via exosomes. Understanding this process and developing agents to modulate it could provide the opportunity to influence senescence and apoptosis in a variety of circumstances.
Transmissible spongiform encephalopathies, diseases such as mad cow disease and Creutzfeld-Jakob disease, involve the spread of abnormally folded prion proteins from cell to cell, with the abnormal protein subsequently causing the misfolding of normal proteins in target cells. The disease is thus propagated by intercellular trafficking proteins. This process can be viewed as a primitive intracrine system. Similar propagation of misfolded proteins occurs in Alzheimer disease, Parkinson disease, amyotrophic lateral sclerosis, and chronic traumatic encephalopathy, among other diseases. One could ask if a more robust intracrine action is involved in these disorders. For example, intracellular trafficking occurs not only by secretion of abnormal proteins or the release of the proteins from dying cells, but rather the evidence shows that intercellular trafficking occurs via exosomes and nanotubes, modalities also associated with intracrine trafficking.43 A more important issue is whether intracrine-like protein feed-forward amplification occurs in these neurodegenerative disorders. If so, interruption of these loops could provide a framework for the development of novel therapies for these disorders.31,32,43
CONCLUSION
Intracrinology is 30 years old and yet is only now becoming widely accepted. However, the evidence to support its validity is overwhelming, suggesting that in the near future intracrine principles will be widely applied to the investigation of physiological processes and to the development of novel therapies. Although this cursory review cannot fully explore all aspects of intracrine biology, it can serve as a guide for interested readers.
Finally, it is appropriate to recognize the researchers at Ochsner Clinic Foundation who have contributed to this effort. Initially, I had the opportunity to work with a talented technician, Meera Parab, without whom the project would have never gotten off the ground. After several years, I had the good fortune of establishing a collaboration with the late Dr Sara Bryan, professor of biology at the University of New Orleans. Along with Jean Brown, Dale Seth, Michael Saucier (who now is a physician on the Ochsner staff), and others, we made great progress in understanding the genomic actions of intracrine angiotensin II. This collaboration only ended with Dr Bryan's death. After several more years, Dr Julia Cook, co-director of the Ochsner Molecular Genetics Laboratory, agreed to collaborate on the study of intracrine angiotensin II. This marked a turning point in the work on intracrine action here and nationwide. Dr Cook's participation led to a near-explosive increase in understanding the operative mechanisms of intracrine angiotensin II action. Dr Cook is an enormously talented and creative scientist whose application of molecular genetic techniques led the way to understanding much of the basic biology of intracrine angiotensin II. Thanks to Dr Cook, during this period Ochsner remained the leader in the emerging field of intracrinology. Collectively, this team has accomplished a great deal.
Footnotes
The author has no financial or proprietary interest in the subject matter of this article.
This article meets the Accreditation Council for Graduate Medical Education and the American Board of Medical Specialties Maintenance of Certification competencies for Patient Care and Medical Knowledge.
