

Abstract
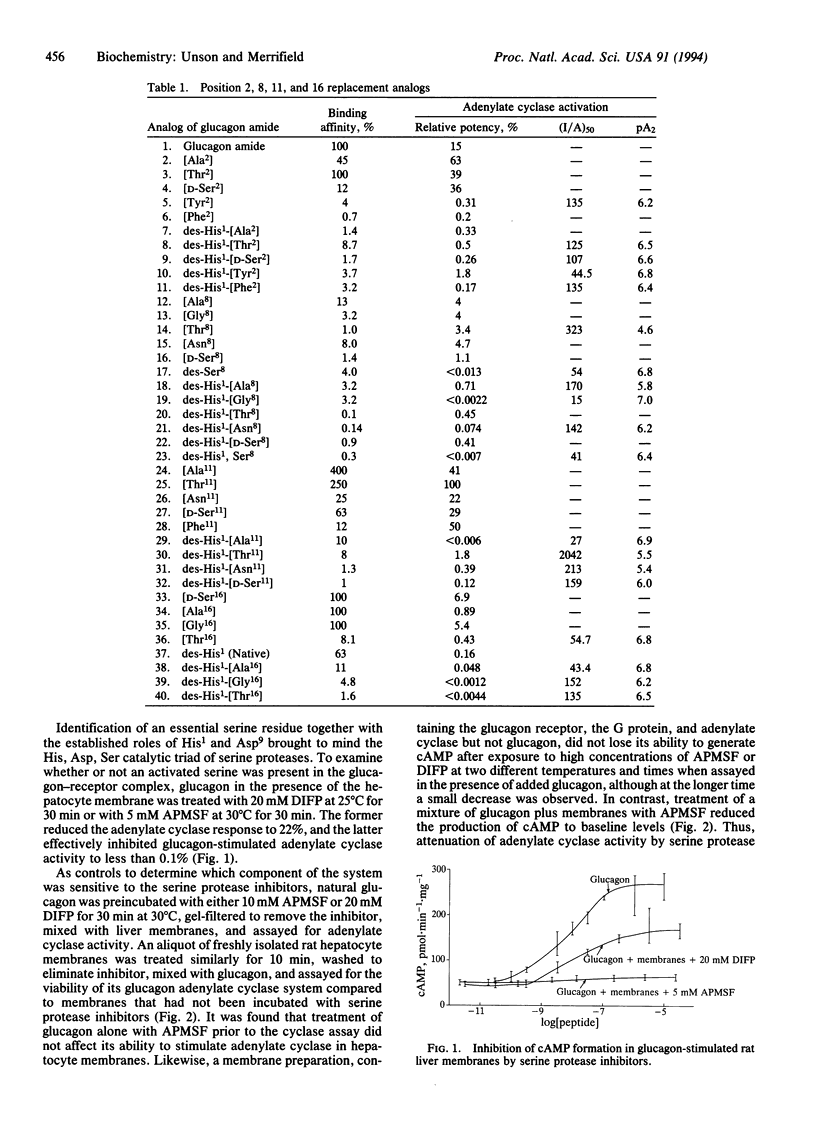
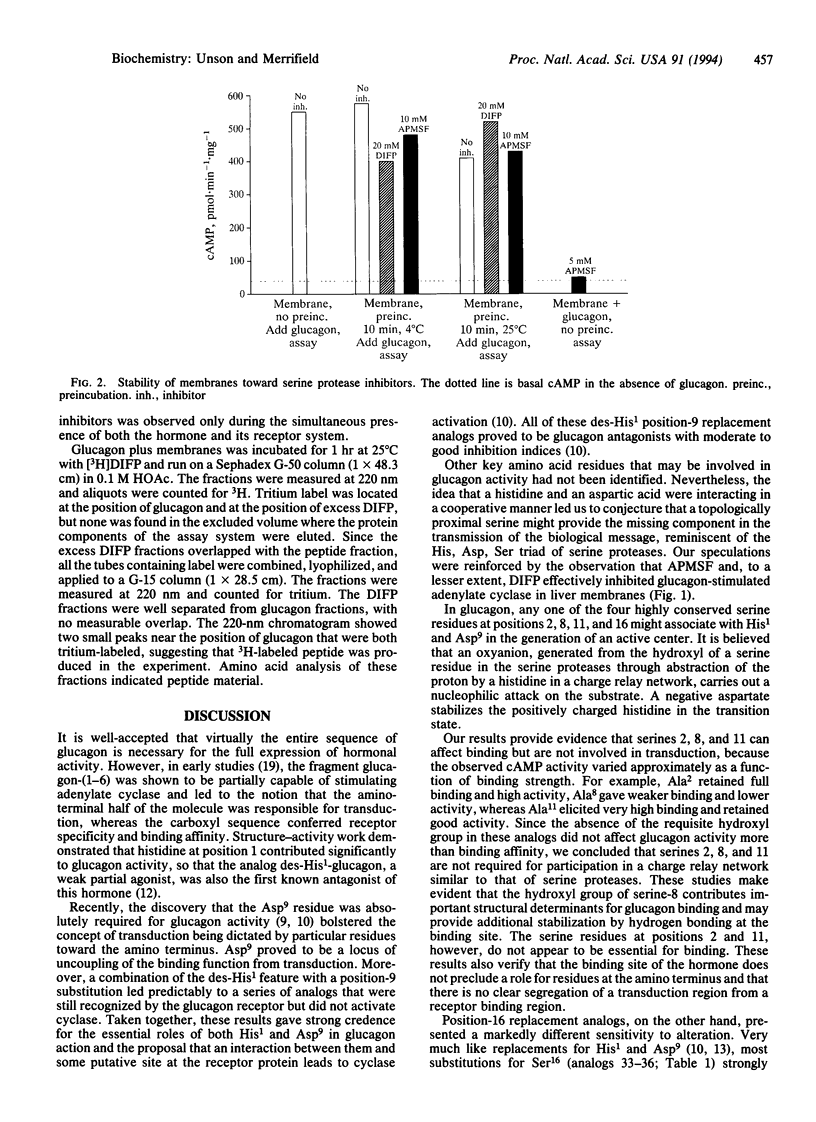
Several glucagon analogs containing substitutions for serine have been synthesized to assess the role of the four serine residues in the hormone. The strategic importance of His1 has been confirmed, and we have previously identified an aspartic acid critical for activity at position 9. While these findings have led to a series of pure glucagon antagonists, the details of specific glucagon-receptor interactions that switch on the ensuing signaling events are still not readily apparent. The requirement for serine was tested by the chemical synthesis of a series of analogs containing substitutions for the hydrophilic hydroxyl group in each of the highly conserved serine residues at positions 2, 8, 11, and 16 of glucagon. The resulting analogs were analyzed in rat hepatocyte membranes for their receptor-binding affinities as well as their abilities to stimulate adenylate cyclase. Positions 2 and 8 were the most sensitive to modification, where both binding and activity were adversely affected. This is consistent with the notion that although the sequence responsible for transduction lies in the amino-terminal half of glucagon, some residues at that end also contribute to binding affinity. Modifications at position 11 generated high-binding-affinity derivatives that were full or moderate agonists. In contrast, position 16 replacement analogs maintained significant receptor binding affinities while the agonist properties were almost completely lost, thus separating binding and transduction functions. Therefore, Ser16 is a third critical residue that determines glucagon activity. It is postulated, but not proven, that a serine residue, together with His1 and Asp9, may participate in the putative active center of glucagon, which, upon initial recognition and binding to receptor, leads to transduction of the biological signal. A dependence of the glucagon action on a three-residue cooperative mechanism might be analogous to the charge-relay scheme of serine proteases. It is suggested that, after binding to its receptor, glucagon becomes activated and functions like a coenzyme in catalyzing the specific hydrolysis of a peptide bond in the receptor, generating new