

Abstract
Background
Corona radiata cells (CRCs) refer to the fraction of cumulus cells just adjacent to the oocyte. The CRCs are closely connected to the oocyte throughout maturation and their gene expression profiles might reflect oocyte quality. Polycystic ovary syndrome (PCOS) is a common cause of infertility. It is controversial whether PCOS associate with diminished oocyte quality. The purpose of this study was to compare individual human CRC samples between PCOS patients and controls.
Methods
All patients were stimulated by the long gonadotropin-releasing hormone (GnRH) agonist protocol. The CRC samples originated from individual oocytes developing into embryos selected for transfer. CRCs were isolated in a two-step denudation procedure, separating outer cumulus cells from the inner CRCs. Extracted RNA was amplified and transcriptome profiling was performed with Human Agilent® arrays.
Results
The transcriptomes of CRCs showed no individual genes with significant differential expression between PCOS and controls, but gene set enrichment analysis identified several cell cycle- and DNA replication pathways overexpressed in PCOS CRCs (FDR < 0.05). Five of the genes contributing to the up-regulated cell cycle pathways in the PCOS CRCs were selected for qRT-PCR validation in ten PCOS and ten control CRC samples. qRT-PCR confirmed significant up-regulation in PCOS CRCs of cell cycle progression genes HIST1H4C (FC = 2.7), UBE2C (FC = 2.6) and cell cycle related transcription factor E2F4 (FC = 2.5).
Conclusion
The overexpression of cell cycle-related genes and cell cycle pathways in PCOS CRCs could indicate a disturbed or delayed final maturation and differentiation of the CRCs in response to the human chorionic gonadotropin (hCG) surge. However, this had no effect on the in vitro development of the corresponding embryos. Future studies are needed to clarify whether the up-regulated cell cycle pathways in PCOS CRCs have any clinical implications.
Electronic supplementary material
The online version of this article (doi:10.1186/s13048-014-0110-6) contains supplementary material, which is available to authorized users.
Keywords: Corona radiata cells, Transcriptome, Gene expression, PCOS, Oocyte quality
Background
Polycystic Ovary Syndrome (PCOS) is the most prevalent endocrine disorder of women in the reproductive age and represents a combination of polycystic ovaries, oligo/anovulation and hyperandrogenism [1]. The follicular microenvironment previously found to be altered in PCOS women might influence oocyte maturation and oocyte developmental competence [2,3]. Previous studies of PCOS women compared to healthy women revealed gene expression differences in metaphase ІІ (MII) oocytes [4], in cumulus cells of individual MІІ oocytes with unknown developmental potential [5] and pooled, cultured cumulus cells [6]. Ribosomal RNA content was increased in cumulus cells of PCOS women [7], which could indicate a higher rate of proliferation; also granulosa cells from PCOS women have been shown to be hyperproliferative [3]. These alterations may suggest an altered oocyte quality in PCOS patients compared to controls. However, they do not necessarily extrapolate to the clinical situation, where only oocytes developing into top quality embryos are used for transfer. A meta-analysis showed that PCOS women had a similar number of top quality embryos and similar rates of pregnancy and live births compared to healthy women undergoing in vitro fertilization (IVF), but PCOS patients had more oocytes retrieved and a significantly lower fertilization rate [8].
Corona radiata cells (CRCs) refer to the innermost layer of the cumulus cells, which is in direct contact with the zona pellucida of the oocyte. Throughout folliculogenesis and until the luteinizing hormone (LH) surge for final oocyte maturation, transzonal projections exist between the oocyte and the CRCs, allowing exchange of substances between the oocyte and the CRCs [9]. We hypothesize that transcriptomic analysis of CRCs would serve as a non-invasive method of gaining deeper understanding of the microenvironment of the oocyte. Since PCOS and non-PCOS women undergoing IVF had the same clinical outcome [8], we wanted to find out whether the transcriptomic profile of CRCs would differ between PCOS and controls in clinically relevant samples of CRCs from embryos chosen for transfer.
Materials and methods
This study was approved by The Danish Ethical Science Committee (SJ-156) and conducted in accordance with the Helsinki Declaration and all participants gave informed consent before inclusion in the study.
Study population
Ten women with PCOS and ten healthy, regularly cycling women without known disease (controls) were included. Exclusion criteria were diabetes type 1 or 2, impaired thyroid, renal or hepatic function, congenital adrenal hyperplasia, endometriosis, premature ovarian failure, hypothalamic amenorrhea or age >35 years.
Diagnosis of PCOS was made according to the Rotterdam Consensus Criteria [1]. For all control women, indication for intracytoplasmic sperm injection (ICSI) was a partner with infertility (defined as <5 million progressively moving spermatozoa/ml). For the PCOS women, half (5/10) of the couples were referred to ICSI because of male infertility and the rest after 4–6 failed attempts of intra-uterine insemination (IUI).
Baseline examination
Participants were included in the study based on a focused gynecological history and objective examination including transvaginal ultrasound of the ovaries and uterus. Blood samples were drawn after an overnight fast at 08.30-09.00 a.m. on cycle day (cd) 3–5 for regularly cycling women and on a random day for amenorrhoeic women. All androgen analyses were done at the same laboratory (Statens Serum Institut, SSI, Copenhagen, Denmark) in order to minimize variability. Total testosterone was measured by the CHS™ MSMS Steroids Kit (PerkinElmers®, Waltham, Massachusetts, USA) with intra-assay variation of 9.6% and inter-assay variation of 10.6%. Sex hormone-binding globulin (SHBG) was measured by Architect i2000 analyzer (Abbott®, Abbott Park, Illinois, USA) with an intra-assay variation of 2.8% and an inter-assay variation of 5.8%. Free testosterone was calculated from total testosterone and SHBG [10]. LH and follicle-stimulating hormone (FSH) were measured by immunoassay (LH: ref 11732234, FSH: ref 11775863, Roche Diagnostics, Mannheim, Germany).
