

Abstract
Purpose
Keratoconus (KC) is characterized by progressive vision loss due to corneal thinning and structural abnormalities. It is hypothesized that KC is caused by deregulated collagen levels and collagen fibril-maturating enzyme lysyl oxidase (LOX). Further, it is currently not understood whether the gene expression deregulated by the corneal epithelium influences KC pathogenesis. We studied (i) the expressions of the LOX, collagen I (COL IA1), collagen IV (COL IVA1), MMP9, and IL6 genes in KC corneal epithelia, (ii) validated their expression levels in patient tissues, and (iii) correlated expression levels with KC disease severity. The primary goal of this study was to evaluate the importance of these genes in the progression of KC.
Methods
We analyzed the gene expression levels of the key proteins LOX, collagens (COL IA1 and COL IVA1), MMP9, and IL6 in debrided corneal epithelia from a large cohort of KC patients (90 eyes) and compared them to control patients (52 eyes) without KC. We measured the total LOX activity in the tears of KC patients compared to controls. We also correlated the protein expression levels of LOX and collagens by immunohistochemistry (IHC) in primary tissues from KC patients (27 eyes) undergoing keratoplasty compared to healthy donor corneas (15 eyes).
Results
We observed a significant reduction in LOX transcript levels in KC corneal epithelia, and LOX activity in KC tears correlated with disease severity. Collagen transcripts were also reduced in KC while MMP9 transcript levels were upregulated and correlated with disease severity. IL6 was moderately increased in KC patients. IHC demonstrated a reduction in the protein expression levels of LOX in the epithelium and collagen IV in the basement membrane of KC patients compared to healthy donor corneas.
Conclusions
The data demonstrates that the structural deformity of the KC cornea may be dependent on reduced expressions of collagens and LOX, as well as on MMP9 elevated by the corneal epithelium.
Introduction
Keratoconus (KC) is a corneal disorder characterized by progressive corneal thinning leading to astigmatism and visual loss [1,2], often affecting young adults. KC onset typically occurs in the second decade of life, and it has a variable progression lasting until about the third or fourth decade of life, after which the disease stabilizes. The Collaborative Longitudinal Evaluation of Keratoconus (CLEK) study reported a decrease in high- and low-contrast visual acuity accompanied by progressive corneal steepening [3]. This disease is characterized by corneal distortion, as measured by keratometry, as well as by a displaced, thin corneal apex, isolated corneal steepening, deposits of iron in a ring segment in corneal epithelia termed Fleischer’s ring, posterior corneal stroma demonstrating Vogt’s striae, and some scarring in the central cornea from the progressive disease [2,4,5]. In addition, the KC cornea demonstrates breaks in the Bowman’s layer, an abnormal stromal collagen structure, and focal fibrotic deposits [6]. KC is bilateral, though not symmetric, but there are differences neither between the left and right eyes nor between males and females [3,7]. Corneal scarring due to abnormal eye rubbing and contact lens wear has been suggested as important in KC pathogenesis [8]. The CLEK study data reports the 5-year corneal scarring incidence at 13.7% overall and 16.7% for patients wearing contact lenses [9]. In addition, atopy, Down syndrome, connective tissue disorders, and genetic factors have also been associated with KC [10,11].
KC is currently treated by various types of contact lenses or surgical procedures, such as corneal collagen crosslinking (C3R), accelerated collagen crosslinking (KXL), topography-guided photo refractive keratectomy (T-PRK), intrastromal corneal ring segments (ICRS), etc. [12-14]. These treatment options rely on maintaining or enhancing the corneal stiffness in an attempt to retard KC progression. Despite evolving knowledge about the nature of KC, the molecular factors driving this loss of corneal structure integrity are poorly understood. Recently, various inflammatory factors, such as IL6, TNFα, MMP9, cathepsins, etc., have been shown to be elevated in the tears of KC patients [15-18], suggesting a deregulation of underlying molecular pathways.
The disease process in KC induces abnormal gene expressions [19], as well as biochemical and pathologic changes [6,20,21] in the cornea that consequently affect corneal structure [22]. Thus, the structural integrity of the cornea is disrupted in KC [6], indicating there may be changes in the collagen content or its modifying enzymes in the cornea [20]. Epithelial or stromal proteins that are deregulated in KC might be a response to the thinning process or may be drivers of the process itself [23-25]. The tears of KC patients have higher levels of proinflammatory cytokines, cell adhesion molecules, and matrix metalloproteinases (MMPs) compared to controls [15,17,26]. Enzymes of the MMP family [27] mediate the degradation of extracellular matrix proteins, typically in response to stress or injury [28]. Matrix metalloproteinase-9 (MMP9) is one of the matrix-degrading enzymes produced by the human corneal epithelium [29]. MMP9 activity is regulated by cytokine Interleukin 6 (IL6) and has been implicated in the degradation of type I and IV collagen fibers in ocular disorders [30], as well as in certain cancers [31]. As collagen forms an integral part of the cornea, which is most affected in the ectasia that characterizes KC [6,20-22], it is important to understand the factors that regulate different forms of collagen.
Lysyl oxidase (LOX) [32,33], a copper-dependent amine oxidase, is responsible for the development of lysine-derived crosslinks in extracellular matrix proteins, such as collagen and elastin [34]. LOX oxidizes the epsilon amino groups of peptidyl lysines into reactive aldehydes. The resulting aldehyde groups then undergo spontaneous condensation with unreacted epsilon amino groups or neighboring aldehyde groups, converting collagen or elastin monomers into insoluble fibers [34]. Recent molecular studies have revealed the existence of a human LOX family consisting of the five paralogues [35]. Each LOX paralogue contains a conserved copper-binding domain, residues for lysyl-tyrosyl quinone (LTQ), and a cytokine receptor-like (CRL) domain in the carboxyl (C) termini. The amino (N)-terminal regions of the LOX family members, in contrast, show little sequence homology, except that the N termini of LOXL2–4 contain four copies of scavenger receptor cysteine-rich domains [35]. Several new functions, such as tumor suppression, cellular senescence, and chemotaxis, have been attributed to LOX [36-38], the impairment of which contributes to diseases like atherosclerosis, aortic aneurysms, pulmonary or hepatic fibrosis, cutis laxa, proliferative diabetic retinopathy [39], and hypertrophic scars [37,40].
The role of LOX in KC is proposed by two conflicting reports, suggesting reductions [41] and increases in KC [42]. Additionally, a linkage study of familial and case-control KC patients suggests that the LOX gene containing genomic loci may be associated with KC [43], although pathogenic mutants were not found [44]. However, it has yet to be determined how LOX activity is altered in KC and whether the deregulation contributes to the pathogenesis of this progressive disorder. It is important to understand the correlation of LOX activity with the expressions of different forms of collagen or other inflammatory genes. Therefore, this study is designed to investigate whether the corneal epithelium has a functional role in KC progression. The expression of select genes associated with the corneal structure, such as LOX, collagen I (COL IA1), collagen IV (COL IVA1), MMP9, and IL6, were measured in debrided corneal epithelia from patients. The correlations of gene expression data with different clinical grades of KC is expected to help elucidate any potential association of these genes with disease severity.
Methods
Clinical study design and endpoints
This study was approved by the Narayana Nethralaya Institutional Review Board and performed as per the Indian Council for Medical Research (ICMR). In addition, institutional ethics guidelines were in accordance with the tenets of the Declaration of Helsinki. All patient samples were collected after obtaining informed, written consent as per institutional and ethics board guidelines. For minor patients, informed, written consent was obtained from their guardians or next-of-kin. This study has also been registered at ClinicalTrials.gov (Identifier: NCT01746823).