REFERENCES
- 1.Re RN, MacPhee AA, Fallon JT. Specific nuclear binding of angiotensin II by rat liver and spleen nuclei. Clin Sci (Lond) 1981 Dec;61(Suppl 7):245s–247s. doi: 10.1042/cs061245s. [DOI] [PubMed] [Google Scholar]
- 2.Re RN. Changes in nuclear initiation sites after the treatment of isolated nuclei with angiotensin II. Clin Sci (Lond) 1982 Oct;63(Suppl 8):191s–193s. [Google Scholar]
- 3.Re RN, LaBiche RA, Bryan SE. Nuclear-hormone mediated changes in chromatin solubility. Biochem Biophys Res Commun. 1983 Jan 14;110(1):61–68. doi: 10.1016/0006-291x(83)91260-3. [DOI] [PubMed] [Google Scholar]
- 4.Re R, Parab M. Effect of angiotensin II on RNA synthesis by isolated nuclei. Life Sci. 1984 Feb 13;34(7):647–651. doi: 10.1016/0024-3205(84)90228-5. [DOI] [PubMed] [Google Scholar]
- 5.Re RN, Vizard DL, Brown J, Bryan SE. Angiotensin II receptors in chromatin fragments generated by micrococcal nuclease. Biochem Biophys Research Commun. 1984 Feb 29;119(1):220–227. doi: 10.1016/0006-291x(84)91641-3. [DOI] [PubMed] [Google Scholar]
- 6.Re R, Bryan SE. Functional intracellular renin-angiotensin systems may exist in multiple tissues. Clin Exp Hypertens A. 1984;6(10-11):1739–1742. doi: 10.3109/10641968409046069. [DOI] [PubMed] [Google Scholar]
- 7.Bryan SE, Legros L, Brown J, Byrne C, Re RN. Copper-rich nucleoprotein generated by micrococcal nuclease. Biol Trace Elem Res. 1985 Nov;8(3):219–229. doi: 10.1007/BF02917461. [DOI] [PubMed] [Google Scholar]
- 8.Saucier MA, Wang X, Re RN, Brown J, Bryan SE. Effects of ionic strength on endogenous nuclease activity in chelated and nonchelated chromatin. J Inorg Biochem. 1991 Feb 1;41(2):117–124. doi: 10.1016/0162-0134(91)80005-3. [DOI] [PubMed] [Google Scholar]
- 9.Cook JL, Bhandaru S, Giardina JF, Claycomb W, Ré RN. Identification and antisense inhibition of a renin-angiotensin system in transgenic cardiomyocytes. Am J Physiol. 1995 Apr;268(4 Pt 2):H1471–H1482. doi: 10.1152/ajpheart.1995.268.4.H1471. [DOI] [PubMed] [Google Scholar]
- 10.Re R. The nature of intracrine peptide hormone action. Hypertension. 1999 Oct;34(4 Pt 1):534–538. doi: 10.1161/01.hyp.34.4.534. [DOI] [PubMed] [Google Scholar]
- 11.Re RN. On the biological actions of intracellular angiotensin. Hypertension. 2000 Jun;35(6):1189–1190. doi: 10.1161/01.hyp.35.6.1189. [DOI] [PubMed] [Google Scholar]
- 12.Cook JL, Zhang Z, Re RN. In vitro evidence for an intracellular site of angiotensin action. Circ Res. 2001 Dec;89(12):1138–1146. doi: 10.1161/hh2401.101270. [DOI] [PubMed] [Google Scholar]
- 13.Re RN. The origins of intracrine hormone action. Am J Med Sci. 2002 Jan;323(1):43–48. doi: 10.1097/00000441-200201000-00008. [DOI] [PubMed] [Google Scholar]
- 14.Re RN. Toward a theory of intracrine hormone action. Regul Pept. 2002 Jun 15;106(1-3):1–6. doi: 10.1016/s0167-0115(02)00031-9. [DOI] [PubMed] [Google Scholar]
- 15.Cook JL, Giardina JF, Zhang Z, Re RN. Intracellular angiotensin II increases the long isoform of PDGF mRNA in rat hepatoma cells. J Mol Cell Cardiol. 2002 Nov;34(11):1525–1537. doi: 10.1006/jmcc.2002.2106. [DOI] [PubMed] [Google Scholar]
- 16.Re RN. The intracrine hypothesis and intracellular peptide hormone action. Bioessays. 2003 Apr;25(4):401–409. doi: 10.1002/bies.10248. [DOI] [PubMed] [Google Scholar]
- 17.Re RN. Implications of intracrine hormone action for physiology and medicine. Am J Physiol Heart Circ Physiol. 2003 Mar;284(3):H751–H757. doi: 10.1152/ajpheart.00935.2002. [DOI] [PubMed] [Google Scholar]