Ovarian stimulation
Ovarian stimulation was achieved by the long gonadotropin-releasing hormone (GnRH) agonist protocol. Pituitary desensitization with buserelin 0.5 mg (Suprefact®, Sanofi-Aventis, Paris, France) was started on cd 21 in regularly menstruating women and at cd 15 for oligo/amenorrhoeic women starting with ethinylestradiol 30 mg/desogestrel 150 mg daily from the 1st day of bleeding (Marvelon®, Organon/Microgyn®, Bayer Pharma, Leverkusen, Germany) and until cd 21. Controlled ovarian stimulation with recombinant FSH (rFSH) (Puregon®, Organon, Oss, Netherlands) was started after at least 14 days of desensitization. Follicle growth was monitored by transvaginal ultrasound. Recombinant human chorionic gonadotropin (rhCG) 6500 IU (Ovitrelle®, Modugno, Italy) was administered when at least three follicles reached the size of 17 mm. Oocyte Pick-up (OPU) was performed 36 hours later under transvaginal ultrasound guidance. Luteal phase support (Lutinus®, Ferring©, Copenhagen, Denmark) was given from the day of transfer and until the pregnancy test. We adhered to the Danish National Criteria of elective single transfer for all patients <37 years of age. Transvaginal ultrasound was performed three weeks after a positive hCG blood test to confirm intrauterine clinical pregnancy.
Isolation of corona radiata cells, fertilization and time lapse incubation of embryos
Following OPU, cumulus-oocyte complexes were washed several times in Fertilization medium (Cook, Eight Mile Plains, Queensland, Australia) to remove cell debris, and incubated for two hours in Fertilization medium (Cook, Eight Mile Plains, Queensland, Australia). Then the oocytes were transferred to a droplet of Cleavage medium (Cook, Eight Mile Plains, Queensland, Australia) and denudated in a two-step procedure: First, the cumulus cells were removed by gentle pipetting in 20 μl Cumulase® (Origio, Måløv, Denmark) with a 1–10 μl Eppendorf Pipette using a Dual filter PCR clean 20 μl tip (Eppendorf, Hamburg, Germany). Then the oocyte with the remaining CRCs was transferred to 10 μl Cumulase® (Origio, Måløv, Denmark) and the CRCs were removed by gentle pipetting with a Denudation pipette 0.134-0.145 mm (Vitrolife, Göteborg, Sweden). Immediately after oocyte denudation, individual droplets containing the CRCs were transferred to DNA Lobind Eppendorf tubes (Eppendorf, Hamburg, Germany), snap frozen in liquid nitrogen and stored at −80°C until RNA extraction. MΙΙ oocytes were fertilized by ICSI within ten minutes after denudation, and incubated in individual wells in an Embryoscope® (Unisense Fertilitech, Aarhus, Denmark), which gave the opportunity to track the development of all fertilized oocytes until transfer (day 2), or, for the untransferred embryos, until vitrification as top quality blastocyst or disposal at day 5/6. Analysis of the images was done with the EmbryoViewer® Software (Unisense FertiliTech Aarhus, Denmark). The time from ICSI to the following events were annotated: Pronuclei breakdown (defined as the first picture frame where the pronuclei disappeared), 1st cleavage (defined as the first picture frame where the zygote turned into two cells) and cleavage to four cells (defined as the first picture frame where four cells were observed the first time).
Selection of CRC samples used in the study
The CRC samples used in this study came from oocytes developing into top quality embryos: All transferred embryos were top quality embryos according to the ALPHA/ESHRE consensus [11] scoring points in short: Four cells at day 2, low fragmentation (cut-off 25% fragmentation), cell cycle specific cell size, no multinucleation. Vitrified top quality blastocysts day 5/6 had score 3–6 AA/AB according to the blastocyst classification proposed by Gardner and Schoolcraft [12]. The fate of the ten oocytes corresponding to the CRC samples used were as follows: In the PCOS group, ten transferred embryos gave three clinical pregnancies, resulting in two live births and one missed abortion in gestational week 7 and seven negative hCG tests 14 days after oocyte retrieval; In the control group, three top quality blastocysts were vitrified for later use, and seven transferred embryos gave three clinical pregnancies with two live births and one missed abortion in gestational week 7.
RNA extraction and amplification
Total RNA was extracted by the RNAequeous® micro-kit from Ambion (Life Technologies, Paisley, UK) according to manufacturer’s instructions. The samples were analyzed for total RNA concentration by Qubit® (Life Technologies, Paisley, UK) and total RNA quality and level of degradation using an Agilent 2100 Bioanalyzer and RNA 6000 Pico LabChip according to the manufacturer’s instructions (Agilent Technologies, Waldbronn, Germany). All of the RNA samples showed two distinct peaks representing 18S and 28S rRNA, which indicated good quality RNA and presented RNA Integrity Number (RIN) from 6–9.4. Total RNA of 50 ng was amplified and converted into cDNA using the Ovation Pico WTA System V2 RNA Amplification System from NuGEN® Inc. (NuGEN®, San Carlos, California, USA).