The study cohort was selected from patients who reported to the cornea clinic at Narayana Nethralaya, Bangalore. KC patients of Indian origin undergoing standard-of-care surgical intervention for the correction of visual defects were included in the study. In total, 80 KC patients undergoing topography guided photorefractive keratectomy (T-PRK) or corneal crosslinking were included for the collection of corneal epithelia. Additionally, 29 subjects with normal corneal topography undergoing photorefractive keratectomy (PRK) for the correction of refractive errors were used as controls for the corneal epithelia samples. Patients using contact lenses, any type of anti-inflammatory ocular or systemic medications (e.g., anti-allergic, anti-inflammatory drugs), or having undergone any prior surgical intervention (e.g., penetrating keratoplasty/corneal collagen crosslinking, cataract surgery, etc.) for either eye were excluded from the study. Patients with a recent (less than 3 months) allergic or infection history in either eye were excluded. All subjects also underwent a dry eye evaluation using Schirmer’s test and corneal staining. Subjects with concurrent symptoms of dry eye were excluded. Patients with systemic inflammatory or autoimmune diseases were also excluded. Tear samples were also collected from patients before surgery. Additionally, immunohistochemical (IHC) studies were performed on a separate group of archived tissue samples, as described below.
The experimental endpoints included analyses of the biomarkers LOX, MMP9, IL6, collagen I, and collagen IV in patient-derived samples. There were no safety endpoints, as there were no experimental interventions or any deviations from the standard of care.
Patient diagnosis and grading
Patient diagnosis for KC was done using retinoscopy, slit lamp biomicroscopy, and corneal refraction measurements. Corneal topographic images of all the patients were acquired with a Pentacam (OCULUS Optikgeräte GmbH, Germany) upon first presentation at the clinic and were used for the diagnosis and grading of KC patients [2,45] in this study. KC grades were determined from cumulative analyses of biomicroscopy data, KC features observed by slit lamp, spherical, and cylindrical refraction changes, mean central keratometry measurements, and corneal thickness, as defined by the Amsler–Krumeich classification [46,47]. However, grades 2 and 4 were combined as the severe KC group, as reported earlier [48]. No drug-based tests or analyses were performed during this study.
The KC subjects were graded based on methods reported in recent studies [4]. Mean keratometry was the primary classification variable. The mean keratometry reading was calculated as the average of the steep and flat axis keratometry readings. KC eyes with a mean keratometry reading of less than 48D were classified as grade 1. KC eyes with a mean keratometry reading of greater than 48D and less than 53D were classified as grade 2. KC eyes with a mean keratometry reading of greater than 53D were classified as grade 3, with grade 3 being the most severe.
Patient-derived corneal epithelium
Epithelial cells were collected using the epithelial debridement procedure routinely performed during ocular surface surgeries, such as corneal collagen crosslinking or T-PRK in KC patients. Control epithelial cells were collected from subjects undergoing PRK who demonstrated no corneal surface distortions and had no clinical signs of KC. The corneal epithelium study cohort had a total size of 142 eyes, with 52 control and 90 KC subjects (Grade 1 = 44, grade 2 = 29, and grade 3 = 17) with a mean age of 22.7±5.7 years. Surgical procedures for all patients and controls were performed under topical anesthesia with a proparacaine 0.5% ophthalmic solution. Debrided epithelial cells were immediately transferred to −80 °C for storage until processing for RNA extraction.
Isolation of RNA, cDNA synthesis, and real-time PCR
The isolation of mRNA and the analysis of gene expressions have been described before [49]. Total RNA was extracted from patient epithelia using a TRIZOL reagent according to the manufacturer’s instructions (Invitrogen, Carlsbad, CA), followed by quantification and a quality assessment. In addition, cDNA was synthesized using a Superscript III (Life Technologies, Carlsbad, CA) cDNA conversion kit. Quantitative real-time PCR was performed as previously reported [50]. The quantitative real-time PCR cycle includes preincubation at 95 °C for 5 min, as well as 40 amplification cycles at 95 °C for 10 s, 60 °C for 15 s, and 72 °C for 30 s using a CFX ConnectTM real-time PCR detection system (Bio-Rad, Philadelphia, PA). The total levels of LOX, MMP9, IL6, COL IA1, and COL IVA1 were estimated after normalization to actin.
Patient tear collection
Tear samples were collected using capillary micropipette tubes, as described earlier [17]. The tears were collected from the exterior one-third of the lower fornix, taking care not to touch the conjunctiva or produce any reflex tearing. The tears were extracted from the micropipette tubes into sterile microfuge tubes followed by storage at −80 °C until analysis.
Lysyl oxidase activity assay
LOX activity in 20 µl tear samples from 30 control and 62 KC samples (grade 1 = 17, grade 2 = 17, grade 3 = 13, and grade 4 = 15) was measured using an Amplite™ Fluorimetric Lysyl Oxidase Assay Kit (AAT Bioquest Inc., Sunnyvale, CA) in a 96-well plate, as per the manufacturer’s instructions and as reported in the literature [51,52]. This kit and protocol were validated by the manufacturer using β-aminopropionitrile for the specific inhibition of LOX activity. Briefly, the fluorometric assay utilized a proprietary substrate that releases hydrogen peroxide using Amplite™ HRP substrate-coupled reactions to determine LOX activity. Enzyme activity was reported as relative fluorescence units (RFU/ml) from the mean of two independent readings. Blank wells without tear samples were used for determination of the baseline.
Immunohistochemical (IHC) studies
IHC studies have been reported in the literature for a variety of tissues, including KC corneas [49,53]. A cohort of 27 formalin-fixed, paraffin-embedded (FFPE) corneas from KC patients undergoing keratoplasty was obtained from the Narayana Nethralaya pathology department. In total, 15 unused donor corneas from individuals without clinical symptoms of KC were obtained from the Narayana Nethralaya Eye Bank to be used as normal controls for the IHC studies. All KC patient samples were classified as grade 4 KC. IHC experiments were performed using the Leica NovolinkTM Polymer Detection System (Leica Biosystems, Newcastle, UK, Cat # RE7159-K) as per the manufacturer’s protocols. Briefly, antigen retrieval was performed from 4-µm thick sections of FFPE tissue blocks in citrate buffer by boiling, followed by appropriate washing and blocking for endogenous peroxidase. Thereafter, primary antibodies for LOX (1:100 dilution; Merck Millipore, Billerca, MA; Cat # ABT112), collagen I (1:100 dilution; Novus biologic, Littleton, CO; Cat # NB600–408), and collagen IV (1:250 dilution; Abcam, Cambridge, MA; Cat # AB6586) were applied at 4 °C overnight, followed by washing and incubation with secondary antibodies as per the IHC kit directions. Counterstaining with hematoxylin was performed before the final substrate reaction step. Control and KC sample slides stained without a primary antibody were used as negative controls, and staining with an actin antibody was used as a positive control for the standardization of the IHC protocol. Photomicrographs were taken using a Leica Microsystems microscope and image acquisition software (DM 3000; LAS version 3.1.0). Protein expression levels were classified as high or low upon analysis of staining intensities (brown color) by two independent, blinded observers.