- 18.Re RN. Cardiac angiotensin II: an intracrine hormone? Am J Hypertens. 2003 May;16(5 Pt 1):426–427. doi: 10.1016/s0895-7061(03)00038-4. [DOI] [PubMed] [Google Scholar]
- 19.Re RN. Intracellular renin and the nature of intracrine enzymes. Hypertension. 2003 Aug;42(2):117–122. doi: 10.1161/01.HYP.0000082495.93495.5B. [DOI] [PubMed] [Google Scholar]
- 20.Cook JL, Re R, Alam J, Hart M, Zhang Z. Intracellular angiotensin II fusion protein alters AT1 receptor fusion protein distribution and activates CREB. J Mol Cell Cardiol. 2004 Jan;36(1):75–90. doi: 10.1016/j.yjmcc.2003.09.021. [DOI] [PubMed] [Google Scholar]
- 21.Re RN. A proposal regarding the biology of memory: participation of intracrine peptide networks. Med Hypotheses. 2004;63(5):887–894. doi: 10.1016/j.mehy.2004.04.027. [DOI] [PubMed] [Google Scholar]
- 22.Re RN, Cook JL. The intracrine hypothesis: an update. Regul Pept. 2006 Jan 15;133(1-3):1–9. doi: 10.1016/j.regpep.2005.09.012. [DOI] [PubMed] [Google Scholar]
- 23.Cook JL, Mills SJ, Naquin R, Alam J, Re RN. Nuclear accumulation of the AT1 receptor in a rat vascular smooth muscle cell line: effects upon signal transduction and cellular proliferation. J Mol Cell Cardiol. 2006 May;40(5):696–707. doi: 10.1016/j.yjmcc.2005.11.014. [DOI] [PubMed] [Google Scholar]
- 24.Re RN, Cook JL. An intracrine view of angiogenesis. Bioessays. 2006 Sep;28(9):943–953. doi: 10.1002/bies.20459. [DOI] [PubMed] [Google Scholar]
- 25.Re RN, Cook JL. Potential therapeutic implications of intracrine angiogenesis. Med Hypotheses. 2007;69(2):414–421. doi: 10.1016/j.mehy.2006.10.065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Cook JL, Mills SJ, Naquin RT, Alam J, Re RN. Cleavage of the angiotensin II type 1 receptor and nuclear accumulation of the cytoplasmic carboxy-terminal fragment. Am J Physiol Cell Physiol. 2007 Apr;292(4):C1313–C1322. doi: 10.1152/ajpcell.00454.2006. [DOI] [PubMed] [Google Scholar]
- 27.Re R. Intracellular renin-angiotensin system: the tip of the intracrine physiology iceberg. Am J Physiol Heart Circ Physiol. 2007 Aug;293(2):H905–H906. doi: 10.1152/ajpheart.00552.2007. [DOI] [PubMed] [Google Scholar]
- 28.Re RN, Cook JL. Mechanisms of disease: Intracrine physiology in the cardiovascular system. Nat Clin Pract Cardiovasc Med. 2007 Oct;4(10):549–557. doi: 10.1038/ncpcardio0985. [DOI] [PubMed] [Google Scholar]
- 29.Re RN, Cook JL. The basis of an intracrine pharmacology. J Clin Pharmacol. 2008 Mar;48(3):344–350. doi: 10.1177/0091270007312155. [DOI] [PubMed] [Google Scholar]
- 30.Cook JL, Re RN, deHaro DL, Abadie JM, Peters M, Alam J. The trafficking protein GABARAP binds to and enhances plasma membrane expression and function of the angiotensin II type 1 receptor. Circ Res. 2008 Jun 20;102(12):1539–1547. doi: 10.1161/CIRCRESAHA.108.176594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Re RN, Cook JL. The physiological basis of intracrine stem cell regulation. Am J Physiol Heart Circ Physiol. 2008 Aug;295(2):H447–H453. doi: 10.1152/ajpheart.00461.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Re RN, Cook JL. Senescence, apoptosis, and stem cell biology: the rationale for an expanded view of intracrine action. Am J Physiol Heart Circ Physiol. 2009 Sep;297(3):H893–H901. doi: 10.1152/ajpheart.00414.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Alam J, Deharo D, Redding KM, Re RN, Cook JL. C-terminal processing of GABARAP is not required for trafficking of the angiotensin II type 1A receptor. Regul Pept. 2010 Jan 8;159(1-3):78–86. doi: 10.1016/j.regpep.2009.09.