Microarray experiment
cDNA was coupled to a Cyanine 3-dUTP fluorescent dye (Cy3) using the Oligonucleotide Array-Based CGH for Genomic DNA Analysis, Enzymatic Labeling for Blood, Cells or Tissues (protocol version 6.2, Agilent Technologies, Santa Clara, California, USA). Cy3-labeled cDNA was hybridized to Agilent Human Gene Expression Microarrays 4 × 44k v2 (G4845A) using the One-Color Microarray-Based Gene Expression Analysis, Quick Amp Labeling (version 5.7 protocol, Agilent Technologies, Santa Clara, California, USA) and scanned using an Agilent DNA Microarray scanner (Agilent Technologies, Santa Clara, California, USA). For microarray analysis, we used six CRC samples from PCOS women, and six from healthy controls (12 arrays in total).
Quantitative reverse-transcriptase PCR
The following TaqMan® Gene Expression Assays (pre-designed) (Applied Biosystems, Life Technologies Europe, Nærum, Denmark) were used (Assay ID-No: Hs00168719_m1 (Cyclophillin B/PPIB), Hs00171034_m1 (Cyclin T2, CCNT2), Hs00543883_s1 (histone cluster 1, H4c/HIST1H4C), Hs00608098_m1 (E2F transcription factor 4, p107/p130-binding/E2F4), and Hs00964100_g1 (ubiquitin-conjugating enzyme E2C/UBE2C)). Sample triplicates were prepared according to the manufacturer’s instructions. A total reaction volume of 20 μL was prepared on ice containing 10 μL TaqMan® Gene Expression Master Mix (2X), 1 μL TaqMan® Gene Expression Assay Mix (20X), 4 μL cDNA 5 ng/μl, and 5 μL RNase-free water. The samples were then centrifuged at 1,100 × g at 4°C for 5 minutes.
Gene expression was quantified using the MX3005 qPCR system (Agilent Technologies Denmark, Hørsholm, Denmark) under the following thermal cycling conditions: 95°C for 10 minutes, 40 cycles of 95°C for 15 seconds and 60°C for 1 minute. The data was normalized to cyclophillin (PPIB) [13], and relative quantification calculated according to the Comparative CT Method (ABI user bulletin # 2, 2001). qRT-PCR was performed on ten PCOS CRC samples and ten control CRC samples.
Gene expression microarray processing and analysis
Array quality was assessed with the arrayQualityMetrics R package [14], which used a variety of statistical tests combined with data visualization to mark outliers. Evaluation was done manually on a per array basis. Pre-processing of microarray data was done with the LIMMA software [15,16] available from the Bioconductor project [17]. Normalization between arrays was done by quantile normalization [18]. For genes with multiple probes, the intensity was defined as the median of all probes mapping to that gene.
Statistically significant differences between PCOS and control CRC samples in expression of individual transcripts were assessed by the LIMMA moderated t-test [16] using the Benjamin-Hochberg method of correction of P-values for multiple testing.
Statistical significance of biological themes was investigated for the entire dataset with the Gene Set Enrichment Analysis (GSEA) software version 2.0.12 (http://www.broadinstitute.org/gsea/index.jsp) [19,20]. In short, the GSEA algorithm ranked genes according to their expression level. By default, genes were ranked using the Signal-2-Noise metric, a more robust measure than both mean and median values, and very robust against outliers:
The enrichment of a pathway was assessed by walking through the list of ranked genes, incrementing a running sum score when encountering a gene found in the pathway, and decreasing it when encountering a gene not in the pathway. According to the authors, this corresponded to a weighted Kolmogorov-Smirnov-like statistic. Statistical significance was assessed by permutating the ranked list of genes a thousand times. Genes which were either poorly expressed, had high intra-variation in the group or had low variance between the PCOS and control group populated the middle of the ranked list, and thus did not contribute to the running sum score. We specifically investigated the pathways contained in the databases KEGG [21] and REACTOME [22]. Normalized enrichment score (NES) is defined as:
NES was reported together with the false discovery rate (FDR). We reported pathways with FDR ≤0.05.
All samples were MIAME compliant and were handled according to SOP in the microarray Center. The 12 arrays were submitted to ArrayExpress (http://www.ebi.ac.uk/arrayexpress/) at EMBL using MIAMExpress. The experiment accession number is E-MEXP-3985.
Statistics
Differences in baseline parameters between groups were tested by Mann–Whitney test (Graphpad Prism v. 6, San Diego, California, USA). Differences between groups in developmental timing of the embryos corresponding to the selected CRC samples were evaluated by Students t-test (Graphpad Prism v. 6, San Diego, California, USA). Differences between PCOS and control CRC samples in gene expression of selected genes in the qRT-PCR experiment were evaluated by Bayesian parameter estimation [23]. A comparison was considered significant if the 95% HDI (Highest Density Interval) did not contain the value 0.
Results
Baseline parameters of study groups
The PCOS and control group differed according to PCOS status. The PCOS group exhibited oligomenorrhea, polycystic ovaries and hyperandrogenaemia (Table 1). Age, BMI and basal FSH were similar across groups (Table 1). Embryo kinetic timings of the embryos corresponding to the selected CRC samples did not differ between groups (Table 1).
Table 1.