Statistical analysis
All data were compiled and grouped according to the stages of the diseases. The Friedman test for a comparison of group means was used. If the test was statistically significant, a post-hoc comparison of the group means was performed. The mean value of the individual groups was reported as mean ± standard error of mean (SEM). A linear regression analysis was performed among LOX, COL IA1, and COL IVA1. As the gene expression changed by a factor of 2 or more within the cohort, the expression data were converted to logarithmic values followed by a regression analysis. All statistical analyses were performed with MedCalc v12.5.0 (MedCalc Inc., Belgium) and GraphPad Prizm 6.0.
All continuous variables were assessed for normality of distribution. A one-way ANOVA was used to assess the difference between normal and KC grade means.
Results
Clinical analysis of patient study cohort
The study cohort clinical characteristics are summarized in Table 1. KC patients were graded into four groups numbered 1–4, while controls were labeled as grade 0. As expected, the gradation reflected the concordant increase in the corneal curvature measured by keratometry; the mean K values were grade 0: 44.7+0.25, grade 1: 44.2+0.22, grade 2: 47.3+0.18, and grade 3: 51.5+0.39. The corneal refraction spherical measurements were grade 1: −1.1+0.35, grade 2: −2.14+0.55, and grade 3: −3.67+0.56. Consequently, visual acuity (logMAR) was reduced across the grades, as follows: grade 1: 0.11+0.02, grade 2: 0.17+0.03, and grade 3: 0.23+0.03. As a function of grade (Table 1), the best corrected distance acuity (BCDA), sphere, cylinder, spherical equivalent, K1, K2, Kmean, central corneal thickness (CCT), and thinnest regions were significantly different (p<0.001). The BCDA of the control was significantly different from all KC grades (p<0.05). Within the KC grades, grade 1 was similar to grade 2 (p>0.05), but it was significantly different from grade 3 (p<0.05). The cylinders of the control and all the KC grades were significantly different from each other (p<0.05). The sphere of the control was similar to those of grades 2 and 3 (p>0.05), but it was significantly different from that of grade 1 (p<0.05). The spheres of grades 1 and 2 were significantly different from that of grade 3 (p<0.05). The spherical equivalents of the control and of grade 1 were similar (p>0.05), while the spherical equivalent of grade 3 was significantly different from those of all the grades and the control (p<0.05). K1, K2, and Kmean were significantly different among all the grades and the control (p<0.05). The CCT of the control was significantly different from those of all the KC grades (p<0.05), and the CCTs of grades 1 and 2 were similar (p>0.05), but they differed from that of grade 3 (p<0.05). The thinnest region showed a trend similar to CCT. The data expectedly showed a strong correlation of the clinical grades with the corneal curvature and vision loss. Therefore, this data establishes the basis for the stratification of gene expression levels from the same group of controls and KC patients. The various gene expression and LOX activity values are summarised with their corresponding significance values across the different grades of KC in Table 2.
Table 1. Summary statistics table of the patient cohort.
| Clinical observations | Control | Grade 1 | Grade 2 | Grade 3 | ANOVA p value |
|---|---|---|---|---|---|
| AGE |
24.59±0.44 |
25.46±1.19 |
21.11±0.94 |
21.84±0.93 |
0.06 |
| BCDA (logMAR) |
0.01±0.01 |
0.11±0.02 |
0.17±0.03 |
0.23±0.02 |
<0.001 |
| Sphere (D) |
−2.75±0.23 |
−1.09±0.35 |
−2.14±0.55 |
−3.67±0.56 |
<0.001 |
| Cylinder (D) |
−0.54±0.06 |
−2.08±0.33 |
−2.96±0.37 |
−4.43±0.36 |
<0.001 |
| K1 (D) |
44.25±0.24 |
42.97±0.28 |
45.76±0.18 |
48.92±0.37 |
<0.001 |
| K2 (D) |
45.25±0.26 |
45.67±0.30 |
49.0±0.33 |
54.43±0.50 |
<0.001 |
| Km (D) |
44.74±0.25 |
44.24±0.23 |
47.30±0.18 |
51.53±0.39 |
<0.001 |
| CCT (micrometer) |
519.20±3.42 |
490.2±5.21 |
479.0±6.0 |
455.9±4.63 |
<0.001 |
| Thinnest Point Thickness (micrometer) | 513.11±3.30 | 477.9±4.91 | 470.32±5.82 | 447.1±4.56 | <0.001 |
The table provides information about the corneal thickness based on pachymetry data from Pentacam, refraction measurements and mean values of gene expression data. VA: Visual acuity; Sphere and Cylinder: Spherical and Cylindrical refraction values; Pachy Apex: corneal thickness by pachymetry at the corneal apex; Pachy Thinnest: corneal thickness at the thinnest point of the cornea; K1, K2: Independent readings of corneal curvature by keratometry; Km and K-Max: Mean keratometry value and maximum keratometry value respectively. The ANOVA p value column shows group statistics; pairwise statistics are discussed in the results section.
Table 2. Summary statistics of the gene expression and LOX activity values in the patient cohort.
| Gene | Control | Grade 1 | Grade 2 | Grade 3 | ANOVA p value |
|---|---|---|---|---|---|
| MMP9 |
1.773±0.621 |
2.812±1.036 |
5.999±1.236 |
6.246±1.697 |
0.007 |
| Lox |
2.122±0.385 |
1.284±0.268 |
0.888±0.199 |
0.602±0.105 |
0.006 |
| IL6 |
6.488±1.534 |
6.444±2.063 |
6.68±1.742 |
7.405±1.482 |
0.98 |
| COL I AI |
24.21±5.549 |
14.325±5.726 |
12.813±4.579 |
8.245±2.153 |
0.12 |
| COL IV AI |
6.771±2.162 |
6.453±3.482 |
4.222±1.422 |
2.881±0.640 |
0.51 |
| Lox Activity | 6055±305 | 5627±512 | 4956±392 | 4753±382 | 0.04 |
The table provides the average values and SEM for the measured values for every gene from each grade of KC respectively. The ANOVA p value column shows group statistics; pairwise statistics are discussed in the results section.
Lysyl oxidase mRNA expression is reduced in KC patient corneal epithelia and correlates with clinical grade of disease
We addressed the question of whether the LOX expression in the corneal epithelium undergoes any change in KC. Figure 1A demonstrates that the LOX expression is significantly reduced in KC corneal epithelial cells compared to controls. Furthermore, when we stratified the entire cohort based on the clinical grades of KC, the LOX expression values were grade 0: 2.12 ± 0.39 grade 1: 1.28 ± 0.27, grade 2: 0.89 ± 0.20, and grade 3: 0.6 ± 0.1. We clearly observed a reducing trend for LOX mRNA levels across the grades (p = 0.006), suggesting it may play a role in disease progression.
Figure 1.
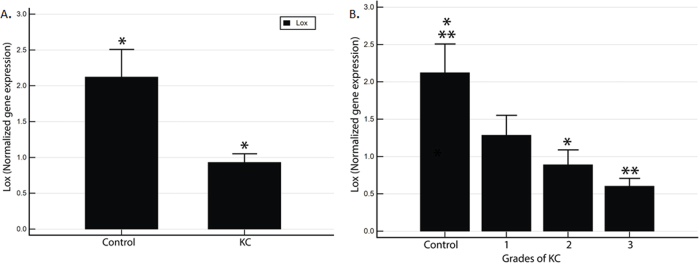
KC patients exhibit reduced LOX mRNA expression levels in corneal epithelial cells. Transcript levels were measured in debrided corneal epithelial cells from 52 control and 90 KC samples. Relative LOX gene expression in (A) control versus KC epithelium; (B) patients stratified by clinical grades of KC (Grade 0 = control).