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Redding KM, Chen BL, Singh A, et al. Transgenic mice expressing an intracellular fluorescent fusion of angiotensin II demonstrate renal thrombotic microangiopathy and elevated blood pressure. Am J Physiol Heart Circ Physiol. 2010 Jun;298(6):H1807–H1818. doi: 10.1152/ajpheart.00027.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Re RN, Cook JL. The mitochondrial component of intracrine action. Am J Physiol Heart Circ Physiol. 2010 Sep;299(3):H577–H583. doi: 10.1152/ajpheart.00421.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Re RN. Lysosomal action of intracrine angiotensin II. Focus on “Intracellular angiotensin II activates rat myometrium”. Am J Physiol Cell Physiol. 2011 Sep;301(3):C553–C554. doi: 10.1152/ajpcell.00232.2011. [DOI] [PubMed] [Google Scholar]
- 37.Cook JL, Singh A, DeHaro D, Alam J, Re RN. Expression of a naturally occurring angiotensin AT(1) receptor cleavage fragment elicits caspase-activation and apoptosis. Am J Physiol Cell Physiol. 2011 Nov;301(5):C1175–C1185. doi: 10.1152/ajpcell.00040.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Re RN, Cook JL. Noncanonical intracrine action. J Am Soc Hypertens. 2011 Nov-Dec;5(6):435–448. doi: 10.1016/j.jash.2011.07.001. [DOI] [PubMed] [Google Scholar]
- 39.Vitko JR, Re RN, Alam J, Cook JL. Cell-penetrating peptides corresponding to the angiotensin II Type 1 receptor reduce receptor accumulation and cell surface expression and signaling. Am J Hypertens. 2012 Jan;25(1):24–28. doi: 10.1038/ajh.2011.162. [DOI] [PubMed] [Google Scholar]
- 40.Cook JL, Re RN. Lessons from in vitro studies and a related intracellular angiotensin II transgenic mouse model. Am J Physiol Regul Integr Comp Physiol. 2012 Mar 1;302(5):R482–R493. doi: 10.1152/ajpregu.00493.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Re RN, Chen B, Alam J, Cook JL. Reduction of blood pressure by AT1 receptor decoy peptides. Ochsner J. 2013 Spring;13(1):33–36. [PMC free article] [PubMed] [Google Scholar]
- 42.Singh A, Vitko J, Re RN, Cook JL. Intracellular enhanced cyan fluorescent protein/angiotensin II does not modify angiotensinogen accumulation in transgenic mice. Ochsner J. 2013 Spring;13(1):37–41. [PMC free article] [PubMed] [Google Scholar]
- 43.Re RN. Could intracrine biology play a role in the pathogenesis of transmissible spongiform encephalopathies, Alzheimer's disease and other neurodegenerative diseases? Am J Med Sci. 2014 Apr;347(4):312–320. doi: 10.1097/MAJ.0b013e3182a28af3. [DOI] [PubMed] [Google Scholar]
- 44.Gerber HP, Malik AK, Solar GP, et al. VEGF regulates haematopoietic stem cell survival by an internal autocrine loop mechanism. Nature. 2002 Jun 27;417(6892):954–958. doi: 10.1038/nature00821. [DOI] [PubMed] [Google Scholar]
- 45.Gerber HP, Ferrara N. The role of VEGF in normal and neoplastic hematopoiesis. J Mol Med (Berl) 2003 Jan;81(1):20–31. doi: 10.1007/s00109-002-0397-4. [DOI] [PubMed] [Google Scholar]
- 46.Vincent L, Jin DK, Karajannis MA, et al. Fetal stromal-dependent paracrine and intracrine vascular endothelial growth factor-a/vascular endothelial growth factor receptor-1 signaling promotes proliferation and motility of human primary myeloma cells. Cancer Res. 2005 Apr 15;65(8):3185–3192. doi: 10.1158/0008-5472.CAN-04-3598. [DOI] [PubMed] [Google Scholar]
- 47.Nijholt I, Farchi N, Kye M, et al. Stress-induced alternative splicing of acetylcholinesterase results in enhanced fear memory and long-term potentiation. Mol Psychiatry. 2004 Feb;9(2):174–183. doi: 10.1038/sj.mp.4001446. [DOI] [PubMed] [Google Scholar]