Baseline descriptive parameters of the study participants and the average developmental timings of the embryos corresponding to the CRC samples used in the study
| PCOS | Controls | p-value | |
|---|---|---|---|
| Age | 27,3±3,4 | 27,8±4 | ns |
| BMI | 24±4,8 | 22,4±3,5 | ns |
| No of antral follicles/ one plane | 15,8±5,4 | 8,2±1,7 | <0,0001 |
| No of menstrual bleedings/year | 1,8±3,2 | 12±0 | <0,0001 |
| Total testosterone | 2,8±1,1 | 1,2±0,3 | 0,0003 |
| Free testosterone | 0,03931±0,02532 | 0,014±0,0042 | 0,0029 |
| SHBG | 78±26 | 79±21 | ns |
| LH/FSH | 1,6±0,8 | 0,8±0,3 | 0,03 |
| Time of 2PN breakdown (h) | 23.2±3.8 | 21.8±2.4 | ns |
| Time of 1st Cleavage (h) | 26±4.1 | 24.3±2.5 | ns |
| Time of cleavage to 4 cells (h) | 38.3±5 | 36.4±4.2 | ns |
p-value <0,05 considered significant. Not significant = ns.
Quality of the arrays
There were no apparent outliers based on the arrayQualityMetrics reports.
Differentially expressed genes
After correction for multiple testing, no individual genes in the microarray experiment showed significant differential expression between PCOS CRC samples and control CRC samples (Additional file 1).
Gene set enrichment analysis
GSEA showed upregulation of 24 pathways with FDR < 0.05 in PCOS CRCs compared to control CRCs (Table 2). Especially pathways involved in cell cycle and DNA replication were up-regulated in PCOS CRCs (Table 2). Additional file 2 shows the full pathway list with the genes contributing to the up-regulation of these pathways in PCOS CRC samples.
Table 2.
Gene set enrichment analysis of the transcriptome of CRCs of individual oocytes developing into embryos selected for transfer
| Pathway | No of genes enriched/total number of genes in the pathway | FDR | NES |
|---|---|---|---|
| Reactome Cell Cycle | 127/421 | 0.001 | 0.42 |
| Reactome G2 M Checkpoints | 17/45 | 0.001 | 0.61 |
| KEGG DNA Replication | 20/36 | 0.001 | 0.65 |
| Reactome Meiotic Recombination | 35/86 | 0.001 | 0.59 |
| Reactome Packaging of Telomere Ends | 25/48 | 0.001 | 0.63 |
| Reactome Activation of the Pre Replicative Complex | 14/31 | 0.001 | 0.60 |
| Reactome RNA POL Ι RNA POL ΙΙΙ and Mitochondrial Transcription | 45/122 | 0.001 | 0.53 |
| Reactome RNA POL Ι Transcription | 35/89 | 0.001 | 0.55 |
| Reactome RNA POL Ι Promotor Opening | 32/62 | 0.001 | 0.64 |
| Reactome Activation of ATR in Response to Replication Stress | 15/38 | 0.001 | 0.63 |
| Reactome Deposition of new CENPA containing Nucleosomes at the Centromere | 34/64 | 0.001 | 0.60 |
| Reactome DNA strand Elongation | 20/30 | 0.001 | 0.69 |
| Reactome Telomere Maintenance | 39/75 | 0.001 | 0.61 |
| Reactome Cell Cycle Mitotic | 94/325 | 0.01 | 0.41 |
| Reactome Meiosis | 45/116 | 0.01 | 0.49 |
| Reactome Amyloids | 34/83 | 0.01 | 0.51 |
| Reactome lagging strand synthesis | 12/19 | 0.01 | 0.69 |
| Reactome Mitotic M M G1 Phases | 55/137 | 0.03 | 0.42 |
| Reactome Mitotic Prometaphase | 32/87 | 0.03 | 0.47 |
| Reactome DNA replication | 56/192 | 0.03 | 0.42 |
| Reactome Meiotic Synapsis | 34/73 | 0.03 | 0.50 |
| Reactome Chromosome Maintenance | 55/122 | 0.03 | 0.42 |
| Reactome Extension of Telomeres | 13/27 | 0.03 | 0.59 |
| Reactome Transcription | 61/210 | 0.04 | 0.41 |
PCOS women compared controls (n = 6 PCOS arrays vs n = 6 control arrays). Enriched pathways with FDR <0.05 were listed. NES = normalized enrichment score. All enriched pathways were upregulated in PCOS CRC samples.
Quantitative reverse transcriptase PCR
Five of the genes contributing to the up-regulated cell cycle pathways in the PCOS CRCs were selected for qRT-PCR validation. We selected genes with different functions in the cell cycle. Gene expression by qRT-PCR were in accordance with the microarray data for three out of five genes tested (Figure 1). According to the qRT-PCR results, PCOS CRC samples showed significant up-regulation of HIST1H4C (FC = 2.7, 95% HDI 0.436-4.49), UBE2C (FC (Fold Change) = 2.6, 95% HDI 0.00438-3.21) and E2F4 (FC = 2.5, 95% HDI 0.571-5.33). CCND2 and CCNT2 showed equal expression between groups in the qRT-PCR experiment while the microarray data showed a 1.4 higher expression in the PCOS CRC samples (Figure 1).
Figure 1.