Reduction of the collagen gene expression in KC corneas shows a correlation with disease severity
As the LOX expression is reduced in KC, we investigated the levels of the collagen I (COL IA1) and collagen IV (COL IVA1) transcripts (Figure 2) in the same patient cohort. We observed that both collagen I and IV transcript levels were reduced in KC corneas compared to controls (Figure 2A). Although collagen I shows a reducing trend across grades 1–4 (grade 0: 24.2+5.5 grade 1: 14.3+5.73, grade 2: 12.8+4.58, and grade 3: 8.2+2.15), we did not observe a uniform reduction (p = 0.12) among the higher grades (Figure 2B). The collagen IV expression also reduced with the clinical grade of KC (p = 0.77) (grade 0: 6.77+2.16 grade 1: 6.5+3.48, grade 2: 4.22+1.42, and grade 3: 2.8+0.64; Figure 2B). However, the collagen IV expression demonstrated a significant reduction in grades 3 and 4, while grade 1 does not show a reduction compared to the control and grade 2 shows a modest reduction (Figure 2B).
Figure 2.

The collagen gene expression is attenuated in the KC corneal epithelium. The relative gene expression values for COL IA1 and COL IVA1 were measured from 52 control and 90 KC corneal epithelium samples. The relative gene expression of (A) COL IA1 and COL IVA1 in the control versus KC; (B) COL IA1 and COL IVA1 mRNA levels stratified by clinical grades of KC. C: A linear regression analysis of the relative gene expression levels of LOX (y-axis) and COL IA1 (x-axis). Each circle represents an individual eye. D: A linear regression analysis of the relative gene expression levels of LOX (y-axis) and COL IVA1 (x-axis). Each circle represents individual eyes. E: A linear regression analysis of the relative gene expression levels of COLI A1 (y-axis) and COL IVA1 (x-axis). Each circle represents an individual eye.
The data suggests that along with the reduction in the LOX transcript levels, collagen expressions are also attenuated during KC disease progression. We conducted a statistical analysis using linear regression, as well as the correlation of the LOX expression with collagen I (Figure 2C) and collagen IV (Figure 2D) and of collagen I with collagen IV. The LOX expression demonstrated a strong positive correlation with collagen I (r = 0.35; p<0.0001). A strong positive correlation was also observed between LOX and collagen IV (r = 0.38; p<0.0001). Similarly, a strong positive correlation was also observed between collagen I and collagen IV (r = 0.72; p<0.0001).
KC corneal epithelium expresses high levels of MMP9
In the same cohort, the transcript levels of MMP9 were increased in the KC patients, while IL6 also showed a slight increase in KC patients over controls (Figure 3A). Figure 3B illustrates the differences in the gene expression of each MMP9 and IL6 marker when grouped according to the clinical disease severity of KC. MMP9 demonstrated a clear KC severity-dependent increase in expression (p = 0.007) (grade 0: 1.77+0.62 grade 1: 2.8+1.04, grade 2: 5.99+1.23, and grade 3: 6.2+1.69; Figure 3B). The IL6 expression, alternatively, showed a trend of increasing over the controls only in grades 2 and 3 (p = 0.98) (grade 0: 6.49+1.53 grade 1: 6.4+2.06, grade 2: 6.68+1.74, and grade 3: 7.4+1.48; Figure 3B).
Figure 3.
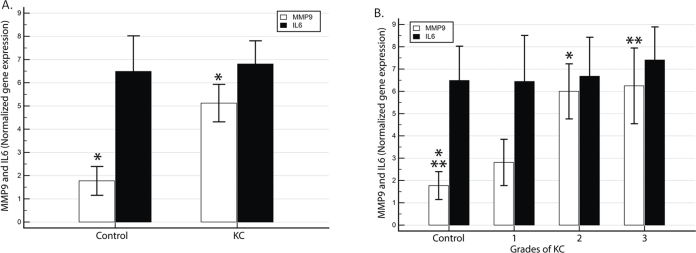
The MMP9 and IL6 gene expressions are elevated in KC, and the mRNA levels of MMP9 and IL6 were measured in the corneal epithelia of 52 control and 90 KC samples. The relative expression values for (A) MMP9 and IL6 in the control versus KC; (B) The MMP9 and IL6 mRNA levels stratified by clinical grades of KC.
Enzymatic activity of LOX in tears from KC patients is reduced compared to nonectatic controls
The corneal epithelium is known to secrete various factors in tears [54,55]. It has been reported that LOX enzymes are secreted by corneal epithelial cultures [41]. Therefore, we hypothesized that the human corneal epithelium may also secrete LOX, which could be analyzed in the tear film that bathes the cornea. We measured the total LOX activity in tear samples collected from the patients before their respective surgeries (Figure 4). Enzymatic activity was determined in 30 control and 62 KC patients using a commercially available assay kit that provides a functional readout in a fluorometric assay based on substrate oxidation. The total LOX activity was observed to be significantly reduced in the KC group (Figure 4A). There was a significant difference (p = 0.016) in the mean LOX activity between the control (6055 ± 16690 and KC (5039 ± 1928) groups. A reduction in LOX activity was also observed across KC grades (Figure 4B). When the control was compared with the grades of KC, there was a significant difference (p = 0.04) between the control and grade 3 (4753 ± 2021). However, the LOX activity of grades 1 (4956 ± 1619) and 2 (4753 ± 2021) were similar to the control (p>0.05). These data are reflective of the LOX transcript data (Figure 1B), suggesting the total LOX protein levels may be decreasing across different KC grades (Figure 4B).
Figure 4.
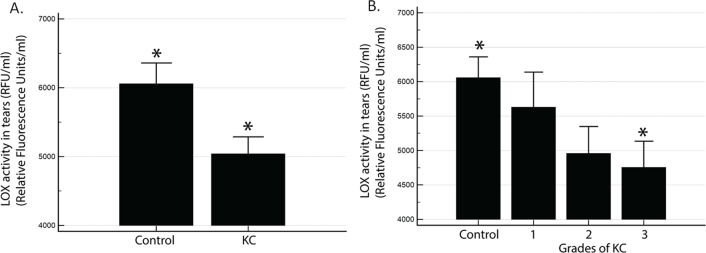
LOX activity is reduced in tears from KC patients. LOX activity in tears from 30 control and 62 KC samples is represented as relative fluorescence units/ml (RFU/ml) in (A) the control versus KC (*: p = 0.016 for KC activity compared to controls); B) different clinical grades of KC.
Expression of LOX protein is reduced in corneal epithelium of KC patients
To determine whether the LOX protein levels were indeed reduced in KC cornea, we analyzed LOX levels by IHC in KC patient corneal tissues (Figure 5). A cohort of 27 KC and 15 healthy donor corneas in FFPE blocks were stained with a LOX antibody. Figure 5A–C are representative photomicrographs of corneas from three individual KC patients. Similarly, Figure 5D–F are photomicrographs of three individual healthy donor control corneas. The LOX expression was observed in the corneal epithelial layer, but not in the basement membrane or stroma (as indicated by arrows). We observed that of the 27 KC corneas, 70% demonstrated highly reduced levels of LOX, while only 20% of the control donor corneas showed reductions in LOX expression. There was considerably more variability in the expression levels in KC samples compared to the controls, as evident in the three representative individuals in Figure 5 (compare A and C). As controls for IHC, we performed staining with actin as a positive control and no primary antibody as a negative control (Figure 6). Actin staining (brown color) was of similar intensity in both the control and KC corneas, while the negative controls did not provide any background brown color.