- 48.Deutsch VR, Pick M, Perry C, et al. The stress-associated acetylcholinesterase variant AChE-R is expressed in human CD34(+) hematopoietic progenitors and its C-terminal peptide ARP promotes their proliferation. Exp Hematol. 2002 Oct;30(10):1153–1161. doi: 10.1016/s0301-472x(02)00900-1. [DOI] [PubMed] [Google Scholar]
- 49.Ventura C, Branzi A. Autocrine and intracrine signaling for cardiogenesis in embryonic stem cells: a clue for the development of novel differentiating agents. Handb Exp Pharmacol. 2006;(174):123–146. [PubMed] [Google Scholar]
- 50.Zhou H, Wu S, Joo JY, et al. Generation of induced pluripotent stem cells using recombinant proteins. Cell Stem Cell. 2009 May 8;4(5):381–384. doi: 10.1016/j.stem.2009.04.005. Erratum in: Cell Stem Cell. 2009 Jun 5;4(6):581. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Ferrer-Soler L, Vazquez-Martin A, Brunet J, Menendez JA, De Llorens R, Colomer R. An update of the mechanisms of resistance to EGFR-tyrosine kinase inhibitors in breast cancer: Gefitinib (Iressa)-induced changes in the expression and nucleo-cytoplasmic trafficking of HER-ligands (Review) Int J Mol Med. 2007 Jul;20(1):3–10. [PubMed] [Google Scholar]
- 52.Ibaragi S, Yoshioka N, Li S, et al. Neamine inhibits prostate cancer growth by suppressing angiogenin-mediated rRNA transcription. Clin Cancer Res. 2009 Mar 15;15(6):1981–1988. doi: 10.1158/1078-0432.CCR-08-2593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Li S, Ibaragi S, Hu GF. Angiogenin as a molecular target for the treatment of prostate cancer. Curr Cancer Ther Rev. 2011 May;7(2):83–90. doi: 10.2174/1573394711107020083. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Kumar R, Yong QC, Thomas CM, Baker KM. Intracardiac intracellular angiotensin system in diabetes. Am J Physiol Regul Integr Comp Physiol. 2012 Mar 1;302(5):R510–R517. doi: 10.1152/ajpregu.00512.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Gressner OA. Intracrine signaling mechanisms of activin A and TGF-β. Vitam Horm. 2011;85:59–77. doi: 10.1016/B978-0-12-385961-7.00004-4. [DOI] [PubMed] [Google Scholar]
- 56.Sorci G, Riuzzi F, Arcuri C, et al. The many faces of S100B protein: when an extracellular factor inactivates its own receptor and activates another one. Ital J Anat Embryol. 2010;115(1-2):147–151. [PubMed] [Google Scholar]
- 57.Tsoporis JN, Mohammadzadeh F, Parker TG. Intracellular and extracellular effects of S100B in the cardiovascular response to disease. Cardiovasc Psychiatry Neurol. 2010;2010:206073. doi: 10.1155/2010/206073. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Iosef C, Gkourasas T, Jia CY, Li SS, Han VK. A functional nuclear localization signal in insulin-like growth factor binding protein-6 mediates its nuclear import. Endocrinology. 2008 Mar;149(3):1214–1226. doi: 10.1210/en.2007-0959. [DOI] [PubMed] [Google Scholar]
- 59.Saha AK, Kappes F, Mundade A, et al. Intercellular trafficking of the nuclear oncoprotein DEK. Proc Natl Acad Sci U S A. 2013 Apr 23;110(17):6847–6852. doi: 10.1073/pnas.1220751110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Tomas E, Stanojevic V, Habener JF. GLP-1-derived nonapeptide GLP-1(28-36)amide targets to mitochondria and suppresses glucose production and oxidative stress in isolated mouse hepatocytes. Regul Pept. 2011 Apr 11;167(2-3):177–184. doi: 10.1016/j.regpep.2011.01.003. [DOI] [PubMed] [Google Scholar]
- 61.Felin JE, Mayo JL, Loos TJ, et al. Nuclear variants of bone morphogenetic proteins. BMC Cell Biol. 2010 Mar;15(11):20. doi: 10.1186/1471-2121-11-20. doi: 10.1186/1471-2121-11-20. [DOI] [PMC free article] [PubMed] [Google Scholar]