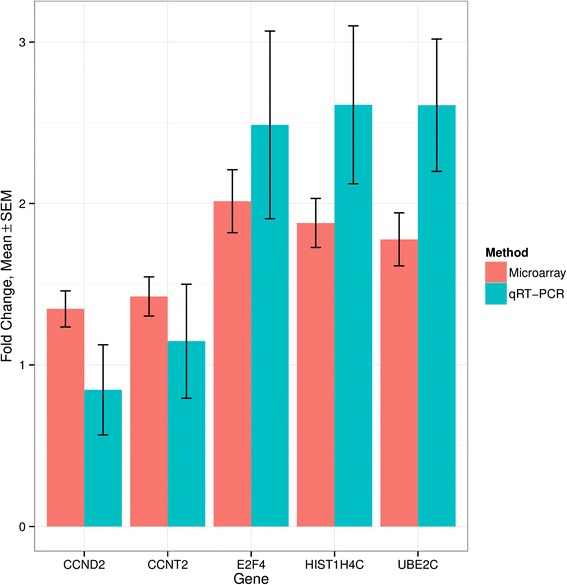
Comparison between microarray and qRT-PCR results for 5 selected genes in PCOS and control CRC samples. Fold change (PCOS/controls) ± Standard Error of the Mean (SEM). Red bars represented microarray results and blue bars represented qRT-PCR results. Microarray experiment: 6 PCOS CRC arrays vs. 6 control CRC arrays. qRT-PCR experiment: 10 PCOS CRC samples vs. 10 control CRC samples. The up-regulation of UBE2C, HIST1H4C and E2F4 in PCOS CRCs found in the microarray experiment was in line with the qRT-PCR results, whereas the 1.4 fold up-regulation of CCND2 and CCNT2 in PCOS CRCs found in the microarray experiment could not be confirmed by qRT-PCR.
Discussion
In this study, we present for the first time a transcriptomic analysis of individual human CRC samples from oocytes used for transfer or blastocyst vitrification in PCOS and controls. We did not find any significantly differentially expressed individual genes after correction for multiple testing in the present microarray study of PCOS CRCs and control CRCs from MІІ oocytes developing into top quality embryos. This is in contrast to the study by Haouzi et al. [5] who found 3,700 significantly differentially expressed genes between PCOS and control cumulus cells from MІІ oocytes of unspecified developmental potential with the same number of arrays per group as in the present study (six PCOS vs. six control arrays). However, the apparent inconsistency between the present study and the study by Haouzi et al. [5] could be explained by the different study designs and experimental approaches: Firstly, we used enzymatically isolated CRCs, which constitute the innermost cells closest to the oocyte, whereas Haouzi et al. [5] used mechanically cut cumulus cells. Several previous studies have successfully used enzymatically isolated cumulus cells [24–29] and CRCs [30,31] to investigate gene expression differences according to e.g. oocyte quality. Cumulase® used for denudation is a pure, recombinant and specific hyaluronidase and we would not expect the short exposure to mask gene expression differences. To the best of our knowledge, no studies have investigated gene expression differences according to isolation procedure of cumulus cells. Secondly, we used CRC samples from MІІ oocytes capable of developing into top quality embryos, whereas in former studies on gene expression in cumulus cells from PCOS patients [5,6], the developmental potential of the oocytes was not tracked. The strict use of CRCs from MІІ oocytes developing into top quality embryos in the present microarray study might explain why there were no individual genes with significant differential expression between PCOS and controls. Previous studies have shown that the transcriptome of cumulus cells from MІІ oocytes of poor developmental competence differed from cumulus cells of MІІ oocytes with high developmental competence [24,26,29,31–36]. These previous findings underlined the importance of oocyte selection, when comparing cumulus cells or CRCs in relation to a certain condition, such as PCOS.
During the study period, 10% (5/51) of women diagnosed with PCOS did not develop embryos for transfer or blastocyst vitrification. In the present study, we did not examine CRCs from this minority of the PCOS patients. This might have led to an underestimation of the differences in CRC transcriptomic profile between PCOS patients and controls since all controls developed embryos useful for transfer during the study period. Nevertheless, our data were applicable to the majority of the PCOS patients who developed embryos suitable for transfer: Transcriptomic aberrations found in the cumulus cells of MІІ oocytes of unknown developmental potential from PCOS women [5] or in the cumulus cells of the entire oocyte pool from PCOS women [6] did not extrapolate to the transcriptome of CRCs of MІІ oocytes developing into embryos used for transfer or blastocyst vitrification. This indicated that the microenvironment constituted by the CRCs did not differ substantially between PCOS and controls for oocytes with good developmental potential and it is in line with previous findings of similar implantation and pregnancy rates in PCOS patients and non-PCOS patients undergoing IVF [8].
The minority of PCOS patients with no embryos suitable for transfer constituted a clinically interesting subgroup with clearly impaired oocyte quality, and future studies on oocyte quality in PCOS should focus on this subgroup as a distinct entity within the PCOS population.
The GSEA showed up-regulation of pathways involved in cell cycle and DNA replication in PCOS CRCs. This is in line with previous studies showing hyperproliferative cumulus [7] and granulosa [3] cells in PCOS. The LH/hCG surge for final oocyte maturation dramatically down-regulate cell cycle genes in human granulosa cells [37] as well as in rodent cumulus-oocyte-complexes [38]. In agreement with this, cell cycle pathways were up-regulated in human cumulus cells of MІ oocytes compared to cumulus cells of MІІ oocytes [39]. We speculate that the up-regulation of cell cycle pathways in PCOS CRCs could be an indicator of disturbed or delayed final maturation of the cumulus cells/CRCs in response to the LH/hCG trigger.
The three cell cycle-related genes which showed significant up-regulation in PCOS CRCs represented different aspects of cell cycle: HIST1H4C is a replication-dependent histone exclusively transcribed during the S-phase of the cell cycle [40,41], UBE2C is required for destruction of mitotic cyclins and cell cycle progression [42], and E2F4 is a transcription factor and exerted a range of functions, mainly in cell cycle, DNA repair, ubiquitination and stress response pathways [43]. In light of the multitude of regulatory functions exerted by E2F4, the significant 2.5 fold up-regulation in PCOS CRCs according to the qRT-PCR experiment might have interesting biological implications, which should be explored in future studies.