Figure 5.
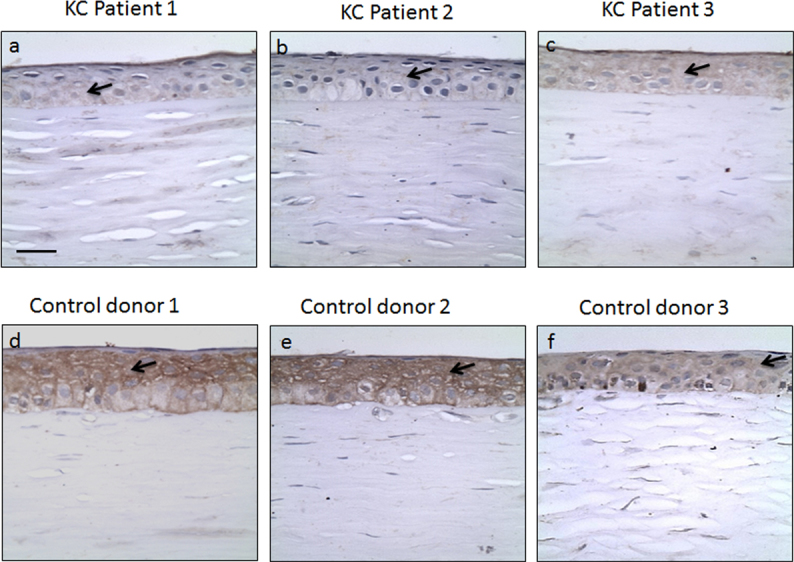
LOX protein levels are reduced in KC corneas. The representative photomicrographs show staining for LOX (Brown color) with counterstain hematoxylin (blue color) for nuclei. The staining is observed in the epithelial layer as indicated by the arrows. Three independent representative samples for KC and control corneas are presented at 40X magnification. Scale bar: 10 µm.
Figure 6.

Control staining for immunohistochemistry (IHC) experiments. Photomicrographs of control staining slides for IHC studies at 40X magnification; A-C: KC patient cornea; D-F: Control donor cornea. Actin staining (A and D); negative control without primary antibody, secondary antibody alone (B and E); hematoxylin and eosin (H & E) stain (C and F). Scale bar: 10 µm.
Collagen IV is significantly reduced in the corneal basement membrane of KC patients, while the collagen I protein expression is slightly reduced in the corneal stroma
We next determined the levels of collagen I and collagen IV in the same cohort of KC and control corneal tissue samples (Figure 7 and Figure 8, respectively). Photomicrographs of the same patient samples represented in Figure 5 are shown in both Figure 7 and Figure 8. The collagen I expression was observed across the entire corneal stroma in all samples (as reported earlier [20]), but it was slightly higher in the healthy controls (Figure 7D–F) compared to the KC patients (Figure 7A–C), as indicated by the arrows. A reduced collagen I staining intensity was observed in 33% of the KC corneas compared to 13% of the healthy donor controls. It should be noted that we observed an increased intensity of collagen I staining near the corneal basement membrane, which is known [56] (Figure 7D, F). However, some of the control corneas do not show the same increased intensity near the basement membrane (Figure 7E). The donor corneas are obtained from subjects that expired of natural causes and are typically older in age, which may have led to a reduction in condensed collagen fibrils in some cases, as reported earlier [56].
Figure 7.

Collagen I protein levels are reduced in KC corneas. The representative photomicrographs show staining for COL I (Brown color) with counterstain hematoxylin (blue color) for nuclei. The staining is observed in the corneal stroma, as indicated by the arrows. Three independent representative samples for KC and control corneas are presented at 40X magnification. Scale bar: 10 µm.
Figure 8.

Collagen IV protein levels are reduced in KC corneas. The representative photomicrographs show staining for COL IV (Brown color) with counterstain hematoxylin (blue color) for nuclei. The staining is observed in the basement membrane and immediate sub-epithelium, as indicated by the arrows. Three independent representative samples for KC and control corneas are presented at 40X magnification. Scale bar: 10 µm.
We observed a clear expression of the collagen IV protein in the epithelial basement membrane, as reported previously [57,58] (indicated in the figure by the arrows). The collagen IV expression was significantly reduced or absent in 81% of the KC patient samples (Figure 8A–C) compared to 26% in controls (Figure 8D–F). These data mirror the gene expression levels observed in Figures 1 and Figure 2.
Discussion
The role of collagen in maintaining the corneal structure is paramount [20,21,57,59]. Collagen-degrading enzymes, such as MMP9, or crosslinking enzymes, such as LOX, are reported to be deregulated in ocular disorders [30,39,41]. The formation of a fibrillar extracellular matrix through the oxidative linkage of collagen by LOX occurs in many tissue types, including the cornea [34]. A previous study suggested that the LOX expression is reduced in a small cohort of KC cornea tissues [41]. However, these data are opposed by gene expression microarray studies that suggest an upregulation of LOX in KC [42]. The reasons for the observed discrepancy in the two studies could be a cohort bias due to the small number of patients (8 eyes in the Dudakova et al. study and 10 eyes in Neilsen et al.), as well as due to the nature of the investigation (corneal tissue protein expression by immunofluorescent staining in Dudakova et al. and corneal epithelium mRNA measurement in Neilsen et al.). In addition, these studies did not analyze the expressions of the different contributing factors/genes in the corneal epithelia of KC patients to correlate their activities in disease. Furthermore, no specific association of such markers with the clinical severity of the disease was identified. Thus, our study sought to clarify the source and role of LOX, as well as the structural components of collagens I and IV and the inflammation-associated factors MMP9 and IL6 in a large cohort of Indian patients (142 eyes; 52 eyes from 29 control patients and 90 eyes from 80 KC patients). The correlation of the gene expressions of these factors with clinical severity will enhance our understanding of KC biology.
The LOX transcript levels were reduced in KC patients in our cohort (Figure 1A) and correlated with the significantly reduced enzymatic activity in patient tears (Figure 4A). When we measured the LOX gene expression across different grades, we observed a reducing trend across the grades, suggesting LOX levels may be a biomarker for KC grades (Figure 1B). The transcript levels also correlated clearly with the LOX activity assay in tear samples from KC patients (Figure 4). Furthermore, IHC staining for LOX in KC corneas demonstrated a significant reduction compared to healthy controls (Figure 5). The data therefore supports the observations by Dudakova et al., who found reduced levels of the LOX protein in most KC corneas. The gene expression data also suggest significant heterogeneity in the LOX levels, as observed by the wide range of expression values in each grade. This variation is also reflected in the IHC data set, suggesting that additional regulatory mechanisms apart from inflammation may be driving KC pathology. This variation may explain the observed increase in LOX mRNA levels observed in the Neilsen et al. report.