amino and carboxyl end groups, and that one of these exposed chains may contact the GTP-binding protein and activate it for further interaction with adenylate cyclase. This idea was supported by inhibition experiments with 4-amidinophenylmethanesulfonyl fluoride (APMSF), a specific and irreversible inhibitor of serine proteases, which at a concentration of 5 mM completely suppressed cAMP formation by glucagon in liver membranes. cAMP formation was not affected if either glucagon or membranes were separately pretreated with APMSF and then assayed.
Full text
PDF


Selected References
These references are in PubMed. This may not be the complete list of references from this article.
- Iyengar R., Herberg J. T. Structural analysis of the hepatic glucagon receptor. Identification of a guanine nucleotide-sensitive hormone-binding region. J Biol Chem. 1984 Apr 25;259(8):5222–5229. [PubMed] [Google Scholar]
- Neer E. J., Clapham D. E. Roles of G protein subunits in transmembrane signalling. Nature. 1988 May 12;333(6169):129–134. doi: 10.1038/333129a0. [DOI] [PubMed] [Google Scholar]
- Okamoto T., Murayama Y., Hayashi Y., Inagaki M., Ogata E., Nishimoto I. Identification of a Gs activator region of the beta 2-adrenergic receptor that is autoregulated via protein kinase A-dependent phosphorylation. Cell. 1991 Nov 15;67(4):723–730. doi: 10.1016/0092-8674(91)90067-9. [DOI] [PubMed] [Google Scholar]
- Pilch P. F., Axelrod J. D., Colello J., Czech M. P. Unimpaired signal transduction by the adipocyte insulin receptor following its partial proteolytic fragmentation. J Biol Chem. 1981 Feb 25;256(4):1570–1575. [PubMed] [Google Scholar]
- Sheetz M. J., Tager H. S. Receptor-linked proteolysis of membrane-bound glucagon yields a membrane-associated hormone fragment. J Biol Chem. 1988 Jun 15;263(17):8509–8514. [PubMed] [Google Scholar]
- Unger R. H. The Banting Memorial Lecture 1975. Diabetes and the alpha cell. Diabetes. 1976 Feb;25(2):136–151. doi: 10.2337/diab.25.2.136. [DOI] [PubMed] [Google Scholar]
- Vu T. K., Hung D. T., Wheaton V. I., Coughlin S. R. Molecular cloning of a functional thrombin receptor reveals a novel proteolytic mechanism of receptor activation. Cell. 1991 Mar 22;64(6):1057–1068. doi: 10.1016/0092-8674(91)90261-v. [DOI] [PubMed] [Google Scholar]