Conclusion
The transcriptomic analysis of CRCs from oocytes developing into embryos selected for transfer showed up-regulation of cell cycle pathways and DNA replication pathways in PCOS CRC samples, however, this had no detectable effect on in vitro development of the corresponding embryos. Future studies are needed to clarify whether the up-regulated cell cycle pathways in PCOS CRCs have any clinical implications.
Acknowledgements
We thank the staff in the IVF lab of Holbæk Fertility Clinic for their assistance in isolating corona cell samples. The study was supported by a grant from Nordic InfuCare.
Additional files
Comparison of the corona radiata cell transcriptome between PCOS and controls (6 PCOS arrays vs. 6 control arrays). Full gene list showing Log fold change and adjusted p-values.
Comparison of the corona radiata cell transcriptome between PCOS and controls (6 PCOS arrays vs. 6 control arrays). Here is the full list of genes contributing to the upregulation of cell cycle pathways in PCOS corona radiata cells. Mean intensity and fold change of each gene is shown.
Footnotes
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
MLWI planned the study, recruited patients, collected materials, extracted RNA, performed qRT-PCR and wrote the manuscript. SBS extracted RNA and revised the manuscript. DW and KB performed the bio-informatical analyses and revised the manuscript. KDN performed the microarray experiment. THT planned the study and helped with collecting materials. ALM planned the study and revised the manuscript. All authors have read and approved the final version of the manuscript.
Contributor Information
Marie Louise Wissing, Email: mlwi@regionsjaelland.dk.
Si Brask Sonne, Email: ssonne@bio.ku.dk.
David Westergaard, Email: David@harsk.dk.
Kho do Nguyen, Email: khodo@cbs.dtu.dk.
Kirstine Belling, Email: belling@cbs.dtu.dk.
Thomas Høst, Email: tht@regionsjaelland.dk.
Anne Lis Mikkelsen, Email: aleg@regionsjaelland.dk.
References
- 1.Rotterdam ESHRE/ASRM-Sponsored PCOS Consensus Workshop Group Revised 2003 consensus on diagnostic criteria and long-term health risks related to polycystic ovary syndrome. Fertil Steril. 2004;81:19–25. doi: 10.1016/j.fertnstert.2003.10.004. [DOI] [PubMed] [Google Scholar]
- 2.Desforges-Bullet V, Gallo C, Lefebvre C, Pigny P, Dewailly D, Catteau-Jonard S. Increased anti-Müllerian hormone and decreased FSH levels in follicular fluid obtained in women with polycystic ovaries at the time of follicle puncture for in vitro fertilization. Fertil Steril. 2010;94:198–204. doi: 10.1016/j.fertnstert.2009.03.004. [DOI] [PubMed] [Google Scholar]
- 3.Stubbs SA, Stark J, Dilworth SM, Franks S, Hardy K. Abnormal preantral folliculogenesis in polycystic ovaries is associated with increased granulosa cell division. J Clin Endocrinol Metab. 2007;92:4418–4426. doi: 10.1210/jc.2007-0729. [DOI] [PubMed] [Google Scholar]
- 4.Wood JR, Dumesic DA, Abbott DH, Strauss JF. Molecular abnormalities in oocytes from women with polycystic ovary syndrome revealed by microarray analysis. J Clin Endocrinol Metab. 2007;92:705–713. doi: 10.1210/jc.2006-2123. [DOI] [PubMed] [Google Scholar]
- 5.Haouzi D, Assou S, Monzo C, Vincens C, Dechaud H, Hamamah S. Altered gene expression profile in cumulus cells of mature MII oocytes from patients with polycystic ovary syndrome. Hum Reprod. 2012;27:3523–3530. doi: 10.1093/humrep/des325. [DOI] [PubMed] [Google Scholar]
- 6.Kenigsberg S, Bentov Y, Chalifa-Caspi V, Potashnik G, Ofir R, Birk OS. Gene expression microarray profiles of cumulus cells in lean and overweight-obese polycystic ovary syndrome patients. Mol Hum Reprod. 2009;15:89–103. doi: 10.1093/molehr/gan082. [DOI] [PubMed] [Google Scholar]
- 7.Polzikov M, Yakovenko S, Voznesenskaya J, Troshina M, Zatsepina O. Overexpression of ribosomal RNA in cumulus cells of patients with polycystic ovary syndrome. J Assist Reprod Genet. 2012;29:1141–1145. doi: 10.1007/s10815-012-9827-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Heijnen EMEW, Eijkemans MJC, Hughes EG, Laven JSE, Macklon NS, Fauser BCJM. A meta-analysis of outcomes of conventional IVF in women with polycystic ovary syndrome. Hum Reprod Update. 2006;12:13–21. doi: 10.1093/humupd/dmi036. [DOI] [PubMed] [Google Scholar]
- 9.Albertini DF, Combelles CM, Benecchi E, Carabatsos MJ. Cellular basis for paracrine regulation of ovarian follicle development. Reproduction (Cambridge, England) 2001;121:647–653. doi: 10.1530/rep.0.1210647. [DOI] [PubMed] [Google Scholar]
- 10.Vermeulen A, Verdonck L, Kaufman JM. A critical evaluation of simple methods for the estimation of free testosterone in serum. J Clin Endocrinol Metab. 1999;84:3666–3672. doi: 10.1210/jcem.84.10.6079. [DOI] [PubMed] [Google Scholar]