We also measured the expressions of collagen IA1 and collagen IVA1, which are required to maintain corneal structure (Figure 2). We observed that collagen levels were also reduced across the different grades of KC (Figure 2B). The linear regression analysis of gene expression correlations between LOX and collagens I and IV (Figure 2C, D, respectively) also suggest the expression levels of these genes are biologically linked and indicative of disease progression. Interestingly, while collagen IA1 was reduced in grade 1, collagen IVA1 was reduced in KC grade 3 (Figure 2). This suggests a possible regulatory network where collagen I is more important in the initial stages of the disease, while a reduction in collagen IV may be an effect of disease progression. Our IHC data for collagen IV appear to support this idea, as the expression of collagen IV in the KC basement membrane was drastically reduced compared to the controls (Figure 8). However, the data did not reveal whether the lack of collagen IV is due to the reduction in the Bowman’s layer associated with KC [60,61]. Conversely, it is plausible that the reduced production of collagen IV by the epithelium may cause the thinning of the basement membrane, thereby affecting the corneal structure and distorting vision. Collagen I was also mildly reduced in staining intensity in KC patient tissues compared to healthy donors (Figure 7). Positive staining of a similar intensity was observed for actin in both KC and controls, while no primary antibody staining served as the negative controls (Figure 6). The IHC data suggest the structural components of KC (LOX, collagen I, and collagen IV) are all reduced in grade 4 KC corneas, thereby rendering the corneas highly compromised for visual function. The data raise the possibility that a common molecular regulatory network exists for the genes that may be the real driver of KC. Alternatively, a reduction in LOX may trigger a reduced production of collagens through a signaling module hitherto unknown.
It has been reported that inflammatory cytokines and extracellular matrix-degrading enzymes are increased in the tears of KC patients in western populations [15-17,24]. In particular, studies by Lema et al. have shown a strong concordance of elevated IL6, TNFα, and MMP9 in tears from KC patients [16,17]. We demonstrate that the corneal epithelia in KC patients may be a source of these inflammatory factors (Figure 3). The inflammatory cytokine IL6 increases slightly in KC patients (Figure 3A), while the matrix-degrading enzyme and signaling protein MMP9 increases significantly (Figure 3A). One reason for the differential profile of IL6 could be that in both KC and controls, the epithelium is obtained intraoperatively by debridement, a procedure that may be causing an inflammatory stress response. Importantly, the concordance of increasing MMP9 levels with KC disease severity demonstrates that the corneal epithelium could indeed be an important source of these factors (Figure 3B).
Conclusions
In summary, we demonstrate for the first time in corneal epithelial cells of a large patient cohort that mRNA expression levels of the corneal structure-related genes collagen I, Collagen IV, and LOX are reduced in KC and correlate with disease severity. Consequently, both LOX activity and protein levels are decreased in KC, suggesting a crucial role for this enzyme in disease pathogenesis. The corresponding reduction in collagen protein levels has direct implications on corneal function, and it may be used as a biomarker of disease progression. A reduction in the expression of corneal structure-related genes correlates strongly with a concomitant increase in MMP9 levels, suggesting a possible regulatory signaling loop between these genes. The data strongly suggest that the regulation of the expression of these genes in the corneal epithelium should be explored as a treatment target for KC.
Acknowledgments
We would like to thank Dr. Chaitra Jaydev for critical review of the paper. We thank Krina Mihir, Shaniba Naizul and Devika Mahajan for technical assistance with laboratory experiments. Author contributions: Conceived and designed the experiments: AG, RS. Performed the experiments: AG, RAR, AS. Collected clinical samples, associated clinical data RS, VA, PR, NP. Analyzed the data: AG, ASR, AG, RS, RRM. Wrote the manuscript: RRM, AG. Funding: The authors would like to acknowledge funding by the Narayana Nethralaya Foundation for A.G. and Ruth M. Kraeuchi Missouri Endowed Chair Ophthalmology fund (RRM) for partial, support. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. Dr. Arkasubhra Ghosh (arkasubhra@narayananethralaya.com) and Dr Rajiv Mohan have contributed equally to this paper.
References
- 1.Sugar J, Macsai MS. What causes keratoconus? Cornea. 2012;31:716–9. doi: 10.1097/ICO.0b013e31823f8c72. [DOI] [PubMed] [Google Scholar]
- 2.Rabinowitz YS. Keratoconus. Surv Ophthalmol. 1998;42:297–319. doi: 10.1016/s0039-6257(97)00119-7. [DOI] [PubMed] [Google Scholar]
- 3.Zadnik K, Barr JT, Edrington TB, Everett DF, Jameson M, McMahon TT, Shin JA, Sterling JL, Wagner H, Gordon MO. Baseline findings in the Collaborative Longitudinal Evaluation of Keratoconus (CLEK) Study. Invest Ophthalmol Vis Sci. 1998;39:2537–46. [PubMed] [Google Scholar]
- 4.Piñero DP, Nieto JC, Lopez-Miguel A. Characterization of corneal structure in keratoconus. J Cataract Refract Surg. 2012;38:2167–83. doi: 10.1016/j.jcrs.2012.10.022. [DOI] [PubMed] [Google Scholar]
- 5.Romero-Jiménez M, Santodomingo-Rubido J, Wolffsohn JS. Keratoconus: a review. Cont Lens Anterior Eye. 2010;33:157–66. doi: 10.1016/j.clae.2010.04.006. [DOI] [PubMed] [Google Scholar]
- 6.Kenney MC, Nesburn AB, Burgeson RE, Butkowski RJ, Ljubimov AV. Abnormalities of the extracellular matrix in keratoconus corneas. Cornea. 1997;16:345–51. [PubMed] [Google Scholar]
- 7.Nichols JJ, Steger-May K, Edrington TB, Zadnik K. group Cs. The relation between disease asymmetry and severity in keratoconus. Br J Ophthalmol. 2004;88:788–91. doi: 10.1136/bjo.2003.034520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Korb DR, Finnemore VM, Herman JP. Apical changes and scarring in keratoconus as related to contact lens fitting techniques. J Am Optom Assoc. 1982;53:199–205. [PubMed] [Google Scholar]
- 9.Barr JT, Wilson BS, Gordon MO, Rah MJ, Riley C, Kollbaum PS, Zadnik K, Group CS. Estimation of the incidence and factors predictive of corneal scarring in the Collaborative Longitudinal Evaluation of Keratoconus (CLEK) Study. Cornea. 2006;25:16–25. doi: 10.1097/01.ico.0000164831.87593.08. [DOI] [PubMed] [Google Scholar]