- 11.Alpha Scientists in Reproductive Medicine and ESHRE Special Interest Group of Embryology The Istanbul consensus workshop on embryo assessment: proceedings of an expert meeting. Hum Reprod. 2011;26:1270–1283. doi: 10.1093/humrep/der037. [DOI] [PubMed] [Google Scholar]
- 12.Gardner DK, Schoolcraft WB. Culture and transfer of human blastocysts. Curr Opin Obstet Gynecol. 1999;11:307–311. doi: 10.1097/00001703-199906000-00013. [DOI] [PubMed] [Google Scholar]
- 13.Teixeira Filho FL, Baracat EC, Lee TH, Suh CS, Matsui M, Chang RJ, Shimasaki S, Erickson GF. Aberrant expression of growth differentiation factor-9 in oocytes of women with polycystic ovary syndrome. J Clin Endocrinol Metab. 2002;87:1337–1344. doi: 10.1210/jcem.87.3.8316. [DOI] [PubMed] [Google Scholar]
- 14.Kauffmann A, Gentleman R, Huber W. ArrayQualityMetrics--a bioconductor package for quality assessment of microarray data. Bioinformatics (Oxford, England) 2009;25:415–416. doi: 10.1093/bioinformatics/btn647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Bolstad BM, Irizarry RA, Astrand M, Speed TP. A comparison of normalization methods for high density oligonucleotide array data based on variance and bias. Bioinformatics (Oxford, England) 2003;19:185–193. doi: 10.1093/bioinformatics/19.2.185. [DOI] [PubMed] [Google Scholar]
- 16.Smyth GK. Linear models and empirical bayes methods for assessing differential expression in microarray experiments. Stat Appl Genet Mol Biol. 2004;3:Article3. doi: 10.2202/1544-6115.1027. [DOI] [PubMed] [Google Scholar]
- 17.Gentleman RC, Carey VJ, Bates DM, Bolstad B, Dettling M, Dudoit S, Ellis B, Gautier L, Ge Y, Gentry J, Hornik K, Hothorn T, Huber W, Iacus S, Irizarry R, Leisch F, Li C, Maechler M, Rossini AJ, Sawitzki G, Smith C, Smyth G, Tierney L, Yang JYH, Zhang J. Bioconductor: open software development for computational biology and bioinformatics. Genome Biol. 2004;5:R80. doi: 10.1186/gb-2004-5-10-r80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Smyth GK, Speed T. Normalization of cDNA microarray data. Methods (San Diego, Calif) 2003;31:265–273. doi: 10.1016/S1046-2023(03)00155-5. [DOI] [PubMed] [Google Scholar]
- 19.Subramanian A, Tamayo P, Mootha VK, Mukherjee S, Ebert BL, Gillette MA, Paulovich A, Pomeroy SL, Golub TR, Lander ES, Mesirov JP. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc Natl Acad Sci U S A. 2005;102:15545–15550. doi: 10.1073/pnas.0506580102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Mootha VK, Lindgren CM, Eriksson K-F, Subramanian A, Sihag S, Lehar J, Puigserver P, Carlsson E, Ridderstråle M, Laurila E, Houstis N, Daly MJ, Patterson N, Mesirov JP, Golub TR, Tamayo P, Spiegelman B, Lander ES, Hirschhorn JN, Altshuler D, Groop LC. PGC-1alpha-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat Genet. 2003;34:267–273. doi: 10.1038/ng1180. [DOI] [PubMed] [Google Scholar]
- 21.Kanehisa M, Goto S, Sato Y, Furumichi M, Tanabe M. KEGG for integration and interpretation of large-scale molecular data sets. Nucleic Acids Res. 2012;40(Database issue):D109–D114. doi: 10.1093/nar/gkr988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Matthews L, Gopinath G, Gillespie M, Caudy M, Croft D, de Bono B, Garapati P, Hemish J, Hermjakob H, Jassal B, Kanapin A, Lewis S, Mahajan S, May B, Schmidt E, Vastrik I, Wu G, Birney E, Stein L, D’Eustachio P. Reactome knowledgebase of human biological pathways and processes. Nucleic Acids Res. 2009;37(Database issue):D619–D622. doi: 10.1093/nar/gkn863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Kruschke JK. Bayesian estimation supersedes the t test. J Exp Psychol Gen. 2013;142:573–603. doi: 10.1037/a0029146. [DOI] [PubMed] [Google Scholar]
- 24.Anderson RA, Sciorio R, Kinnell H, Bayne RAL, Thong KJ, de Sousa PA, Pickering S. Cumulus gene expression as a predictor of human oocyte fertilisation, embryo development and competence to establish a pregnancy. Reproduction (Cambridge, England) 2009;138:629–637. doi: 10.1530/REP-09-0144. [DOI] [PubMed] [Google Scholar]
- 25.Adriaenssens T, Segers I, Wathlet S, Smitz J. The cumulus cell gene expression profile of oocytes with different nuclear maturity and potential for blastocyst formation. J Assist Reprod Genet. 2011;28:31–40. doi: 10.1007/s10815-010-9481-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Feuerstein P, Puard V, Chevalier C, Teusan R, Cadoret V, Guerif F, Houlgatte R, Royere D. Genomic assessment of human cumulus cell marker genes as predictors of oocyte developmental competence: impact of various experimental factors. PLoS One. 2012;7:e40449. doi: 10.1371/journal.pone.0040449. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Wathlet S, Adriaenssens T, Segers I, Verheyen G, Janssens R, Coucke W, Devroey P, Smitz J. New candidate genes to predict pregnancy outcome in single embryo transfer cycles when using cumulus cell gene expression. Fertil Steril. 2012;98(2):432–439. doi: 10.1016/j.fertnstert.2012.05.007. [DOI] [PubMed] [Google Scholar]
- 28.Ekart J, McNatty K, Hutton J, Pitman J. Ranking and selection of MII oocytes in human ICSI cycles using gene expression levels from associated cumulus cells. Hum Reprod. 2013;28:2930–2942. doi: 10.1093/humrep/det357. [DOI] [PubMed] [Google Scholar]
- 29.Wathlet S, Adriaenssens T, Segers I, Verheyen G, Van Landuyt L, Coucke W, Devroey P, Smitz J. Pregnancy prediction in single embryo transfer cycles after ICSI using QPCR: validation in oocytes from the same cohort. PLoS One. 2013;8:e54226. doi: 10.1371/journal.pone.0054226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.May-Panloup P, Ferré-L’Hôtellier V, Morinière C, Marcaillou C, Lemerle S, Malinge M-C, Coutolleau A, Lucas N, Reynier P, Descamps P, Guardiola P. Molecular characterization of corona radiata cells from patients with diminished ovarian reserve using microarray and microfluidic-based gene expression profiling. Hum Reprod. 2012;27:829–843. doi: 10.1093/humrep/der431. [DOI] [PubMed] [Google Scholar]
- 31.Fragouli E, Wells D, Iager AE, Kayisli UA, Patrizio P. Alteration of gene expression in human cumulus cells as a potential indicator of oocyte aneuploidy. Hum Reprod. 2012;27:2559–2568. doi: 10.1093/humrep/des170. [DOI] [PubMed] [Google Scholar]
- 32.Ouandaogo ZG, Haouzi D, Assou S, Dechaud H, Kadoch IJ, De Vos J, Hamamah S. Human cumulus cells molecular signature in relation to oocyte nuclear maturity stage. PLoS One. 2011;6:e27179. doi: 10.1371/journal.pone.0027179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Van Montfoort APA, Dumoulin JCM, Kester ADM, Evers JLH. Early cleavage is a valuable addition to existing embryo selection parameters: a study using single embryo transfers. Hum Reprod. 2004;19:2103–2108. doi: 10.1093/humrep/deh385. [DOI] [PubMed] [Google Scholar]
- 34.Gebhardt KM, Feil DK, Dunning KR, Lane M, Russell DL. Human cumulus cell gene expression as a biomarker of pregnancy outcome after single embryo transfer. Fertil Steril. 2011;96:47–52. doi: 10.1016/j.fertnstert.2011.04.033. [DOI] [PubMed] [Google Scholar]
- 35.Assou S, Haouzi D, De Vos J, Hamamah S. Human cumulus cells as biomarkers for embryo and pregnancy outcomes. Mol Hum Reprod. 2010;16:531–538. doi: 10.1093/molehr/gaq032. [DOI] [PubMed] [Google Scholar]
- 36.Iager AE, Kocabas AM, Otu HH, Ruppel P, Langerveld A, Schnarr P, Suarez M, Jarrett JC, Conaghan J, Rosa GJM, Fernández E, Rawlins RG, Cibelli JB, Crosby JA. Identification of a novel gene set in human cumulus cells predictive of an oocyte’s pregnancy potential. Fertil Steril. 2013;99:745–752. doi: 10.1016/j.fertnstert.2012.10.041. [DOI] [PubMed] [Google Scholar]
- 37.Wissing ML, Kristensen SG, Andersen CY, Mikkelsen AL, Høst T, Borup R, Grøndahl ML. Identification of new ovulation-related genes in humans by comparing the transcriptome of granulosa cells before and after ovulation triggering in the same controlled ovarian stimulation cycle. Hum Reprod. 2014;29:997–1010. doi: 10.1093/humrep/deu008. [DOI] [PubMed] [Google Scholar]
- 38.Agca C, Yakan A, Agca Y. Estrus synchronization and ovarian hyper-stimulation treatments have negligible effects on cumulus oocyte complex gene expression whereas induction of ovulation causes major expression changes. Mol Reprod Dev. 2012;80:102–117. doi: 10.1002/mrd.22141. [DOI] [PubMed] [Google Scholar]
- 39.Devjak R, Fon Tacer K, Juvan P, Virant Klun I, Rozman D, Vrtačnik Bokal E. Cumulus cells gene expression profiling in terms of oocyte maturity in controlled ovarian hyperstimulation using GnRH agonist or GnRH antagonist. PLoS One. 2012;7:e47106. doi: 10.1371/journal.pone.0047106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Harshman SW, Young NL, Parthun MR, Freitas MA. H1 histones: current perspectives and challenges. Nucleic Acids Res. 2013;41:9593–9609. doi: 10.1093/nar/gkt700. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Plumb M, Marashi F, Green L, Zimmerman A, Zimmerman S, Stein J, Stein G. Cell cycle regulation of human histone H1 mRNA. Proc Natl Acad Sci U S A. 1984;81:434–438. doi: 10.1073/pnas.81.2.434. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Hao Z, Zhang H, Cowell J. Ubiquitin-conjugating enzyme UBE2C: molecular biology, role in tumorigenesis, and potential as a biomarker. Tumour Biol. 2012;33:723–730. doi: 10.1007/s13277-011-0291-1. [DOI] [PubMed] [Google Scholar]
- 43.Lee B-K, Bhinge AA, Iyer VR. Wide-ranging functions of E2F4 in transcriptional activation and repression revealed by genome-wide analysis. Nucleic Acids Res. 2011;39:3558–3573. doi: 10.1093/nar/gkq1313. [DOI] [PMC free article] [PubMed] [Google Scholar]