- 10.Krachmer JH, Feder RS, Belin MW. Keratoconus and related noninflammatory corneal thinning disorders. Surv Ophthalmol. 1984;28:293–322. doi: 10.1016/0039-6257(84)90094-8. [DOI] [PubMed] [Google Scholar]
- 11.Jeyabalan N, Shetty R, Ghosh A, Anandula VR, Ghosh AS, Kumaramanickavel G. Genetic and genomic perspective to understand the molecular pathogenesis of keratoconus. Indian J Ophthalmol. 2013;61:384–8. doi: 10.4103/0301-4738.116055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Keating A, Pineda R, 2nd, Colby K. Corneal cross linking for keratoconus. Semin Ophthalmol. 2010;25:249–55. doi: 10.3109/08820538.2010.518503. [DOI] [PubMed] [Google Scholar]
- 13.Tuwairqi WS, Sinjab MM. Safety and efficacy of simultaneous corneal collagen cross-linking with topography-guided PRK in managing low-grade keratoconus: 1-year follow-up. J Refract Surg. 2012;28:341–5. doi: 10.3928/1081597X-20120316-01. [DOI] [PubMed] [Google Scholar]
- 14.Coskunseven E, Jankov MR, 2nd, Grentzelos MA, Plaka AD, Limnopoulou AN, Kymionis GD. Topography-guided transepithelial PRK after intracorneal ring segments implantation and corneal collagen CXL in a three-step procedure for keratoconus. J Refract Surg. 2013;29:54–8. doi: 10.3928/1081597X-20121217-01. [DOI] [PubMed] [Google Scholar]
- 15.Balasubramanian SA, Mohan S, Pye DC, Willcox MD. Proteases, proteolysis and inflammatory molecules in the tears of people with keratoconus. Acta Ophthalmol. 2012;90:e303–9. doi: 10.1111/j.1755-3768.2011.02369.x. [DOI] [PubMed] [Google Scholar]
- 16.Lema I, Duran JA. Inflammatory molecules in the tears of patients with keratoconus. Ophthalmology. 2005;112:654–9. doi: 10.1016/j.ophtha.2004.11.050. [DOI] [PubMed] [Google Scholar]
- 17.Lema I, Sobrino T, Duran JA, Brea D, Diez-Feijoo E. Subclinical keratoconus and inflammatory molecules from tears. Br J Ophthalmol. 2009;93:820–4. doi: 10.1136/bjo.2008.144253. [DOI] [PubMed] [Google Scholar]
- 18.Nemet AY, Vinker S, Bahar I, Kaiserman I. The association of keratoconus with immune disorders. Cornea. 2010;29:1261–4. doi: 10.1097/ICO.0b013e3181cb410b. [DOI] [PubMed] [Google Scholar]
- 19.Ghosh A, Zhou L, Ghosh A, Shetty R, Beuerman R. Proteomic and gene expression patterns of keratoconus. Indian J Ophthalmol. 2013;61:389–91. doi: 10.4103/0301-4738.116056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Nakayasu K, Tanaka M, Konomi H, Hayashi T. Distribution of types I, II, III, IV and V collagen in normal and keratoconus corneas. Ophthalmic Res. 1986;18:1–10. doi: 10.1159/000265406. [DOI] [PubMed] [Google Scholar]
- 21.Newsome DA, Foidart JM, Hassell JR, Krachmer JH, Rodrigues MM, Katz SI. Detection of specific collagen types in normal and keratoconus corneas. Invest Ophthalmol Vis Sci. 1981;20:738–50. [PubMed] [Google Scholar]
- 22.Yue BY, Sugar J, Schrode K. Histochemical studies of keratoconus. Curr Eye Res. 1988;7:81–6. doi: 10.3109/02713688809047024. [DOI] [PubMed] [Google Scholar]
- 23.Joseph R, Srivastava OP, Pfister RR. Differential epithelial and stromal protein profiles in keratoconus and normal human corneas. Exp Eye Res. 2011;92:282–98. doi: 10.1016/j.exer.2011.01.008. [DOI] [PubMed] [Google Scholar]
- 24.Srivastava OP, Chandrasekaran D, Pfister RR. Molecular changes in selected epithelial proteins in human keratoconus corneas compared to normal corneas. Mol Vis. 2006;12:1615–25. [PubMed] [Google Scholar]
- 25.Lackner EM, Matthaei M, Meng H, Ardjomand N, Eberhart CG, Jun AS. Design and analysis of keratoconus tissue microarrays. Cornea. 2014;33:49–55. doi: 10.1097/ICO.0000000000000012. [DOI] [PubMed] [Google Scholar]
- 26.Jun AS, Cope L, Speck C, Feng X, Lee S, Meng H, Hamad A, Chakravarti S. Subnormal cytokine profile in the tear fluid of keratoconus patients. PLoS ONE. 2011;6:e16437. doi: 10.1371/journal.pone.0016437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Jackson BC, Nebert DW, Vasiliou V. Update of human and mouse matrix metalloproteinase families. Hum Genomics. 2010;4:194–201. doi: 10.1186/1479-7364-4-3-194. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Malemud CJ. Matrix metalloproteinases (MMPs) in health and disease: an overview. Front Biosci. 2006;11:1696–701. doi: 10.2741/1915. [DOI] [PubMed] [Google Scholar]
- 29.Sobrin L, Liu Z, Monroy DC, Solomon A, Selzer MG, Lokeshwar BL, Pflugfelder SC. Regulation of MMP-9 activity in human tear fluid and corneal epithelial culture supernatant. Invest Ophthalmol Vis Sci. 2000;41:1703–9. [PubMed] [Google Scholar]
- 30.Ollivier FJ, Gilger BC, Barrie KP, Kallberg ME, Plummer CE, O'Reilly S, Gelatt KN, Brooks DE. Proteinases of the cornea and preocular tear film. Vet Ophthalmol. 2007;10:199–206. doi: 10.1111/j.1463-5224.2007.00546.x. [DOI] [PubMed] [Google Scholar]
- 31.Fan HX, Li HX, Chen D, Gao ZX, Zheng JH. Changes in the expression of MMP2, MMP9, and ColIV in stromal cells in oral squamous tongue cell carcinoma: relationships and prognostic implications. Journal of experimental & clinical cancer research. CR (East Lansing, Mich) 2012;31:90. doi: 10.1186/1756-9966-31-90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Finney J, Moon HJ, Ronnebaum T, Lantz M, Mure M. Human copper-dependent amine oxidases. Arch Biochem Biophys. 2014;546:19–32. doi: 10.1016/j.abb.2013.12.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Knott L, Bailey AJ. Collagen cross-links in mineralizing tissues: a review of their chemistry, function, and clinical relevance. Bone. 1998;22:181–7. doi: 10.1016/s8756-3282(97)00279-2. [DOI] [PubMed] [Google Scholar]
- 34.Kagan HM, Trackman PC. Properties and function of lysyl oxidase. Am J Respir Cell Mol Biol. 1991;5:206–10. doi: 10.1165/ajrcmb/5.3.206. [DOI] [PubMed] [Google Scholar]
- 35.Mäki JM, Kivirikko KI. Cloning and characterization of a fourth human lysyl oxidase isoenzyme. Biochem J. 2001;355:381–7. doi: 10.1042/0264-6021:3550381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Li W, Liu G, Chou IN, Kagan HM. Hydrogen peroxide-mediated, lysyl oxidase-dependent chemotaxis of vascular smooth muscle cells. J Cell Biochem. 2000;78:550–7. [PubMed] [Google Scholar]
- 37.Mäki JM. Lysyl oxidases in mammalian development and certain pathological conditions. Histol Histopathol. 2009;24:651–60. doi: 10.14670/HH-24.651. [DOI] [PubMed] [Google Scholar]
- 38.Cano A, Santamaria PG, Moreno-Bueno G. LOXL2 in epithelial cell plasticity and tumor progression. Future Oncol. 2012;8:1095–108. doi: 10.2217/fon.12.105. [DOI] [PubMed] [Google Scholar]
- 39.Coral K, Angayarkanni N, Madhavan J, Bharathselvi M, Ramakrishnan S, Nandi K, Rishi P, Kasinathan N, Krishnakumar S. Lysyl oxidase activity in the ocular tissues and the role of LOX in proliferative diabetic retinopathy and rhegmatogenous retinal detachment. Invest Ophthalmol Vis Sci. 2008;49:4746–52. doi: 10.1167/iovs.07-1550. [DOI] [PubMed] [Google Scholar]
- 40.Lucero HA, Kagan HM. Lysyl oxidase: an oxidative enzyme and effector of cell function. Cellular and molecular life sciences. Cell Mol Life Sci. 2006;63:2304–16. doi: 10.1007/s00018-006-6149-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Dudakova L, Liskova P, Trojek T, Palos M, Kalasova S, Jirsova K. Changes in lysyl oxidase (LOX) distribution and its decreased activity in keratoconus corneas. Exp Eye Res. 2012;104:74–81. doi: 10.1016/j.exer.2012.09.005. [DOI] [PubMed] [Google Scholar]
- 42.Nielsen K, Birkenkamp-Demtroder K, Ehlers N, Orntoft TF. Identification of differentially expressed genes in keratoconus epithelium analyzed on microarrays. Invest Ophthalmol Vis Sci. 2003;44:2466–76. doi: 10.1167/iovs.02-0671. [DOI] [PubMed] [Google Scholar]
- 43.Bisceglia L, De Bonis P, Pizzicoli C, Fischetti L, Laborante A, Di Perna M, Giuliani F, Delle Noci N, Buzzonetti L, Zelante L. Linkage analysis in keratoconus: replication of locus 5q21.2 and identification of other suggestive Loci. Invest Ophthalmol Vis Sci. 2009;50:1081–6. doi: 10.1167/iovs.08-2382. [DOI] [PubMed] [Google Scholar]
- 44.Bykhovskaya Y, Li X, Epifantseva I, Haritunians T, Siscovick D, Aldave A, Szczotka-Flynn L, Iyengar SK, Taylor KD, Rotter JI, Rabinowitz YS. Variation in the lysyl oxidase (LOX) gene is associated with keratoconus in family-based and case-control studies. Invest Ophthalmol Vis Sci. 2012;53:4152–7. doi: 10.1167/iovs.11-9268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Rao SN, Raviv T, Majmudar PA, Epstein RJ. Role of Orbscan II in screening keratoconus suspects before refractive corneal surgery. Ophthalmology. 2002;109:1642–6. doi: 10.1016/s0161-6420(02)01121-1. [DOI] [PubMed] [Google Scholar]
- 46.Ishii R, Kamiya K, Igarashi A, Shimizu K, Utsumi Y, Kumanomido T. Correlation of corneal elevation with severity of keratoconus by means of anterior and posterior topographic analysis. Cornea. 2012;31:253–8. doi: 10.1097/ICO.0B013E31823D1EE0. [DOI] [PubMed] [Google Scholar]
- 47.Miháltz K, Kovacs I, Takacs A, Nagy ZZ. Evaluation of keratometric, pachymetric, and elevation parameters of keratoconic corneas with pentacam. Cornea. 2009;28:976–80. doi: 10.1097/ICO.0b013e31819e34de. [DOI] [PubMed] [Google Scholar]
- 48.Piñero DP, Alio JL, Barraquer RI, Michael R, Jimenez R. Corneal biomechanics, refraction, and corneal aberrometry in keratoconus: an integrated study. Invest Ophthalmol Vis Sci. 2010;51:1948–55. doi: 10.1167/iovs.09-4177. [DOI] [PubMed] [Google Scholar]
- 49.Teo H, Ghosh S, Luesch H, Ghosh A, Wong ET, Malik N, Orth A, de Jesus P, Perry AS, Oliver JD, Tran NL, Speiser LJ, Wong M, Saez E, Schultz P, Chanda SK, Verma IM, Tergaonkar V. Telomere-independent Rap1 is an IKK adaptor and regulates NF-kappaB-dependent gene expression. Nat Cell Biol. 2010;12:758–67. doi: 10.1038/ncb2080. [DOI] [PubMed] [Google Scholar]
- 50.Ghosh A, Saginc G, Leow SC, Khattar E, Shin EM, Yan TD, Wong M, Zhang Z, Li G, Sung WK, Zhou J, Chng WJ, Li S, Liu E, Tergaonkar V. Telomerase directly regulates NF-kappaB-dependent transcription. Nat Cell Biol. 2012;14:1270–81. doi: 10.1038/ncb2621. [DOI] [PubMed] [Google Scholar]
- 51.López B, Gonzalez A, Lindner D, Westermann D, Ravassa S, Beaumont J, Gallego I, Zudaire A, Brugnolaro C, Querejeta R, Larman M, Tschope C, Diez J. Osteopontin-mediated myocardial fibrosis in heart failure: a role for lysyl oxidase? Cardiovasc Res. 2013;99:111–20. doi: 10.1093/cvr/cvt100. [DOI] [PubMed] [Google Scholar]
- 52.Vonk WI, de Bie P, Wichers CG, van den Berghe PV, van der Plaats R, Berger R, Wijmenga C, Klomp LW, van de Sluis B. The copper-transporting capacity of ATP7A mutants associated with Menkes disease is ameliorated by COMMD1 as a result of improved protein expression. Cellular and molecular life sciences. Cell Mol Life Sci. 2012;69:149–63. doi: 10.1007/s00018-011-0743-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Messmer EM, Meyer P, Herwig MC, Loeffler KU, Schirra F, Seitz B, Thiel M, Reinhard T, Kampik A, Auw-Haedrich C. Morphological and immunohistochemical changes after corneal cross-linking. Cornea. 2013;32:111–7. doi: 10.1097/ICO.0b013e31824d701b. [DOI] [PubMed] [Google Scholar]
- 54.Rao K, Farley WJ, Pflugfelder SC. Association between high tear epidermal growth factor levels and corneal subepithelial fibrosis in dry eye conditions. Invest Ophthalmol Vis Sci. 2010;51:844–9. doi: 10.1167/iovs.09-3875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Chotikavanich S, de Paiva CS. Li de Q, Chen JJ, Bian F, Farley WJ, Pflugfelder SC. Production and activity of matrix metalloproteinase-9 on the ocular surface increase in dysfunctional tear syndrome. Invest Ophthalmol Vis Sci. 2009;50:3203–9. doi: 10.1167/iovs.08-2476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Germundsson J, Karanis G, Fagerholm P, Lagali N. Age-related thinning of Bowman's layer in the human cornea in vivo. Invest Ophthalmol Vis Sci. 2013;54:6143–9. doi: 10.1167/iovs.13-12535. [DOI] [PubMed] [Google Scholar]
- 57.Cameron JD, Skubitz AP, Furcht LT. Type IV collagen and corneal epithelial adhesion and migration. Effects of type IV collagen fragments and synthetic peptides on rabbit corneal epithelial cell adhesion and migration in vitro. Invest Ophthalmol Vis Sci. 1991;32:2766–73. [PubMed] [Google Scholar]
- 58.Cleutjens JP, Havenith MG, Kasper M, Vallinga M, Bosman FT. Absence of type IV collagen in the centre of the corneal epithelial basement membrane. Histochem J. 1990;22:688–94. doi: 10.1007/BF01047454. [DOI] [PubMed] [Google Scholar]
- 59.Ishizaki M, Shimoda M, Wakamatsu K, Ogro T, Yamanaka N, Kao CW, Kao WW. Stromal fibroblasts are associated with collagen IV in scar tissues of alkali-burned and lacerated corneas. Curr Eye Res. 1997;16:339–48. doi: 10.1076/ceyr.16.4.339.10684. [DOI] [PubMed] [Google Scholar]
- 60.Abou Shousha M, Perez VL, Fraga Santini Canto AP, Vaddavalli PK, Sayyad FE, Cabot F, Feuer WJ, Wang J, Yoo SH. The use of Bowman's layer vertical topographic thickness map in the diagnosis of keratoconus. Ophthalmology. 2014;121:988–93. doi: 10.1016/j.ophtha.2013.11.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Yadav R, Kottaiyan R, Ahmad K, Yoon G. Epithelium and Bowman's layer thickness and light scatter in keratoconic cornea evaluated using ultrahigh resolution optical coherence tomography. J Biomed Opt. 2012;17:116010. doi: 10.1117/1.JBO.17.11.116010. [DOI] [PMC free article] [PubMed] [Google Scholar]